
Characterization of Soluble Sugar Content, Related Enzyme Activity and Gene Expression in the Fruits of ‘Minihyang’ Mandarin on Different Rootstocks

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Measurement of Scion Tree Growth, Yield, and Fruit Quality Characteristics
2.3. Fruit Sugar Analysis
2.4. Enzyme Extraction and Assay
2.5. Quantitative Real-Time (qRT)-PCR Reaction Analysis
2.6. Statistical Analysis
3. Results and Discussion
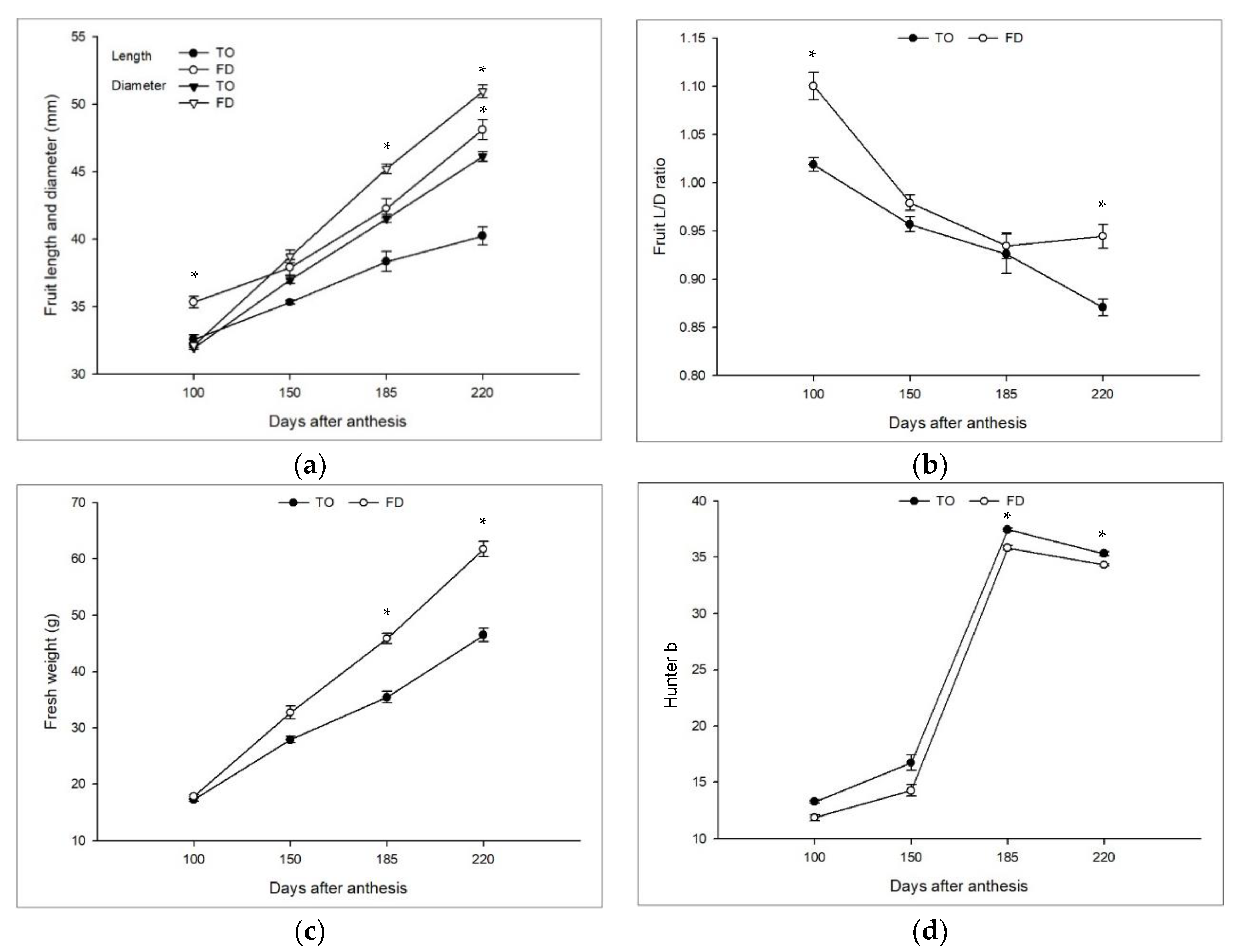
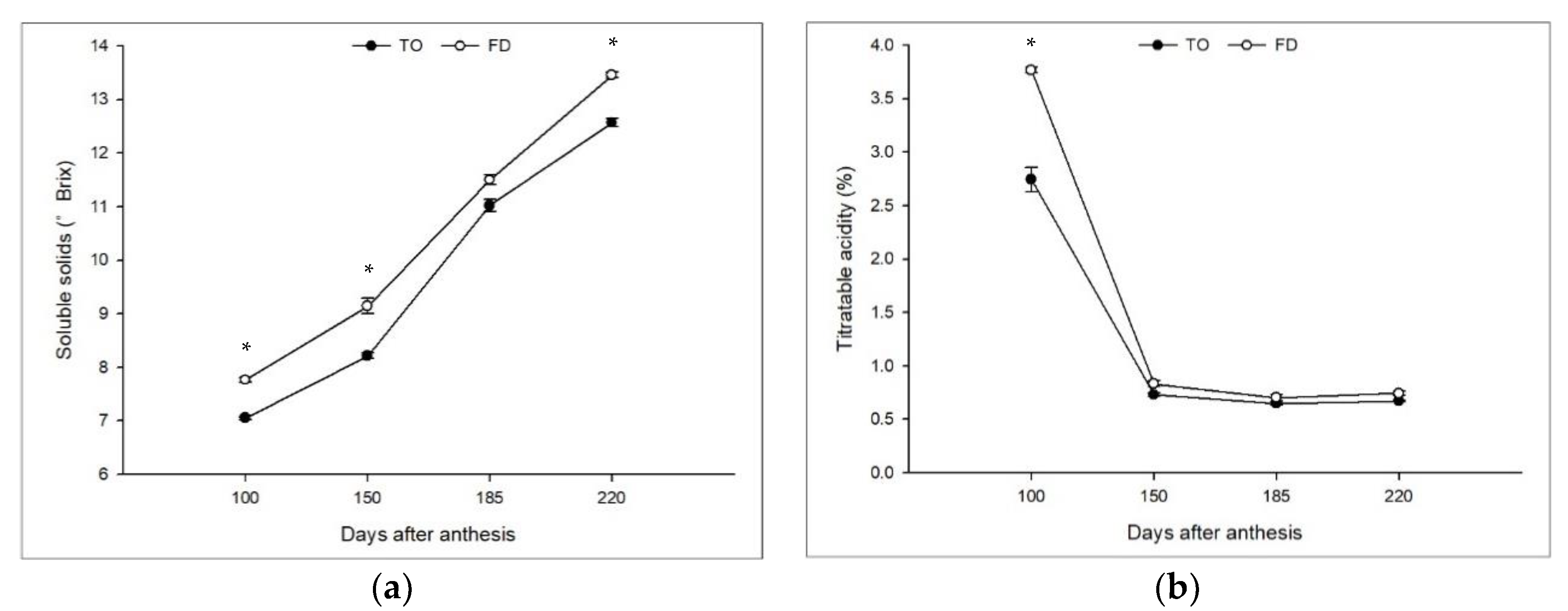
3.1. Effect of Different Rootstocks on Scion Tree Vegetative Growth, Yield, and Fruit Quality
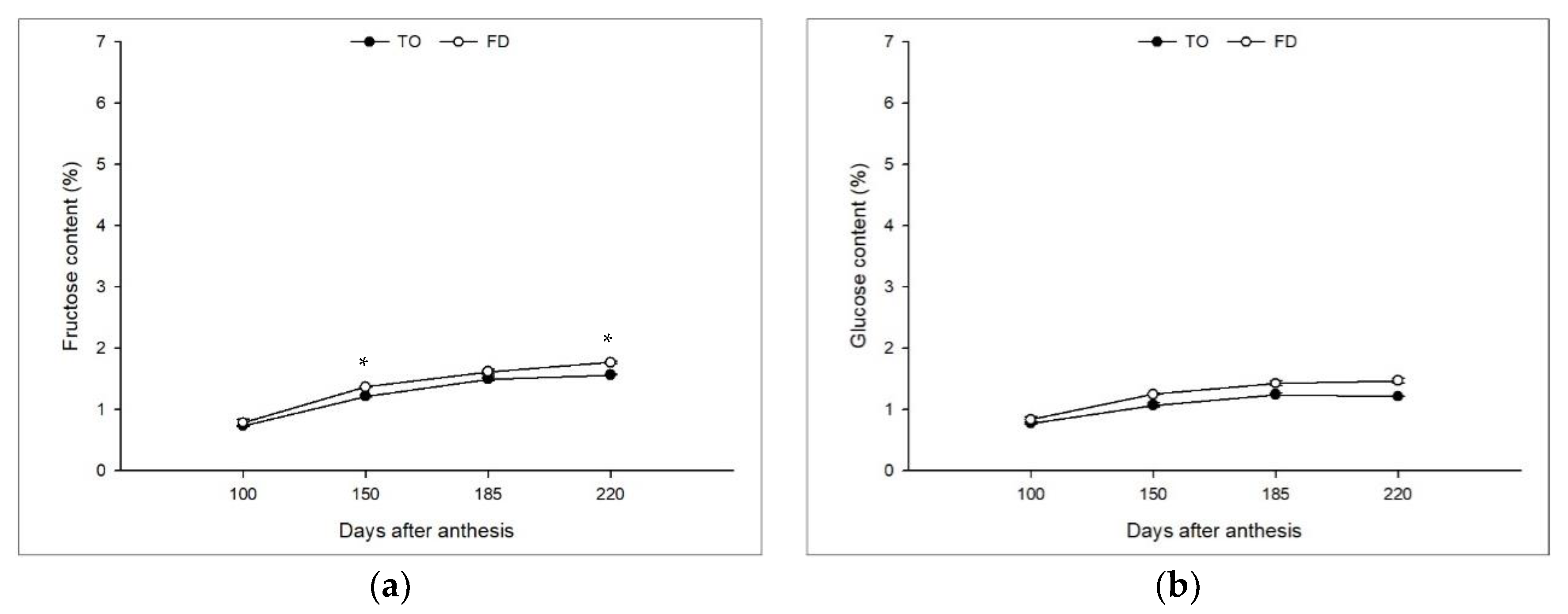
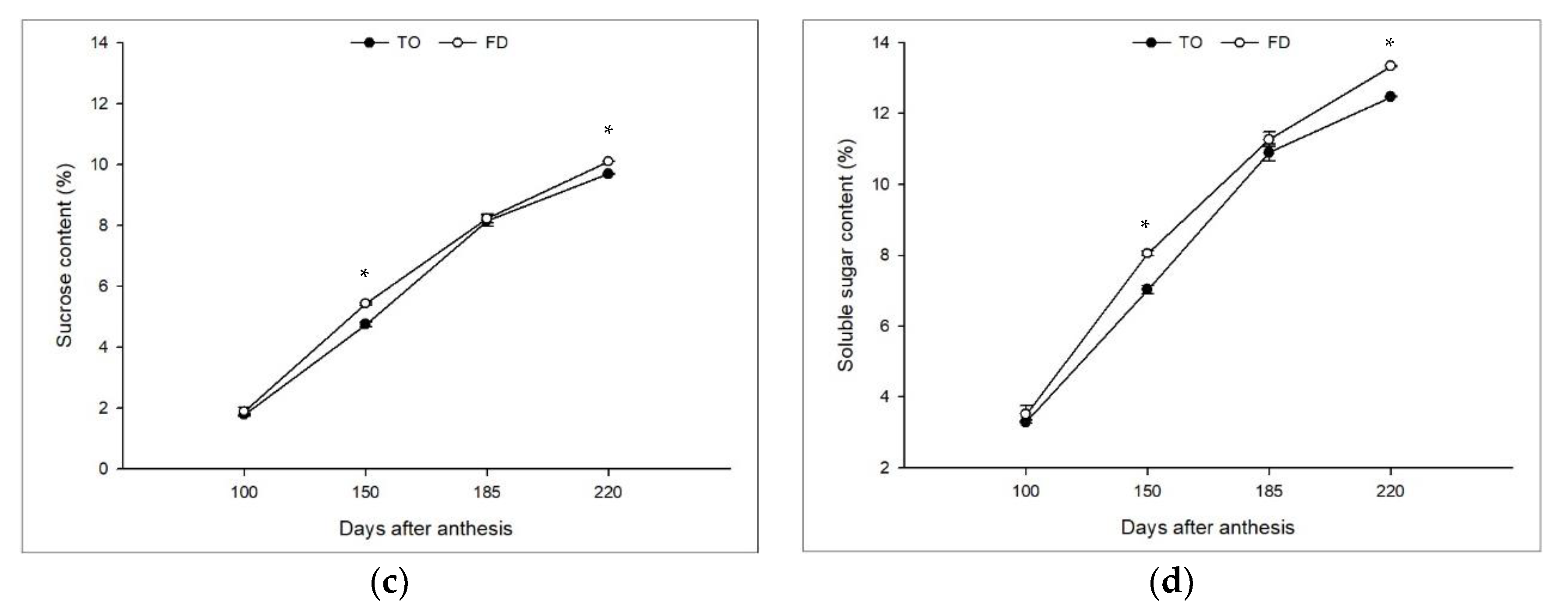
3.2. Effect of Different Rootstocks on Sugar Contents
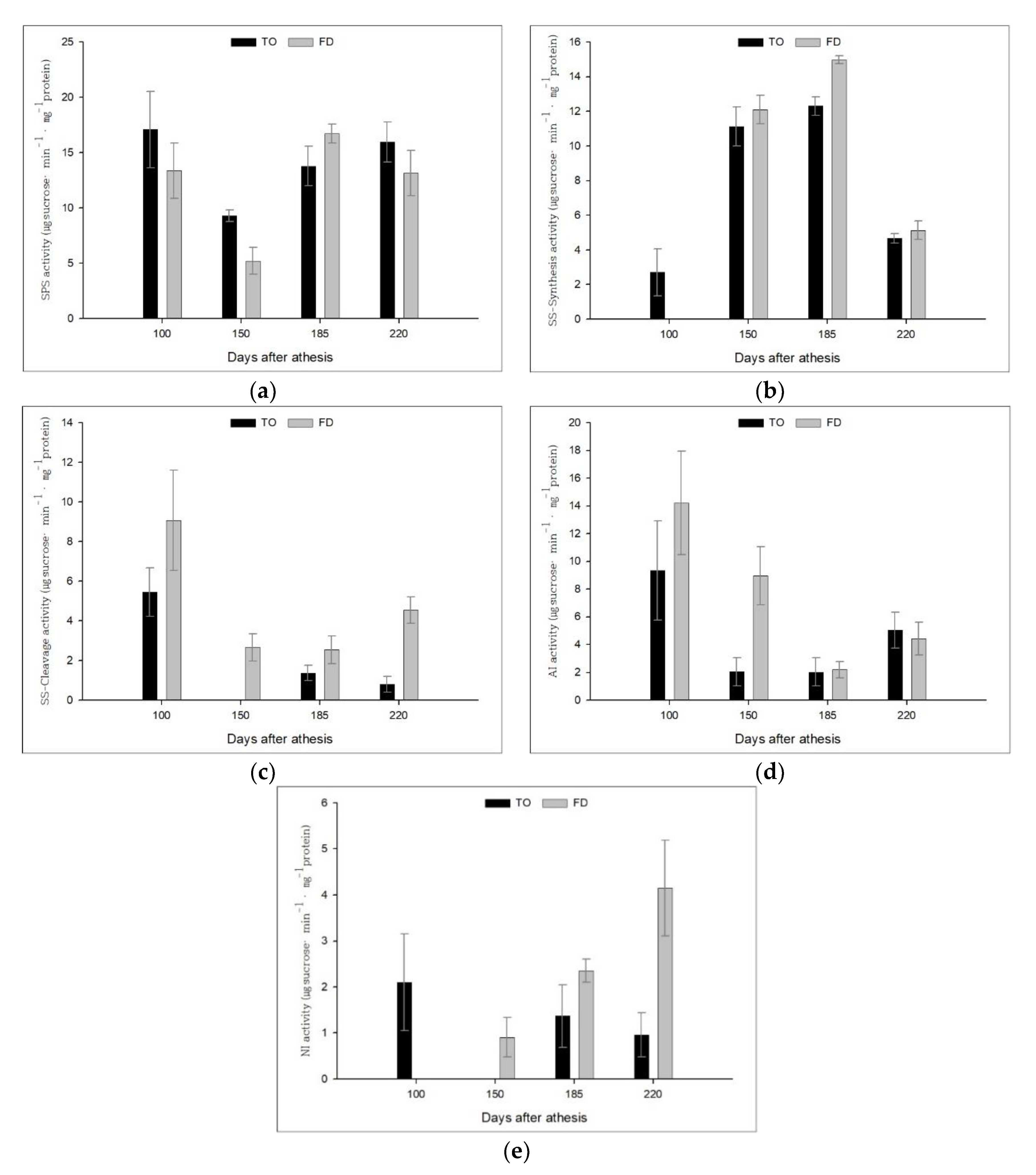
3.3. Effect of Different Rootstocks on Sucrose Metabolic Enzyme Activities
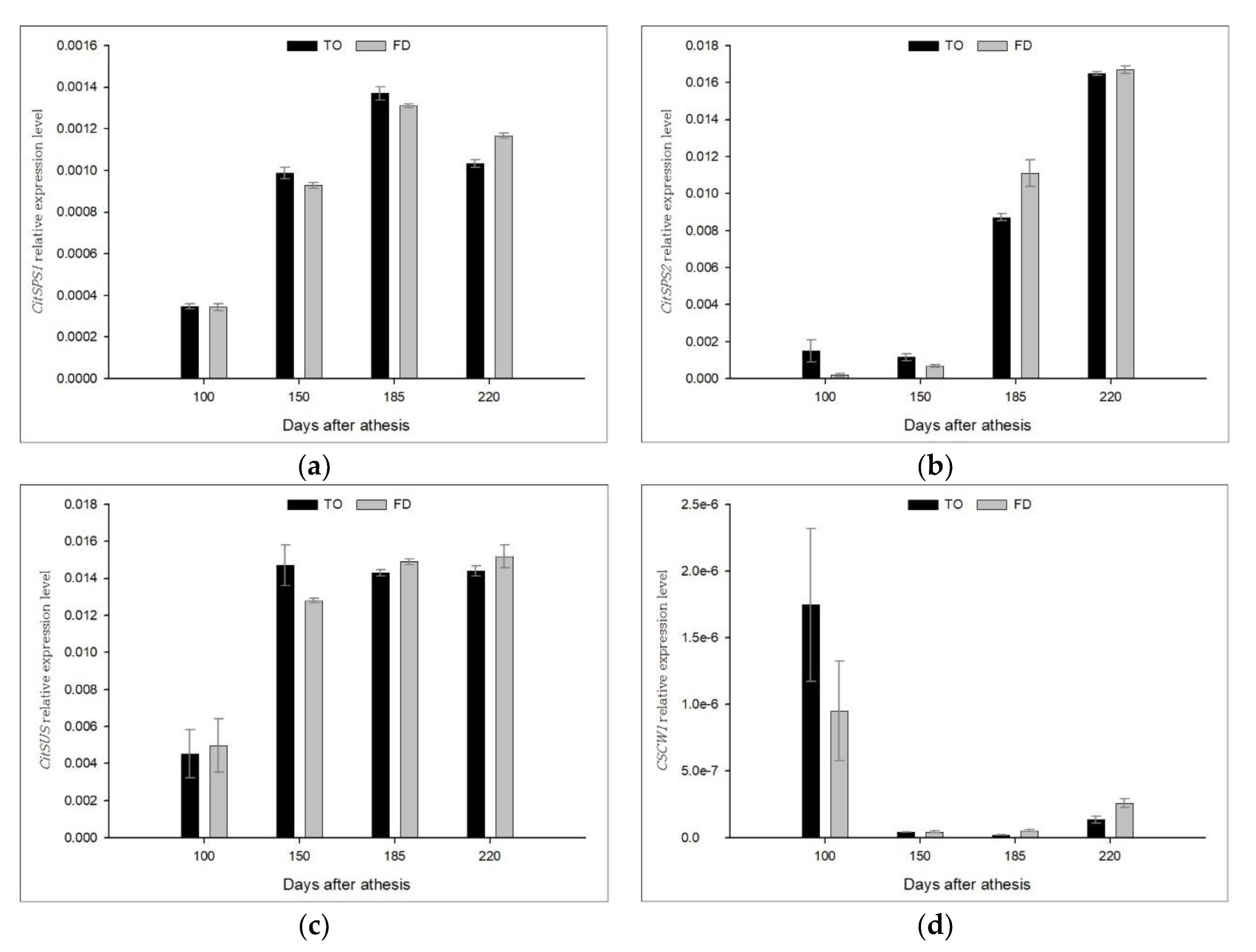
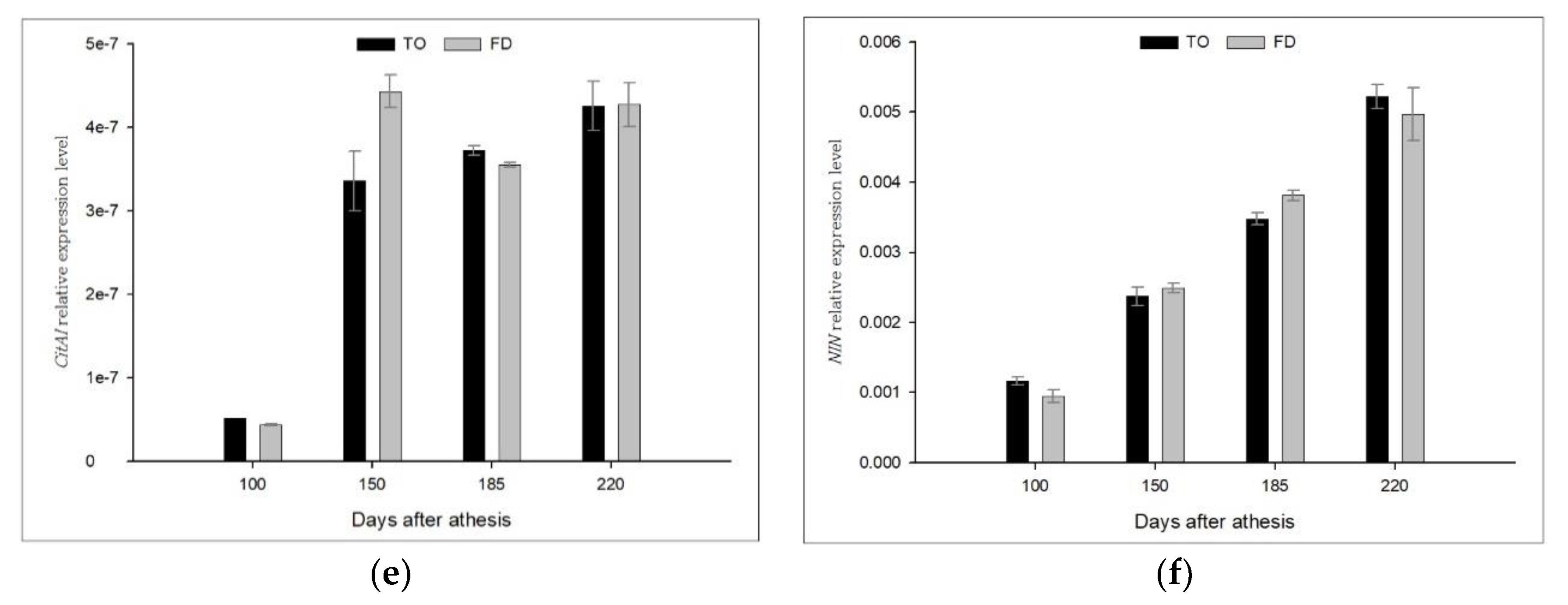
3.4. Effect of Different Rootstocks on the Expression of Sucrose Metabolic Enzyme Genes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sadka, A.; Shlizerman, L.; Kamara, I.; Blumwald, E. Primary metabolism in citrus fruit as affected by its unique structure. Front. Plant Sci. 2019, 10, 1167. [Google Scholar] [CrossRef] [PubMed]
- KOSIS. Download Data from the KOSIS Database of the Korean Statistical Information Service. Available online: https://kosis.kr/ (accessed on 17 November 2021).
- Castle, W.S. Rootstock as a fruit quality factor in citrus and deciduous tree crops. N. Z. J. Crop Hortic. 1995, 23, 383–394. [Google Scholar] [CrossRef]
- Zhou, G.F.; Liu, Y.Z.; Sheng, O.; Wei, Q.J.; Yang, C.Q.; Peng, S.A. Transcription profiles of boron-deficiency-responsive genes in citrus rootstock root by suppression subtractive hybridization and cDNA microarray. Front. Plant Sci. 2015, 5, 795. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Riaz, M.; Yan, L.; Du, C.; Liu, Y.; Jiang, C. Boron deficiency in trifoliate orange induces changes in pectin composition and architecture of components in root cell walls. Front. Plant Sci. 2017, 8, 1882. [Google Scholar] [CrossRef]
- Mademba-Sy, F.; Lemerre-Desprez, Z.; Lebegin, S. Use of Flying Dragon trifoliate orange as dwarfing rootstock for citrus under tropical climatic conditions. Hortic. Sci. 2012, 47, 11–17. [Google Scholar] [CrossRef]
- Dong, T.; Xiong, B.; Huang, S.; Liao, L.; Qiu, X.; Sun, G.; He, Y.; Duan, C.; Wang, X.; Zhang, X.; et al. Investigation of the cause of reduced sugar content in Kiyomi tangor fruit of Ziyang xiangcheng (Citrus junos Sieb. ex Tanaka) rootstock. Sci. Rep. 2019, 9, 19263. [Google Scholar] [CrossRef]
- Noda, K.; Okuda, H.; Kihara, T.; Iwagaki, I.; Kawase, K. Effects of rootstocks on tree growth and fruit quality in very early ripening satsuma mandarin ‘Yamakawa’. J. Jpn. Soc. Hortic. Sci. 2001, 70, 78–82. [Google Scholar] [CrossRef][Green Version]
- Lin, Q.; Wang, C.; Dong, W.; Jiang, Q.; Wang, D.; Li, S.; Chen, M.; Liu, C.; Sun, C.; Chen, K. Transcriptome and metabolome analyses of sugar and organic acid metabolism in Ponkan (Citrus reticulata) fruit during fruit maturation. Gene 2012, 554, 64–74. [Google Scholar] [CrossRef]
- Cardini, C.E.; Leloir, L.F.; Chiriboga, J. The biosynthesis of sucrose. J. Biol. Chem. 1995, 214, 149–155. [Google Scholar] [CrossRef]
- Stein, O.; Granot, D. An overview of sucrose synthases in plants. Front. Plant Sci. 2019, 10, 95. [Google Scholar] [CrossRef]
- Zhang, X.M.; Wang, W.; Du, L.Q.; Xie, J.H.; Yao, Y.L.; Sun, G.M. Expression patterns, activities and carbohydrate-metabolizing regulation of sucrose phosphate synthase, sucrose synthase and neutral invertase in pineapple fruit during development and ripening. Int. J. Mol. Sci. 2012, 13, 9460–9477. [Google Scholar] [CrossRef]
- Kubo, T.; Hohjo, I.; Hiratsuka, S. Sucrose accumulation and its related enzyme activities in the juice sacs of satsuma mandarin fruit from trees with different crop loads. Sci. Hortic. 2001, 91, 215–225. [Google Scholar] [CrossRef]
- Song, K.J.; Echeverria, E.; Lee, H.S. Distribution of Sugars and Related Enzymes in the Stem and Blossom Halves of ‘Valencia’ Oranges. J. Am. Soc. Hortic. Sci. 1998, 123, 416–420. [Google Scholar] [CrossRef]
- Van Handel, E. Direct microdetermination of sucrose. Anal. Biochem. 1968, 22, 280–283. [Google Scholar] [CrossRef]
- Brandford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cantuarias-Avilés, T.; Mourão Filho, F.D.A.A.; Stuchi, E.S.; da Silva, S.R.; Espinoza-Núñez, E.; Neto, H.B. Rootstocks for high fruit yield and quality of ‘Tahiti’ lime under rain-fed conditions. Sci. Hortic. 2012, 142, 105–111. [Google Scholar] [CrossRef]
- Andersen, P.C.; Brodbeck, B.V. Yield, tree size, and fruit quality of mature “owari” and “brown select” satsuma on Poncirus trifoliata “Rubidoux” and ‘Flying Dragon’ Rootstocks in North Florida. HortScience 2015, 50, 1650–1653. [Google Scholar] [CrossRef]
- Domingues, A.R.; Marcolini, C.D.M.; Goncalves, C.H.S.; Resende, J.T.V.; Roberto, S.R.; Carlos, E.F. Rootstocks genotypes impact on tree development and industrial properties of ‘Valencia’ sweet orange juice. Horticulturae 2021, 7, 141. [Google Scholar] [CrossRef]
- Marsh, K.B.; Richardson, A.C.; Macrae, E.A. Early-and mid-season temperature effects on the growth and composition of satsuma mandarins. J. Hortic. Sci. Biotechnol. 1999, 74, 443–451. [Google Scholar] [CrossRef]
- Zhang, J.; Ritenour, M.A. Sugar composition analysis of commercial citrus juice products. Proc. Fla. State Hortic. Soc. 2016, 129, 178–180. [Google Scholar]
- Moon, Y.E.; Kang, S.B.; Han, S.G.; Kim, Y.H.; Choi, Y.H.; Koh, S.C.; Oh, S.J. Effects of elevated spring temperatures on the growth and fruit quality of the Mandarin hybrid ‘Shiranuhi’. Kor. J. Hortic. Sci. Technol. 2015, 33, 459–469. [Google Scholar]
- Bowman, K.D.; McCollum, G. Five new citrus rootstocks with improved tolerance to huanglongbing. Hortic. Sci. 2015, 50, 1731–1734. [Google Scholar]
- Liu, Y.Z.; Liu, Q.; Xiong, J.J.; Deng, X.X. Difference of a citrus late-maturing mutant (Citrus sinensis) from its parental line in sugar and acid metabolism at the fruit ripening stage. Sci. China Ser. C. 2007, 50, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.B.; Oh, E.U.; Park, J.H.; Yun, S.H.; Oh, H.W.; Kang, J.H.; Koh, S.W.; Oh, H.J.; Song, K.J. Fruit growth, sugar, and acid characteristic in leafy and leafless fruits of Satsuma mandarin. Kor. J. Hortic. Sci. Technol. 2012, 30, 642–649. [Google Scholar]
- Raddatz-Mota, D.; Franco-Mora, O.; Mendoza-Espinoza, J.A.; Rodríguez-Verástegui, L.L.; de León-Sánchez, F.D.; Rivera-Cabrera, F. Effect of different rootstocks on Persian lime (Citrus latifolia T.) postharvest quality. Sci. Hortic. 2019, 257, 108716. [Google Scholar] [CrossRef]
- Albrigo, L.G.; Stelinski, L.L.; Timmer, L.W. Citrus; CABI: Oxfordshire, UK, 2019. [Google Scholar]
- Reese, R.I.; Loo, R.C.J. Effect of N and K fertilization on internal and external fruit quality of three major Florida orange cultivars. J. Am. Soc. Hortic. Sci. 1975, 100, 425–428. [Google Scholar]
- Liao, L.; Dong, T.; Qiu, X.; Rong, Y.; Qang, Z.; Zhu, J. Nitrogen nutrition is a key modulator of the sugar and organic acid content in citrus fruit. PLoS ONE 2019, 14, e0223356. [Google Scholar] [CrossRef]
- Komatsu, A.; Moriguchi, T.; Koyama, K.; Omura, M.; Akihama, T. Analysis of sucrose synthase genes in citrus suggests different roles and phylogenetic relationships. J. Exp. Bot. 2002, 53, 61–71. [Google Scholar] [CrossRef]
- Barry, G.H.; Castle, W.S.; Davies, F.S. Rootstocks and plant water relations affect sugar accumulation of citrus fruit via osmotic adjustment. J. Am. Soc. Hortic. Sci. 2014, 129, 881–889. [Google Scholar] [CrossRef]
- Hockema, B.R.; Etxeberria, E. Metabolic contributors to drought-enhanced accumulation of sugars and acids in orange. J. Am. Soc. Hortic. Sci. 2001, 126, 599–605. [Google Scholar] [CrossRef]
- Islam, M.Z.; Hu, X.M.; Jin, L.F.; Liu, Y.Z.; Peng, S.A. Genome-wide identification and expression profile analysis of citrus sucrose synthase genes: Investigation of possible roles in the regulation of sugar accumulation. PLoS ONE 2014, 9, e113623. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, A.; Takanokura, Y.; Akihama, T.; Omura, M. Cloning and molecular analysis of cDNAs encoding three sucrose phosphate synthase isoforms from a citrus fruit (Citrus unshiu Marc.). Mol. Gen. Genet. 1996, 252, 346–351. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rootstock | Tree Size | One-Year-Old Shoots | Water Sprout | |||||
|---|---|---|---|---|---|---|---|---|
| Height (m) | Width (m) | Length (cm) | Thickness (mm) | No. of Thorn | Thorn Length (cm) | Number | Length (cm) | |
| TO | 4.0 ± 0.1 | 1.5 ± 0.1 | 24.8 ± 0.7 | 2.99 ± 0.05 | 8.5 ± 0.4 | 1.5 ± 0.1 | 25.0 ± 3.3 | 221.1 ± 4.8 |
| FD | 1.4 ± 0.2 | 0.7 ± 0.1 | 9.8 ± 0.9 | 2.15 ± 0.11 | 1.2 ± 0.3 | 0.5 ± 0.0 | 10.5 ± 0.5 | 81.8 ± 15.8 |
| Significance | * | * | * | * | * | * | * | * |
| Rootstock | Number of Fruits Per Tree | Fresh Weight (g) | Yield (kg·m−2) |
|---|---|---|---|
| TO | 426 ± 51.7 | 46.5 ± 2.5 | 1.3 ± 0.2 |
| FD | 183 ± 47.0 | 61.8 ± 2.8 | 5.7 ± 0.8 |
| Significance | * | * | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, H.R.; Oh, E.U.; Han, S.G.; Yun, S.H.; Kim, H.B.; Song, K.J. Characterization of Soluble Sugar Content, Related Enzyme Activity and Gene Expression in the Fruits of ‘Minihyang’ Mandarin on Different Rootstocks. Horticulturae 2022, 8, 47. https://doi.org/10.3390/horticulturae8010047
Hong HR, Oh EU, Han SG, Yun SH, Kim HB, Song KJ. Characterization of Soluble Sugar Content, Related Enzyme Activity and Gene Expression in the Fruits of ‘Minihyang’ Mandarin on Different Rootstocks. Horticulturae. 2022; 8(1):47. https://doi.org/10.3390/horticulturae8010047
Chicago/Turabian StyleHong, Ha Rim, Eun Ui Oh, Seung Gab Han, Su Hyun Yun, Ho Bang Kim, and Kwan Jeong Song. 2022. "Characterization of Soluble Sugar Content, Related Enzyme Activity and Gene Expression in the Fruits of ‘Minihyang’ Mandarin on Different Rootstocks" Horticulturae 8, no. 1: 47. https://doi.org/10.3390/horticulturae8010047
APA StyleHong, H. R., Oh, E. U., Han, S. G., Yun, S. H., Kim, H. B., & Song, K. J. (2022). Characterization of Soluble Sugar Content, Related Enzyme Activity and Gene Expression in the Fruits of ‘Minihyang’ Mandarin on Different Rootstocks. Horticulturae, 8(1), 47. https://doi.org/10.3390/horticulturae8010047