
Use of Thermal Imaging to Assess Water Status in Citrus Plants in Greenhouses



Abstract
1. Introduction
2. Materials and Methods
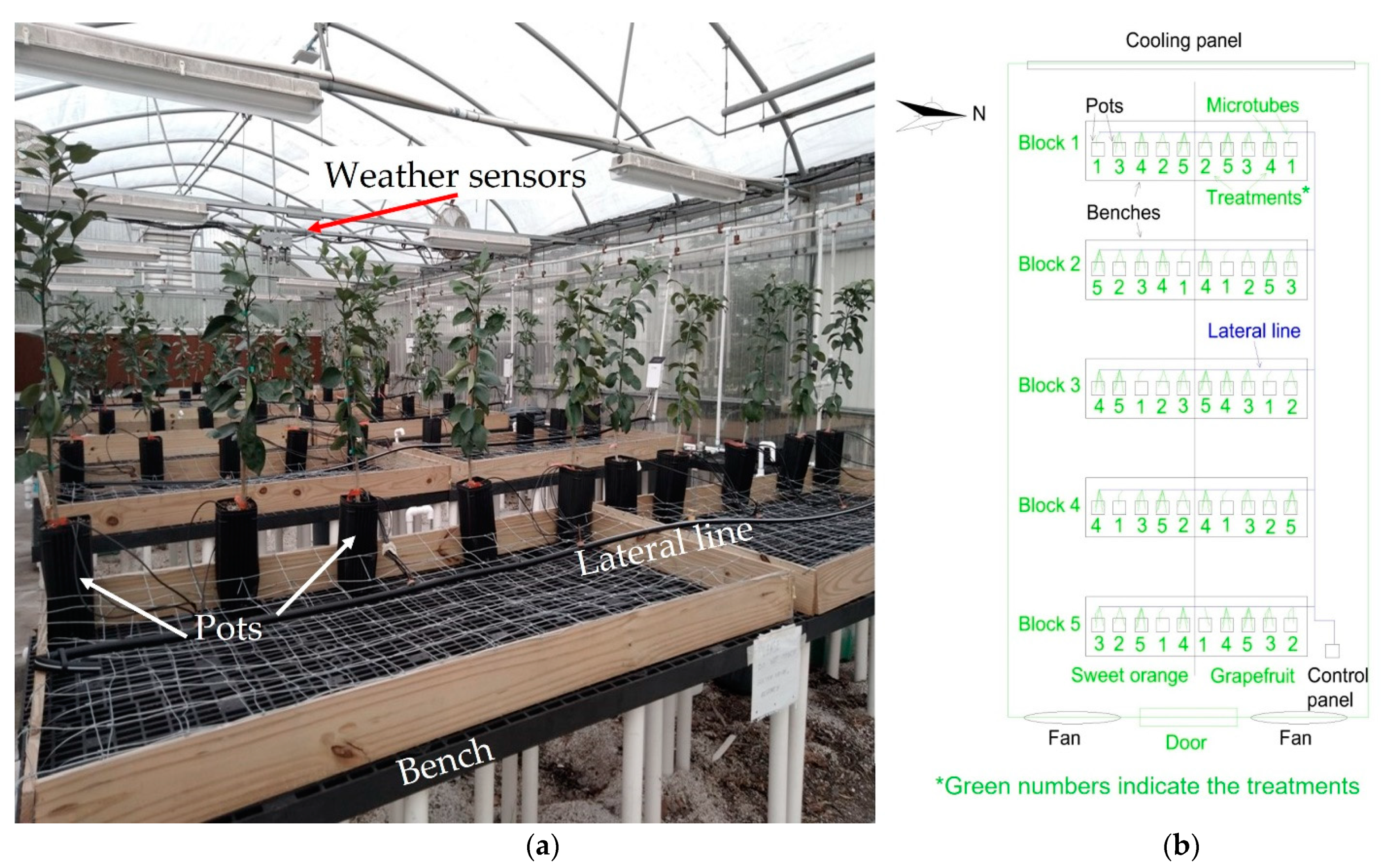
2.1. Location and Plant Material
2.2. Irrigation and Treatments
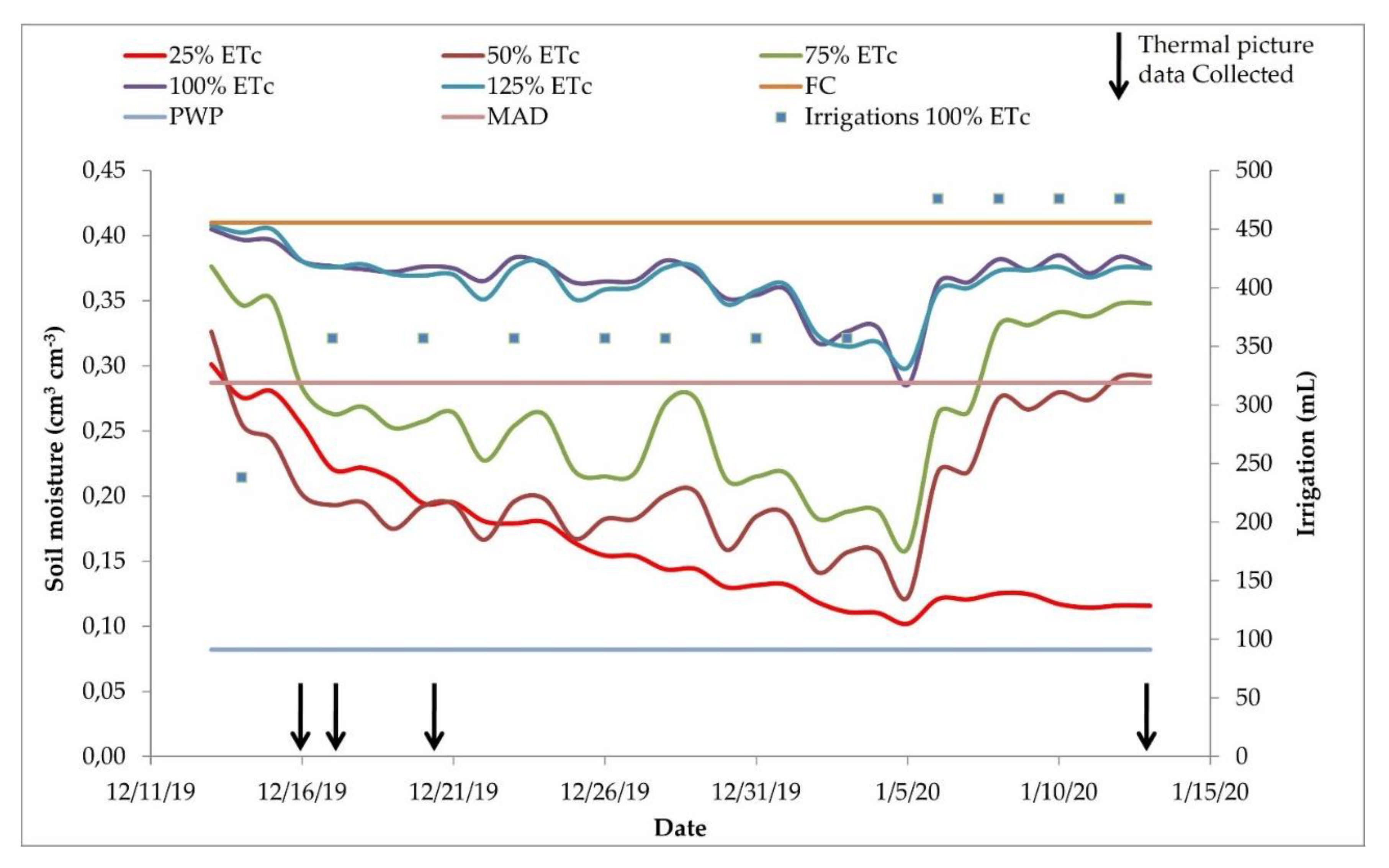
2.3. Soil Moisture
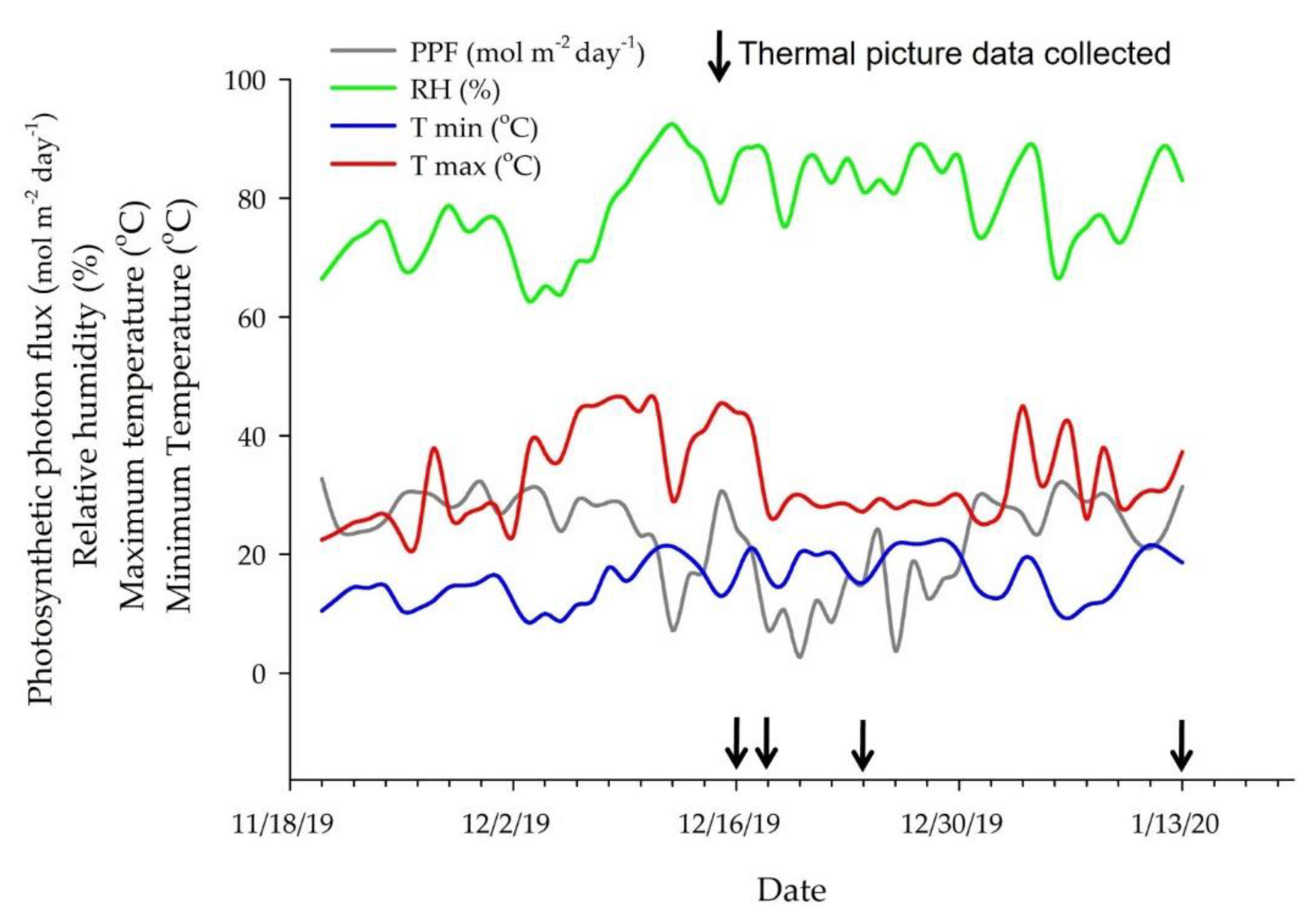
2.4. Measurements
2.4.1. Thermal Imaging
2.4.2. Photosynthesis and Transpiration
2.4.3. Biomass
2.4.4. Experimental Design and Statistical Analysis
3. Results
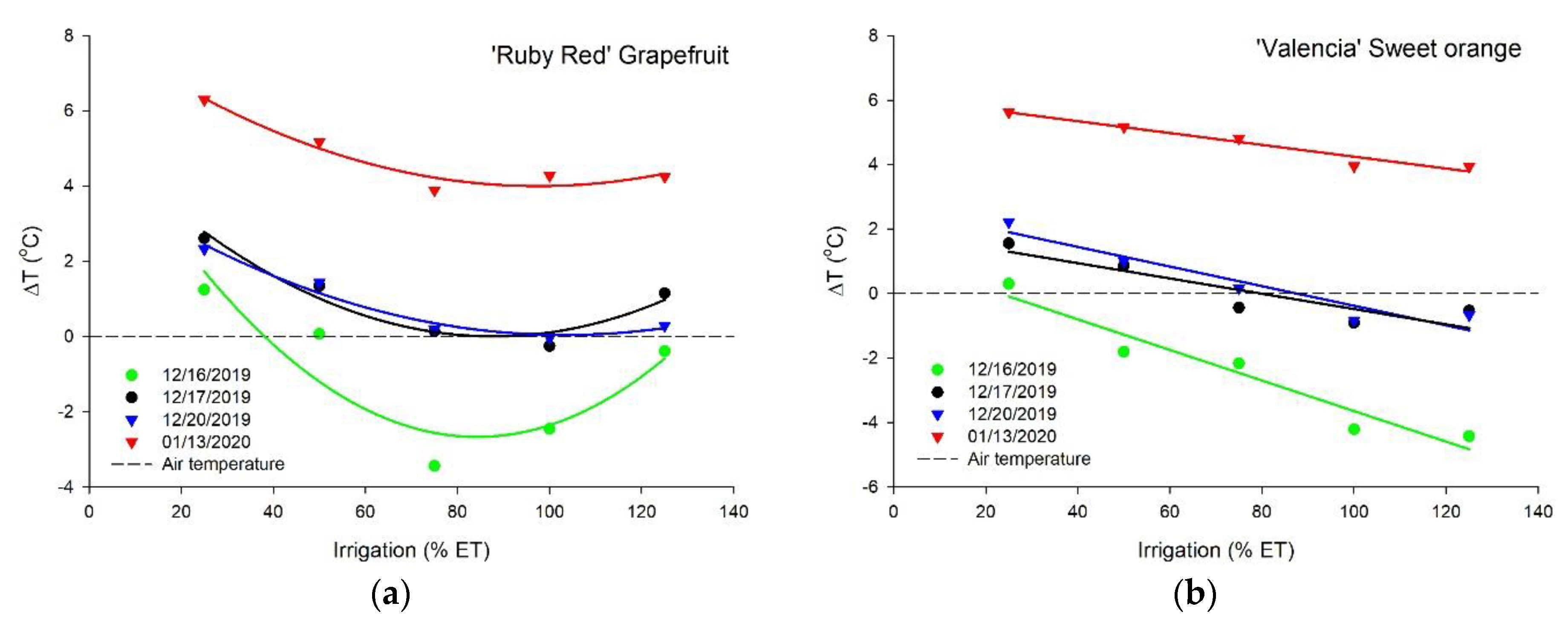
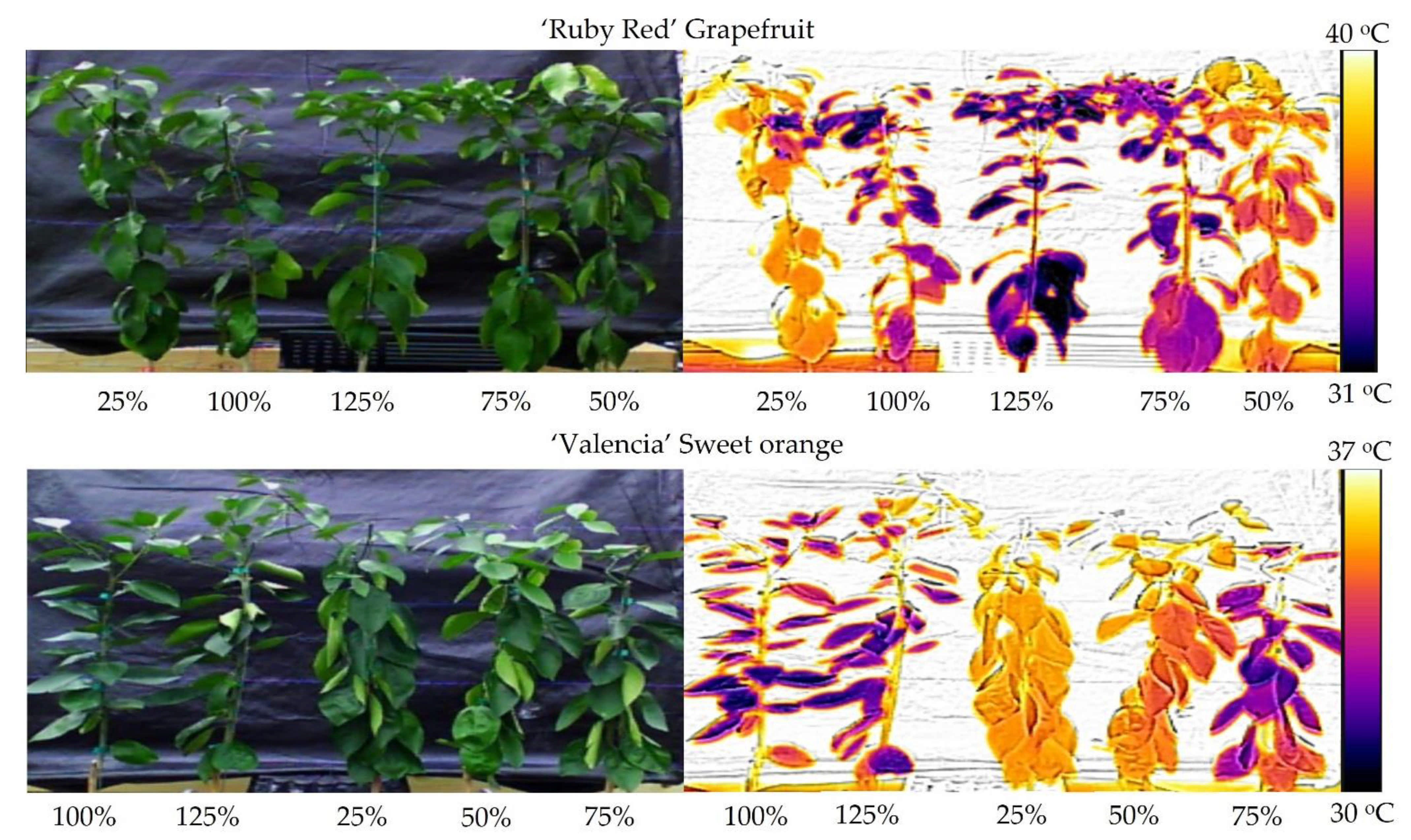
3.1. Thermal Imaging
3.2. Soil Moisture Monitoring
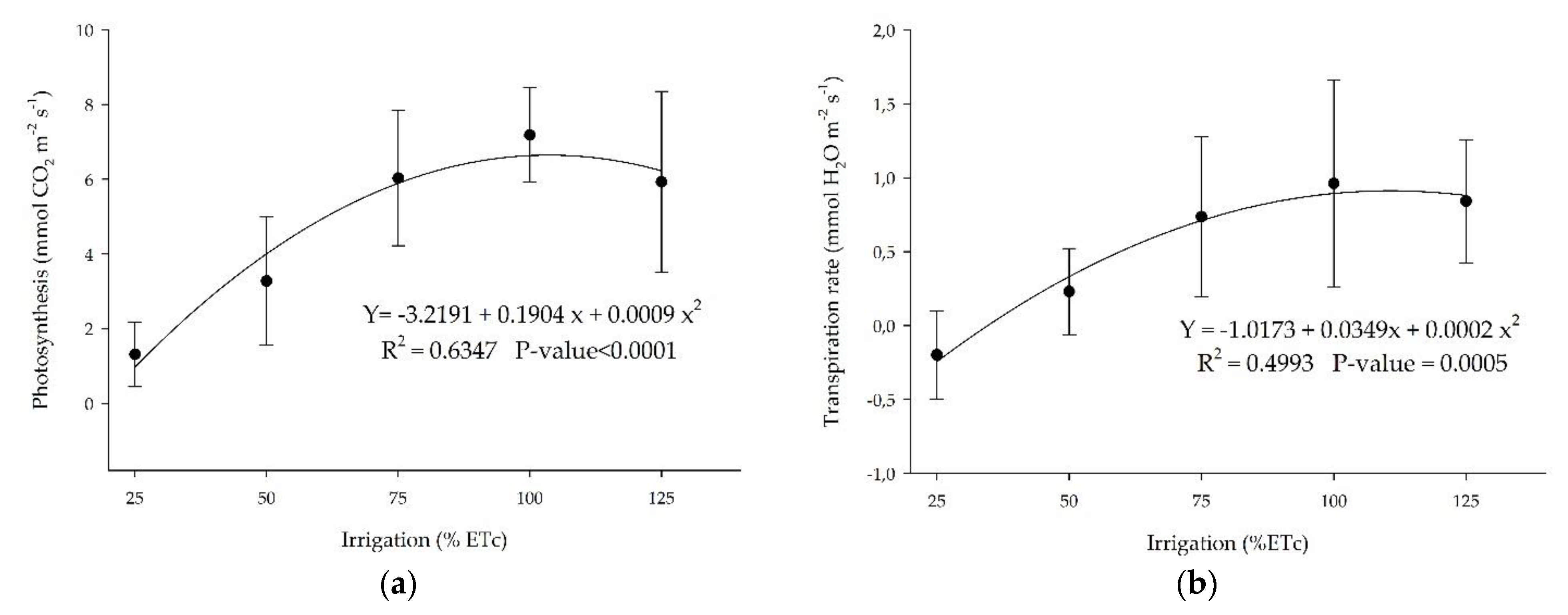
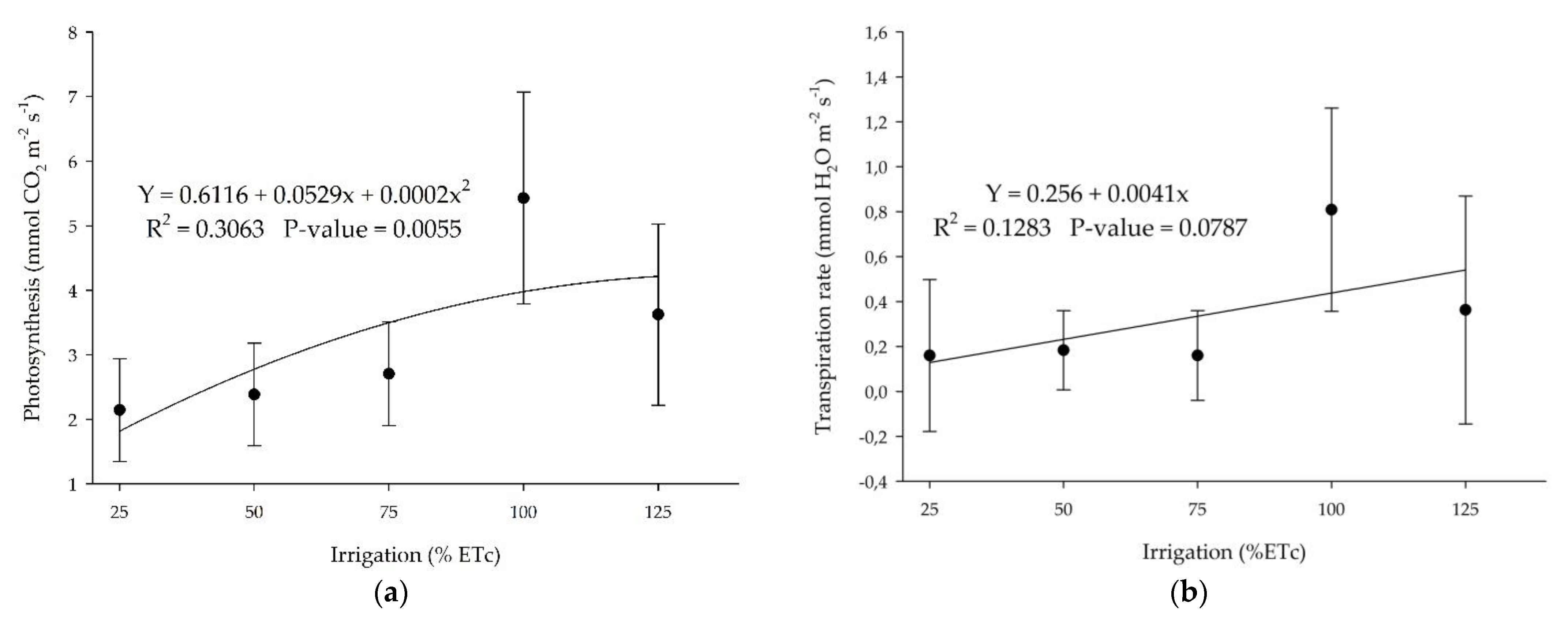
3.3. Photosynthesis and Transpiration
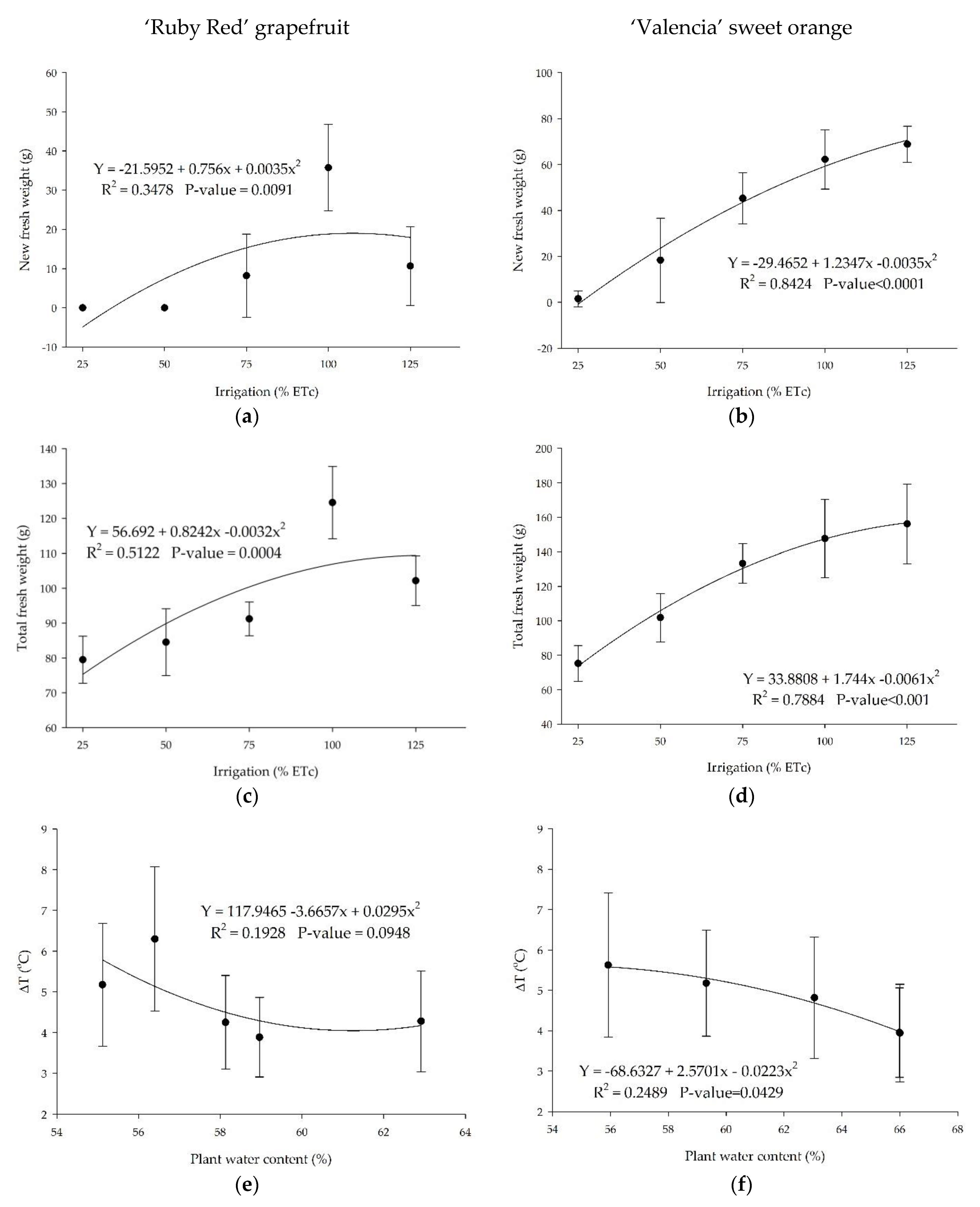
3.4. Biomass and Difference of Temperature
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García-Tejero, I.F.; Durán-Zuazo, V.H.; Muriel-Fernández, J.L.; Jiménez-Bocanegra, J.A. Linking canopy temperature and trunk diameter fluctuations with other physiological water status tools for water stress management in citrus orchards. Funct. Plant Biol. 2011, 38, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Naor, A. Irrigation scheduling and evaluation of tree water status in deciduous orchards. In Horticultural Reviews, 32nd ed.; Janick, J., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2006; pp. 111–165. [Google Scholar] [CrossRef]
- Velez, J.E.; Intrigliolo, D.S.; Castel, J.R. Scheduling deficit irrigation of citrus trees with maximum daily trunk shrinkage. Agric. Water Manag. 2007, 90, 197–204. [Google Scholar] [CrossRef]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J., Jr.; Reginato, R.J.; Hatfield, J.L. Normalizing the stress-degree-day parameter for environmental variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Clawson, K.L.; Blad, B.L. Infrared thermometry for scheduling irrigation of corn. Agron. J. 1982, 74, 311–316. [Google Scholar] [CrossRef]
- Testi, L.; Goldhamer, D.A.; Iniesta, F.; Salinas, M. Crop water stress index is a sensitive water stress indicator in pistachio trees. Irrig. Sci. 2008, 26, 395–405. [Google Scholar] [CrossRef]
- Jackson, R.D.; Reginato, R.J.; Idso, S.B. Wheat canopy temperature: A practical tool for evaluating water requirements. Water Resour. Res. 1977, 13, 651–656. [Google Scholar] [CrossRef]
- Trentin, R.; Zolnier, S.; Ribeiró, A.; Steidle Neto, A.J. Transpiration and leaf temperature of sugarcane under different matric potential values. Eng. Agríc. 2011, 31, 1085–1095, (In Portuguese with English Abstract). [Google Scholar] [CrossRef]
- Gardner, B.R.; Blad, B.L.; Watts, D.G. Plant and air temperatures in differentially irrigated corn. Agric. Meteorol. 1981, 25, 207–217. [Google Scholar] [CrossRef]
- Jackson, R.D. Canopy temperature and crop water stress. In Advances in Irrigation; Hillel, D., Ed.; Academic Press: New York, NY, USA, 1982; pp. 43–85. [Google Scholar]
- Fuchs, M.; Tanner, C.B. Infrared thermometry of vegetation. Agron. J. 1966, 58, 597–601. [Google Scholar] [CrossRef]
- Lobo, F.A.; Oliva, M.A.; Resende, M.; Lopes, N.F.; Maestri, M. Infrared thermometry to schedule irrigation of common bean. Pesqui. Agropecu. Bras. 2004, 39, 113–121. [Google Scholar] [CrossRef][Green Version]
- Johnson, K.; Sankaran, S.; Ehsani, R. Identification of water stress in citrus leaves using sensing technologies. Agronomy 2013, 3, 747–756. [Google Scholar] [CrossRef]
- Ballester, C.; Castel, J.; Intrigliolo, D.S.; Castel, J.R. Response of Clementina de Nules citrus trees to regulated deficit irrigation. Yield components and fruit composition. Agric. Water Manag. 2011, 98, 1027–1032. [Google Scholar] [CrossRef]
- Jimenez-Bello, M.A.; Ballester, C.; Castel, J.R.; Intrigliolo, D.S. Development and validation of an automatic thermal imaging process for assessing plant water status. Agric. Water Manag. 2011, 98, 1497–1504. [Google Scholar] [CrossRef]
- Romano, G.; Zia, S.; Spreer, W.; Sanchez, C.; Cairns, J.; Araus, J.L.; Müller, J. Use of thermography for high throughput phenotyping of tropical maize adaptation in water stress. Comput. Electron. Agric. 2011, 79, 67–74. [Google Scholar] [CrossRef]
- Ghazouani, H.; Capodici, F.; Ciraolo, G.; Maltese, A.; Rallo, G.; Provenzano, G. Potential of Thermal Images and Simulation Models to Assess Water and Salt Stress: Application to Potato Crop in Central Tunisia. Chem. Eng. Trans. 2017, 58, 709–714. [Google Scholar] [CrossRef]
- González-Dugo, V.; Zarco-Tejada, P.J.; Nicolás, E.; Nortes, P.A.; Alarcón, J.J.; Intrigliolo, D.S.; Fereres, E. Using high-resolution UAV thermal imagery to assess the variability in the water status of five fruit tree species within a commercial orchard. Precis. Agric. 2013, 14, 660–678. [Google Scholar] [CrossRef]
- Ballester, C.; Jiménez-Bello, M.; Castel, J.; Intrigliolo, D. Usefulness of thermography for plant water stress detection in citrus and persimmon trees. Agric. For. Meteorol. 2013, 168, 120–129. [Google Scholar] [CrossRef]
- Bellvert, J.; Zarco-Tejada, P.J.; Fereres, E.; Girona, J. Mapping crop water stress index in a ‘Pinot-noir’ vineyard: Comparing ground measurements with thermal remote sensing imagery from an unmanned aerial vehicle. Precis. Agric. J. 2014, 15, 361–376. [Google Scholar] [CrossRef]
- O’Shaughnessy, S.A.; Evett, S.R.; Colaizzi, P.D.; Howell, T.A. Using radiation thermography and thermometry to evaluate crop water stress in soybean and cotton. Agric. Water Manag. 2011, 98, 1523–1535. [Google Scholar] [CrossRef]
- Viana, L.A.; Zambolim, L.; Sousa, T.V.; Tomaz, D.C. Potential use of thermal camera coupled in UAV for crop monitoring. Braz. J. Biosyst. Eng. 2018, 12, 286–298, (In Portuguese with English Abstract). [Google Scholar] [CrossRef]
- Xu, J.; Lv, Y.; Liu, X.; Dalson, T.; Yang, S.; Wu, J. Diagnosing Crop Water Stress of Rice using Infrared Thermal Imager under Water Deficit Condition. Int. J. Agric. Biol. 2016, 18, 565–572. [Google Scholar] [CrossRef]
- Xu, H.; Ying, Y. Application of infrared thermal imaging in the identification of citrus on trees. J. Infrared Millim. Waves 2004, 23, 353–356. [Google Scholar]
- Bulanon, D.; Burks, T.F.; Alchanatis, V. Study on temporal variation in citrus canopy using thermal imaging for citrus fruit detection. Biosyst. Eng. 2008, 101, 161–171. [Google Scholar] [CrossRef]
- Alchanatis, V.; Cohen, Y.; Cohen, S.; Moller, M.; Sprinstin, M.; Meron, M.; Tsipris, J.; Saranga, Y.; Sela, E. Evaluation of different approaches for estimating and mapping crop water status in cotton with thermal imaging. Precis. Agric. 2010, 11, 27–41. [Google Scholar] [CrossRef]
- Belfiore, N.; Vinti, R.; Lovat, L.; Chitarra, W.; Tomasi, D.; de Bei, R.; Meggio, F.; Gaiotti, F. Infrared Thermography to Estimate Vine Water Status: Optimizing Canopy Measurements and Thermal Indices for the Varieties Merlot and Moscato in Northern Italy. Agronomy 2019, 9, 821. [Google Scholar] [CrossRef]
- Gutiérrez, S.; Diago, M.P.; Fernández-Novales, J.; Tardaguila, J. Vineyard water status assessment using on-the-go thermal imaging and machine learning. PLoS ONE 2018, 13, e0192037. [Google Scholar] [CrossRef]
- Waller, P.; Yitayew, M. Irrigation and Drainage Engineering; Springer: New York, NY, USA, 2016; p. 742. [Google Scholar] [CrossRef]
- Keller, J.; Bliesner, R.D. Sprinkle and Trickle Irrigation; Springer: New York, NY, USA, 1990; p. 652. [Google Scholar]
- American Society of Agricultural Engineers (ASAE). Field Evaluation of Micro-Irrigation Systems (EP 458); ASAE: St. Joseph, MI, USA, 2001; pp. 792–797. [Google Scholar]
- Gonzalez-Dugo, V.; Zarco-Tejada, P.J.; Fereres, E. Applicability and limitations of using the crop water stress index as an indicator of water deficits in citrus orchards. Agric. For. Meteorol. 2014, 198–199, 94–104. [Google Scholar] [CrossRef]
- Pou, A.; Diago, M.P.; Medrano, H.; Baluja, J.; Tardaguila, J. Validation of thermal indices for water stress status identification in grapevine. Agric. Water Manag. 2014, 134, 60–72. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Rubio, A.E.; Viñuela, I.; Hernández, A.; Gutiérrez-Gordillo, S.; Rodríguez-Pleguezuelo, C.R.; Durán-Zuazo, V.H. Thermal imaging at plant level to assess the crop-water status in almond trees (cv. Guara) under deficit irrigation strategies. Agric. Water Manag. 2018, 208, 176–186. [Google Scholar] [CrossRef]
- Balfagón, D.; Zandalinas, S.I.; Baliño, P.; Muriach, M.; Gómez-Cadenas, A. Involvement of ascorbate peroxidase and heat shock proteins on citrus tolerance to combined conditions of drought and high temperatures. Plant Physiol. Biochem. 2018, 27, 194–199. [Google Scholar] [CrossRef]
- Shafqat, W.; Naqvi, S.A.; Maqbool, R.; Haider, M.S.; Jaskani, M.J.; Khan, I.A. Climate change and citrus. In Citrus; Intech Open: London, UK, 2021; 24p. [Google Scholar] [CrossRef]
- Shafqat, W.; Jaskani, M.J.; Maqbool, R.; Khan, A.S.; Ali, Z. Evaluation of citrus rootstocks against drought, heat, and combined stress based on growth and photosynthetic pigments. Int. J. Agric. Biol. 2019, 22, 1001–1009. [Google Scholar] [CrossRef]
- Ihuoma, S.O.; Madramootoo, C.A. Recent advances in crop water stress detection. Comput. Electron. Agric. 2017, 141, 267–275. [Google Scholar] [CrossRef]
- Vieira, G.H.S.; Mantovani, E.C.; Sediyama, G.C.; Delazari, F.T. Morpho-physiological indicators of water stress on sugarcane as a function of irrigation depths. Biosci. J. 2014, 30, 65–75, (In Portuguese with English Abstract). Available online: http://www.seer.ufu.br/index.php/biosciencejournal/article/view/15026 (accessed on 26 July 2021).
- Zandalinas, S.I.; Rivero, R.M.; Martínez, V.; Gómez-Cadenas, A.; Arbona, V. Tolerance of citrus plants to the combination of high temperatures and drought is associated with the increase in transpiration modulated by a reduction in abscisic acid levels. BMC Plant Biol. 2016, 16, 105. [Google Scholar] [CrossRef]
- Klein, T. The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviors. Funct. Ecol. 2014, 28, 1313–1320. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Regulation of citrus responses to the combined action of drought and high temperatures depends on the severity of water deprivation. Physiol. Plant. 2018, 162, 427–438. [Google Scholar] [CrossRef]
- Martin-St. Paul, N.; Delzon, S.; Cochard, H. Plant resistance to drought depends on timely stomatal closure. Ecol. Lett. 2017, 20, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Zaer-Ara, T.; Boroomand, N.; Sadat-Hosseini, M. Physiological and morphological response to drought stress in seedlings of ten citruses. Trees 2016, 30, 985–993. [Google Scholar] [CrossRef]
- De Souza, C.R.; Maroco, J.P.; dos Santos, T.P.; Rodrigues, M.L.; Lopes, C.M.; Pereira, J.S.; Chaves, M.M. Impact of deficit irrigation on water use efficiency and carbon isotope composition of field-grown grapevines under Mediterranean climate. J. Exp. Bot. 2005, 56, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; McAdam, S.A.M. Evolution of the stomatal regulation of plant water content. Plant Physiol. 2017, 174, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.G.; Stoll, M.; Santos, T.; de Sousa, C.; Chaves, M.M.; Grant, O.M. Use of infrared thermography for monitoring stomatal closure in the field: Application to the grapevine. J. Exp. Bot. 2002, 53, 2249–2260. [Google Scholar] [CrossRef]
- Möller, M.; Alchanatis, V.; Cohen, Y.; Meron, M.; Tsipris, J.; Ostrovsky, V. Use of thermal and visible imagery for estimating crop water status of irrigated grapevine. J. Exp. Bot. 2007, 58, 827–838. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Wei, Z.; Liu, F. ABA-mediated modulation of elevated CO2 on stomatal response to drought. Curr. Opin. Plant Biol. 2020, 56, 174–180. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Number of Drippers Per Pot | Emitter Discharge (L h−1) | Total Water Applied (L) |
|---|---|---|---|
| 25% ETc | 1 | 1.81 | 1.66 |
| 50% ETc | 2 | 3.43 | 3.14 |
| 75% ETc | 3 | 5.20 | 4.76 |
| 100% ETc | 4 | 7.15 | 6.55 |
| 125% ETc | 5 | 8.73 | 8.00 |
| Date | ‘Ruby Red’ Grapefruit | n | ||
|---|---|---|---|---|
| 16 December 2019 | Y = 6.2316 − 0.2114x + 0.0013x2 | R2 = 0.4490 | p = 0.0014 | 25 |
| 17 December 2019 | Y = 5.4340 − 0.1236x + 0.0007x2 | R2 = 0.4862 | p = 0.0007 | 25 |
| 20 December 2019 | Y = 4.2288 − 0.0812x + 0.0004x2 | R2 = 0.5331 | p = 0.0002 | 25 |
| 13 January 2019 | Y = 8.2156 − 0.0865x + 0.0004x2 | R2 = 0.3183 | p = 0.0148 | 25 |
| All dates | Y = 6.0275 − 0.1257x + 0.0007x2 | R2 = 0.1506 | p < 0.0001 | 100 |
| ‘Valencia’ sweet orange | ||||
| 16 December 2019 | Y = 1.0936 − 0.0474x | R2 = 0.7209 | p < 0.0001 | 25 |
| 17 December 2019 | Y = 1.8910 − 0.0237x | R2 = 0.2793 | p = 0.0066 | 25 |
| 20 December 2019 | Y = 2.6618 − 0.0304x | R2 = 0.6003 | p < 0.0001 | 25 |
| 13 January 2019 | Y = 6.0794 − 0.0183x | R2 = 0.2082 | p = 0.0219 | 25 |
| All dates | Y = 2.9315 − 0.0300x | R2 = 0.1220 | p < 0.0001 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, G.H.S.; Ferrarezi, R.S. Use of Thermal Imaging to Assess Water Status in Citrus Plants in Greenhouses. Horticulturae 2021, 7, 249. https://doi.org/10.3390/horticulturae7080249
Vieira GHS, Ferrarezi RS. Use of Thermal Imaging to Assess Water Status in Citrus Plants in Greenhouses. Horticulturae. 2021; 7(8):249. https://doi.org/10.3390/horticulturae7080249
Chicago/Turabian StyleVieira, Gustavo Haddad Souza, and Rhuanito Soranz Ferrarezi. 2021. "Use of Thermal Imaging to Assess Water Status in Citrus Plants in Greenhouses" Horticulturae 7, no. 8: 249. https://doi.org/10.3390/horticulturae7080249
APA StyleVieira, G. H. S., & Ferrarezi, R. S. (2021). Use of Thermal Imaging to Assess Water Status in Citrus Plants in Greenhouses. Horticulturae, 7(8), 249. https://doi.org/10.3390/horticulturae7080249