
Effect of Ethylene Sletr1-2 Receptor Allele on Flowering, Fruit Phenotype, Yield, and Shelf-Life of Four F1 Generations of Tropical Tomatoes (Solanum lycopersicum L.)

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Development of F1 Populations
2.2. Confirmation of Mutant Hybrid
2.3. Evaluation of Fruit Shelf-Life
2.4. Evaluation of Flowering and Fruit Phenotypes
2.5. Evaluation of Yield
2.6. Statistical Analysis
3. Results
3.1. Hybridization
3.2. Transfer of the Sletr1-2 Allele
3.3. Heterozygous Sletr1-2 Alleles
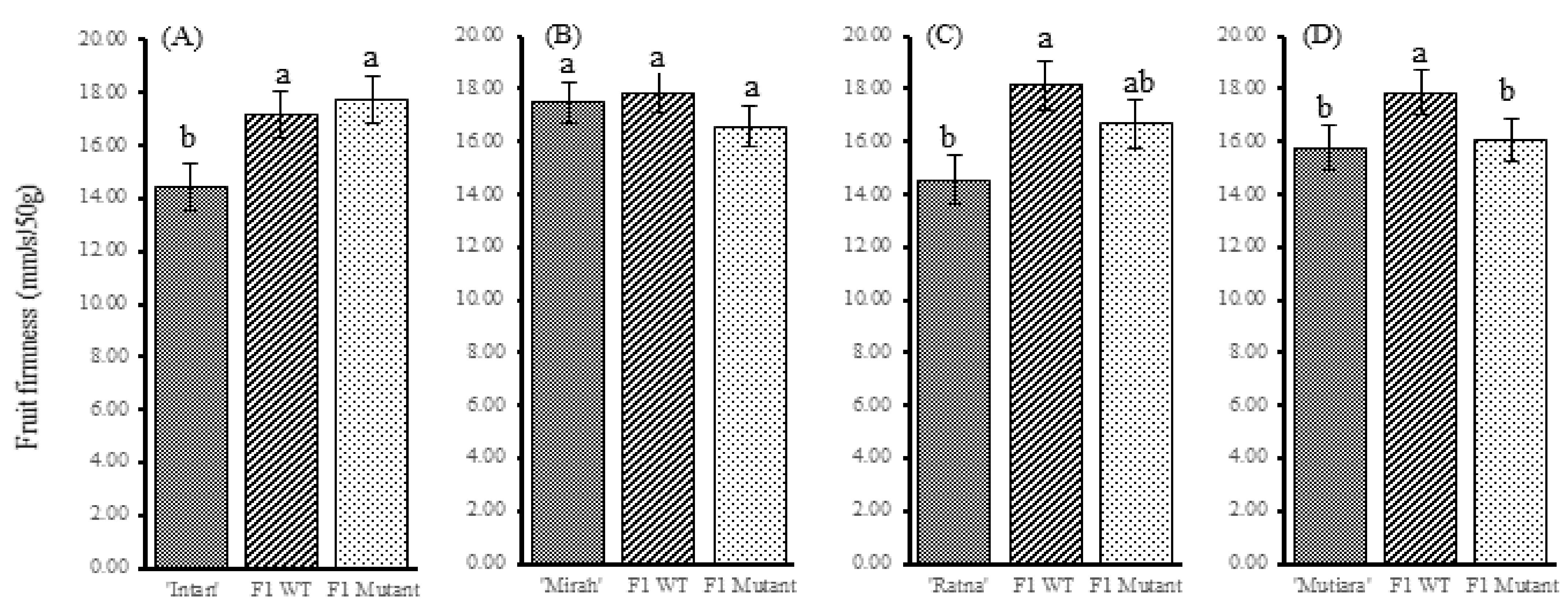
3.4. Effect of the Sletr1-2 Allele on Flowering and Fruit Phenotypes
3.5. Effect of the Sletr1-2 Allele on Yield-Related Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pinela, J.; Barros, L.; Carvalho, A.M.; Ferreira, I.C.F.R. Nutritional composition and antioxidant activity of four tomato (Lycopersicon esculentum L.) farmer’ varieties in Northeastern Portugal homegardens. Food Chem. Toxicol. 2011, 50, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Frusciante, L.; Carli, P.; Ercolano, M.R.; Pernice, R.; Di Matteo, A.; Fogliano, V.; Pellegrini, N. Antioxidant nutritional quality of tomato. Mol. Nutr. Food Res. 2007, 51, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Guil-Guerrero, J.L.; Rebolloso-Fuentes, M.M. Nutrient composition and antioxidant activity of eight tomato (Lycopersicon esculentum) varieties. J. Food Compos. Anal. 2009, 22, 123–129. [Google Scholar] [CrossRef]
- Delian, E.; Chira, A.; Badulescu, L.; Chira, L.L.L.; Bădulescu, L.; Chira, L.L.L. A brief overview of ethylene management to extend the shelf life of tomatoes. Horticulture 2013, 57, 423–428. [Google Scholar]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R.R. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [PubMed]
- Pierik, R.; Sasidharan, R.; Voesenek, L.A.C.J. Growth control by ethylene: Adjusting phenotypes to the environment. J. Plant Growth Regul. 2007, 26, 188–200. [Google Scholar] [CrossRef]
- Schaller, G.E. Ethylene and the regulation of plant development. BMC Biol. 2012, 10, 9. [Google Scholar] [CrossRef]
- Alexander, L.; Grierson, D. Ethylene biosynthesis and action in tomato: A model for climacteric fruit ripening. J. Exp. Bot. 2002, 53, 2039–2055. [Google Scholar] [CrossRef]
- Payasi, A.; Sanwal, G.G. Ripening of Climacteric Fruits and Their Control. J. Food Biochem. 2010, 34, 679–710. [Google Scholar] [CrossRef]
- Wu, X.; Yu, M.; Huan, C.; Ma, R.; Yu, Z. Regulation of the protein and gene expressions of ethylene biosynthesis enzymes under different temperature during peach fruit ripening. Acta Physiol. Plant. 2018, 40, 52. [Google Scholar] [CrossRef]
- Mutari, A.; Debbie, R. The effects of postharvest handling and storage temperature on the quality and shelf of tomato. Afr. J. Food Sci. 2011, 5, 446–452. [Google Scholar]
- Lin, Z.; Zhong, S.; Grierson, D. Recent advances in ethylene research. J. Exp. Bot. 2009, 60, 3311–3336. [Google Scholar] [CrossRef] [PubMed]
- Schaller, B.; Clarke, W.T.; Neubauer, S.; Robson, M.D.; Rodgers, C. Myocardial skeletal muscle signal spoiling using a crusher coil: A human cardiac phosphorus (31P) MR spectroscopic imaging study at 7 Tesla. J. Cardiovasc. Magn. Reson. 2015, 17, 962–972. [Google Scholar] [CrossRef]
- Etana, M.B. Review on the effects of ethylene (C2H4) on quality of fresh fruit and vegetable. The case of banana and tomato. Basic Res. J. Agric. Sci. Rev. 2018, 6, 34–38. [Google Scholar]
- Dubois, M.; Van den Broeck, L.; Inzé, D. The Pivotal Role of Ethylene in Plant Growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Owino, W.O.; Ezura, H. Ethylene Perception and Gene Expression. In Postharvest Biology and Technology of Fruits, Vegetables, and Flowers; Paliyath, G., Murr, D.P., Handa, A.K., Lurie, S., Eds.; Wiley-Black Well: Ames, IA, USA, 2008; Volume 1, pp. 125–138. ISBN 9772081415. [Google Scholar]
- Okabe, Y.; Ariizumi, T.; Ezura, H. Updating the Micro-Tom TILLING platform. Breed. Sci. 2013, 63, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Mubarok, S.; Okabe, Y.; Fukuda, N.; Ariizumi, T.; Ezura, H. Potential Use of a Weak Ethylene Receptor Mutant, Sletr1-2, as Breeding Material To Extend Fruit Shelf Life of Tomato. J. Agric. Food Chem. 2015, 63, 7995–8007. [Google Scholar] [CrossRef] [PubMed]
- Mubarok, S.; Okabe, Y.; Fukuda, N.; Ariizumi, T.; Ezura, H. Favorable effects of the weak ethylene receptor mutation Sletr1-2 on postharvest fruit quality changes in tomatoes. Postharvest. Biol. Technol. 2016, 120, 1–9. [Google Scholar] [CrossRef]
- Pavel, A.B.; Vasile, C.I. PyElph—A software tool for gel images analysis and phylogenetics. BMC Bioinf. 2012, 13, 9. [Google Scholar] [CrossRef]
- Cvikić, D.; Zdravković, J.; Pavlović, N.; Adžić, S.; Dordević, M. Postharvest shelf life of tomato (Lycopersicon esculentum mill.) Mutanats (nor and rin) and their hybrids. Genetika 2012, 44, 449–456. [Google Scholar] [CrossRef]
- Okabe, Y.; Asamizu, E.; Saito, T.; Matsukura, C.; Ariizumi, T.; Brès, C.; Rothan, C.; Mizoguchi, T.; Ezura, H.; Brs, C.; et al. Tomato TILLING technology: Development of a reverse genetics tool for the efficient isolation of mutants from micro-tom mutant libraries. Plant Cell Physiol. 2011, 52, 1994–2005. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Rodríguez, G.R.; Pratta, G.R.; Liberatti, D.R.; Zorzoli, R.; Picardi, L.A. Inheritance of shelf life and other quality traits of tomato fruit estimated from F1’s, F2’s and backcross generations derived from standard cultivar, nor homozygote and wild cherry tomato. Euphytica 2010, 176, 137–147. [Google Scholar] [CrossRef]
- Garg, N.; Cheema, D.S.; Pathak, D. Heterosis breeding in tomato involving rin, nor and alc alleles: A review of literature. Adv. Hortic. Sci. 2008, 22, 54–62. [Google Scholar]
- Adiyoga, W.; Asandhi, A.A.; Laksanawati, A.; Nurhartuti; Sulastrini, I. Rantai Pasokan Sayuran dan Persepsi Partisipan Rantai terhadap Pentingnya Keamanan Pangan (Vegetable Supply Chain and Participants’ Perceptions of the Importance of Food Safety Chain). J. Hortik. 2007, 17, 285–296. [Google Scholar]
- Wijayani, A.; Widodo, W. Usaha Meningkatkan Kualitas Beberapa Varietas Tomat Dengan Sistem Budidaya Hidroponik (Increasing Of Tomatoes Quality In Hydroponic Culture). Ilmu Pertan. 2005, 12, 77–83. [Google Scholar]
- Seymour, G.B.; Chapman, N.H.; Chew, B.L.; Rose, J.K.C. Regulation of ripening and opportunities for control in tomato and other fruits. Plant Biotechnol. J. 2013, 11, 269–278. [Google Scholar] [CrossRef]
- Abera Teka, T.; Teka, T.A. Analysis of the effect of maturity stage on the postharvest biochemical quality characteristics of tomato (Lycopersicon esculentum Mill.) fruit. Int. Res. J. Pharm. Appl. Sci. 2013, 3, 180–186. [Google Scholar]
- Rahayu, Y.D.; Murti, R.H. Pengujian Keunggulan Empat Galur Harapan Tomat (Solanum lycopersicum L.) (Superiority Assessment of Four Tomato Promising Lines (Solanum lycopersicum L.)). Vegetalika 2020, 9, 330. [Google Scholar] [CrossRef][Green Version]
- Purwanti, E.; Jaya, B.; Permadi, A.H. Deskripsi Tomat Varietas Mirah; Decree No. 714/Kpts/TP.240/6/99; Ministry Of Agriculture Republic Indonesia: Jakarta, Indonesia, 22 June 1999; 3p.
- Purwanti, E. Deskripsi Tomat Varietas Mutiara; Decree No. 14/Kpts/TP.240/1/1987; Ministry Of Agriculture Republic Indonesia: Jakarta, Indonesia, 14 January 1987; pp. 130–131.
- Achard, P.; Baghour, M.; Chapple, A.; Hedden, P.; Van Der Straeten, D.; Genschik, P.; Moritz, T.; Harberd, N.P. The plant stress hormone ethylene controls floral transition via DELLA-dependent regulation of floral meristem-identity genes. Proc. Natl. Acad. Sci. USA 2007, 104, 6484–6489. [Google Scholar] [CrossRef] [PubMed]
- Ariizumi, T.; Shinozaki, Y.; Ezura, H. Genes that influence yield in tomato. Breed. Sci. 2013, 63, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Causse, M.; Stevens, R.; Amor, B.B.; Faurobert, M.; Muños, S. Breeding for fruit quality in tomato. In Breeding for Fruit Quality; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 279–305. ISBN 9780813810720. [Google Scholar]
- Ventura Faria, M.; Maluf, W.R.R.; Márcio De Azevedo, S.; Carvalho Andrade-Júnior, V.; Augusto Gomes, L.A.A.; Moretto, P.; Licursi, V. Yield and post-harvest quality of tomato hybrids heterozygous at the loci alcobaça, old gold-crimson or high pigment. Genet. Mol. Res. 2003, 2, 317–327. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Origin | Growth Type | First Harvest (dap) | Fruit Weight (g) | Fruit Diameter (cm) | Pericarp Thickness (cm) | Fruit Weight Loss (20 dah) |
|---|---|---|---|---|---|---|---|
| ‘Intan’ | AVRDC Taiwan, China, introduction variety | Determinate | 80–85 | 40–65 | 3.8–5.0 | 0.3–0.4 | 4.15 |
| ‘Mirah’ | Indonesia local variety | Determinate | 75–85 | 20–40 | 3.0–4.0 | 0.2–0.3 | 6.68 |
| ‘Ratna’ | Philippine, introduction variety | Determinate | 80–85 | 45–85 | 5.0–6.0 | 0.3–0.4 | 5.17 |
| ‘Mutiara’ | Progeny of South American tomato | Indeterminate | 80–90 | 35–50 | 3.8–4.5 | 0.3–0.5 | 7.92 |
| Genotype | Days to Flowering | Days to Maturity |
|---|---|---|
| ‘Intan’ (parent ♀) | 30.25 ± 0.75 a | 81.70 ± 0.68 a |
| ‘Intan’ F1 WT | 28.00 ± 0.41 b | 67.10 ± 0.77 b |
| ‘Intan’ F1 mutant | 28.00 ± 0.41 b | 65.60 ± 0.38 b |
| ‘Mirah’ (parent ♀) | 30.00 ± 0.71 a | 78.80 ± 2.81 a |
| ‘Mirah’ F1 WT | 28.50 ± 0.50 ab | 72.30 ± 1.52 ab |
| ‘Mirah’ F1 mutant | 28.00 ± 0.41 b | 67.45 ± 1.34 b |
| ‘Ratna’ (parent ♀) | 31.00 ± 0.41 a | 82.45 a ± 1.07 a |
| ‘Ratna’ F1 WT | 28.00 ± 0.41 a | 69.35 b ± 0.64 b |
| ‘Ratna’ F1 mutant | 28.00 ± 1.22 a | 68.70 b ± 1.88 b |
| ‘Mutiara’ (parent ♀) | 34.75 ± 1.70 a | 84.65 a ± 1.45 a |
| ‘Mutiara’ F1 WT | 29.00 ± 0.58 b | 75.65 b ± 1.58 b |
| ‘Mutiara’ F1 mutant | 28.00 ± 0.41 b | 69.00 c ± 1.39 c |
| Genotype | Fruit Diameter (cm) | Locule Number | Pericarp Thickness (mm) |
|---|---|---|---|
| ‘Intan’ (parent ♀) | 4.37 ± 0.14 a | 4.60 ± 0.18 a | 3.35 ± 0.16 a |
| ‘Intan’ F1 WT | 4.16 ± 0.11 a | 3.55 ± 0.16 b | 3.04 ± 0.12 ab |
| ‘Intan’ F1 mutant | 3.48 ± 0.13 b | 4.25 ± 0.27 ab | 2.79 ± 0.12 b |
| ‘Mirah’ (parent ♀) | 3.78 ± 0.15 a | 4.33 ± 0.21 a | 2.65 ± 0.13 a |
| ‘Mirah’ F1 WT | 3.71 ± 0.14 a | 4.00 ± 0.17 a | 2.80 ± 0.11 a |
| ‘Mirah’ F1 mutant | 3.58 ± 0.11 a | 4.07 ± 0.12 a | 2.91 ± 0.09 a |
| ‘Ratna’ (parent ♀) | 5.54 ± 0.19 a | 5.93 ± 0.27 a | 3.70 ± 0.21 a |
| ‘Ratna’ F1 WT | 3.53 ± 0.08 b | 4.09 ± 0.14 b | 3.01 ± 0.14 b |
| ‘Ratna’ F1 mutant | 3.30 ± 0.07 b | 3.85 ± 0.95 b | 2.65 ± 0.10 b |
| ‘Mutiara’ (parent ♀) | 4.25 ± 0.15 a | 3.54 ± 0.20 a | 3.98 ± 0.17 a |
| ‘Mutiara’ F1 WT | 3.28 ± 0.08 b | 3.54 ± 0.15 a | 2.70 ± 0.11 b |
| ‘Mutiara’ F1 mutant | 3.57 ± 0.11 b | 4.08 ± 0.23 a | 2.64 ± 0.13 b |
| Genotype | Fruit Weight (g) | Number of Fruit per Plant | Fruit Weight per Plant (g) |
|---|---|---|---|
| ‘Intan’ (parent ♀) | 53.28 ± 3.07 a | 27.30 ± 1.41 b | 1287.52 ± 82.52 a |
| ‘Intan’ F1 WT | 18.24 ± 0.88 b | 47.30 ± 3.45 a | 847.05 ± 69.84 b |
| ‘Intan’ F1 mutant | 18.08 ± 0.64 b | 45.20 ± 2.94 a | 719.45 ± 51.05 b |
| ‘Mirah’ (parent ♀) | 30.21 ± 4.06 a | 44.40 ± 4.38 a | 1105.45 ± 84.53 a |
| ‘Mirah’ F1 WT | 21.63 ± 2.29 b | 47.90 ± 3.62 a | 959.90 ± 76.76 a |
| ‘Mirah’ F1 mutant | 23.58 ± 3.05 b | 44.85 ± 3.63 a | 933.45 ± 61.24 a |
| ‘Ratna’ (parent ♀) | 67.10 ± 6.20 a | 27.05 ± 1.91 b | 1391.20 ± 102.55 a |
| ‘Ratna’ F1 WT | 17.95 ± 1.15 b | 49.95 ± 2.96 a | 823.90 ± 56.44 a |
| ‘Ratna’ F1 mutant | 15.73 ± 0.86 b | 50.60 ± 3.13 a | 723.35 ± 69.06 a |
| ‘Mutiara’ (parent ♀) | 42.97 ± 2.62 a | 36.75 ± 3.01 c | 1394.65 ± 96.74 a |
| ‘Mutiara’ F1 WT | 19.23 ± 0.95 b | 54.20 ± 4.70 b | 940.35 ± 84.13 b |
| ‘Mutiara’ F1 mutant | 21.00 ± 1.46 b | 69.35 ± 4.32 a | 1214.55 ± 117.26 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anas, A.; Wiguna, G.; Damayanti, F.; Mubarok, S.; Setyorini, D.; Ezura, H. Effect of Ethylene Sletr1-2 Receptor Allele on Flowering, Fruit Phenotype, Yield, and Shelf-Life of Four F1 Generations of Tropical Tomatoes (Solanum lycopersicum L.). Horticulturae 2022, 8, 1098. https://doi.org/10.3390/horticulturae8121098
Anas A, Wiguna G, Damayanti F, Mubarok S, Setyorini D, Ezura H. Effect of Ethylene Sletr1-2 Receptor Allele on Flowering, Fruit Phenotype, Yield, and Shelf-Life of Four F1 Generations of Tropical Tomatoes (Solanum lycopersicum L.). Horticulturae. 2022; 8(12):1098. https://doi.org/10.3390/horticulturae8121098
Chicago/Turabian StyleAnas, Anas, Gungun Wiguna, Farida Damayanti, Syariful Mubarok, Dwi Setyorini, and Hiroshi Ezura. 2022. "Effect of Ethylene Sletr1-2 Receptor Allele on Flowering, Fruit Phenotype, Yield, and Shelf-Life of Four F1 Generations of Tropical Tomatoes (Solanum lycopersicum L.)" Horticulturae 8, no. 12: 1098. https://doi.org/10.3390/horticulturae8121098
APA StyleAnas, A., Wiguna, G., Damayanti, F., Mubarok, S., Setyorini, D., & Ezura, H. (2022). Effect of Ethylene Sletr1-2 Receptor Allele on Flowering, Fruit Phenotype, Yield, and Shelf-Life of Four F1 Generations of Tropical Tomatoes (Solanum lycopersicum L.). Horticulturae, 8(12), 1098. https://doi.org/10.3390/horticulturae8121098