
Effects of Potassium Application on Lilium davidii var. unicolor Growth, Polysaccharide Accumulation, and Metabolism

Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Chemicals and Reagents
2.3. Experimental Design
2.4. Determination of Growth Indicators
2.5. Extracts and Determination of Polysaccharide Contents
2.6. HPLC Analysis for Polysaccharide
2.7. Data Analysis
3. Results and Analysis
3.1. Effects of Potassium Application on Growth and Development of L. davidii var. unicolor
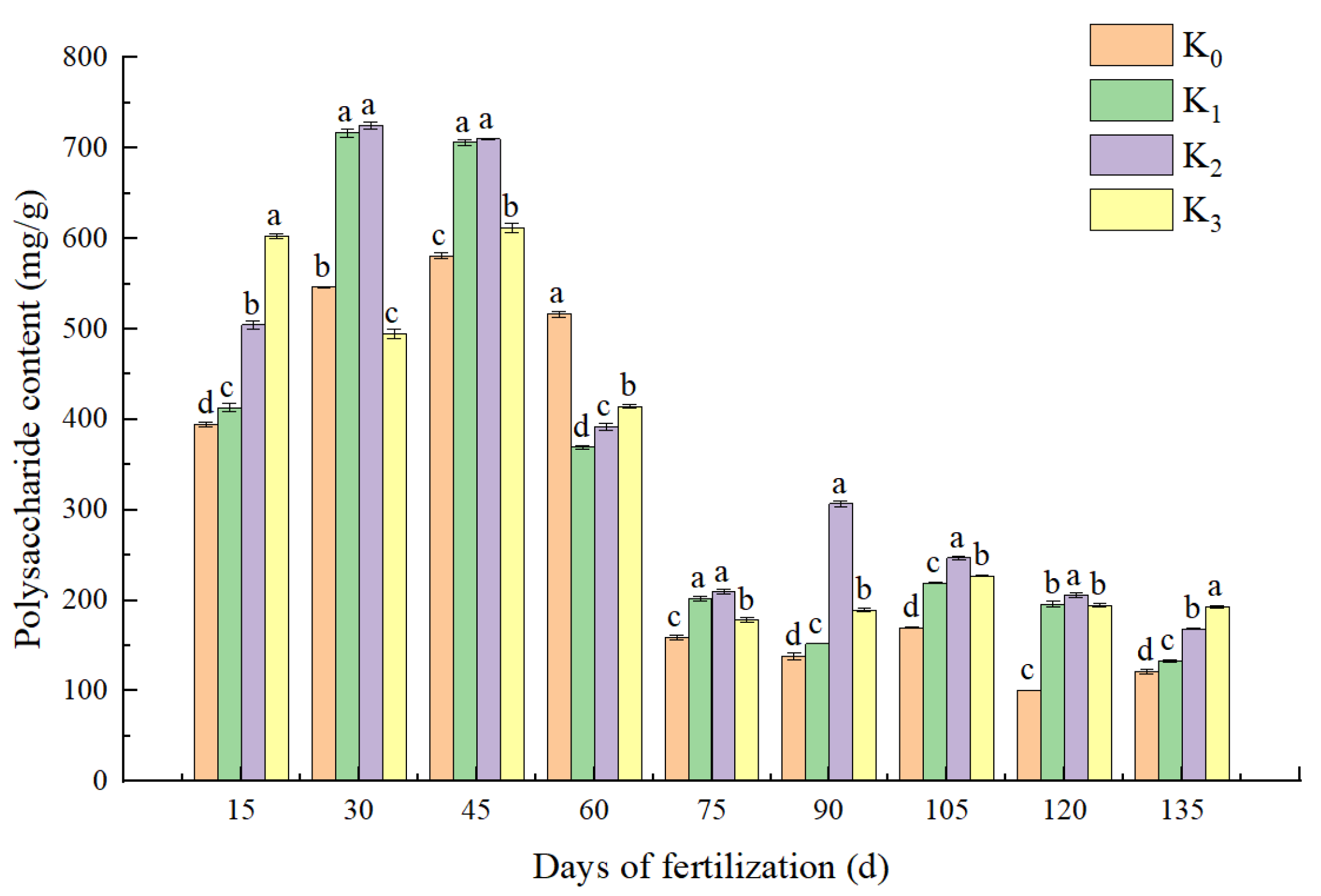
3.2. Effect of Potassium Application on Polysaccharide Content in Bulbs of L. davidii var. unicolor
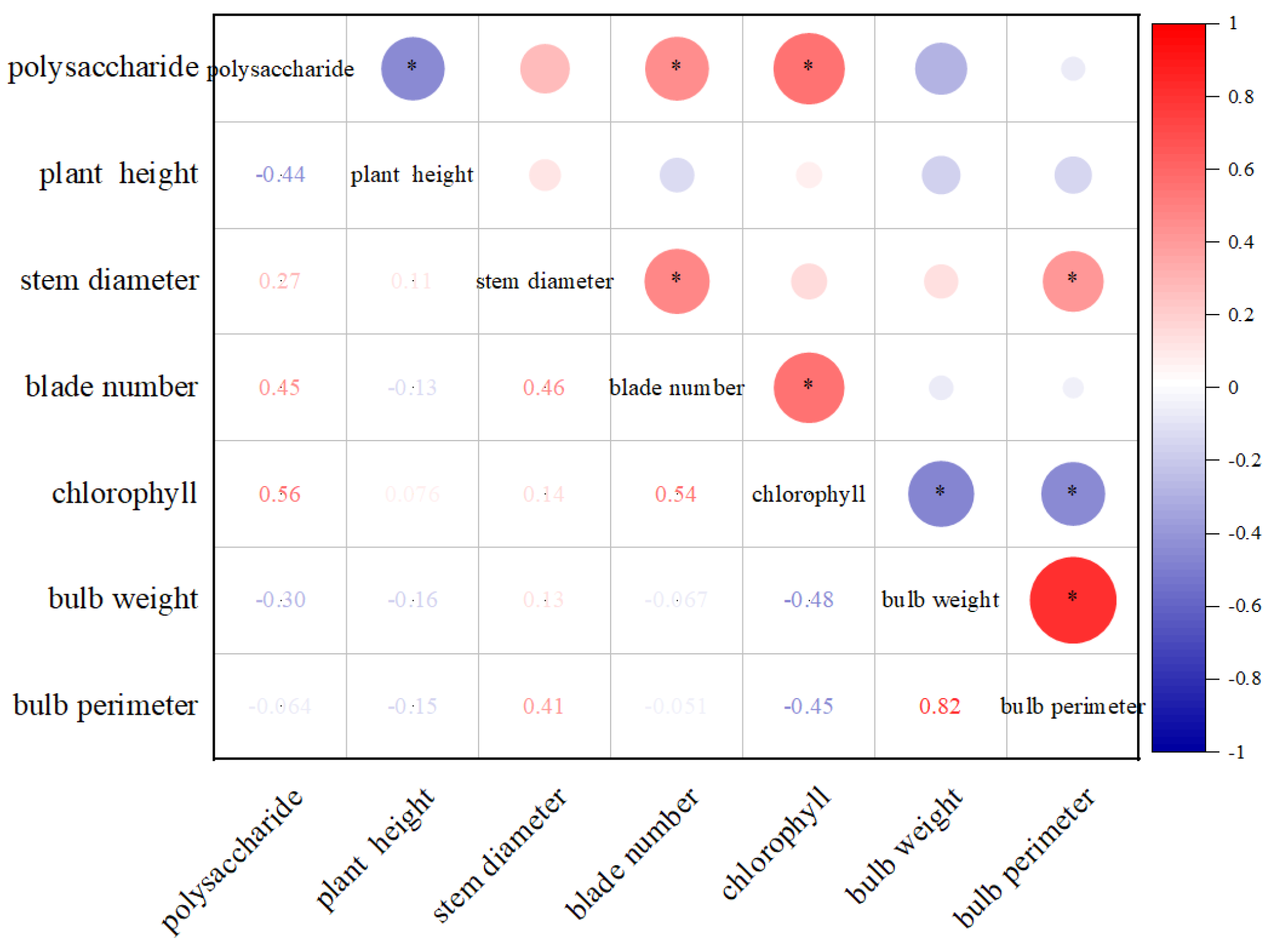
3.3. Correlations between Agronomic Traits and Polysaccharide Contents of L. davidii var. unicolor
3.4. Metabolomics Analysis of Polysaccharides in Bulb Extract of L. davidii var. unicolor
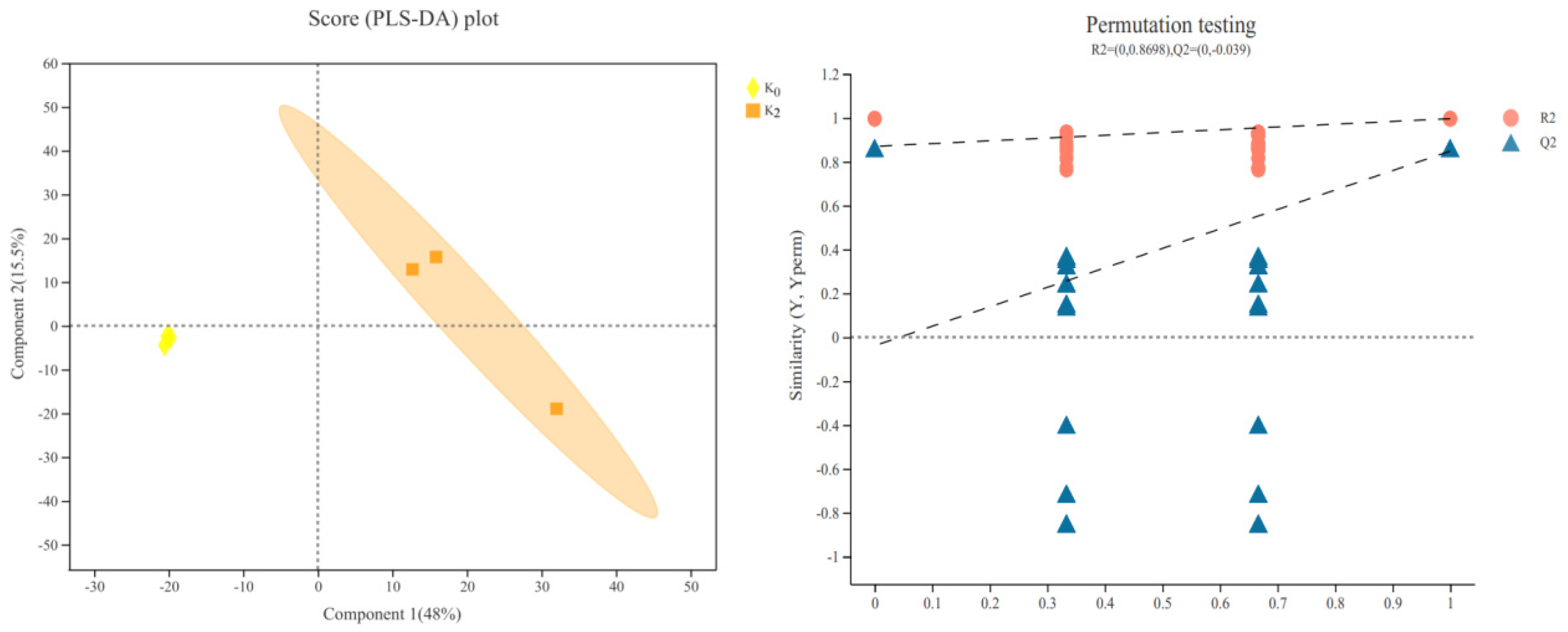
3.4.1. PLS-DA Results
3.4.2. Composition of Polysaccharides and Related Compounds
3.4.3. Differential Metabolites of Polysaccharides and Related Compounds
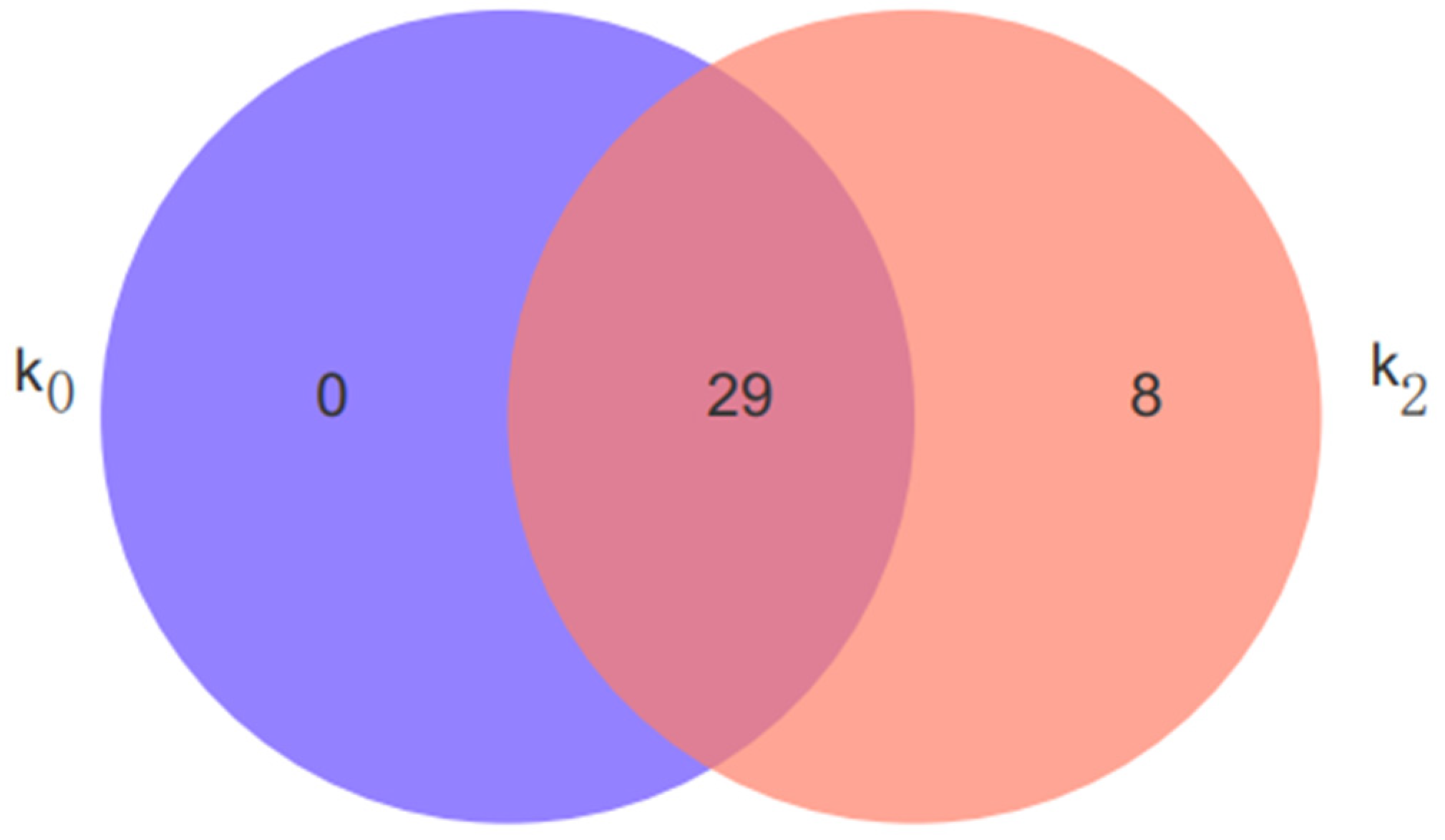
3.4.4. Analysis of Differential Metabolites Using Venn Diagrams
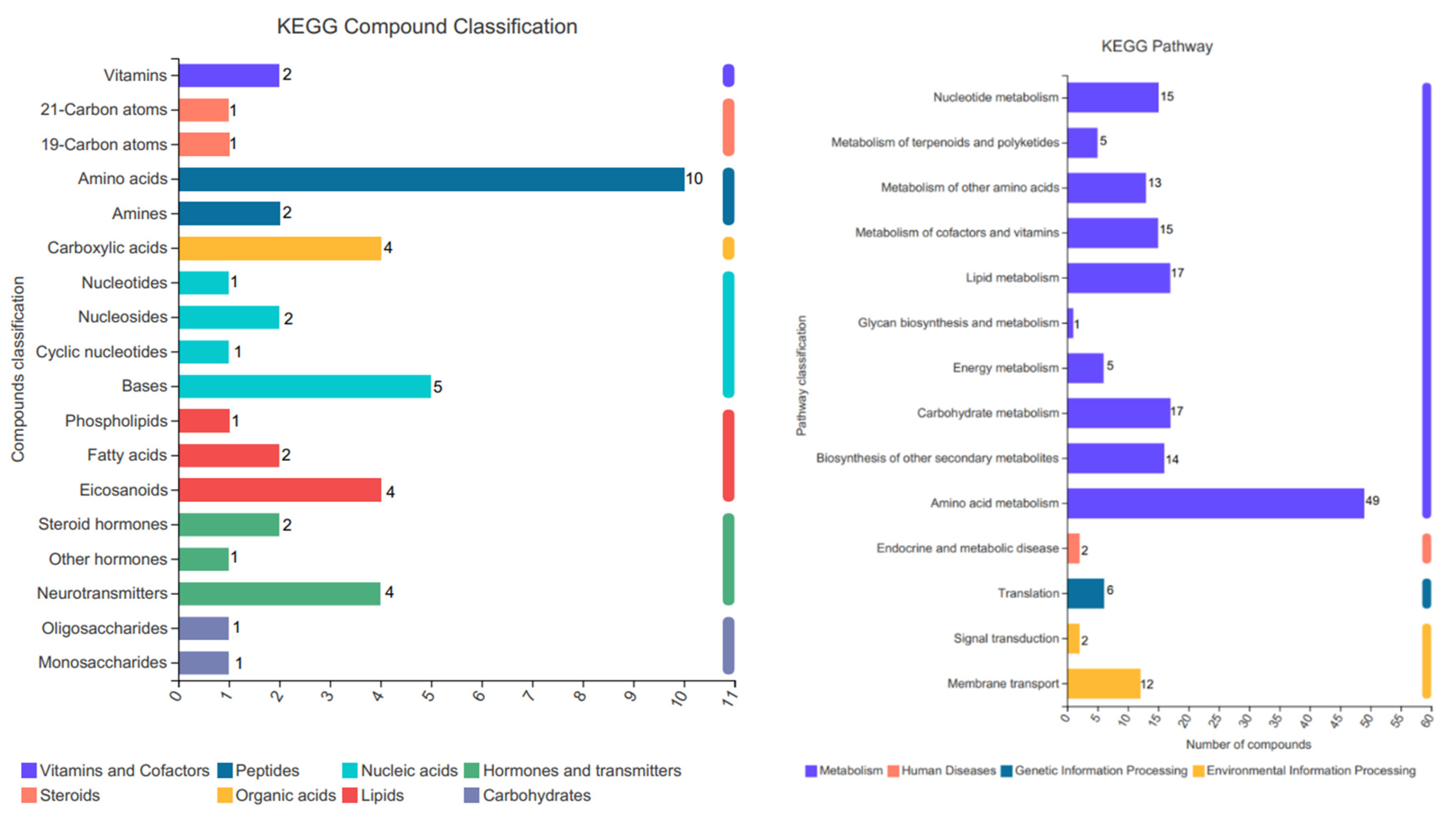
3.4.5. Kyoto Encyclopedia of Genes and Genomes (KEGG) Classifications and Functional Pathways
3.4.6. Analysis of Pathways for Differential Metabolites
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wang, C.L.; Hou, X.M.; Qi, N.N.; Li, C.X.; Luo, Y.Y.; Hu, D.L.; Li, Y.H.; Liao, W.B. An optimized method to obtain high-quality RNA from different tissues in Lilium davidii var. unicolor. Sci. Rep. 2022, 12, 2825. [Google Scholar] [CrossRef] [PubMed]
- Li, W.M.; Wang, Y.J.; Wei, H.L.; Zhang, Y.B.; Guo, Z.H.; Qiu, Y.; Wen, L.R.; Xie, Z.K. Structural characterization of Lanzhou lily (Lilium davidii var. unicolor) polysaccharides and determination of their associated antioxidant activity. J. Sci. Food Agr. 2020, 100, 5603–5616. [Google Scholar] [CrossRef] [PubMed]
- Francis, J.A.; Rumbeiha, W.; Nair, M.G. Constituents in Easter lily flowers with medicinal activity. Life Sci. 2004, 76, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Huang, G.L. Preparation and immunological activity of polysaccharides and their derivatives. Int. J. Biol. Macromol. 2018, 112, 211–216. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, T.; Jin, Z.Y.; Xu, X.M.; Wang, J.H.; Zha, X.Q.; Chen, H.Q. Structural characterisation, physicochemical properties and antioxidant activity of polysaccharide from Lilium lancifolium Thunb. Food Chem. 2015, 169, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Huang, G.L. The antioxidant activities of carboxymethylated garlic polysaccharide and its derivatives. Int. J. Biol. Macromol. 2019, 140, 1054–1063. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.P.; Ma, X.; Ma, L.; Zhang, J.; Zhang, W.M.; Song, X.P. Antioxidative and immunological activities of ophiopogon polysaccharide liposome from the root of Ophiopogon japonicus. Carbohyd. Polym. 2016, 135, 110–120. [Google Scholar] [CrossRef]
- Han, H.P.; Xie, H.C. A study on the extraction and purification process of lily polysaccharide and its anti-tumor effect. Afr. J. Tradit. Complem. 2013, 10, 485–489. [Google Scholar] [CrossRef]
- Pan, G.F.; Xie, Z.W.; Huang, S.X.; Tai, Y.L.; Cai, Q.S.; Jiang, W.; Sun, J.M.; Yuan, Y. Immune-enhancing effects of polysaccharides extracted from Lilium lancifolium Thunb. Int. Immunopharmacol. 2017, 52, 119–126. [Google Scholar] [CrossRef]
- Hou, J.; Zhu, Y.X.; Li, Y.; Zhang, B.N. Synergistic antitumor activity of Lily polysaccharide combined with metformin on human liver cancer hepG2 cells. J. Liaoning Univ. Tradit. Chin. Med. 2017, 19, 30–32. [Google Scholar] [CrossRef]
- Sun, X.; Gao, R.L.; Xiong, Y.K.; Huang, Q.C.; Xu, M. Antitumor and immunomodulatory effects of a water-soluble polysaccharide from Lilii Bulbus in mice. Carbohyd. Polym. 2014, 102, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.; Cui, G.; Feng, Q.; Xiao, Y. Evaluation of Hypoglycemic Activity of the Polysaccharides Extracted from Lycium Barbarum. Afr. J. Tradit. Complem. 2010, 6, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.W.; Chen, Z.Q.; Ramesh, K.S.; Xu, L.L.; Gao, X.D.; Ma, Q.Q.; Xue, Z.H.; Chen, H.X. Hypoglycemic effects of polysaccharides from corn silk (Maydis stigma) and their beneficial roles via regulating the PI3K/Akt signaling pathway in L6 skeletal muscle myotubes. Int. J. Biol. Macromol. 2018, 121, 981–988. [Google Scholar] [CrossRef]
- Zhu, M.D.; Luo, J.; Lv, H.W.; Kong, L.Y. Determination of anti-hyperglycaemic activity in steroidal glycoside rich fraction of lily bulbs and characterization of the chemical profiles by LC-Q-TOF-MS/MS. J. Funct. Foods. 2014, 6, 585–597. [Google Scholar] [CrossRef]
- Furtini, A.E.N.; Boldrin, K.V.F.; Mattson, N.S. Nutrion and quality in ornamental plants Ornam Hortic. J. Hortic. Sci. Biotechnol. 2015, 21, 139–150. [Google Scholar]
- Carneiro, D.N.M.; Almeida, E.F.A.; Paiva, P.D.D.; Frazao, J.E.M.; Santos, F.H.D.; Carneiro, L.F. Carneiro Development and dry mass accumulation in calla lily at the initial cultivation stage. Sci. Agric. Technol. 2011, 35, 1085–1092. [Google Scholar] [CrossRef]
- Carneiro, D.N.M.; Coelho, L.L.; Paiva, P.D.O. Carneiro Evaluation of macronutrient demand in calla lily (Zantedeschia aethiopica). Aust. J. Crop Sci. 2015, 9, 761–766. [Google Scholar]
- Yang, Q.Y.; Zong, J.H.; Huang, P. Growth of edible lily (Lilium davidii var. unicolor) and bulb yield responses to potassium fertiliser and plastic film mulching. J. Hortic. Sci. Biotechnol. 2015, 90, 115–120. [Google Scholar] [CrossRef]
- Mao, Y.F.; Li, Z.L.; Duan, Q.; Du, W.W.; Cui, G.F. Study on the differences of nutrients in four species of lilium. J. Yunnan Agric. Univ. 2017, 32, 366–370. [Google Scholar]
- Qu, W.H.; Zhou, R.B.; He, Y.S.; Tong, Q.Z. Efects of different manure on lily quality. J. Chin. Med. Mater. 2005, 28, 79–81. [Google Scholar] [CrossRef]
- Yin, X.J.; Lin, X.Y.; Liu, Y.X.; Irfan, M.; Chen, L.J.; Zhang, L. Integrated metabolic profiling and transcriptome analysis of pigment accumulation in diverse petal tissues in the lily cultivar “Vivian”. BMC Plant Biol. 2020, 20, 446. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.Y.; Yang, L.P.; Yang, T.J.; Fu, Y.Y.; Han, L.; Quan, J. Cultivation of a new polyploid cultivar ”Yubaihe 1”. Mol. Plant Breed. 2020, 18, 4714–4724. [Google Scholar] [CrossRef]
- Chen, Z.G.; Zhu, Q.; Wang, F. Purification and molecular mass determination of lily polysaccharides. Food Sci. 2013, 34, 1–4. [Google Scholar]
- Song, S.; Liu, X.Y.; Zhao, B.T. Effects of Lactobacillus plantarum Fermentation on the Chemical Structure and Antioxidant Activity of Polysaccharides from Bulbs of Lanzhou Lily. ACS Omega 2021, 6, 29839–29851. [Google Scholar] [CrossRef] [PubMed]
- Bishwoyog, B.; Swarnima, K. Effect of potassium on quality and yield of potato tubers-A Review. Int. J. Agric. Environ. 2016, 3, 7–12. [Google Scholar]
- Slameto; Leni, N.; Indri, F.; Riza, Y.; Kacung, H. Effect of potassium fertilizer on growth, capsaicin and ascorbic acid content of local and hybrid chili (Capsicum annum L.). Plant Cell Biotechnol. Mol. Biol. 2021, 22, 337–345. [Google Scholar]
- Soliman, S.S.; Alebidia, A.I.; Al-Obeed, R.S.; Al-Saif, A.M. Effect of potassium fertilizer on fruit quality and mineral composition of fig (Ficus carica L. cv. Brown Turky). Pak. J. Bot. 2018, 50, 1753–1758. [Google Scholar]
- Wang, M.M.; Ye, Y.L.; Chu, X.; Zhao, Y.A.; Zhang, S.H.; Chen, H.; Qin, W.; Wang, Y. Responses of garlic quality and yields to various types and rates of potassium fertilizer applications. Hortscience 2022, 57, 72–80. [Google Scholar] [CrossRef]
- Huu, N.H.; Maneepong, S.; Suranin, P.P. Effects of potassium, calcium, and magnesium ratios in soil on their uptake and fruit quality of pummelo. J. Agr. Sci. Tech.-Iran. 2017, 9, 110. [Google Scholar] [CrossRef]
- Li, Q. Effects of Combined Application of Potassium Fertilizer and Organic Fertilizer on Soil Enzyme Activity, Soil Nutrients and Yield of Edible Lily; Gansu Agricultural University: Gansu, China, 2018. [Google Scholar]
- Zhu, H.; Wang, Z.J.; Wu, Y.L.; Jiang, H.T.; Zhou, F.; Xie, X.H.; Wang, R.L.; Hua, C. Untargeted metabonomics reveals intervention effects of chicory polysaccharide in a rat model of non-alcoholic fatty liver disease. Int. J. Biol. Macromol. 2019, 128, 363–375. [Google Scholar] [CrossRef]
- Zhou, J.; An, R.F.; Huang, X.F. Genus Lilium: A review on traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2021, 270, 113852. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Z.; Zayed, O.; Zeng, F.S.; Liu, C.X.; Zhang, l.; Zhu, P.P.; Hsu, C.C.; Tuncil, Y.E.; Tao, W.E.; Carpita, N.C.; et al. Arabinose biosynthesis is critical for salt stress tolerance in Arabidopsis. New Phytol. 2019, 224, 274–290. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Wang, H.; Lang, L.X.; Dou, X.Y.; Bai, J.R. Analysis of the contents of 13 intrinsic free sugars in different lily samples. Food Sci. 2021, 42, 249–254. [Google Scholar]
- Lei, L.; Wu, T.X.; Wang, C.N. Analysis of metabolic differences in fermentation of Grifola frondosa based on UPLC-QTOF-MS metabolomics. Mycosystema 2020, 39, 1920–1932. [Google Scholar] [CrossRef]
- Zhang, H.M.; Liu, J.Y. Molecular cloning and characterization of a beta-galactosidase gene expressed preferentially in cotton fibers. J. Integr. Plant Biol. 2005, 47, 223–232. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | K0 | K1 | K2 | K3 |
|---|---|---|---|---|
| N (609.8 mg·L−1) +P (88.4 mg·L−1) | 0 | 447.6 | 671.4 | 895.2 |
| Days after Fertilization | Treatment | Plant Height (cm) | Stem Diameter (mm) | Blade Number (Number) | Chlorophyll | Bulb Weight (g) | Bulb Circumference (cm) |
|---|---|---|---|---|---|---|---|
| K0 | 31.22 ± 0.55 b | 7.25 ± 0.13 b | 208.67 ± 8.09 b | 41.90 ± 1.18 c | 20.27 ± 0.97 b | 11.00 ± 0.15 b | |
| K1 | 35.24 ± 0.55 a | 7.48 ± 0.12 ab | 213.00 ± 7.09 b | 47.27 ± 0.69 b | 23.73 ± 0.33 a | 12.43 ± 0.55 a | |
| 15 | K2 | 35.56 ± 0.45 a | 8.19 ± 0.40 a | 243.67 ± 9.33 a | 56.67 ± 2.08 a | 22.23 ± 0.46 a | 11.47 ± 0.30 b |
| K3 | 32.68 ± 0.54 b | 7.69 ± 0.21 ab | 194.00 ± 8.72 b | 57.10 ± 1.47 a | 17.53 ± 0.44 c | 11.70 ± 0.45 b | |
| K0 | 43.52 ± 0.58 c | 6.92 ± 0.07 b | 209.00 ± 5.03 c | 43.73 ± 0.32 c | 14.53 ± 0.49 b | 11.03 ± 0.44 a | |
| K1 | 49.44 ± 1.23 b | 6.69 ± 0.05 b | 233.33 ± 6.64 b | 60.70 ± 0.25 b | 21.53 ± 0.56 a | 10.93 ± 0.72 b | |
| 30 | K2 | 53.96 ± 0.80 a | 7.98 ± 0.45 a | 254.67 ± 8.01 a | 66.60 ± 2.51 a | 22.40 ± 0.99 a | 12.07 ± 0.29 a |
| K3 | 49.94 ± 0.52 b | 7.18 ± 0.21 ab | 165.67 ± 5.55 d | 63.20 ± 1.20 ab | 16.53 ± 0.55 b | 10.83 ± 0.20 b | |
| K0 | 50.22 ± 0.97 c | 6.51 ± 0.04 b | 163.67 ± 9.60 c | 62.20 ± 1.01 b | 13.00 ± 0.31 b | 10.07 ± 0.15 b | |
| K1 | 57.60 ± 1.04 b | 8.19 ± 0.27 a | 196.00 ± 1.00 b | 64.77 ± 0.52 ab | 13.83 ± 0.50 b | 10.40 ± 0.35 ab | |
| 45 | K2 | 62.88 ± 0.63 a | 8.27 ± 0.35 a | 244.67 ± 6.23 a | 65.64 ± 1.04 ab | 16.03 ± 0.48 a | 11.53 ± 0.90 a |
| K3 | 57.62 ± 1.21 b | 7.02 ± 0.19 b | 192.00 ± 9.50 b | 67.50 ± 1.70 a | 15.33 ± 0.49 a | 10.40 ± 0.95 ab | |
| K0 | 60.48 ± 0.28 c | 6.93 ± 0.25 b | 156.67 ± 5.55 c | 60.83 ± 1.50 c | 7.83 ± 0.09 d | 7.88 ± 0.20 b | |
| K1 | 65.64 ± 0.52 b | 7.44 ± 0.27 b | 246.33 ± 7.84 a | 60.87 ± 0.79 c | 9.30 ± 0.58 c | 8.57 ± 0.26 b | |
| 60 | K2 | 72.76 ± 1.84 a | 9.09 ± 0.10 a | 235.67 ± 4.33 a | 71.47 ± 0.30 a | 13.57 ± 0.23 a | 10.03 ± 0.28 a |
| K3 | 64.10 ± 0.96 b | 6.95 ± 0.25 b | 205.33 ± 8.51 b | 66.67 ± 1.11 b | 11.57 ± 0.60 b | 9.83 ± 0.24 a | |
| K0 | 63.10 ± 0.51 c | 6.27 ± 0.04 b | 174.33 ± 7.45 b | 48.90 ± 3.39 b | 11.80 ± 0.31 d | 9.67 ± 0.13 | |
| K1 | 67.92 ± 0.29 b | 7.37 ± 0.43 ab | 213.33 ± 6.69 a | 58.83 ± 0.81 a | 13.00 ± 0.06 c | 9.83 ± 0.28 ab | |
| 75 | K2 | 74.56 ± 1.22 a | 8.19 ± 0.71 a | 211.00 ± 7.55 a | 65.17 ± 1.13 a | 16.27 ± 0.28 a | 10.60 ± 0.29 a |
| K3 | 62.40 ± 0.51 c | 6.13 ± 0.15 b | 175.00 ± 3.21 b | 59.47 ± 1.31 a | 14.97 ± 0.34 b | 9.60 ± 0.20 b | |
| K0 | 61.80 ± 0.49 ab | 5.67 ± 011 c | 173.00 ± 7.09 b | 50.03 ± 0.17 b | 20.60 ± 0.61 b | 10.50 ± 0.15 b | |
| K1 | 65.00 ± 2.35 a | 6.77 ± 0.31 b | 203.33 ± 7.62 a | 56.57 ± 1.19 a | 19.37 ± 0.27 bc | 10.67 ± 0.09 b | |
| 90 | K2 | 63.50 ± 0.61 ab | 8.12 ± 0.25 a | 184.67 ± 7.54 ab | 56.20 ± 0.46 a | 23.37 ± 0.43 a | 11.60 ± 0.17 a |
| K3 | 60.80 ± 0.46 b | 7.37 ± 0.32 ab | 184.00 ± 3.46 ab | 55.30 ± 0.85 a | 18.30 ± 0.61 c | 11.37 ± 0.17 a | |
| K0 | 59.10 ± 1.05 c | 6.84 ± 0.08 a | 201.33 ± 9.53 a | 54.37 ± 0.58 ab | 22.27 ± 0.56 b | 10.27 ± 0.19 b | |
| K1 | 63.02 ± 0.45 b | 6.96 ± 0.04 a | 204.00 ± 7.51 a | 53.57 ± 0.55 b | 22.87 ± 0.20 b | 10.90 ± 0.20 ab | |
| 105 | K2 | 68.98 ± 0.62 a | 7.01 ± 0.11 a | 188.00 ± 3.79 ab | 56.53 ± 0.15 a | 23.30 ± 0.38 a | 11.67 ± 0.09 a |
| K3 | 58.58 ± 0.91 c | 6.50 ± 0.14 b | 165.33 ± 6.36 b | 54.13 ± 1.13 b | 22.50 ± 0.42 b | 11.40 ± 0.35 a | |
| K0 | 52.88 ± 0.47 b | 6.83 ± 0.19 b | 162.67 ± 0.88 a | 27.47 ± 0.22 b | 17.37 ± 0.73 b | 10.63 ± 0.13 c | |
| K1 | 62.70 ± 0.32 a | 7.25 ± 0.21 b | 154.67 ± 2.33 b | 30.10 ± 0.35 a | 22.20 ± 0.55 a | 11.90 ± 0.31 b | |
| 120 | K2 | 66.90 ± 0.43 a | 7.96 ± 0.14 a | 153.33 ± 0.88 b | 29.23 ± 0.32 a | 23.90 ± 0.62 a | 13.33 ± 0.26 a |
| K3 | 51.68 ± 3.78 b | 6.74 ± 0.11 b | 152.33 ± 1.76 b | 25.93 ± 0.84 b | 17.73 ± 0.38 b | 11.13 ± 0.23 bc | |
| K0 | 59.34 ± 0.32 c | 7.27 ± 0.37 b | 190.00 ± 7.00 a | 26.00 ± 0.29 b | 19.57 ± 0.33 b | 10.93 ± 0.09 c | |
| K1 | 61.44 ± 0.22 b | 7.64 ± 0.14 b | 183.67 ± 7.67 b | 28.60 ± 0.49 a | 22.23 ± 1.04 a | 11.97 ± 0.30 b | |
| 135 | K2 | 69.44 ± 0.70 a | 8.58 ± 0.09 a | 171.00 ± 3.06 b | 25.77 ± 0.32 b | 24.00 ± 0.36 a | 13.73 ± 0.12 a |
| K3 | 59.50 ± 0.38 c | 6.44 ± 0.28 c | 129.33 ± 6.77 b | 23.20 ± 0.52 c | 19.73 ± 0.90 b | 11.03 ± 0.15 bc |
| Metabolite | VIP | FC | U/D |
|---|---|---|---|
| Maltopentaose | 2.06 | 1.38 | U |
| Maltotetraose | 0.37 | 1.08 | U |
| 1-Kestose | 1.57 | 1.12 | U |
| B-D-Xylopyranosyl-(1->4)-a-L-rhamnopyranosyl-(1->2)-D-fucose | 0.69 | 0.97 | D |
| Trans-p-Menthane-1,7,8-triol 8-glucoside | 1.84 | 1.15 | U |
| (3b,7b,22x)-Cucurbita-5,24-diene-3,7,23-triol 7-glucoside | 2.06 | 1.29 | U |
| L-Citronellol glucoside | 0.56 | 1.36 | U |
| Linalool oxide D 3-[apiosyl-(1->6)-glucoside] | 1.38 | 1.08 | U |
| Kojibiose | 2.07 | 1.40 | U |
| Alpha-D-Xylopyranosyl-(1->6)-beta-D-glucopyranosyl-(1->4)-D-glucose | 0.54 | 1.05 | U |
| Stachyose | 2.14 | 1.31 | U |
| Alpha-Hydrojuglone 4-O-b-D-glucoside | 1.13 | 0.92 | D |
| Formononetin 7-(6″-malonylglucoside) | 0.86 | 1.11 | U |
| (1xi,2xi)-1-(4-Hydroxyphenyl)-1,2,3-propanetriol 3-O-beta-D-Glucopyranoside | 0.37 | 1.36 | U |
| N-Acetyllactosamine | 0.84 | 1.06 | U |
| Taraxacoside | 1.45 | 0.84 | D |
| Cis-p-Coumaric acid 4-[apiosyl-(1->2)-glucoside] | 0.78 | 0.36 | D |
| Glycerol 2-(9Z,12Z-octadecadienoate) 1-hexadecanoate 3-O-[alpha-D-galactopyranosyl-(1->6)-beta-D-galactopyranoside] | 1.73 | 0.89 | D |
| B-Chlorogenin 3-[4″-(2″-glucosyl-3″-xylosylglucosyl)galactoside] | 1.02 | 0.92 | D |
| Blumenol C glucoside | 0.25 | 1.01 | U |
| Methyl (3x,4E,10R)-3,10-dihydroxy-4,11-dodecadiene-6,8-diynoate 10-glucoside | 1.02 | 1.09 | U |
| 1-O-E-Cinnamoyl-(6-arabinosylglucose) | 0.80 | 1.06 | U |
| 3-Hydroxy-beta-ionol 3-[glucosyl-(1->6)-glucoside] | 1.27 | 1.11 | U |
| 1-O-Feruloyl-beta-D-glucose | 0.60 | 1.03 | U |
| L-DOPA 3′-glucoside | 0.45 | 0.96 | D |
| Todatriol glucoside | 0.90 | 1.08 | U |
| Isopropyl beta-D-glucoside | 0.18 | 0.99 | D |
| 3-Fucosyllactose | 3.14 | 1.90 | U |
| Beta-D-Xylopyranosyl-(1->5)-alpha-L-arabinofuranosyl-(1->3)-L-arabinose | 1.13 | 1.11 | U |
| Maltohexaose | 0.41 | 1.01 | U |
| A-L-Fucopyranosyl-(1->2)-b-D-galactopyranosyl-(1->2)-D-xylose | 1.47 | 0.83 | D |
| Beta-D-Fructofuranosyl alpha-D-glucopyranosyl-(1->4)-D-glucopyranoside | 1.67 | 1.23 | U |
| Maltotriose | 1.73 | 1.18 | U |
| Prenyl glucoside | 1.86 | 1.38 | U |
| (S)-a-Amino-2,5-dihydro-5-oxo-4-isoxazolepropanoic acid N2-glucoside | 0.91 | 0.54 | D |
| Maclurin 3-C-(6″-p-hydroxybenzoyl-glucoside) | 1.06 | 0.93 | D |
| CDP-glucose | 1.41 | 0.87 | D |
| Pathway ID | Pathway | First Category | Second Category |
|---|---|---|---|
| map00052 | Galactose metabolism | Metabolism | Carbohydrate metabolism |
| map00520 | Amino sugar and nucleotide sugar metabolism | Metabolism | Carbohydrate metabolism |
| map00500 | Starch and sucrose metabolism | Metabolism | Carbohydrate metabolism |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sha, X.; Zhang, P.; Yang, Y.; Bu, H.; Ma, Y.; Jin, L. Effects of Potassium Application on Lilium davidii var. unicolor Growth, Polysaccharide Accumulation, and Metabolism. Horticulturae 2022, 8, 940. https://doi.org/10.3390/horticulturae8100940
Sha X, Zhang P, Yang Y, Bu H, Ma Y, Jin L. Effects of Potassium Application on Lilium davidii var. unicolor Growth, Polysaccharide Accumulation, and Metabolism. Horticulturae. 2022; 8(10):940. https://doi.org/10.3390/horticulturae8100940
Chicago/Turabian StyleSha, Xiaorong, Ping Zhang, Ying Yang, Hubai Bu, Ying Ma, and Lei Jin. 2022. "Effects of Potassium Application on Lilium davidii var. unicolor Growth, Polysaccharide Accumulation, and Metabolism" Horticulturae 8, no. 10: 940. https://doi.org/10.3390/horticulturae8100940
APA StyleSha, X., Zhang, P., Yang, Y., Bu, H., Ma, Y., & Jin, L. (2022). Effects of Potassium Application on Lilium davidii var. unicolor Growth, Polysaccharide Accumulation, and Metabolism. Horticulturae, 8(10), 940. https://doi.org/10.3390/horticulturae8100940