
Pomological Traits and Genome Size of Prunus armeniaca L. Considering to Geographical Origin



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Pomological Analysis
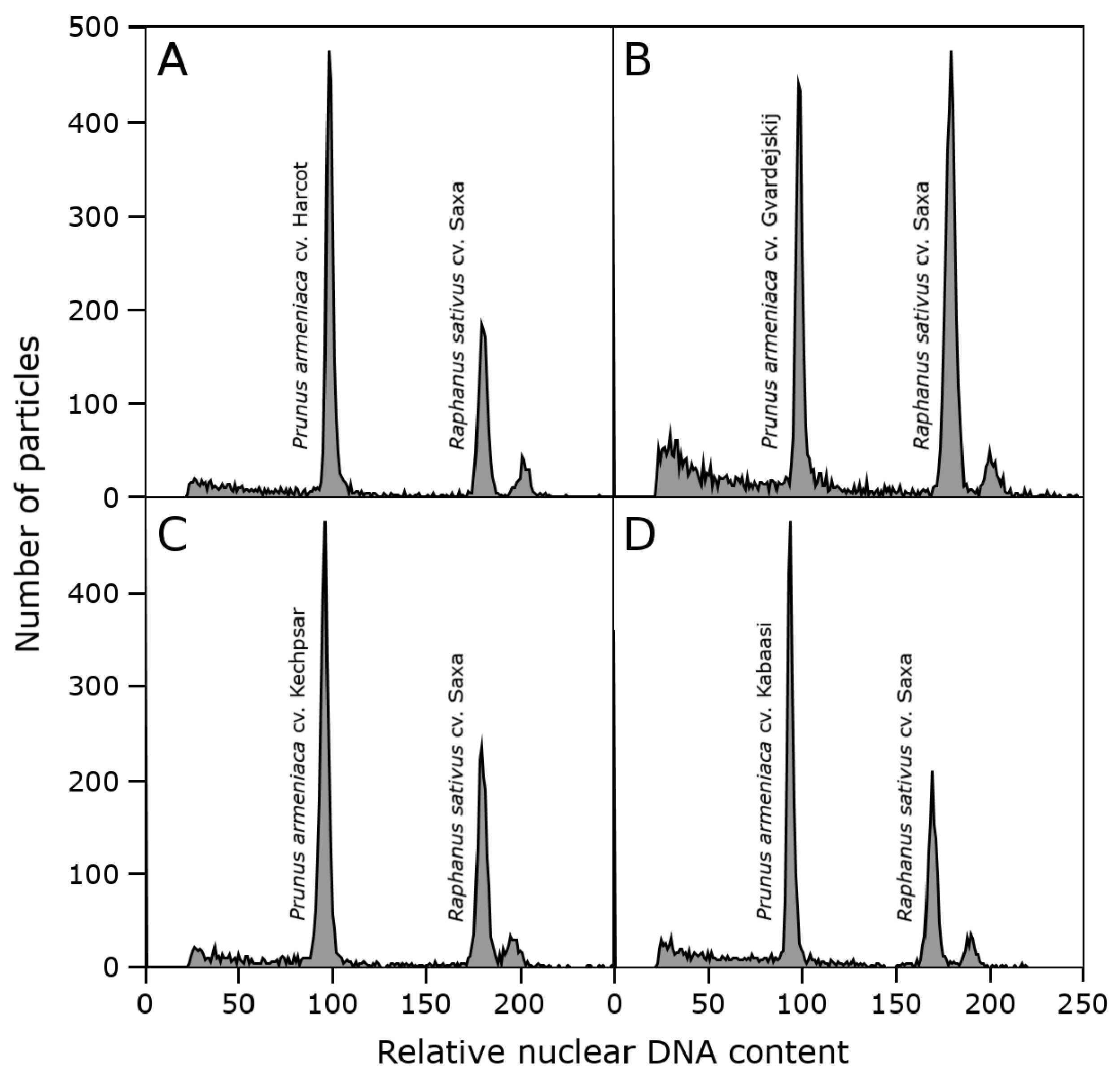
2.3. Genome Size Estimation
2.4. Statistical Analysis
3. Results
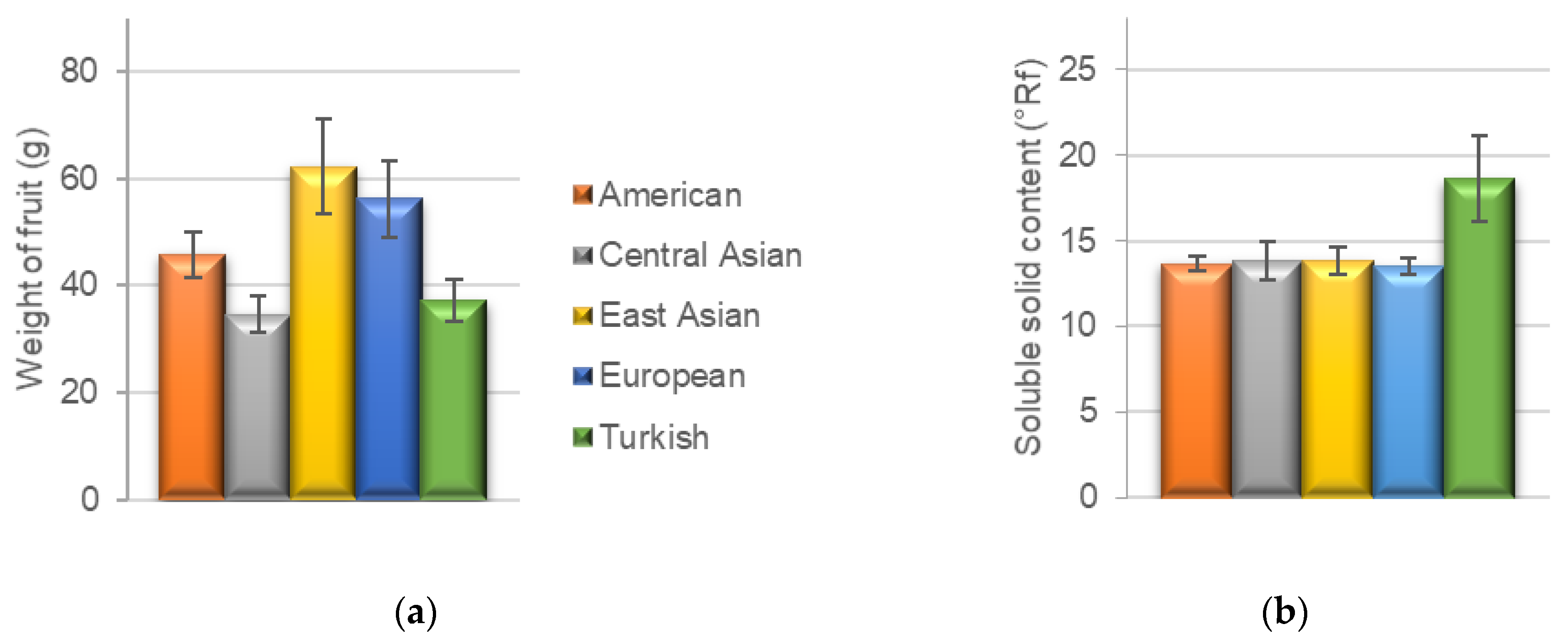
3.1. Pomological Traits
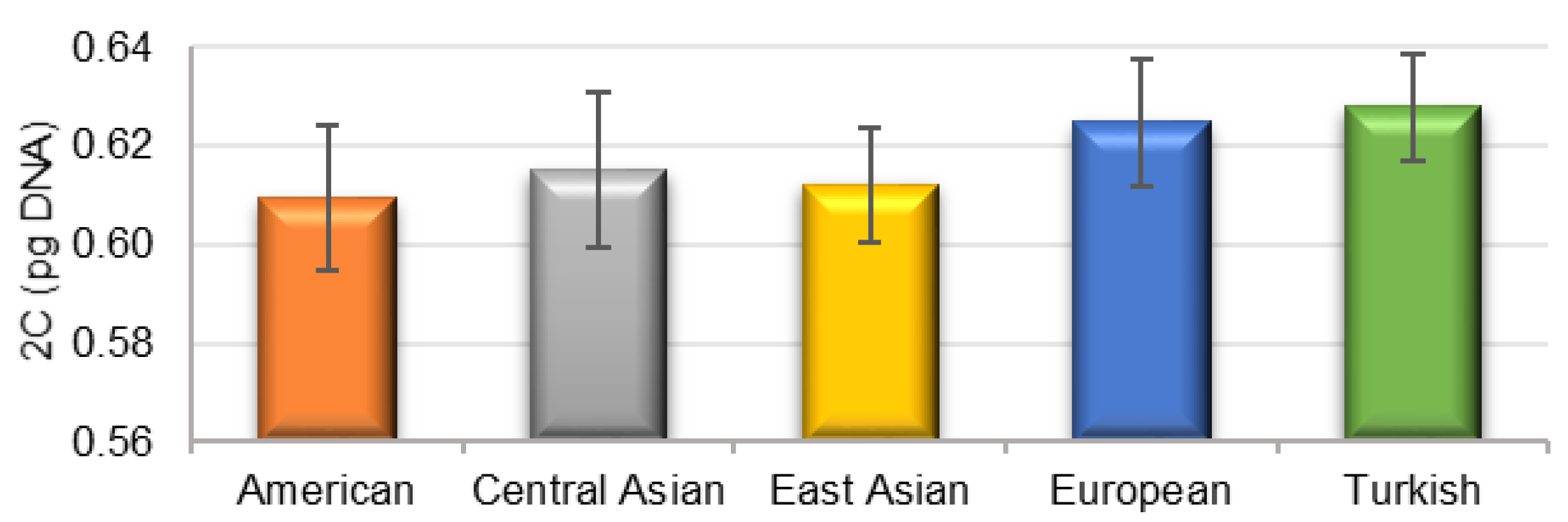
3.2. Genome Size Estimation
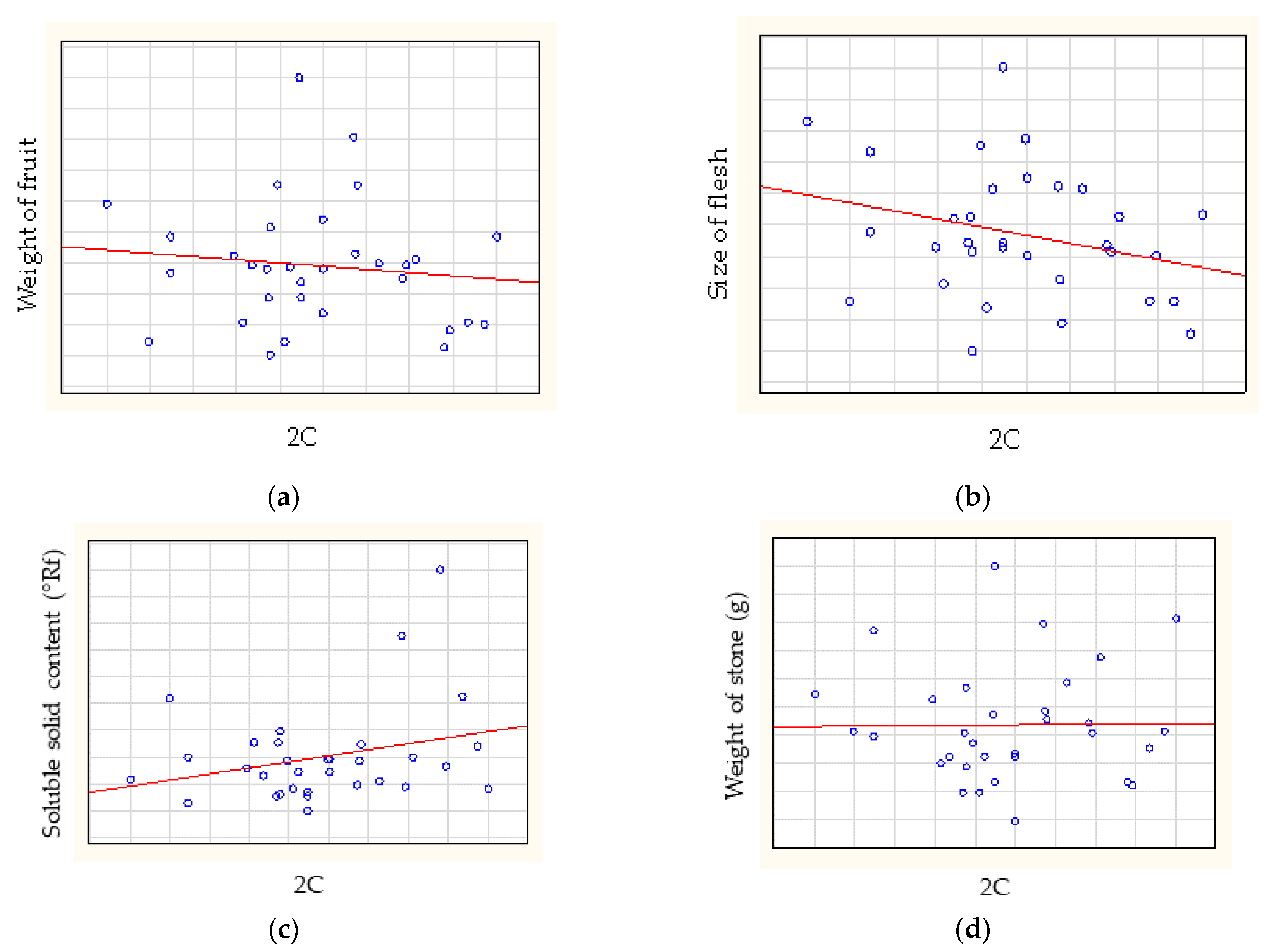
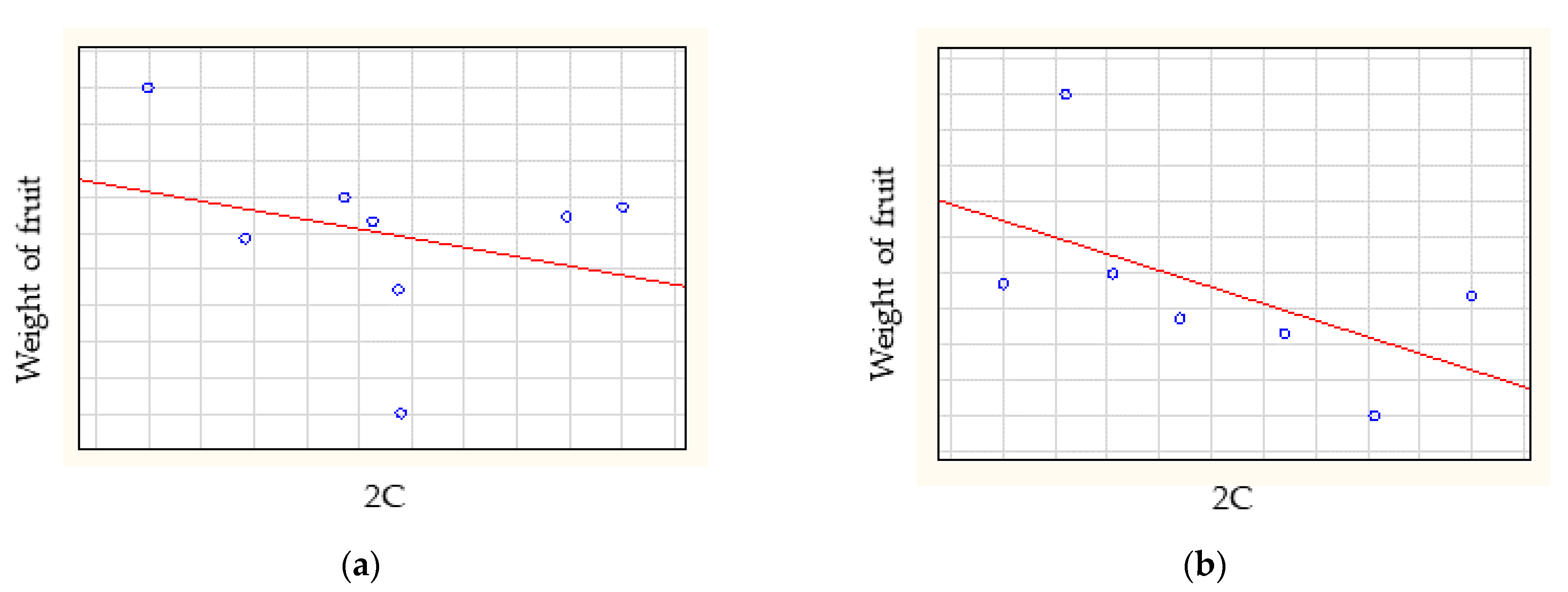
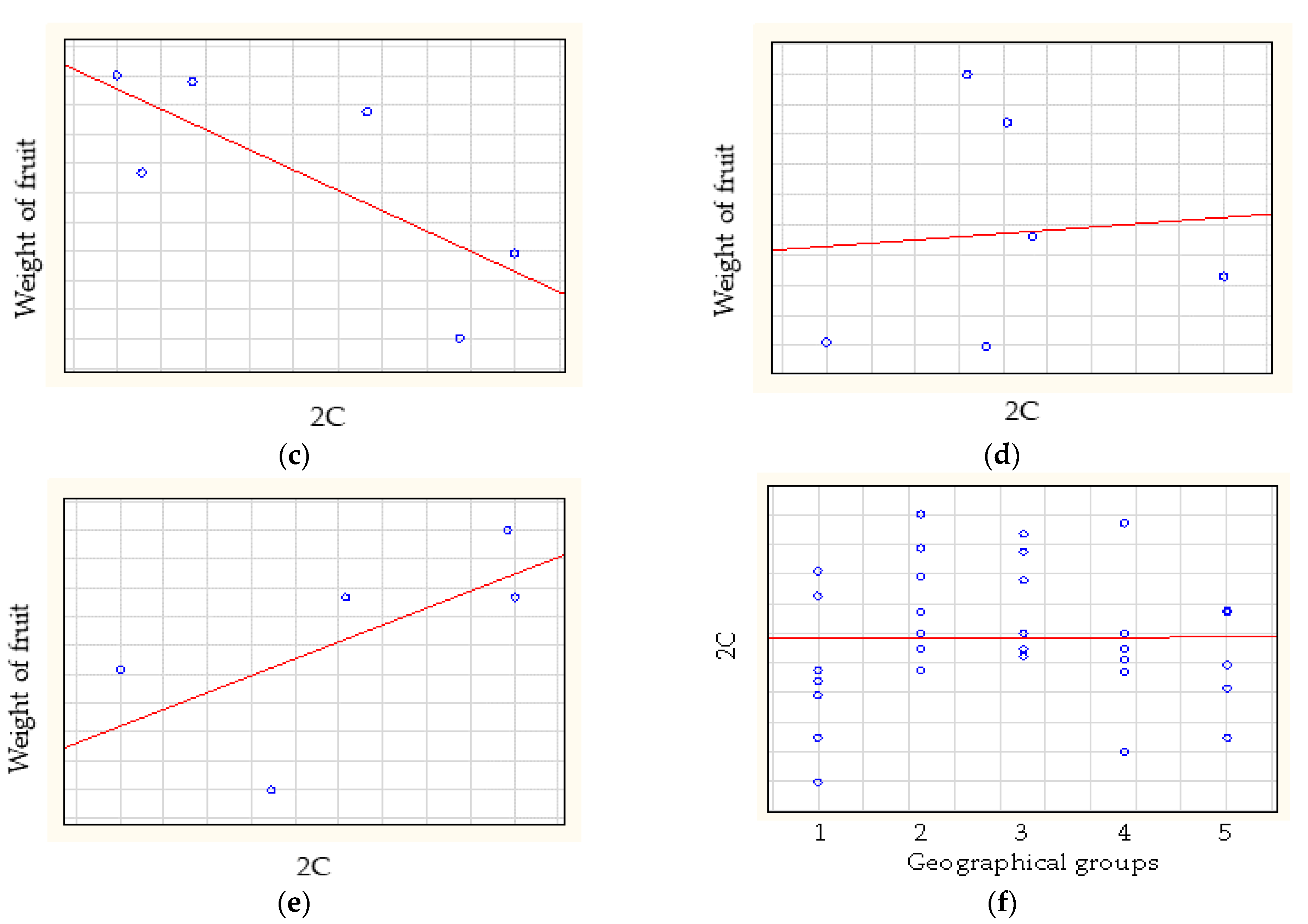
3.3. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rehder, A. Manual of Cultivated Trees and Shrubs, 2nd ed.; MacMillan: New York, NY, USA, 1940; pp. 452–481. [Google Scholar]
- Gharaghani, A.; Solhjoo, S.; Oraguzie, N. A review of genetic resources of almonds and stone fruits (Prunus spp.) in Iran. Genet. Resour. Crop Evol. 2017, 64, 611–640. [Google Scholar] [CrossRef]
- Jiang, F.; Zhang, J.; Wang, S.; Yang, L.; Yinfeng, L.; Gao, S.; Zhang, M.; Wu, S.; Hu, S.; Sun, H.; et al. The apricot (Prunus armeniaca L.) genome elucidates Rosaceae evolution and beta-carotenoid synthesis. Hortic. Res. 2019, 6, 128. [Google Scholar] [CrossRef]
- Löschnig, I.; Passecker, F. Die Marille (Aprikose) und Ihre Kultur; Ost. Agrarverlag: Wienna, Vienna, 1954. [Google Scholar]
- Gatti, E.; Defilippi, G.B.; Predieri, S.; Infante, R. Apricot (Prunus armeniaca L.) quality and breeding perspectives. J. Food. Agric. Environ. 2009, 7, 573–580. [Google Scholar]
- Frantianni, F.; Ombra, M.N.; d’Acierno, A.; Cipriano, L.; Nazzaro, R. Apricots: Biochemistry and functional properties. Curr. Opin. Food Sci. 2018, 19, 23–29. [Google Scholar]
- Milosevic, T.; Milosevic, N.; Glisic, I.; Krska, B. Characteristics of promising apricot (Prunus armeniaca L.) genetic resources in Central Serbia based on blossoming period and fruit quality. Hort. Sci. 2010, 37, 46–55. [Google Scholar]
- Food and Agriculture Organization of the United Nations. Crops and Livestock Products; FAOSTAT. FAO: Rome, Italy, 2022. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 5 February 2022).
- Layne, R.E.C.; Bailey, C.H.; Hough, L.F. Apricots. In Fruit Breeding. Vol II: Tree and Tropical Fruits; Janick, J., Moore, J.N., Eds.; John Wiley and Sons: New York, NY, USA, 1996; pp. 79–111. [Google Scholar]
- Faust, M.; Surány, D.; Nyujtó, F. Origin and dissemination of apricot. Hort. Rev. 1998, 22, 225–266. [Google Scholar]
- Childers, N.; Sherman, W. The Peach; Horticultural Publications: Gainesville, FL, USA, 1988. [Google Scholar]
- Kadkhodaei, S.; Arzani, K.; Yadollahi, A.; Karimzadeh, G.; Abdollahi, H. Genetic Diversity and Similarity of Asian and European Pears (Pyrus Spp.) Revealed by Genome Size and Morphological Traits Prediction. Int. J. Fruit Sci. 2021, 21, 619–633. [Google Scholar] [CrossRef]
- Shearer, K.; Ranney, T.G. Ploidy Levels and Relative Genome Sizes of Species, Hybrids, and Cultivars of Dogwood (Cornus spp.). HortScience 2013, 48, 825–830. [Google Scholar] [CrossRef]
- Li, W.; Liu, L.; Wang, Y.; Fan, G.; Zhang, S.; Wang, Y.; Liao, K. Determination of genome size and chromosome ploidy of selected taxa from Prunus armeniaca by flow cytometry. Sci. Hortic. 2020, 261, 417–432. [Google Scholar] [CrossRef]
- Jatt, T.; Lee, M.; Rayburn, A.L.; Jatol, M.A.; Mirani, A.A. Determination of genome size variations among different date palm cultivars (Phoenix dactylifera L.) by flow cytometry. Biotech 2019, 9, 457. [Google Scholar] [CrossRef]
- Tamarzizt, H.B.; Walker, D.; Mustapha, S.B.; Abdallah, D.; Baraket, G.; Hannachi, A.S.; Azzouzi, S.Z. DNA variation and polymorphism in Tunisian plum species (Prunus spp.): Contribution of flow cytometry and molecular markers. Genet. Mol. Res. 2015, 14, 18034–18046. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.R.; Guo, Z.H.; Xiao, J.Z.; Wang, F.Y. Karyotype analysis of Armeniaca sibirica (L.) Lam. And Armeniaca vulgaris Lam. J. Hebei Agric. Univ. 1986, 6, 14–19. [Google Scholar]
- Arumuganathan, K.; Earle, E.D. Nuclear DNA content of some important plant species. Plant Mol. Biol. Rep. 1991, 9, 208–218. [Google Scholar] [CrossRef]
- Dickson, E.E.; Arumuganathan, K.; Kresovich, S.; Doyle, J.J. Nuclear DNA content variation within the Rosaceae. Am. J. Bot. 1992, 79, 1081–1086. [Google Scholar] [CrossRef]
- Kolano, B.; Siwinska, D.; Gomez Pando, L.; Szymanowska-Pulka, J.; Maluszynska, J. Genome size variation in Chenopodium quinoa (Chenopodiaceae). Plant. Syst. Evol. 2012, 298, 251–255. [Google Scholar] [CrossRef][Green Version]
- Knight, C.A.; Molinary, N.; Petrov, D. The large genome constraint hypothesis:evolution, ecology, and phenotype. Ann. Bot. 2005, 5, 177–190. [Google Scholar] [CrossRef]
- Jadrná, P.; Konza, F.; Plavcová, O. Polyploidization of Pelargonium × hortorum L. H. Bailey in greenhouse conditions. Hort. Sci. 2009, 36, 31–37. [Google Scholar] [CrossRef]
- Dalbato, A.L.; Kobza, F.; Karlsson, L.M. Effect of polyploidy and pollination methods on capsule and seed set of pansies (Viola × wittrockiana Gams). Hort. Sci. 2013, 40, 22–30. [Google Scholar] [CrossRef]
- Müller, L.B.; Zotz, G.; Albach, D.C. Bromeliaceae subfamilies show divergent trends of genome size evolution. Sci. Rep. 2019, 9, 5136. [Google Scholar] [CrossRef]
- Rodrigues, P.S.; Souza, M.M.; Melo, C.A.F.; Pereira, T.N.S.; Corrêa, R.X.C. Karyotype diversity and 2C DNA content in species of the Caesalpinia group. BMC Genet. 2018, 19, 25. [Google Scholar] [CrossRef]
- Sakhanokho, H.F.; Timothy Rinehart, A.; Stringer, S.J.; Islam-Faridi, M.N.; Pounders, C.T. Variation in nuclear DNA content and chromosome numbers in blueberry. Sci. Hortic. 2018, 233, 108–113. [Google Scholar] [CrossRef]
- Swift, H. The constancy of desoxyribose nucleic acid in plant nuclei. Proc. Natl. Acad. Sci. USA 1950, 36, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Nitranský, Š. Klasifikátor—Descriptor List Genus Armeniaca P. Mill. Vyd. Semex ŠP Bratislava; VS Veselé a VÚRV: Praha, Czech Republic, 1992; p. 29. [Google Scholar]
- Doležel, J.; Sgorbatti, S.; Lucretti, S. Comparison of three DNA fluorochromes for flow cytometric estimation of nuclear DNA content in plants. Physiol. Plant. 1992, 85, 625–631. [Google Scholar] [CrossRef]
- Doležel, J.; Liu, L.; Wang, Y.; Fan, G.; Zhang, S.; Wang, Y.; Liao, K. Plant DNA Flow Cytometry and Estimation of Nuclear Genome Size. Ann. Bot. 2005, 95, 99–110. [Google Scholar] [CrossRef]
- Wang, L.; Luo, Z.; Liu, Z.; Zhao, J.; Deng, W.; Wei, H.; Liu, P.; Liu, M. Genome Size Variation within Species of Chinese Jujube (Ziziphus jujube Mill.) and Its Wild Ancestor Sour Jujube (Z. acidojujuba Cheng et Liu). Forests 2019, 10, 460. [Google Scholar] [CrossRef]
- Martínez-Sagarra, G.; Castro, S.; Mota, L.; Loureiro, J.; Devesa, J.A. Genome size, chromosome number and morphological data reveal unexpected infraspecific variability in Festuca (Poaceae). Genes 2021, 12, 906. [Google Scholar] [CrossRef]
- Nečas, T.; Göttingerová, M.; Wolf, J.; Kiss, T.; Rampáčková, E.; Ondrášek, I. New promising apricot hybrids from Faculty of Horticulture in Lednice. Acta Hortic. 2020, 1290, 169–178. [Google Scholar] [CrossRef]
- Saridaş, M.A.; Paydaş Kargı, S.; Kafkas, E. Phenological and pomological characteristics of some important apricot cultivars grown in Turkey. Acta Hortic. 2019, 1242, 553–560. [Google Scholar] [CrossRef]
- Kumar, D.; Lal, S.; Ahmed, N. Morphological and pomological diversity among apricot (Prunus armeniaca) genotypes grown in India. Indian J. Agric. Sci. 2015, 85, 1349–1355. [Google Scholar]
- Ayour, J.; Harrak, H.; Alahyane, A.; Taourirte, M.; Renard, C.M.G.C.; Benichou, M. Pomological and antioxidant properties of apricot (Prunus armeniaca L.): Fruit management and quality analysis. J. Food Microbiol. Saf. Hyg. 2018, 3, 53. [Google Scholar]
- Karatas, N.; Ercisli, S.; Bozhuyuk, M.R. Assessment of Morphological Traits, Nutritional and Nutraceutical Composition in Fruits of 18 Apricot cv. Sekerpare Clones. Sustainability 2021, 13, 11385. [Google Scholar] [CrossRef]
- Göttingerová, M.; Nečas, T. Comparison of selected qualitative characteristic of American, French and Czech apricot cultivars. Acta Hortic. 2020, 1290, 159–168. [Google Scholar]
- Gurrieri, F.; Audergon, J.M.; Albagnac, G.; Reich, M. Soluble sugars and carboxylic acids in ripe apricot fruit as parameters for distinguishing different cultivars. Euphytica 2001, 117, 183–189. [Google Scholar] [CrossRef]
- Gecer, M.K.; Kan, T.; Gundogdu, M.; Ercisli, S.; Ilhan, G.; Sagbas, H.I. Physicochemical characteristics of wild and cultivated apricots (Prunus armeniaca L.) from Aras valley in Turkey. Genet. Resour. Crop Evol. 2020, 67, 935–945. [Google Scholar] [CrossRef]
- Głowacka, A.; Sitarek, M.; Rozpara, E.; Podwyszyńska, M. Pomological Characteristics and Ploidy Levels of Japanese Plum (Prunus salicina Lindl.) Cultivars Preserved in Poland. Plants 2021, 10, 884. [Google Scholar] [CrossRef]
- Baird, W.V.; Estager, A.S.; Wells, J.K. Estimating Nuclear DNA Content in Peach and Related Diploid Species Using Laser Flow Cytometry and DNA Hybridization. J. Am. Soc. Hort. Sci. 1994, 119, 1312–1316. [Google Scholar] [CrossRef]
- Leitch, I.J.; Soltis, D.E.; Soltis, P.S.; Bennet, M.D. Evolution of DNA amounts across land plants (embryophyta). Ann. Bot. 2005, 95, 207–217. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, W.; Sun, L.; Zhao, F.; Huang, B.; Yang, W.; Tao, Y.; Wang, J.; Yuan, Z.; Fan, G.; et al. The genome of Prunus mume. Nat. Commun. 2012, 3, 1318. [Google Scholar] [CrossRef]
- Vitte, C.; Panaud, O. Formation of solo-LTRs through unequal homologous recombination counterbalances amplifications of LTR retrotransposons in rice Oryza sativa L. Mol. Biol. Evol. 2003, 20, 528–540. [Google Scholar] [CrossRef]
- Maghuly, F.; Fernandez, E.B.; Ruthner, S.; Pedryc, A.; Laimer, M. Microsatellite variability in apricots (Prunus armeniaca L.) reflects their geographic origin and breeding history. Tree Genet. Genomes 2005, 1, 151–165. [Google Scholar] [CrossRef]
- Leitch, I.J.; Chase, M.V.; Bennett, M.D. Phylogenetic analysis of DNA C-values provides evidence for a small ancestral genome size in flowering plants. Ann. Bot. 1998, 82, 85–94. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Origin | Date of Maturity (2021) | Weight of Fruit (g) | Colour of Fruit * | Shape of Fruit ** | Size of Flesh (mm) | Colour of Flesh *** | Weight of Stone (g) | Soluble Solid Content (°Rf) | Weight of Kernel (g) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Early Blush | American | 27.7 | 38.6 | ± | 1.7 a,b,c,d,e,f,g,h,i,j,k | 7 | 7 | 11.4 | ± | 0.5 h,i,j,k,l,m,n | 5 | 2.7 | ± | 0.2 f,g,h,i,j,k,l,m,n,o | 15.3 | ± | 0.4 g,h,i,j,k | 0.9 | ± | 0.10 |
| Goldrich | American | 15.7 | 50.5 | ± | 3.1 g,h,i,j,k,l | 4 | 5 | 10.3 | ± | 0.4 a,b,c,d,e,f,g,h,i,j | 6 | 3.9 | ± | 0.2 o,p | 13.7 | ± | 0.6 a,b,c,d,e,f,g,i | 1.0 | ± | 0.03 |
| Harcot | American | 19.7 | 47.5 | ± | 2.0 d,e,f,g,h,i,j,k,l | 4 | 7/8 | 11.3 | ± | 0.2 f,g,h,i,j,k,l,m,n | 6 | 3.1 | ± | 0.1 g,h,i,j,k,l,m,n,o | 13.3 | ± | 0.5 a,b,c,d,e,f,g,h,i | 0.8 | ± | 0.10 |
| Harlayne | American | 19.7 | 45.4 | ± | 2.5 d,e,f,g,h,i,j | 5 | 8 | 10.9 | ± | 0.5 e,f,g,h,i,j,k,l | 6 | 3.4 | ± | 0.1 i,j,k,l,m | 14.4 | ± | 0.3 e,f,g,h,i | 0.9 | ± | 0.10 |
| Orangered | American | 8.7 | 64.7 | ± | 4.0 i,j,k,l | 4 | 5 | 14.9 | ± | 0.3 n | 5 | 2.9 | ± | 0.1 f,g,h,i,j,k,l | 13.1 | ± | 0.4 a,b,c,d,e,f,g,i | 1.1 | ± | 0.10 |
| Scout | American | 19.7 | 22.6 | ± | 1.1 a,b,c | 2 | 4 | 6.5 | ± | 0.3 a | 6 | 2.6 | ± | 0.1 d,e,f,g,h,i,j,k | 12.1 | ± | 0.4 a,b,c,d,e,f,g,i | 0.8 | ± | 0.04 |
| Stark Early Orange | American | 15.7 | 49.5 | ± | 3.3 g,h,i,j,l | 4 | 3 | 11.4 | ± | 0.3 g,h,i,j,k,l,m,n | 5 | 3.5 | ± | 0.2 k,l,m,n,o,p | 14.4 | ± | 0.6 c,d,e,f,g,h,i | 1.2 | ± | 0.05 |
| Stella | American | 19.7 | 48.0 | ± | 2.2 f,g,h,i,j,l | 5 | 5 | 12.4 | ± | 0.5 d,e,f,g,h,i,j,k,l | 6 | 3.9 | ± | 0.2 l,m,n,o,p | 13.0 | ± | 0.4 a,b,c,d,e,f,g,i | 1.1 | ± | 0.07 |
| Achrori | Central Asian | 7.7 | 42.9 | ± | 1.6 c,d,e,f,g,h,i | 5 | 2/5 | 10.5 | ± | 0.3 d,e,f,g,h,i,j,k | 5 | 2.7 | ± | 0.1 e,f,g,h,i,j,k | 11.1 | ± | 0.3 a,f | 0.7 | ± | 0.04 |
| Gvardejskij | Central Asian | 28.6 | 26.2 | ± | 0.8 a | 3 | 8 | 8.1 | ± | 0.2 a,c,d,e,g | 3 | 1.7 | ± | 0.03 a,c,e | 12.4 | ± | 0.4 a,b,c,d,e,f | 0.6 | ± | 0.01 |
| Kechpsar | Central Asian | 24.9 | 26.4 | ± | 0.4 a,b | 4 | 2 | 8.3 | ± | 0.2 a,b,c,d,e,g | 5 | 2.7 | ± | 0.1 d,e,f,g,h,i,j,k | 18.1 | ± | 0.5 i,j,k | 0.9 | ± | 0.03 |
| Lagerdi Mashhad | Central Asian | 19.7 | 34.3 | ± | 1.4 a,b,c,d,e,f | 3 | 3 | 14.2 | ± | 0.3 a,c,d,g | 7 | 2.6 | ± | 0.1 h,i,j,k,l,m | 14.3 | ± | 0.5 e,f,g,h,i | 0.8 | ± | 0.10 |
| Marena | Central Asian | 27.7 | 31.4 | ± | 1.3 a,b,c,d,e,f | 5 | 3/4 | 7.1 | ± | 0.4 a,c,g | 6 | 3.1 | ± | 0.1 h,i,j,k,l,m,n,o | 15.1 | ± | 0.7 f,g,h,i,j | 0.9 | ± | 0.10 |
| Roxana | Central Asian | 25.7 | 46.6 | ± | 5.2 d,e,f,g,h,i,j,k,l | 7 | 2 | 10.4 | ± | 0.3 b,c,d,e,f,g,h,i,j,k,j,l,m,n | 5 | 2.3 | ± | 0.1 a,b,c,d,e,f,g,h,i,j | 12.0 | ± | 0.4 a,b,c,d,e,f,g,i | 0.6 | ± | 0.03 |
| Chuang Zhi Hong | East Asian | 27.7 | 70.1 | ± | 4.4 j,k,l | 5 | 5 | 14.0 | ± | 0.3 l,m,n | 7 | 2.8 | ± | 0.1 f,g,h,i,j,k | 14.2 | ± | 0.6 b,c,d,e,f,g,h,i,j,k | 0.9 | ± | 0.10 |
| Chuangxing | East Asian | 15.7 | 83.6 | ± | 2.7 l | 4 | 3 | 12.5 | ± | 0.6 j,k,l,m,n | 5 | 6.7 | ± | 0.3 p | 12.7 | ± | 0.8 a,b,c,d,e,f | 1.4 | ± | 0.10 |
| Inbeixing | East Asian | 9.7 | 55.7 | ± | 1.8 h,i,j,k,l | 7 | 3 | 13.8 | ± | 0.4 k,l,m,n | 7 | 4.2 | ± | 0.1 n,o,p | 11.5 | ± | 0.3 a,b,d,f | 1.3 | ± | 0.20 |
| Liaoning | East Asian | 21.6 | 70.2 | ± | 3.2 k,l | 4 | 3 | 7.6 | ± | 0.6 m,n | 7 | 3.2 | ± | 0.1 a,b,c,d,e,f,g,h,i | 15.3 | ± | 0.3 b,c,d,e,f,g,h,i | 1.0 | ± | 0.10 |
| Saimaiti | East Asian | 15.7 | 31.9 | ± | 1.4 a,b,c,d | 3 | 5 | 9.0 | ± | 0.4 a,b,c,d,e,f,g,h | 5 | 2.6 | ± | 0.1 c,d,e,f,g,h,i,j | 15.4 | ± | 0.4 h,i,j,k | 0.8 | ± | 0.20 |
| Ananasova | European | 22.7 | 51.1 | ± | 2.2 h,i,j,k,l | 7 | 2 | 9.1 | ± | 0.6 a,b,c,d,e,f,g,h | 7 | 3.4 | ± | 0.1 i,j,k,l,m,n,o,p | 14.2 | ± | 0.3 b,c,d,e,f,g,h,i | 1.0 | ± | 0.04 |
| Bohuticka | European | 19.7 | 55.8 | ± | 2.8 i,j,k,l | 5 | 3 | 11.5 | ± | 0.6 i,j,k,l,m,n | 6 | 4.4 | ± | 0.1 n,o,p | 12.4 | ± | 0.2 a,b,c,d,e,f | 1.4 | ± | 0.10 |
| Karola | European | 7.7 | 29.6 | ± | 1.3 a,b,c,d,e,f,g | 7 | 4 | 10.0 | ± | 0.3 a,b,c,d,e,f,g,h,i,j,k,l | 5 | 2.0 | ± | 0.1 a,b,c,d,e,f,g,h | 13.8 | ± | 0.5 a,b,c,d,e,f,g,h,i,j | 0.7 | ± | 0.04 |
| Pastyrik | European | 30.6 | 99.9 | ± | 4.8 l | 7 | 4 | 16.8 | ± | 0.5 n | 7 | 6.2 | ± | 0.3 p | 12.0 | ± | 0.7 a,b,c,d,e,f,g,i | 1.0 | ± | 0.10 |
| Pozde kvetouci | European | 28.6 | 60.7 | ± | 2.2 i,j,k,l | 4 | 3/5 | 12.8 | ± | 0.3 i,j,k,l,m,n | 6 | 2.6 | ± | 0.1 b,c,d,e,f,g,h,i,j | 13.5 | ± | 0.4 a,b,c,d,e,f,g,i | 0.5 | ± | 0.10 |
| Pozdni chramova | European | 27.7 | 58.3 | ± | 2.3 i,j,k,l | 4 | 5/8 | 10.1 | ± | 0.5 c,d,e,f,g,h,i | 5 | 4.0 | ± | 0.1 k,l,m,n,o | 16.0 | ± | 0.5 d,e,f,g,h,i | 1.1 | ± | 0.04 |
| Velkopavlovicka | European | 19.7 | 47.9 | ± | 1.3 e,f,g,h,i,j,l | 4 | 5 | 10.1 | ± | 0.5 a,b,c,d,e,f,g,h,i | 6 | 3.4 | ± | 0.1 i,j,k,l,m,n,o,p | 12.6 | ± | 0.4 a,b,c,d,e,f,g,i | 0.9 | ± | 0.04 |
| Wondercot | European | 23.6 | 46.9 | ± | 1.1 d,e,f,g,h,i,j,l | 4 | 8 | 12.4 | ± | 0.2 i,j,k,l,m,n | 6 | 2.2 | ± | 0.1 a,b,c,d,e,f,g | 13.5 | ± | 0.9 a,b,c,d,e,f,g,i | 0.8 | ± | 0.04 |
| Hacihaliloglu | Turkish | 27.7 | 32.0 | ± | 1.4 a,b,c,d | 3 | 3/5 | 8.3 | ± | 0.3 a,b,c,d,e,g | 4 | 2.4 | ± | 0.1 a,b,c,d,e,f,g,h | 18.2 | ± | 1.2 j,k | 0.8 | ± | 0.03 |
| Hasanbey | Turkish | 27.7 | 43.8 | ± | 1.1 d,e,f,g,h,i | 3 | 5 | 10.4 | ± | 0.2 a,b,c,d,e,f,g,h,i,j,k | 5 | 3.2 | ± | 0.1 i,j,k,l,m,n,o | 22.0 | ± | 0.7 k | 0.9 | ± | 0.04 |
| Kabaasi | Turkish | 15.7 | 38.7 | ± | 0.8 b,c,d,e,f,g,h | 2 | 3 | 10.3 | ± | 0.0 a,b,c,d,e,f,g,h,i,j | 5 | 4.0 | ± | 0.1 m,n,o,p | 12.3 | ± | 0.4 a,b,c,d,e,f | 1.6 | ± | 0.20 |
| Salak | Turkish | 19.7 | 46.5 | ± | 2.4 d,e,f,g,h,i | 3 | 8 | 9.9 | ± | 0.3 a,b,c,d,e,f,g,h,i | 2 | 2.3 | ± | 0.1 a,b,c,d,e,f,g,h | 14.4 | ± | 0.5 a,b,c,d,e,f,g,i | 0.8 | ± | 0.04 |
| Sekerpare | Turkish | 27.7 | 24.8 | ± | 1.4 a | 3 | 5 | 8.4 | ± | 0.2 a,b,c,d,e,g | 3/4 | 2.1 | ± | 0.1 a,b,c,d,e,f | 26.1 | ± | 0.6 k | 0.7 | ± | 0.04 |
| Group | Date of Maturity (2021) | Weight of Fruit (g) | Colour of Fruit * | Shape of Fruit ** | Size of Flesh (mm) | Colour of Flesh *** | Weight of Stone (g) | Soluble Solid Content (°Rf) | Weight of Kernel (g) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| American | 17.7 | 45.8 | ± | 4.20 | 4 | 5 | 11.1 | ± | 0.80 | 5/6 | 3.3 | ± | 0.20 | 13.7 | ± | 0.40 | 1.0 | ± | 0.10 |
| Central Asian | 12.7 | 34.6 | ± | 3.50 | 4/5 | 4 | 9.8 | ± | 1.00 | 5/6 | 2.5 | ± | 0.20 | 13.8 | ± | 1.10 | 0.8 | ± | 0.10 |
| East Asian | 17.7 | 62.3 | ± | 8.80 | 4/5 | 4 | 11.3 | ± | 1.30 | 6 | 3.9 | ± | 0.80 | 13.8 | ± | 0.80 | 1.1 | ± | 0.10 |
| European | 14.7 | 56.3 | ± | 7.10 | 5 | 4 | 11.6 | ± | 0.90 | 6 | 3.5 | ± | 0.50 | 13.5 | ± | 0.50 | 0.9 | ± | 0.10 |
| Turkish | 23.7 | 37.2 | ± | 4.00 | 3 | 5 | 9.4 | ± | 0.50 | 4 | 2.8 | ± | 0.40 | 18.6 | ± | 2.50 | 1.0 | ± | 0.20 |
| Genotype | Origin | Average 2C (pg DNA) | Average 1Cx (Mbp) | Genotype | Origin | Average 2C (pg DNA) | Average 1Cx (Mbp) |
|---|---|---|---|---|---|---|---|
| Early Blush | American | 0.610 ± 0.001 | 298.5 | Saimaiti | East Asian | 0.607 ± 0.000 | 296.6 |
| Goldrich | American | 0.605 ± 0.002 | 296.0 | Ananasova | European | 0.623 ± 0.004 | 304.7 |
| Harcot | American | 0.608 ± 0.001 | 297.3 | Bohuticka | European | 0.644 ± 0.002 | 314.8 |
| Harlayne | American | 0.596 ± 0.002 | 291.4 | Karola | European | 0.637 ± 0.001 | 311.4 |
| Orangered | American | 0.587 ± 0.002 | 286.9 | Pastyrik | European | 0.615 ± 0.001 | 300.7 |
| Scout | American | 0.611 ± 0.001 | 298.6 | Pozde kvetouci | European | 0.618 ± 0.001 | 302.4 |
| Stark Early Orange | American | 0.632 ± 0.001 | 309.0 | Pozdni chramova | European | 0.611 ± 0.005 | 298.6 |
| Stella | American | 0.626 ± 0.001 | 306.3 | Velkopavlovicka | European | 0.631 ± 0.001 | 308.3 |
| Achrori | Central Asian | 0.615 ± 0.005 | 300.8 | Wondercot | European | 0.614 ± 0.001 | 300.0 |
| Gvardejskij | Central Asian | 0.613 ± 0.001 | 299.6 | Hacihaliloglu | Turkish | 0.640 ± 0.011 | 312.8 |
| Kechpsar | Central Asian | 0.593 ± 0.002 | 289.9 | Hasanbey | Turkish | 0.630 ± 0.001 | 308.1 |
| Lagerdi Mashhad | Central Asian | 0.618 ± 0.002 | 302.4 | Kabaasi | Turkish | 0.615 ± 0.002 | 300.8 |
| Marena | Central Asian | 0.642 ± 0.001 | 313.9 | Salak | Turkish | 0.618 ± 0.006 | 302.4 |
| Roxana | Central Asian | 0.610 ± 0.001 | 298.3 | Sekerpare | Turkish | 0.636 ± 0.000 | 311.0 |
| Chuang Zhi Hong | East Asian | 0.612 ± 0.001 | 299.1 | P. ansu | 0.608 ± 0.001 | 297.2 | |
| Chuangxing | East Asian | 0.623 ± 0.004 | 304.6 | P. mume | 0.581 ± 0.002 | 284.2 | |
| Inbeixing | East Asian | 0.596 ± 0.001 | 291.4 | P. sibirica | 0.621 ± 0.001 | 303.5 | |
| Liaoning | East Asian | 0.623 ± 0.001 | 304.9 | Average | 0.616 ± 0.015 | 301.3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rampáčková, E.; Mrázová, M.; Čížková, J.; Nečas, T. Pomological Traits and Genome Size of Prunus armeniaca L. Considering to Geographical Origin. Horticulturae 2022, 8, 199. https://doi.org/10.3390/horticulturae8030199
Rampáčková E, Mrázová M, Čížková J, Nečas T. Pomological Traits and Genome Size of Prunus armeniaca L. Considering to Geographical Origin. Horticulturae. 2022; 8(3):199. https://doi.org/10.3390/horticulturae8030199
Chicago/Turabian StyleRampáčková, Eliška, Martina Mrázová, Jana Čížková, and Tomáš Nečas. 2022. "Pomological Traits and Genome Size of Prunus armeniaca L. Considering to Geographical Origin" Horticulturae 8, no. 3: 199. https://doi.org/10.3390/horticulturae8030199
APA StyleRampáčková, E., Mrázová, M., Čížková, J., & Nečas, T. (2022). Pomological Traits and Genome Size of Prunus armeniaca L. Considering to Geographical Origin. Horticulturae, 8(3), 199. https://doi.org/10.3390/horticulturae8030199