Physiological Characteristics and Proteome of Dalbergia odorifera in Loam and Gravel Substrates


Abstract
1. Introduction
2. Materials and Methods
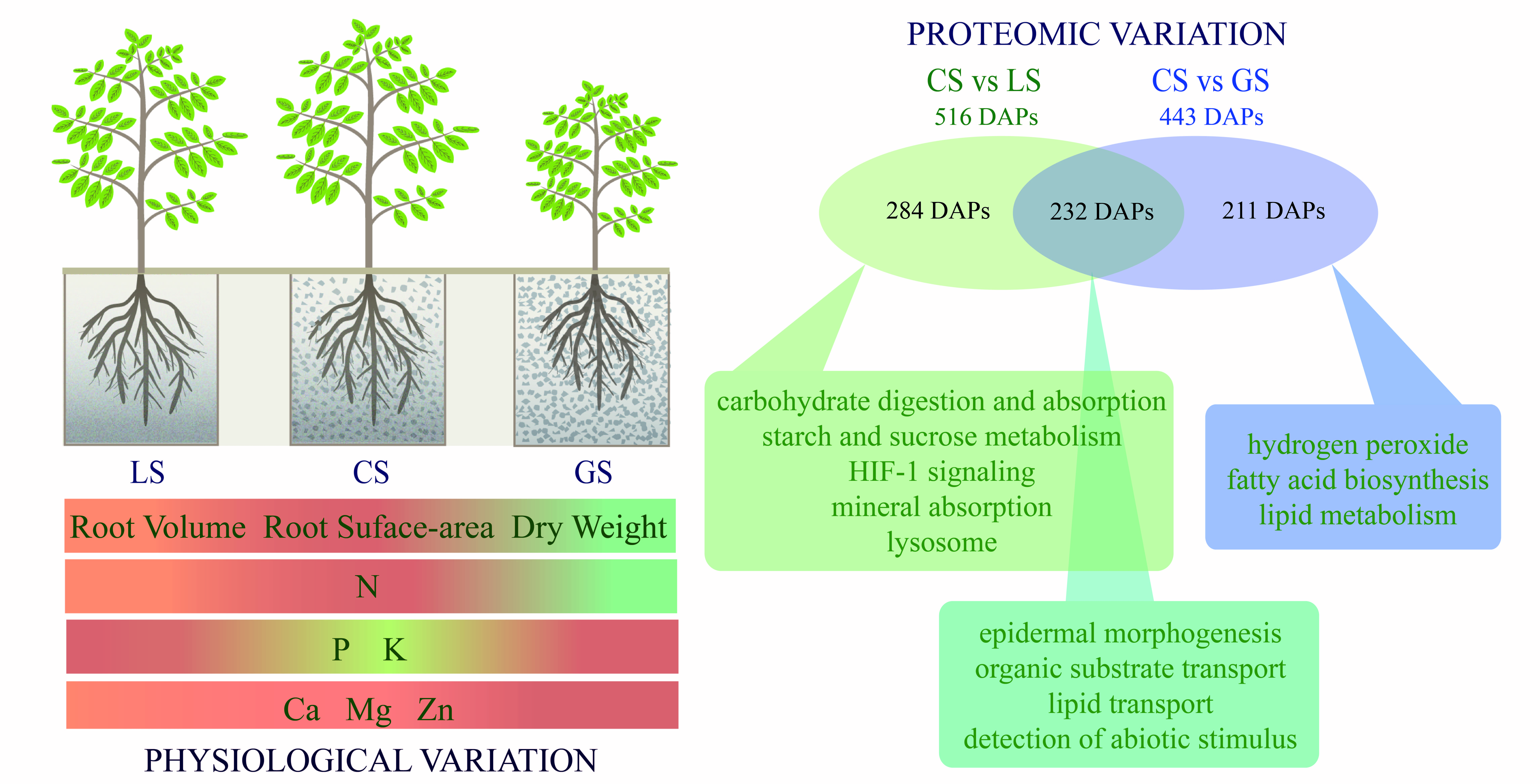
2.1. Plant Materials and Growth Conditions
2.2. Collection of Experimental Materials
2.3. Analysis of Basic Properties and Mineral Element Content of the Substrate and Plant
2.4. Protein Preparation of Root Tip for Proteomic Analysis
2.5. iTRAQ-Based Differential Proteomic Analysis by LC-MS/MS
2.6. Data Processing and Bioinformatics Analysis
2.7. Quantitative Reverse Transcription PCR (RT-qPCR)
2.8. Statistical Analysis
3. Results
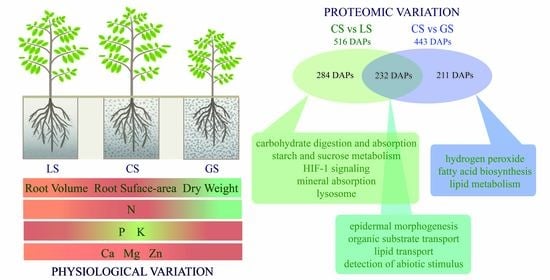
3.1. Comparison of Basic Physicochemical Characteristics of Three Growth Substrates
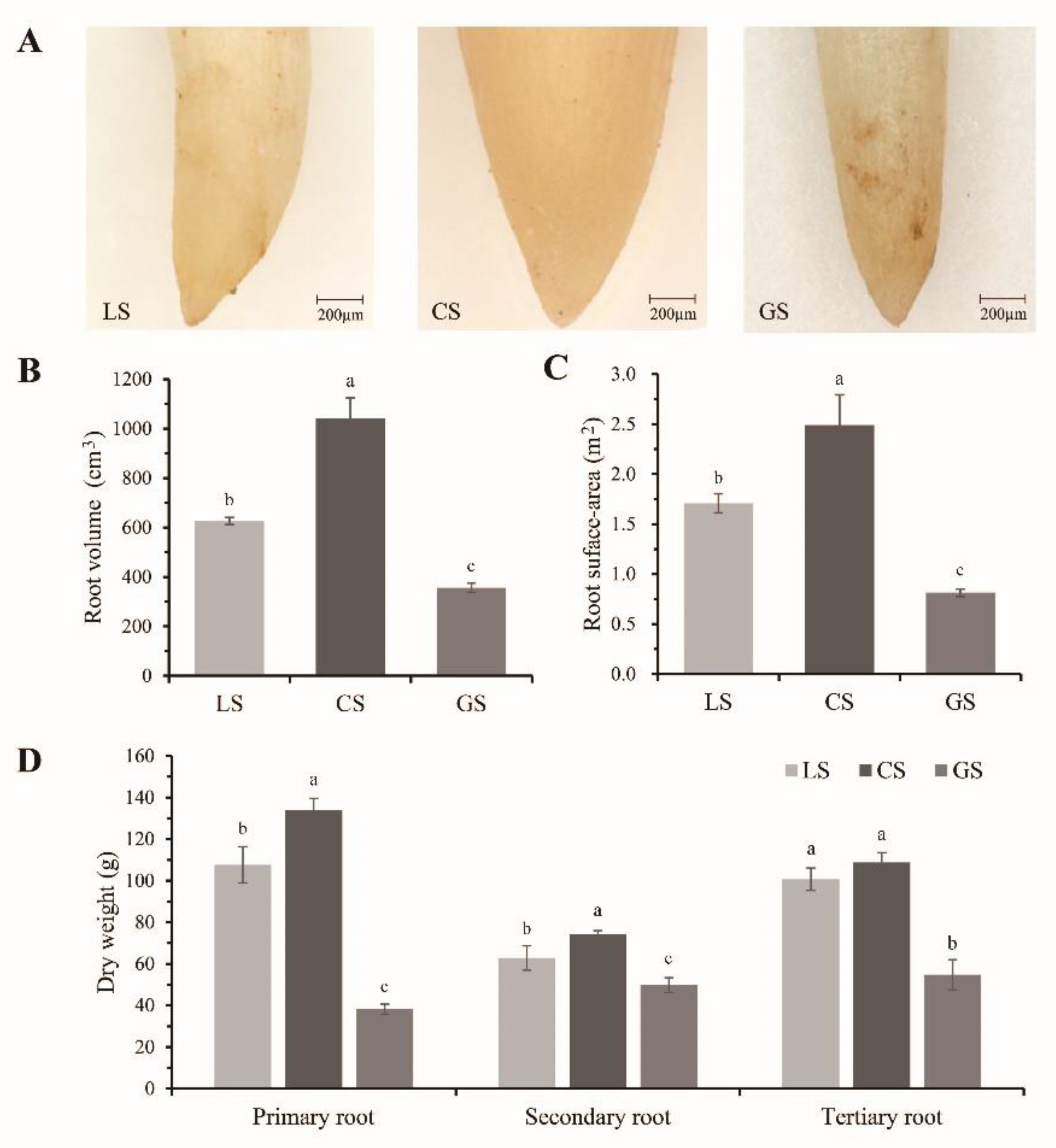
3.2. Root Tip Development of D. odorifera
3.3. Substrate Containing Gravel Changes Root Element Content
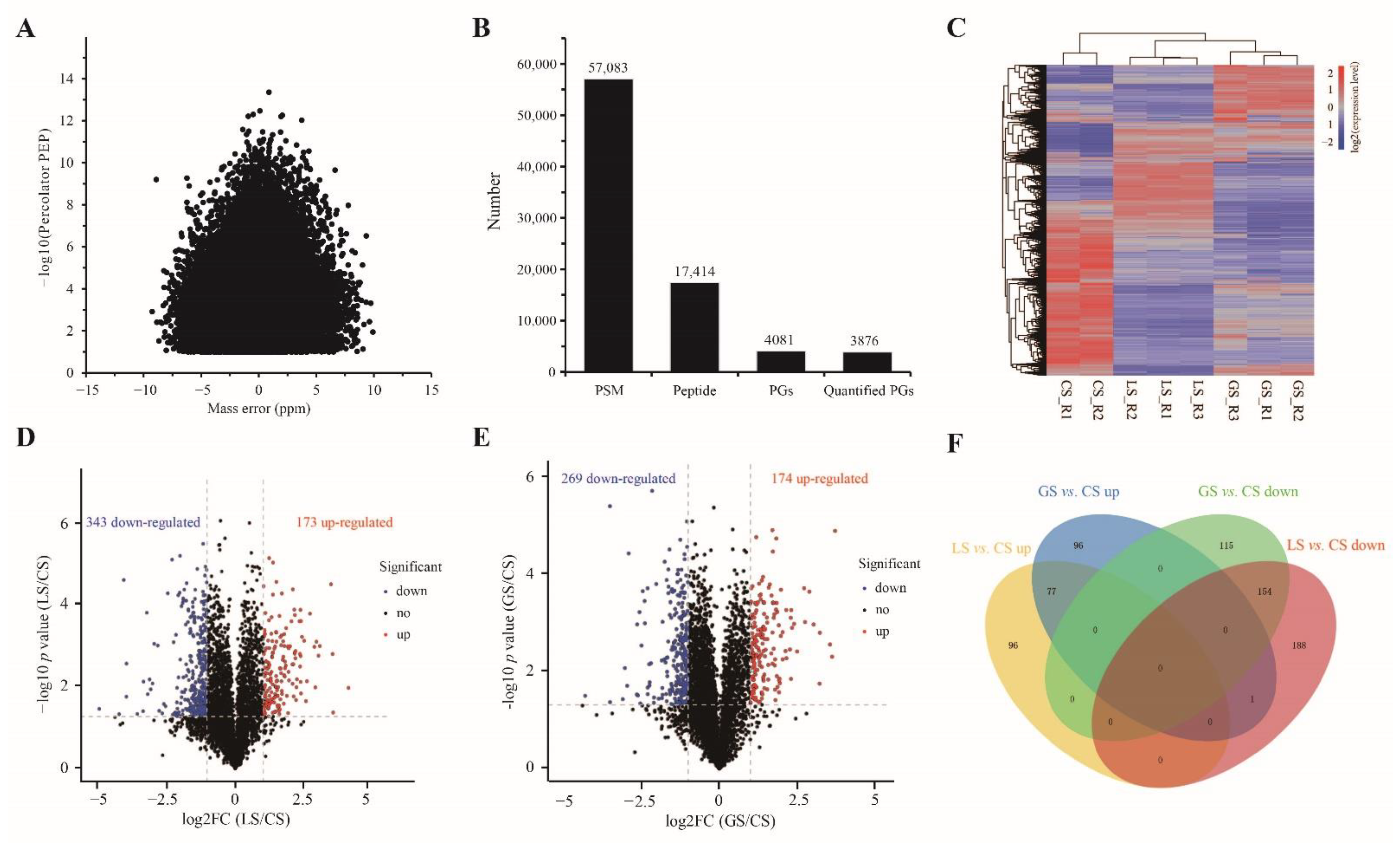
3.4. Differential Proteomics in Root Tips
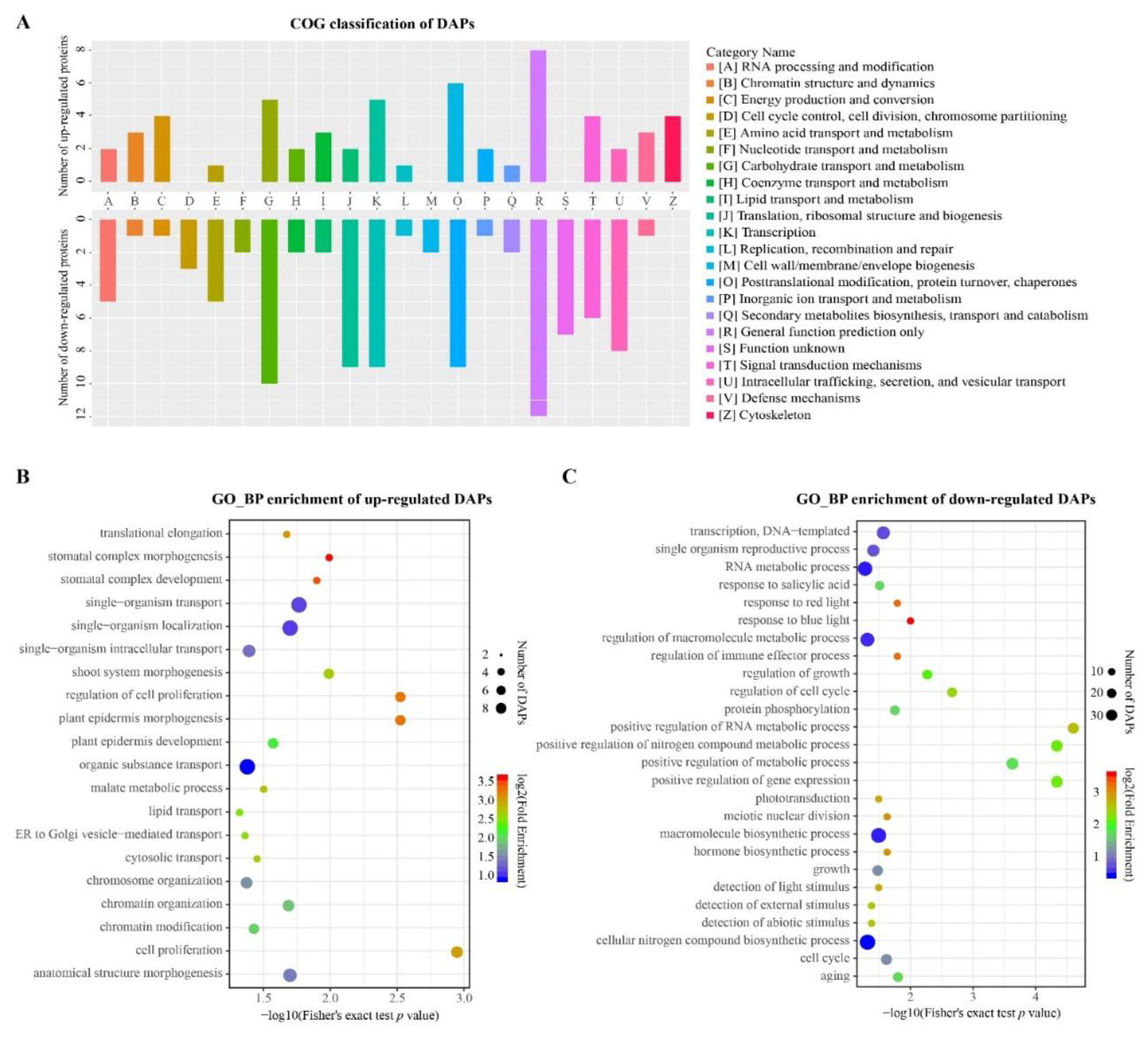
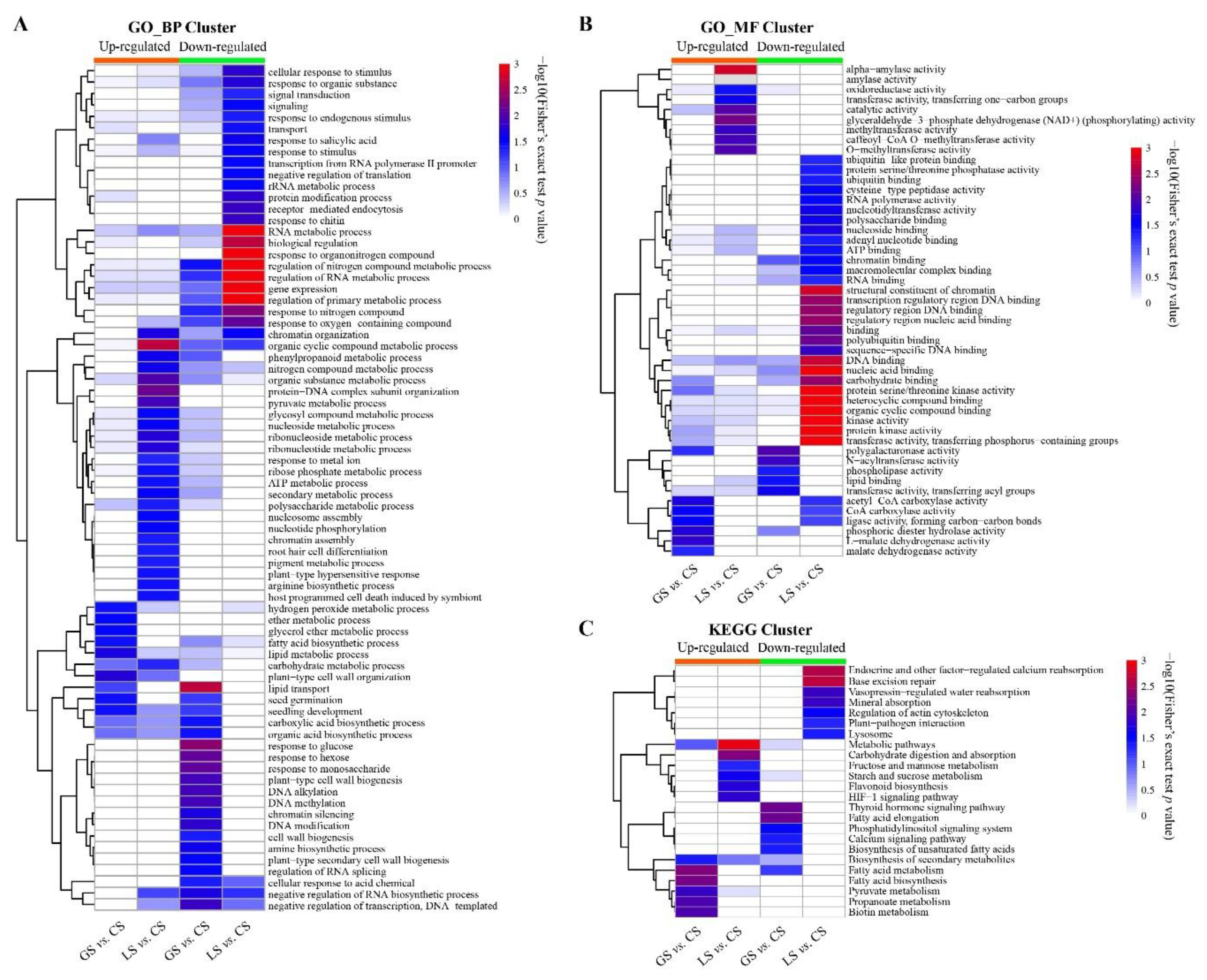
3.5. Functional Analysis of the Overlapped DAPs Regulated by LS and GS Substrates
3.6. Functional Analysis of DAPs Specifically Regulated by LS and GS Substrates
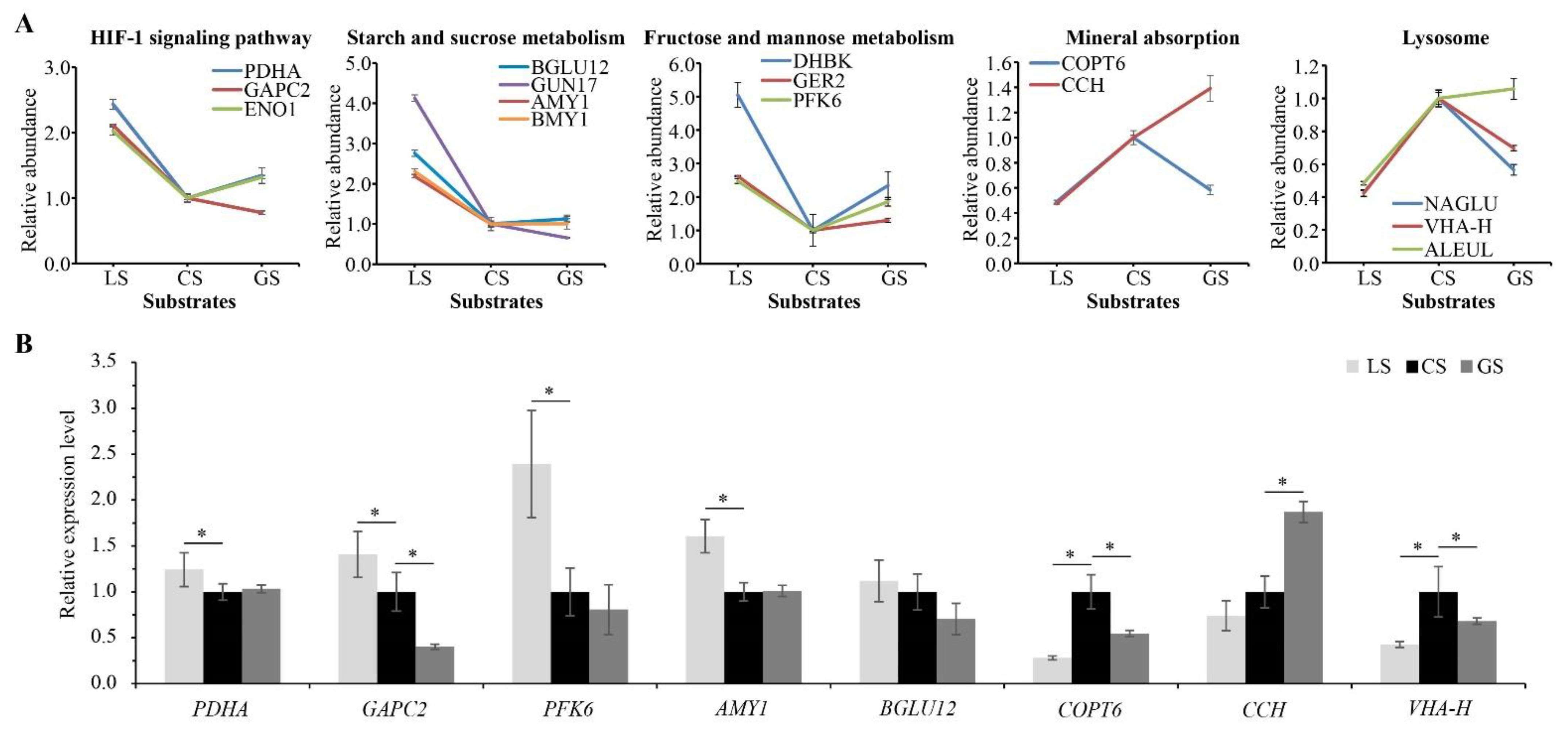
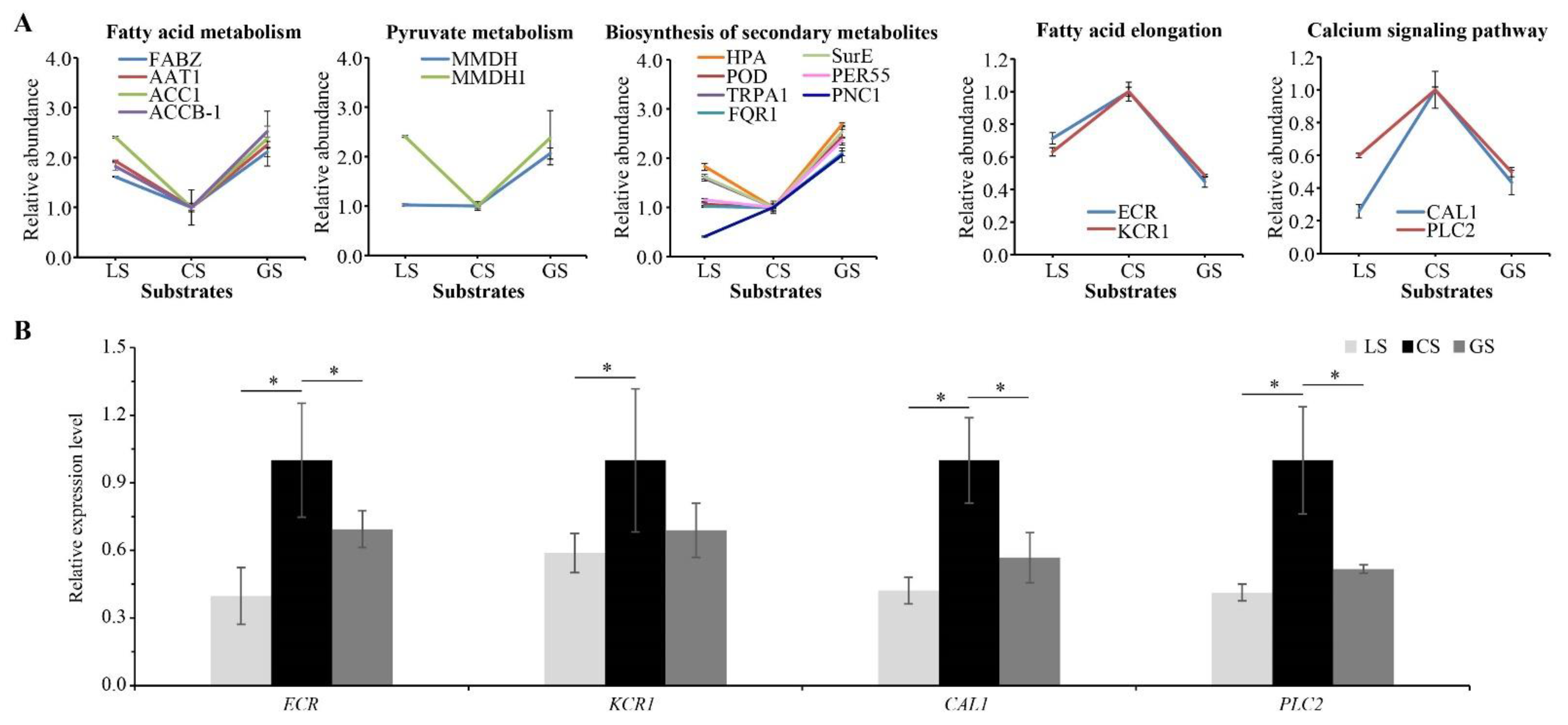
3.7. Effects of Substrate on Proteins Related to Air Permeability, Glucose Metabolism, and Heat Conduction
3.8. Effects of Substrate on Proteins Related to Nutrient Metabolism and Secondary Metabolism
4. Discussion
4.1. Effects of Substrates on Respiration of Root Tips
4.2. Effects of Substrates on Fatty Acid Metabolism of Root Tips
4.3. Effects of Substrates on Cell Wall Remodeling of Root Tips
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, S.; Zeng, X.; Zhang, D.; Guo, S. Diverse fungi associated with partial irregular heartwood of Dalbergia odorifera. Sci. Rep. 2015, 5, 8464. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Li, J.; Liu, X.; Lian, J.; Zhang, N.; Yang, Z.; Niu, Y.; Cui, Z.; Xu, D. The chromosome-level draft genome of Dalbergia odorifera. Gigascience 2020, 9, giaa084. [Google Scholar] [CrossRef] [PubMed]
- Hui, M.; Caixiang, X.; Yun, Y.; Jianhe, W.; Jindong, F.; Shilin, C. Suitable producing areas of Dalbergia odorifera T. Chen. Lishizhen Med. Mater. Med. Res. 2010, 21, 2304–2306. [Google Scholar] [CrossRef]
- Meng, X.W.; Wang, D.Q.; Chen, L.Y.; Shao, F.; Zhang, P.Z.; Hu, S.J.; Zhu, Q.; Liu, R.H. Study on neoflavonoids from heartwood of Dalbergia latifolia. Zhongguo Zhong Yao Za Zhi 2019, 44, 1186–1192. [Google Scholar] [CrossRef] [PubMed]
- Wariss, H.M.; Yi, T.-S.; Wang, H.; Zhang, R. Characterization of the complete chloroplast genome of Dalbergia odorifera (Leguminosae), a rare and critically endangered legume endemic to China. Conserv. Genet. Resour. 2017, 10, 527–530. [Google Scholar] [CrossRef]
- Lee, D.S.; Li, B.; Keo, S.; Kim, K.S.; Jeong, G.S.; Oh, H.; Kim, Y.C. Inhibitory effect of 9-hydroxy-6,7-dimethoxydalbergiquinol from Dalbergia odorifera on the NF-kappaB-related neuroinflammatory response in lipopolysaccharide-stimulated mouse BV2 microglial cells is mediated by heme oxygenase-1. Int. Immunopharmacol. 2013, 17, 828–835. [Google Scholar] [CrossRef]
- Li, X.; Wu, L.; Liu, W.; Jin, Y.; Chen, Q.; Wang, L.; Fan, X.; Li, Z.; Cheng, Y. A network pharmacology study of Chinese medicine QiShenYiQi to reveal its underlying multi-compound, multi-target, multi-pathway mode of action. PLoS ONE 2014, 9, e95004. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, C.; Meng, H.; Yu, Z.; Yang, M.; Wei, J. Dalbergia odorifera: A review of its traditional uses, phytochemistry, pharmacology, and quality control. J. Ethnopharmacol. 2020, 248, 112328. [Google Scholar] [CrossRef]
- Tao, Y.; Wang, Y. Bioactive sesquiterpenes isolated from the essential oil of Dalbergia odorifera T. Chen. Fitoterapia 2010, 81, 393–396. [Google Scholar] [CrossRef]
- Sugiyama, A.; Zhu, B.M.; Takahara, A.; Satoh, Y.; Hashimoto, K. Cardiac effects of salvia miltiorrhiza/Dalbergia odorifera mixture, an intravenously applicable Chinese medicine widely used for patients with ischemic heart disease in China. Circ. J. 2002, 66, 182–184. [Google Scholar] [CrossRef]
- Chan, S.C.; Chang, Y.S.; Wang, J.P.; Chen, S.C.; Kuo, S.C. Three new flavonoids and antiallergic, anti-inflammatory constituents from the heartwood of Dalbergia odorifera. Planta Med. 1998, 64, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Zhang, Q.; Zhou, H.; Nong, Z.; Ye, S.; Deng, Q. Introduction of Dalbergia odorifera enhances nitrogen absorption on Eucalyptus through stimulating microbially mediated soil nitrogen-cycling. For. Ecosyst. 2021, 8, 12. [Google Scholar] [CrossRef]
- Lu, J.; Yang, F.; Wang, S.; Ma, H.; Liang, J.; Chen, Y. Co-existence of Rhizobia and Diverse Non-rhizobial Bacteria in the Rhizosphere and Nodules of Dalbergia odorifera Seedlings Inoculated with Bradyrhizobium elkanii, Rhizobium multihospitium-Like and Burkholderia pyrrocinia-Like Strains. Front. Microbiol. 2017, 8, 2255. [Google Scholar] [CrossRef]
- Yue, X.H.; Miao, L.F.; Yang, F.; Nawaz, M. Morphological and physiological responses of Dalbergia odorifera T. Chen seedlings to different culture substances. PLoS ONE 2020, 15, e0232051. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xia, H.; Wang, J.; Chen, Q. Nutrient uptake and assimilation in fragrant rosewood (Dalbergia odorifera T.C. Chen) seedlings in growing media with un-composted spent mushroom residue. PLoS ONE 2021, 16, e0249534. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.B.; Heuberger, A.L.; Broeckling, C.D.; Jahn, C.E. Non-Targeted Metabolomics Reveals Sorghum Rhizosphere-Associated Exudates are Influenced by the Belowground Interaction of Substrate and Sorghum Genotype. Int. J. Mol. Sci. 2019, 20, 431. [Google Scholar] [CrossRef]
- He, Q.F.; Peng, Y.H.; Cao, Y.Y.; Jiang, Y.; Lu, G.D. Experiment on substrates for container seedling of Dalbergia odorifera. J. For. Eng. 2012, 26, 92–95. [Google Scholar] [CrossRef]
- Li, X.Y.; Zeng, B.S.; Xu, D.P. Effects of Different Transplant Substrate on Survival Ratio and Growth of Dalbergia odorifera T. Chen Tissue-cultured Plantlets. Seed 2019, 38, 16–20. [Google Scholar] [CrossRef]
- Cao, K.L.; Liu, Z.J.; Chen, Y.L.; Liu, C.H. Study on the Container Seedlings of Dalbergia odorifera. J. Anhui Agric. Sci. 2014, 42, 5082–5083, 5087. [Google Scholar] [CrossRef]
- Sun, C.X.; Feng, M.L.; Liu, L.Y.; Chen, W.J.; Chen, H.; Zhang, M.Y.; Li, X.J. Study on the main Physico-chemical Properties of Coir Culture Medium in Hainan. Chin. J. Trop. Crops 2011, 32, 407–411. [Google Scholar] [CrossRef]
- Lin, Y.; Xiong, W.; Xiao, S.; Li, F.; Lu, Z.; Yan, J.; Fang, X.; Cui, X.; Wen, Y.; Liang, J.; et al. Pharmacoproteomics reveals the mechanism of Chinese dragon’s blood in regulating the RSK/TSC2/mTOR/ribosome pathway in alleviation of DSS-induced acute ulcerative colitis. J. Ethnopharmacol. 2020, 263, 113221. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time pcr experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Meng, H.; Yang, Y.; Gao, Z.H.; Wei, J.H. Selection and Validation of Reference Genes for Gene Expression Studies by RT-PCR in Dalbergia odorifera. Sci. Rep. 2019, 9, 3341. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Luan, H.; Shen, H.; Pan, Y.; Guo, B.; Lv, C.; Xu, R. Elucidating the hypoxic stress response in barley (Hordeum vulgare L.) during waterlogging: A proteomics approach. Sci. Rep. 2018, 8, 9655. [Google Scholar] [CrossRef]
- Pan, R.; He, D.; Xu, L.; Zhou, M.; Li, C.; Wu, C.; Xu, Y.; Zhang, W. Proteomic analysis reveals response of differential wheat (Triticum aestivum L.) genotypes to oxygen deficiency stress. BMC Genom. 2019, 20, 60. [Google Scholar] [CrossRef]
- Boaretto, L.F.; Labate, M.T.V.; Franceschini, L.M.; Cataldi, T.R.; Budzinski, I.G.F.; de Moraes, F.E.; Labate, C.A. Proteomics Reveals an Increase in the Abundance of Glycolytic and Ethanolic Fermentation Enzymes in Developing Sugarcane Culms During Sucrose Accumulation. Front. Plant Sci. 2021, 12, 716964. [Google Scholar] [CrossRef]
- Fox, T.C.; Mujer, C.V.; Andrews, D.L.; Williams, A.S.; Cobb, B.G.; Kennedy, R.A.; Rumpho, M.E. Identification and gene expression of anaerobically induced enolase in Echinochloa phyllopogon and Echinochloa cruspavonis. Plant Physiol. 1995, 109, 433–443. [Google Scholar] [CrossRef][Green Version]
- Albrecht, G.; Mustroph, A.; Fox, T.C. Sugar and fructan accumulation during metabolic adjustment between respiration and fermentation under low oxygen conditions in wheat roots. Physiol. Plant. 2004, 120, 93–105. [Google Scholar] [CrossRef]
- Hwang, J.H.; Yu, S.I.; Lee, B.H.; Lee, D.H. Modulation of Energy Metabolism Is Important for Low-Oxygen Stress Adaptation in Brassicaceae Species. Int. J. Mol. Sci. 2020, 21, 1787. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, Z.; Zhu, C.; Yue, J.; Yang, H.; Li, J.; Gao, J.; Xu, R.; Deng, X.; Cheng, Y. Variations of membrane fatty acids and epicuticular wax metabolism in response to oleocellosis in lemon fruit. Food Chem. 2021, 338, 127684. [Google Scholar] [CrossRef]
- Satoh, S.; Ozaki, M.; Matsumoto, S.; Nabatame, T.; Kaku, M.; Shudo, T.; Asayama, M.; Chohnan, S. Enhancement of fatty acid biosynthesis by exogenous acetyl-CoA carboxylase and pantothenate kinase in Escherichia coli. Biotechnol. Lett. 2020, 42, 2595–2605. [Google Scholar] [CrossRef]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2014, 5, 771. [Google Scholar] [CrossRef]
- Francoz, E.; Ranocha, P.; Nguyen-Kim, H.; Jamet, E.; Burlat, V.; Dunand, C. Roles of cell wall peroxidases in plant development. Phytochemistry 2015, 112, 15–21. [Google Scholar] [CrossRef]
- Oliveira, D.M.; Mota, T.R.; Salatta, F.V.; Sinzker, R.C.; Koncitikova, R.; Kopecny, D.; Simister, R.; Silva, M.; Goeminne, G.; Morreel, K.; et al. Cell wall remodeling under salt stress: Insights into changes in polysaccharides, feruloylation, lignification, and phenolic metabolism in maize. Plant Cell Environ. 2020, 43, 2172–2191. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | pH | Moisture Content (%) | Total C (g/kg) | Total N (g/kg) | Available P (mg/kg) | Available K (mg/kg) | Exchangeable Ca (cmol/kg) | Exchangeable Mg (cmol/kg) |
|---|---|---|---|---|---|---|---|---|
| LS | 6.21 ± 0.02 c | 33.50 ± 0.72 a | 11.36 ± 0.38 a | 1.84 ± 1.53 a | 3.72 ± 0.13 a | 55.33 ± 3.38 a | 55.59 ± 5.08 c | 3.79 ± 0.26 a |
| CS | 8.16 ± 0.02 b | 4.17 ± 0.11 b | 11.83 ± 0.09 a | 1.10 ± 0.08 ab | 5.17 ± 0.82 a | 60.67 ± 2.03 a | 95.71 ± 3.19 b | 1.71 ± 0.17 b |
| GS | 9.43 ± 0.04 a | 0.56 ± 0.02 c | 12.00 ± 0.14 a | 0.11 ± 0.01 b | 0.14 ± 0.01 b | 6.43 ± 0.05 b | 176.00 ± 3.79 a | 1.78 ± 0.10 b |
| Substrate | N (mg/kg) | P (mg/kg) | K (mg/kg) | Ca (mg/kg) | Mg (mg/kg) | Zn (mg/kg) |
|---|---|---|---|---|---|---|
| LS | 13,135.33 ± 529.35 b | 1170.35 ± 31.41 a | 5077.74 ± 29.25 ab | 19,363.04 ± 202.53 c | 2220.45 ± 195.23 b | 53.82 ± 4.83 b |
| CS | 15,539.12 ± 685.68 a | 899.16 ± 42.11 b | 4431.10 ± 12.31 b | 26,300.27 ± 849.11 b | 2133.71 ± 134.21 b | 78.09 ± 9.23 a |
| GS | 12,376.57 ± 732.25 b | 1061.02 ± 40.27 a | 6576.24 ± 11.52 a | 36,393.63 ± 517.33 a | 2727.74 ± 172.86 a | 84.83 ± 8.45 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.; Yu, K.; Yang, Z. Physiological Characteristics and Proteome of Dalbergia odorifera in Loam and Gravel Substrates. Horticulturae 2022, 8, 1154. https://doi.org/10.3390/horticulturae8121154
Yu S, Yu K, Yang Z. Physiological Characteristics and Proteome of Dalbergia odorifera in Loam and Gravel Substrates. Horticulturae. 2022; 8(12):1154. https://doi.org/10.3390/horticulturae8121154
Chicago/Turabian StyleYu, Shuzhong, Kai Yu, and Zhende Yang. 2022. "Physiological Characteristics and Proteome of Dalbergia odorifera in Loam and Gravel Substrates" Horticulturae 8, no. 12: 1154. https://doi.org/10.3390/horticulturae8121154
APA StyleYu, S., Yu, K., & Yang, Z. (2022). Physiological Characteristics and Proteome of Dalbergia odorifera in Loam and Gravel Substrates. Horticulturae, 8(12), 1154. https://doi.org/10.3390/horticulturae8121154