Effects of Phospholipase D Inhibitors Treatment on Membrane Lipid Metabolism of Postharvest Banana Fruit in Response to Mechanical Wounding Stress

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Sample Treatment
2.2. Firmness, Total Soluble Solids and Respiration Rate
2.3. Measurement of Cell Membrane Permeability and MDA Content
2.4. Assays of PLD and LOX Activities
2.5. Measurement of the Contents of Cellular Membrane Phospholipids
2.6. Statistical Analysis
3. Results
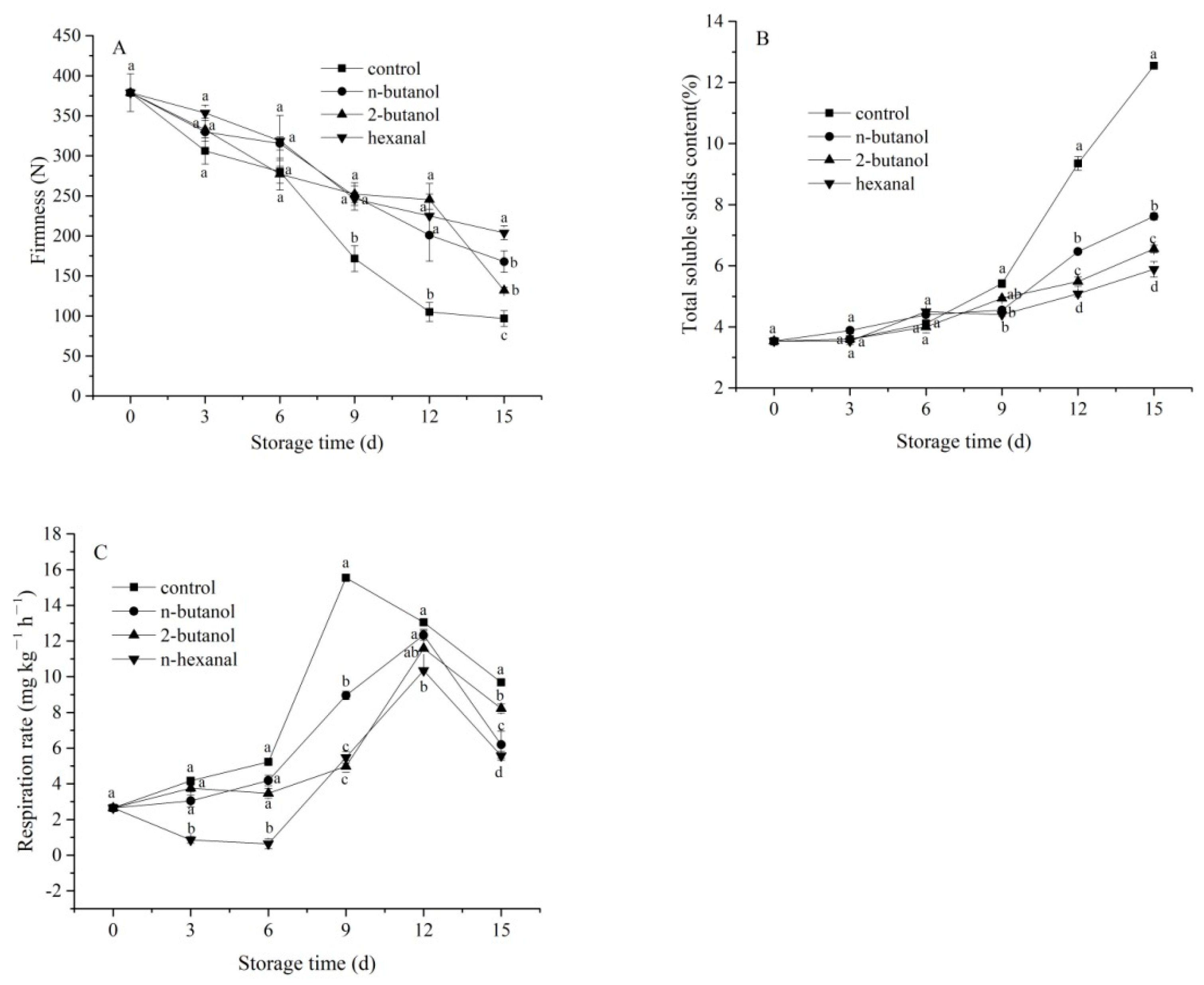
3.1. Firmness, Total Soluble Solids Content and Respiration Rate
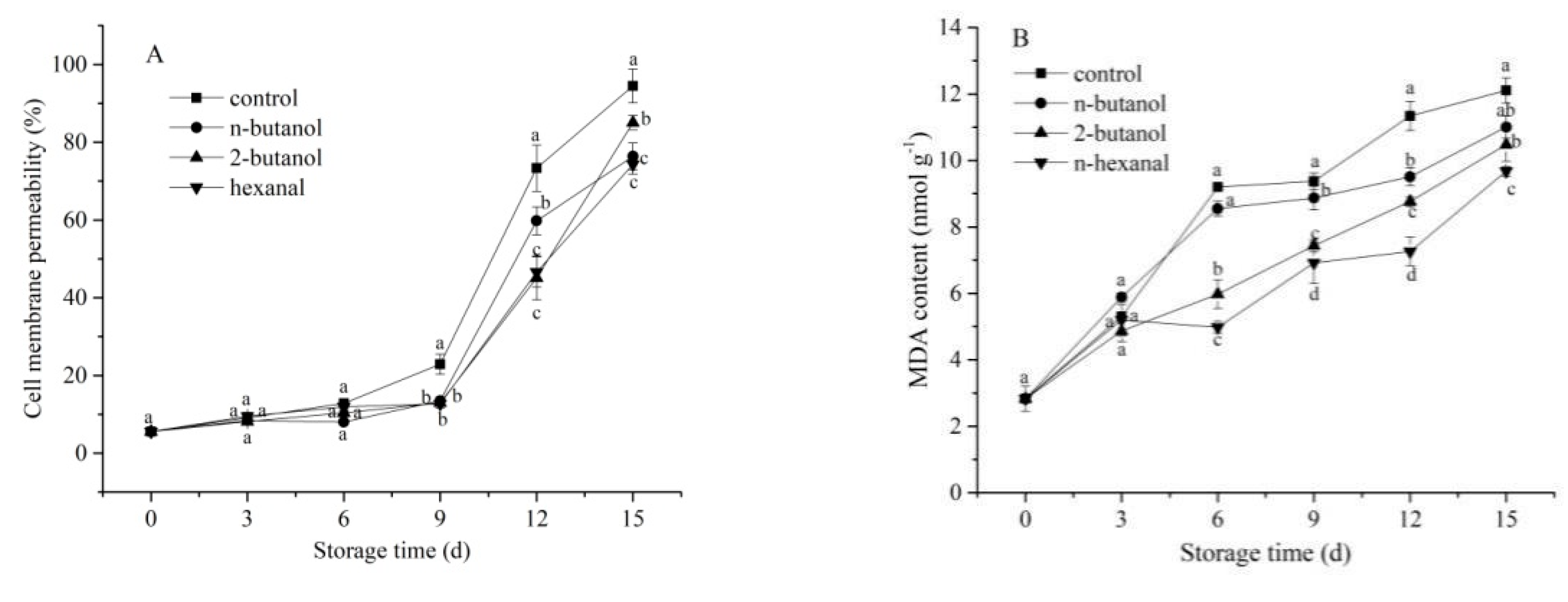
3.2. Cell membrane Permeability and MDA Content
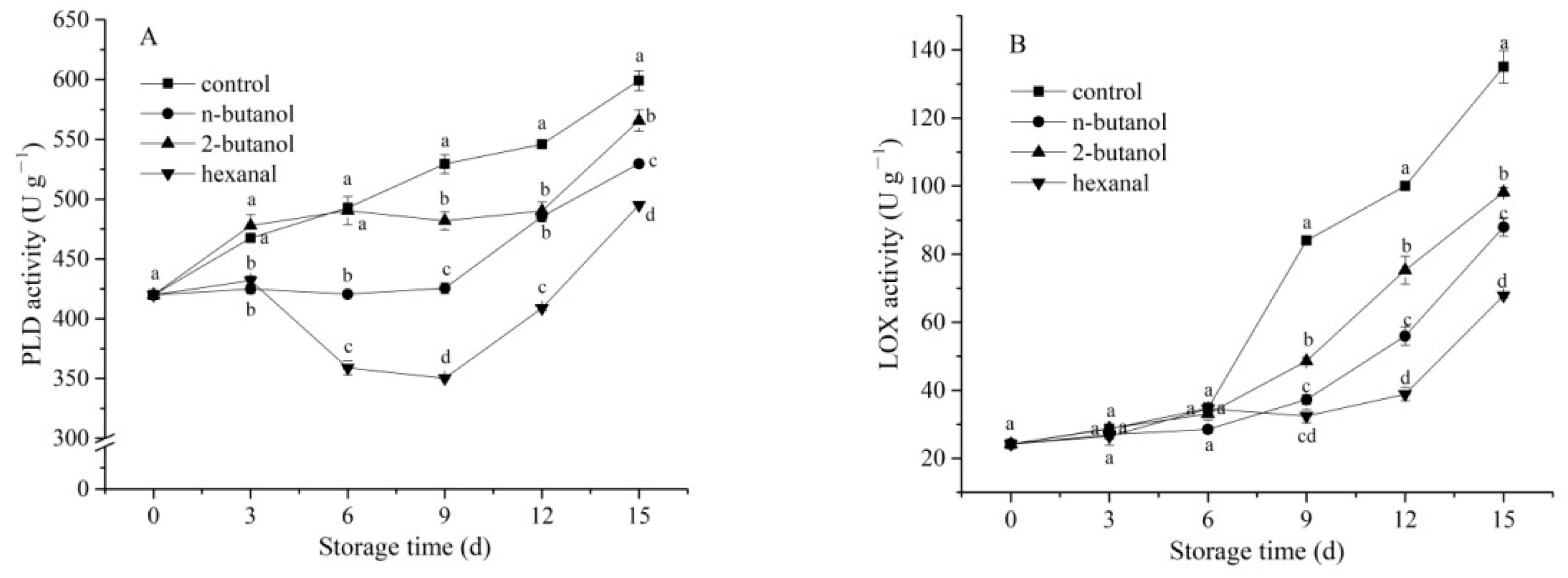
3.3. Activities of PLD and LOX
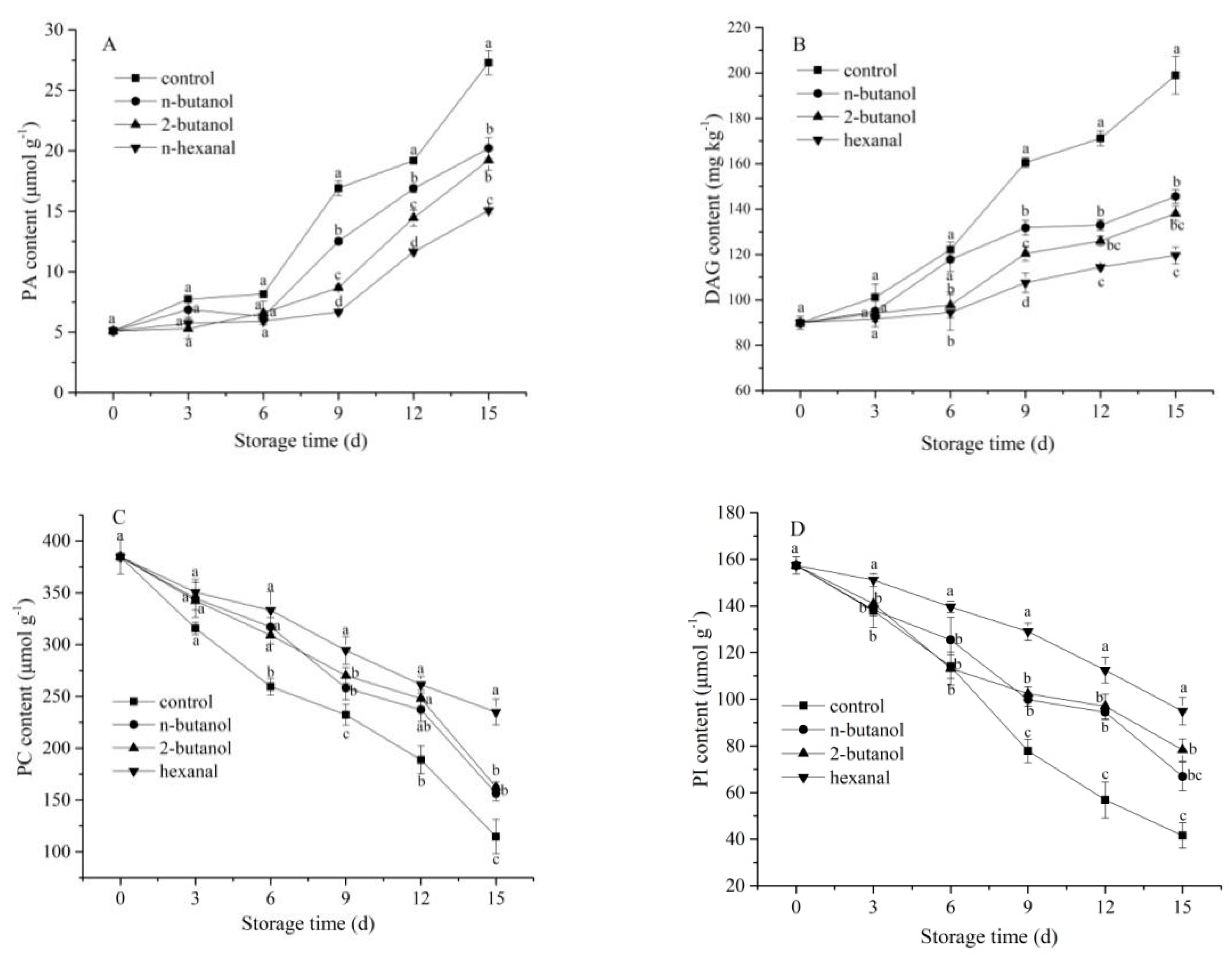
3.4. Changes in Membrane Phospholipids Component
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Z.; Pu, H.; Shan, S.; Zhang, P.; Li, J.; Song, H.; Xu, X. Melatonin enhanced chilling tolerance and alleviated peel browning of banana fruit under low temperature storage. Postharvest Biol. Technol. 2021, 179, 111571. [Google Scholar] [CrossRef]
- Pongprasert, N.; Srilaong, V. A novel technique using 1-MCP microbubbles for delaying postharvest ripening of banana fruit. Postharvest Biol. Technol. 2014, 95, 42–45. [Google Scholar] [CrossRef]
- Liu, J.; Li, F.; Shan, Y.; Zhou, Y.; Liang, L.; Qu, H.; Jiang, Y.; Chen, J. Determination of H+ and Ca2+ fluxes in cold-stored banana fruit using non-invasive micro-test technology. Postharvest Biol. Technol. 2019, 153, 169–175. [Google Scholar] [CrossRef]
- Peng, Y.; Mao, L.C. Salicylic acid, ethephon, and methyl jasmonate induce the expression of phospholipase D in mechanically-wounded cucumber. J. Hortic. Sci. Biotechnol. 2011, 86, 235–240. [Google Scholar] [CrossRef]
- Zien, C.A.; Wang, C.; Wang, X.; Welti, R. In vivo substrates and the contribution of the common phospholipase D, PLDα, to wound-induced metabolism of lipids in Arabidopsis. Biochim. Biophys. Acta BBA—Mol. Cell Biol. Lipids 2001, 1530, 236–248. [Google Scholar] [CrossRef]
- Bourtsala, A.; Farmaki, T.; Galanopoulou, D. Phospholipases Dα and δ are involved in local and systemic wound responses of cotton (G. hirsutum). Biochem. Biophys. Rep. 2017, 9, 133–139. [Google Scholar] [CrossRef]
- Wang, C.; Zien, C.A.; Afitlhile, M.; Welti, R.; Hildebrand, D.F.; Wang, X. Involvement of Phospholipase D in Wound-Induced Accumulation of Jasmonic Acid in Arabidopsis. Plant Cell 2000, 12, 2237–2246. [Google Scholar] [CrossRef]
- Wang, X. Regulatory functions of phospholipase D and phosphatidic acid in plant growth, development, and stress responses. Plant Physiol. 2005, 139, 566–573. [Google Scholar] [CrossRef]
- Whitaker, B.D. Membrane lipid metabolism and oxidative stress involved in postharvest ripening, senescence, and storage disorders of fruits. Acta Hortic. 2012, 945, 269–282. [Google Scholar] [CrossRef]
- Thammawong, M.; Umehara, H.; Kaneta, T.; Nakamura, N.; Ito, Y.; Shiina, T.; Yoshida, M.; Soga, A.; Nakano, K. Changes in Gene Expression of Harvested Cabbage in Response to Mechanical Wound Stress. J. Acta Hortic. 2013, 1005, 117–123. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Qian, C.L.; Chen, J.C.; Peng, Y.; Mao, L.C. Responses of Phospholipase D and Lipoxygenase to Mechanical Wounding in Postharvest Cucumber Fruits. J. Zhejiang Univ. Sci. 2010, 11, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Paliyath, G.; Pinhero, R.G.; Yada, R.Y.; Murr, D.P. Effect of processing conditions on phospholipase D activity of corn kernel subcellular fractions. Agric. Food Chem. 1999, 47, 2579–2588. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, K.; Paliyath, G. Microarray analysis of ripening-regulated gene expression and its modulation by 1-MCP and hexanal. Plant Physiol. Biochem. 2011, 49, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Paliyath, G.; Padmanabhan, P. Preharvest and Postharvest Technologies Based on Hexanal. In Postharvest Biology and Nanotechnology; Paliyath, G., Subramanian, J., Lim, L.-T., Subramanian, K.S., Handa, A.K., Mattoo, A.K., Eds.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2019; pp. 89–101. [Google Scholar]
- Pak Dek, M.S.; Padmanabhan, P.; Subramanian, J.; Paliyath, G. Inhibition of tomato fruit ripening by 1-MCP, wortmannin and hexanal is associated with a decrease in transcript levels of phospholipase D and other ripening related genes. Postharvest Biol. Technol. 2018, 140, 50–59. [Google Scholar] [CrossRef]
- Sun, Y.; Sun, H.; Luo, M.; Zhou, X.; Zhou, Q.; Wei, B.; Cheng, S.; Ji, S. Membrane lipid metabolism in relation to core browning during ambient storage of ‘Nanguo’ pears. Postharvest Biol. Technol. 2020, 169, 111288. [Google Scholar] [CrossRef]
- Sun, J.; You, X.R.; Li, L.; Peng, H.X.; Su, W.Q.; Li, C.B.; He, Q.G.; Liao, F. Effects of a phospholipase D inhibitor on postharvest enzymatic browning and oxidative stress of litchi fruit. Postharvest Biol. Technol. 2011, 62, 288–294. [Google Scholar] [CrossRef]
- Li, L.; Li, J.; Sun, J.; Li, C.; Sheng, J.; Zheng, F.; Liao, F.; He, X.; Liu, G.; Ling, D.; et al. Effects of 2-butanol on quality and physiological characteristics of longan fruit stored at ambient temperature. Postharvest Biol. Technol. 2015, 101, 96–102. [Google Scholar] [CrossRef]
- Sharma, M.; Jacob, J.K.; Subramanian, J.; Paliyath, G. Hexanal and 1-MCP treatments for enhancing the shelf life and quality of sweet cherry (Prunus avium L.). Sci. Hortic. 2010, 125, 239–247. [Google Scholar] [CrossRef]
- Cheema, A.; Padmanabhan, P.; Subramanian, J.; Blom, T.; Paliyath, G. Improving quality of greenhouse tomato (Solanum lycopersicum L.) by pre- and postharvest applications of hexanal-containing formulations. Postharvest Biol. Technol. 2014, 95, 13–19. [Google Scholar] [CrossRef]
- Gill, K.S.; Dhaliwal, H.S.; Mahajan, B.V.C.; Paliyath, G.; Boora, R.S. Enhancing postharvest shelf life and quality of guava (Psidium guajava L.) cv. Allahabad Safeda by pre-harvest application of hexanal containing aqueous formulation. Postharvest Biol. Technol. 2016, 112, 224–232. [Google Scholar] [CrossRef]
- Cheema, A.; Padmanabhan, P.; Amer, A.; Parry, M.J.; Lim, L.-T.; Subramanian, J.; Paliyath, G. Postharvest hexanal vapor treatment delays ripening and enhances shelf life of greenhouse grown sweet bell pepper (Capsicum annum L.). Postharvest Biol. Technol. 2018, 136, 80–89. [Google Scholar] [CrossRef]
- Padmanabhan, P.; Cheema, A.S.; Todd, J.F.; Lim, L.-T.; Paliyath, G. Ripening responses, fruit quality and phospholipase D gene expression in bell peppers exposed to hexanal vapor. Postharvest Biol. Technol. 2020, 170, 111317. [Google Scholar] [CrossRef]
- Li, L.; He, X.; Sun, J.; Li, C.; Ling, D.; Sheng, J.; Zheng, F.; Liu, G.; Li, J.; Tang, Y.; et al. Responses of Phospholipase D and Antioxidant System to Mechanical Wounding in Postharvest Banana Fruits. J. Food Qual. 2017, 2017, 8347306. [Google Scholar] [CrossRef]
- Yi, P.; Li, L.; Sun, J.; He, X.; Li, C.; Sheng, J.; Xin, M.; Ling, D.; Li, Z.; Tang, Y.; et al. Characterization and Expression of Phospholipase D Putatively Involved in Colletotrichum musae Disease Development of Postharvest Banana Fruit. Horticulturae 2022, 8, 312. [Google Scholar] [CrossRef]
- Li, L.; Li, C.; Sun, J.; Xin, M.; Yi, P.; He, X.; Sheng, J.; Zhou, Z.; Ling, D.; Zheng, F.; et al. Synergistic effects of ultraviolet light irradiation and high-oxygen modified atmosphere packaging on physiological quality, microbial growth and lignification metabolism of fresh-cut carrots. Postharvest Biol. Technol. 2021, 173, 111365. [Google Scholar] [CrossRef]
- Song, L.L.; Liu, H.; You, Y.L.; Sun, J.; Yi, C.; Li, Y.B.; Jiang, Y.M.; Wu, J.S. Quality deterioration of cut carnation flowers involves in antioxidant systems and energy status. Sci. Hortic. 2014, 170, 45–52. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, J.; Lin, H.; Lin, M.; Lin, Y.; Wang, H.; Hung, Y.C. Salicylic acid treatment suppresses Phomopsis longanae Chi-induced disease development of postharvest longan fruit by modulating membrane lipid metabolism. Postharvest Biol. Technol. 2020, 164, 111168. [Google Scholar] [CrossRef]
- Lin, Y.; Chen, M.; Lin, H.; Lin, M.; Hung, Y.; Lin, Y.; Chen, Y.; Wang, H.; Ritenourd, M.A. Phomopsis longanae-induced pericarp browning and disease development of longan fruit can be alleviated or aggravated by regulation of ATP-mediated membrane lipid metabolism. Food Chem. 2018, 269, 644–651. [Google Scholar] [CrossRef]
- Kong, X.; Ge, W.; Wei, B.; Zhou, Q.; Zhou, X.; Zhao, Y.; Ji, S. Melatonin ameliorates chilling injury in green bell peppers during storage by regulating membrane lipid metabolism and antioxidant capacity. Postharvest Biol. Technol. 2020, 170, 111315. [Google Scholar] [CrossRef]
- Hou, Q.; Ufer, G.; Bartels, D. Lipid signalling in plant responses to abiotic stress. Plant Cell Environ. 2016, 39, 1029–1048. [Google Scholar] [CrossRef]
- Motes, C.M.; Pechter, P.; Yoo, C.M.; Wang, Y.S.; Chapman, K.D.; Blancaflor, E.B. Differential effects of two phospholipase D inhibitors 1-butanol and Nacylethanolamine, on in vivo cytoskeletal organization and Arabidopsis seedling growth. Protoplasma 2005, 226, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Peters, N.T.; Logan, K.O.; Miller, A.C.; Kropf, D.L. Phospholipase D signaling regulates microtubule organization in the fucoid alga Silvetia compressa. Plant Cell Physiol. 2007, 48, 1764–1774. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sheng, L.; Zhou, X.; Liu, Z.Y.; Wang, J.W.; Zhou, Q.; Wang, L.; Zhang, Q.; Ji, S.J. Changed activities of enzymes crucial to membrane lipid metabolism accompany pericarp browning in ‘Nanguo’ pears during refrigeration and subsequent shelf life at room temperature. Postharvest Biol. Technol. 2016, 117, 1–8. [Google Scholar] [CrossRef]
- Zhang, Q.; Qi, Y.; Li, R.; Yang, Y.; Yan, D.; Liu, X.; Ren, X. Postharvest applications of n-butanol increase greasiness in apple skins by altering wax composition via effects on gene expression. Postharvest Biol. Technol. 2019, 155, 111–119. [Google Scholar] [CrossRef]
- Jincy, M.; Djanaguiraman, M.; Jeyakumar, P.; Subramanian, K.S.; Jayasankar, S.; Paliyath, G. Inhibition of phospholipase D enzyme activity through hexanal leads to delayed mango (Mangifera indica L.) fruit ripening through changes in oxidants and antioxidant enzymes activity. Sci. Hortic. 2017, 218, 316–325. [Google Scholar] [CrossRef]
- Misran, A.; Padmanabhan, P.; Sullivan, J.A.; Khanizadeh, S.; Paliyath, G. Composition of phenolics and volatiles in strawberry cultivars and influence of preharvest hexanal treatment on their profiles. Can. J. Plant Sci. 2015, 95, 115–126. [Google Scholar] [CrossRef]
- Jiang, Y.M.; Jiang, Y.L.; Qu, H.X.; Duan, X.W.; Luo, Y.B.; Jiang, W.B. Energy aspects in ripening and senescence of harvested horticultural crops. Stewart Postharvest Rev. 2007, 3, 1–5. [Google Scholar]
- Lin, Y.F.; Lin, H.T.; Lin, Y.X.; Zhang, S.; Chen, Y.H.; Jiang, X.J. The roles of metabolism of membrane lipids and phenolics in hydrogen peroxide-induced pericarp browning of harvested longan fruit. Postharvest Biol. Technol. 2016, 111, 53–61. [Google Scholar] [CrossRef]
- Mao, L.; Pang, H.; Wang, G.; Zhu, C. Phospholipase D and lipoxygenase activity of cucumber fruit in response to chilling stress. Postharvest Biol. Technol. 2007, 44, 42–47. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Yi, P.; Huang, F.; Tang, J.; Sun, J.; Duan, X.; Li, J.; Su, Z.; Ling, D.; Tang, Y.; et al. Effects of Phospholipase D Inhibitors Treatment on Membrane Lipid Metabolism of Postharvest Banana Fruit in Response to Mechanical Wounding Stress. Horticulturae 2022, 8, 901. https://doi.org/10.3390/horticulturae8100901
Li L, Yi P, Huang F, Tang J, Sun J, Duan X, Li J, Su Z, Ling D, Tang Y, et al. Effects of Phospholipase D Inhibitors Treatment on Membrane Lipid Metabolism of Postharvest Banana Fruit in Response to Mechanical Wounding Stress. Horticulturae. 2022; 8(10):901. https://doi.org/10.3390/horticulturae8100901
Chicago/Turabian StyleLi, Li, Ping Yi, Fang Huang, Jie Tang, Jian Sun, Xuewu Duan, Jiemin Li, Zuxiang Su, Dongning Ling, Yayuan Tang, and et al. 2022. "Effects of Phospholipase D Inhibitors Treatment on Membrane Lipid Metabolism of Postharvest Banana Fruit in Response to Mechanical Wounding Stress" Horticulturae 8, no. 10: 901. https://doi.org/10.3390/horticulturae8100901
APA StyleLi, L., Yi, P., Huang, F., Tang, J., Sun, J., Duan, X., Li, J., Su, Z., Ling, D., Tang, Y., Li, C., He, X., Sheng, J., Li, Z., Huang, M., Xin, M., & Gan, T. (2022). Effects of Phospholipase D Inhibitors Treatment on Membrane Lipid Metabolism of Postharvest Banana Fruit in Response to Mechanical Wounding Stress. Horticulturae, 8(10), 901. https://doi.org/10.3390/horticulturae8100901