
In Vitro Propagation of Cremastra appendiculata var. variabilis by Asymbiotic Seed Germination




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Surface Sterilization
2.2. In Vitro Seed Germination
2.3. Secondary Protocorm Formation and Conversion
2.4. Protocorm Conversion and SLC Development
2.5. Statistical Analysis
3. Results
3.1. Influence of Culture Medium and Seed Age on Asymbiotic SLC Germination
3.2. Impact of AC on Asymbiotic SLC Seed Germination
3.3. Impact of Auxins on Asymbiotic SLC Seed Germination
3.4. Impact of KIN and NAA on Asymbiotic SLC Seed Germination
3.5. Effects of BA and KIN on Secondary Protocorm Formation and Conversion
3.6. Impact of GA3 on the Conversion of SLC Protocorms
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chung, M.Y.; Lopez-Pujol, J.; Moon, M.O.; Maki, M.; Yukawa, T.; Sugiura, N.; Chung, M.G. Population history of the terrestrial orchid Cremastra appendiculata var. variabilis from Korea, inferred from levels and distribution of genetic diversity. Bot. J. Linn. Soc. 2013, 173, 721–732. [Google Scholar] [CrossRef][Green Version]
- Sugiura, N. Pollination biology of Cremastra appendiculata var. variabilis (Orchidaceae). Plant Species Biol. 1996, 11, 185–187. [Google Scholar] [CrossRef]
- Chung, M.Y.; López-Pujol, J.; Son, S.; Suh, G.U.; Yukawa, T.; Chung, M.G. Patterns of genetic diversity in rare and common orchids focusing on the Korean peninsula: Implications for conservation. Bot. Rev. 2018, 84, 1–25. [Google Scholar] [CrossRef]
- Park, H.Y.; Kim, D.H.; Sivanesan, I. Micropropagation of Ajuga species: A mini review. Biotechnol. Lett. 2017, 39, 1291–1298. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Kim, K.-S.; Ak, G.; Zengin, G.; Cziáky, Z.; Jekő, J.; Adaikalam, K.; Song, K.; Kim, D.H.; Sivanesan, I. Establishment of a rapid micropropagation system for Kaempferia parviflora wall. Ex Baker: Phytochemical analysis of leaf extracts and evaluation of biological activities. Plants 2021, 10, 698. [Google Scholar] [CrossRef]
- Zhang, M.S.; Wu, S.J.; Jie, X.J.; Zhang, L.X.; Jiang, X.H.; Du, J.C.; Qi, J.L.; Liu, Z.; Yang, Y.H. Effect of endophyte extract on micropropagation of Cremastra appendiculata (D. Don) Makino (Orchidaceae). Propag. Ornam. Plants 2006, 6, 83–89. [Google Scholar]
- Gao, Y.Y.; Peng, S.J.; Hang, Y.; Xie, G.F.; Ji, N.; Zhang, M.S. Mycorrhizal fungus Coprinellus disseminatus influences seed germination of the terrestrial orchid Cremastra appendicualta (D Don). Makino. Sci. Hortic. 2022, 293, 110724. [Google Scholar] [CrossRef]
- Yang, N.; Wang, D.; Gao, Y.; Hu, E.; Yu, X.; Peng, S.; Ji, J.; Zhang, M.S. An efficient micropropagation protocol, chemical components, and hypoglycemic activity for Cremastra appendiculata (D. Don) Makino pseudobulbs. In Vitro Cell. Dev. Biol. Plant 2022, 58, 213–224. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Kahraman, M.U.; Cullum, F.J. Asymbiotic Germination and Seedling Development of Terrestrial Orchid Bletilla striata Using in vitro and ex vitro Cultures. Hortic. Stud. 2021, 38, 1–14. [Google Scholar] [CrossRef]
- Ferreira, W.D.M.; Oliveira, A.M.D.; Viana, J.C.; Suzuki, R.M.; Oliveira, J.R.G.D. Asymbiotic germination, initial development in vitro and acclimatization of Cyrtopodium paludicolum Hoehne, a Brazilian Savanna orchid species. Rodriguésia 2022, 73, e01272020. [Google Scholar] [CrossRef]
- Barrientos, B.A.B.; Fang, J.Y. Influence of photoperiod and culture medium on the speed of asymbiotic seed germination and seedling development in Spathoglottis plicata. HortScience 2019, 54, 1570–1575. [Google Scholar] [CrossRef]
- Jabin Bello-Bello, J.; Zavala-Ruiz, J.; Cruz-Huerta, N.; Baltazar-Bernal, O. In vitro germination and development of the trumpetist orchid (Myrmecophila grandiflora Lindl.) using ebb-and-flow bioreactor. Propag. Ornam. Plants 2020, 20, 88–95. [Google Scholar]
- Yao, L.; Huang, J.; Zhang, S. An Improved Protocol for Asymbiotic Seed Germination and Seedling Development of Paphiopedilum tigrinum. Horticulturae 2021, 7, 298. [Google Scholar] [CrossRef]
- Arcidiacono, M.; Catalano, C.; Motisi, A.; Sajeva, M.; Carimi, F.; Carra, A. Influence of Culture Conditions on In vitro Asymbiotic Germination of Anacamptis longicornu and Ophrys panormitana (Orchidaceae). Plants 2021, 10, 2543. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Camarena, M.; Ortega-Larrocea, M.P. Mesoamerican Cypripedium: Mycorrhizal Contributions to Promote Their Conservation as Critically Endangered Species. Plants 2022, 11, 1554. [Google Scholar] [CrossRef]
- Mayo-Mosqueda, A.; García-Hernández, E.; Noguera-Savelli, E.; Cetzal-Ix, W.; Alatorre-Cobos, F. Advances in Breeding, Bioprospecting, and In vitro Culture of Laelia Orchid Species. Horticulturae 2022, 8, 103. [Google Scholar] [CrossRef]
- Kaur, S. In vitro Propagation of Vanda testacea (Lindl.) Reichb. F, a Medicinally Important Threatened Orchid. Plant Tissue Cult. Biotechnol. 2021, 31, 153–160. [Google Scholar] [CrossRef]
- Bazzicalupo, M.; Calevo, J.; Adamo, M.; Giovannini, A.; Copetta, A.; Cornara, L. Seed Micromorphology, In vitro Germination, and Early-Stage Seedling Morphological Traits of Cattleya purpurata (Lindl. & Paxton) Van den Berg. Horticulturae 2021, 7, 480. [Google Scholar]
- An, J.; Kim, P.B.; Park, H.B.; Kim, S.; Park, H.J.; Lee, C.W.; Lee, B.-D.; Kim, N.Y.; Hwang, J.E. Effects of Different Growth Media on In vitro Seedling Development of an Endangered Orchid Species Sedirea japonica. Plants 2021, 10, 1193. [Google Scholar] [CrossRef]
- Calevo, J.; Copetta, A.; Marchioni, I.; Bazzicalupo, M.; Pianta, M.; Shirmohammadi, N.; Cornara, L.; Giovannini, A. The use of a new culture medium and organic supplement to improve in vitro early stage development of five orchid species. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2022, 1, 143–151. [Google Scholar] [CrossRef]
- Kim, D.H.; Kang, K.W.; Enkhtaivan, G.; Jan, U.; Sivanesan, I. Impact of activated charcoal, culture medium strength and thidiazuron on non-symbiotic in vitro seed germination of Pecteilis radiata (Thunb.) Raf. S. Afr. J. Bot. 2019, 124, 144–150. [Google Scholar] [CrossRef]
- Knudson, L. A new nutrient solution for germination of orchid seed. Am. Orchid. Soc. Bull. 1946, 15, 214–217. [Google Scholar]
- Kang, H.; Kang, K.W.; Kim, D.H.; Sivanesan, I. In vitro Propagation of Gastrochilus matsuran (Makino) Schltr., an Endangered Epiphytic Orchid. Plants 2020, 9, 524. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Gopal, J.; Sivanesan, I. Nanomaterials in plant tissue culture: The disclosed and undisclosed. RSC Adv. 2017, 7, 36492–36505. [Google Scholar] [CrossRef]
- Dutra, D.; Johnson, T.R.; Kauth, P.J.; Stewart, S.L.; Kane, M.E.; Richardson, L. Asymbiotic seed germination, in vitro seedling development, and greenhouse acclimatization of the threatened terrestrial orchid Bletia purpurea. Plant Cell Tissue Organ Cult. 2008, 94, 11–21. [Google Scholar] [CrossRef]
- Tian, L.; Gao, Y.Y.; Yang, N.X.; Peng, S.J.; Zhang, M.S. Effects of plant growth regulators for seed embryos development of Cremastra appendiculata. Mol. Plant Breed. 2021, 19, 3090–3095. [Google Scholar]
- Lee, Y.-I. The asymbiotic seed germination of six Paphiopedilum species in relation to the time of seed collection and seed pretreatment. Acta Hortic. 2007, 755, 381–386. [Google Scholar] [CrossRef]
- Pant, B.; Swar, S. Micropropagation of Cymbidium iridioides. Nepal. J. Sci. Technol. 2011, 12, 91–96. [Google Scholar] [CrossRef]
- Kunakhonnuruk, B.; Inthima, P.; Kongbangkerd, A. In vitro propagation of Epipactis flava Seidenf, an endangered rheophytic orchid: A first study on factors affecting asymbiotic seed germination, seedling development and greenhouse acclimatization. Plant Cell Tissue Organ Cult. 2018, 135, 419–432. [Google Scholar] [CrossRef]
- Xu, X.; Fang, L.; Li, L.; Ma, G.; Wu, K.; Zeng, S. Abscisic Acid Inhibits Asymbiotic Germination of Immature Seeds of Paphiopedilum armeniacum. Int. J. Mol. Sci. 2020, 21, 9561. [Google Scholar] [CrossRef] [PubMed]
- Kauth, P.J.; Dutra, D.; Johnson, T.R.; Stewart, S.L.; Kane, M.E.; Vendrame, V. Techniques and applications of in vitro orchid seed germination. In Floriculture, Ornamental and Plant Biotechnology: Advances and Topics Issues; da Silva, J.A.T., Ed.; Global Science Books: Isleworth, UK, 2008; pp. 375–391. [Google Scholar]
- Thomas, T.D. The role of activated charcoal in plant tissue culture. Biotechnol. Adv. 2008, 26, 618–631. [Google Scholar] [CrossRef]
- Kumar, A.; Chauhan, S.; Rattan, S.; Warghat, A.R.; Kumar, D.; Bhargava, B. In vitro propagation and phyto-chemical assessment of Cymbidium aloifolium (L.) Sw.: An orchid of pharma-horticultural importance. S. Afr. J. Bot. 2022, 144, 261–269. [Google Scholar] [CrossRef]
- Kim, D.H.; Kang, K.W.; Sivanesan, I. In vitro germination and seedling development of Gastrochilus japonicus (Makino) Schltr. Propag. Ornam. Plants 2019, 19, 61–65. [Google Scholar]
- Godo, T.; Hashimoto, T.; Nakata, M.; Miyoshi, K. The effects of illumination, temperature and 6-benzylaminoprine on asymbiotic seed germination and protocorm development in vitro in the achlorophyllous orchid Gastrodia pubilabiata Sawa. In Vitro Cell. Dev. Biol. Plant 2020, 56, 230–235. [Google Scholar] [CrossRef]
- Fritsche, Y.; Deola, F.; da Silva, D.A.; Holderbaum, D.F.; Guerra, M.P. Cattleya tigrina (Orchidaceae) in vitro regeneration: Main factors for optimal protocorm-like body induction and multiplication, plantlet regeneration, and cytogenetic stability. S. Afr. J. Bot. 2022, 149, 96–108. [Google Scholar] [CrossRef]
- Seon, K.M.; Kim, D.H.; Kang, K.W.; Sivanesan, I. Highly competent in vitro propagation of Thrixspermum japonicum (Miq.) Rchb.f., a rare epiphytic orchid. In Vitro Cell. Dev. Biol. Plant 2018, 54, 302–308. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
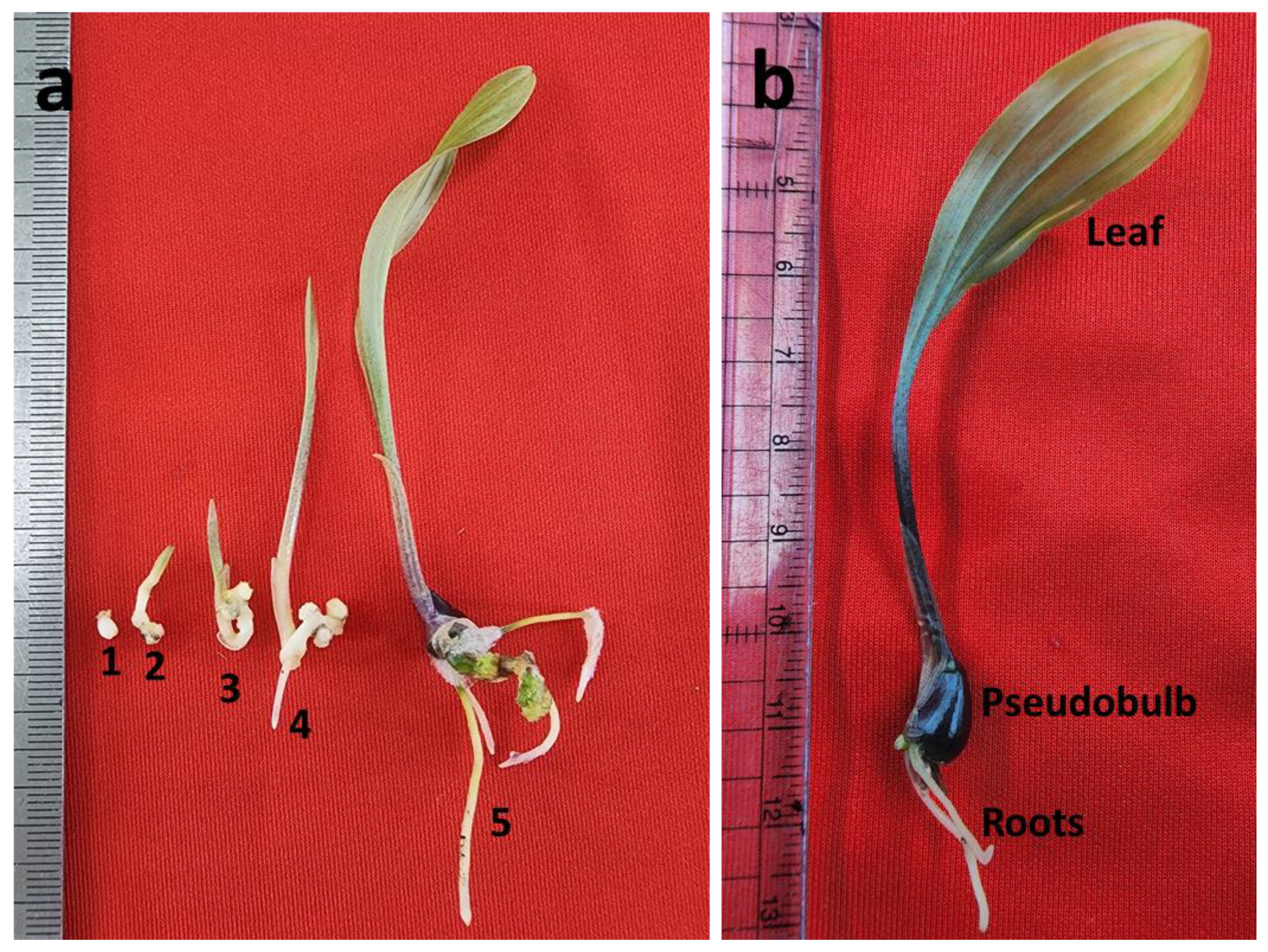{kind=link}
| Media | Seed Age (Days) | Seed Germination (%) |
|---|---|---|
| MS | 132 | 23.7 ± 6.9 c |
| 159 | 39.1 ± 6.5 a | |
| 210 | 14.1 ± 4.8 def | |
| Half-MS | 132 | 17.9 ± 5.5 d |
| 159 | 30.4 ± 5.9 b | |
| 210 | 10.6 ± 4.1 efg | |
| KC | 132 | 15.2 ± 5.2 de |
| 159 | 23.2 ± 5.9 c | |
| 210 | 9.9 ± 2.3 fg | |
| Hyponex | 132 | 6.2 ± 2.2 gh |
| 159 | 10.4 ± 3.5 efg | |
| 210 | 3.7 ± 1.6 h | |
| ANOVA | R-square | 0.825683 |
| Coefficient of variation | 28.53494 | |
| Root mean square error | 4.861508 | |
| F value | Pr > F | |
| Media | 71.08 | <0.0001 |
| Seed age | 102.45 | <0.0001 |
| Media*Seed age | 6.10 | <0.0001 |
| AC (mg/L) | % of Germination | |
|---|---|---|
| 0 | 39.1 ± 6.5 b | |
| 250 | 41.9 ± 8.1 b | |
| 500 | 48.7 ± 6.7 a | |
| 1000 | 35.7 ± 4.8 b | |
| 2000 | 27.3 ± 4.0 c | |
| ANOVA | R-square | 0.546221 |
| Coefficient of variation | 17.66698 | |
| Root mean square error | 6.807675 | |
| F value | 12.04 | |
| Pr > F | <0.0001 | |
| Auxin | Concentration (µM) | % of Germination |
|---|---|---|
| Control | 0.0 | 48.7 ± 6.7 fg |
| IAA | 1.5 | 52.1 ± 3.9 ef |
| 3.0 | 54.7 ± 4.1 de | |
| 5.0 | 58.3 ± 4.9 cd | |
| 10.0 | 43.3 ± 4.9 hi | |
| IBA | 1.5 | 56.2 ± 6.9 cde |
| 3.0 | 65.1 ± 4.0 b | |
| 5.0 | 46.8 ± 6.1 gh | |
| 10.0 | 33.9 ± 4.1 j | |
| NAA | 1.5 | 55.6 ± 5.7 cde |
| 3.0 | 73.4 ± 5.3 a | |
| 5.0 | 60.6 ± 4.6 bc | |
| 10.0 | 38.9 ± 5.9 i | |
| ANOVA | R-Square | 0.829979 |
| Coefficient of variation | 9.621950 | |
| Root mean square error | 5.122798 | |
| F Value | Pr > F | |
| Auxin | 16.30 | <0.0001 |
| Concentration | 117.25 | <0.0001 |
| Auxin*Concentration | 14.05 | <0.0001 |
| KIN (µM) | NAA (µM) | % of Germination |
|---|---|---|
| 0 | 3 | 73.4 ± 5.3 c |
| 1 | 3 | 78.6 ± 7.5 c |
| 2 | 3 | 91.9 ± 4.6 a |
| 4 | 3 | 85.2 ± 7.5 b |
| 8 | 3 | 63.8 ± 8.6 d |
| ANOVA | R-square | 0.688727 |
| Coefficient of variation | 8.767367 | |
| Root mean square error | 6.889203 | |
| F value | 22.13 | |
| Pr > F | <0.0001 |
| BA (µM) | KIN (µM) | Number of Secondary Protocorms | Secondary Protocorm Conversion (%) |
|---|---|---|---|
| 0 | 0 | 0.0 ± 0.0 f | 0.0 ± 0.0 h |
| 2 | 1 | 4.4 ± 1.5 e | 41.6 ± 3.0 a |
| 4 | 1 | 17.8 ± 3.4 b | 28.1 ± 1.8 c |
| 8 | 1 | 8.9 ± 2.1 d | 16.7 ± 2.2 f |
| 2 | 2 | 7.7 ± 1.6 d | 22.2 ± 3.3 d |
| 4 | 2 | 28.9 ± 2.9 a | 18.0 ± 2.1 ef |
| 8 | 2 | 12.1 ± 2.3 c | 12.1 ± 2.7 g |
| 2 | 4 | 4.8 ± 1.7 e | 20.1 ± 2.6 ed |
| 4 | 4 | 8.0 ± 1.7 d | 26.0 ± 2.2 c |
| 8 | 4 | 3.0 ± 1.1 e | 33.7 ± 2.9 b |
| ANOVA | R-square | 0.935623 | 0.926335 |
| Coefficient of variation | 20.22504 | 10.64282 | |
| Root mean square error | 2.147350 | 2.583154 | |
| F value | 130.80 | 113.17 |
| GA3 (µM) | Protocorm Conversion (%) | |
|---|---|---|
| 0 | 19.2 ± 4.9 e | |
| 0.5 | 54.2 ± 5.8 c | |
| 1.0 | 78.7 ± 4.2 a | |
| 2.0 | 62.3 ± 4.0 b | |
| 3.0 | 40.6 ± 4.8 d | |
| ANOVA | R-square | 0.952336 |
| Coefficient of variation | 9.359194 | |
| Root mean square error | 4.773189 | |
| F value | 199.80 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faisal, M.; Seob, P.K.; Kang, K.W.; Sivanesan, I. In Vitro Propagation of Cremastra appendiculata var. variabilis by Asymbiotic Seed Germination. Horticulturae 2022, 8, 926. https://doi.org/10.3390/horticulturae8100926
Faisal M, Seob PK, Kang KW, Sivanesan I. In Vitro Propagation of Cremastra appendiculata var. variabilis by Asymbiotic Seed Germination. Horticulturae. 2022; 8(10):926. https://doi.org/10.3390/horticulturae8100926
Chicago/Turabian StyleFaisal, Mohammad, Park Kwang Seob, Kyung Won Kang, and Iyyakkannu Sivanesan. 2022. "In Vitro Propagation of Cremastra appendiculata var. variabilis by Asymbiotic Seed Germination" Horticulturae 8, no. 10: 926. https://doi.org/10.3390/horticulturae8100926
APA StyleFaisal, M., Seob, P. K., Kang, K. W., & Sivanesan, I. (2022). In Vitro Propagation of Cremastra appendiculata var. variabilis by Asymbiotic Seed Germination. Horticulturae, 8(10), 926. https://doi.org/10.3390/horticulturae8100926