
The Effect of Mycorrhizal Inoculum and Phosphorus Treatment on Growth and Flowering of Ajania (Ajania pacifica (Nakai) Bremer et Humphries) Plant


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Measurements of Plant Growth and Flowering Parameters
2.3. Assessment of Root Mycorrhizal Colonization
2.4. Statistical Analysis
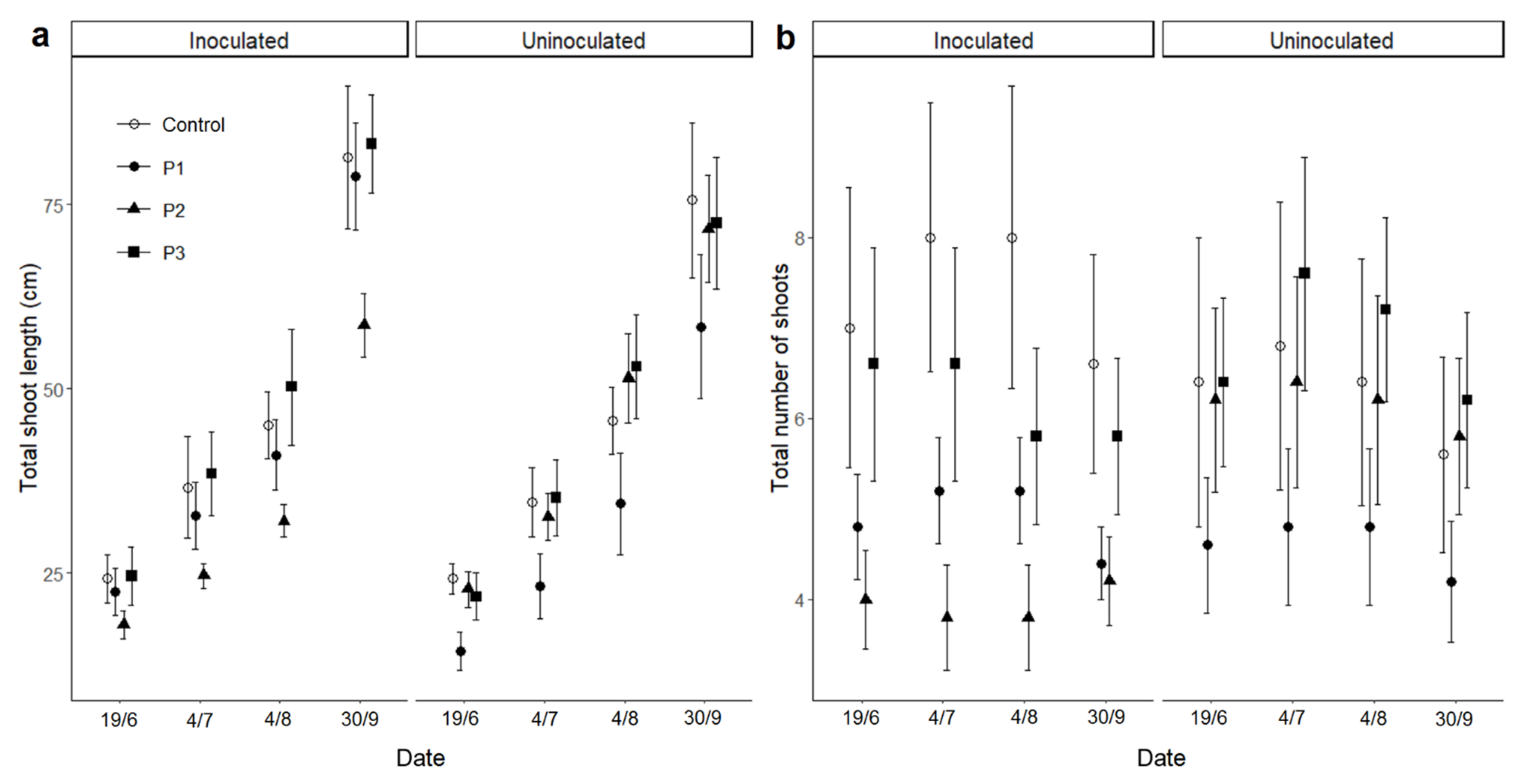
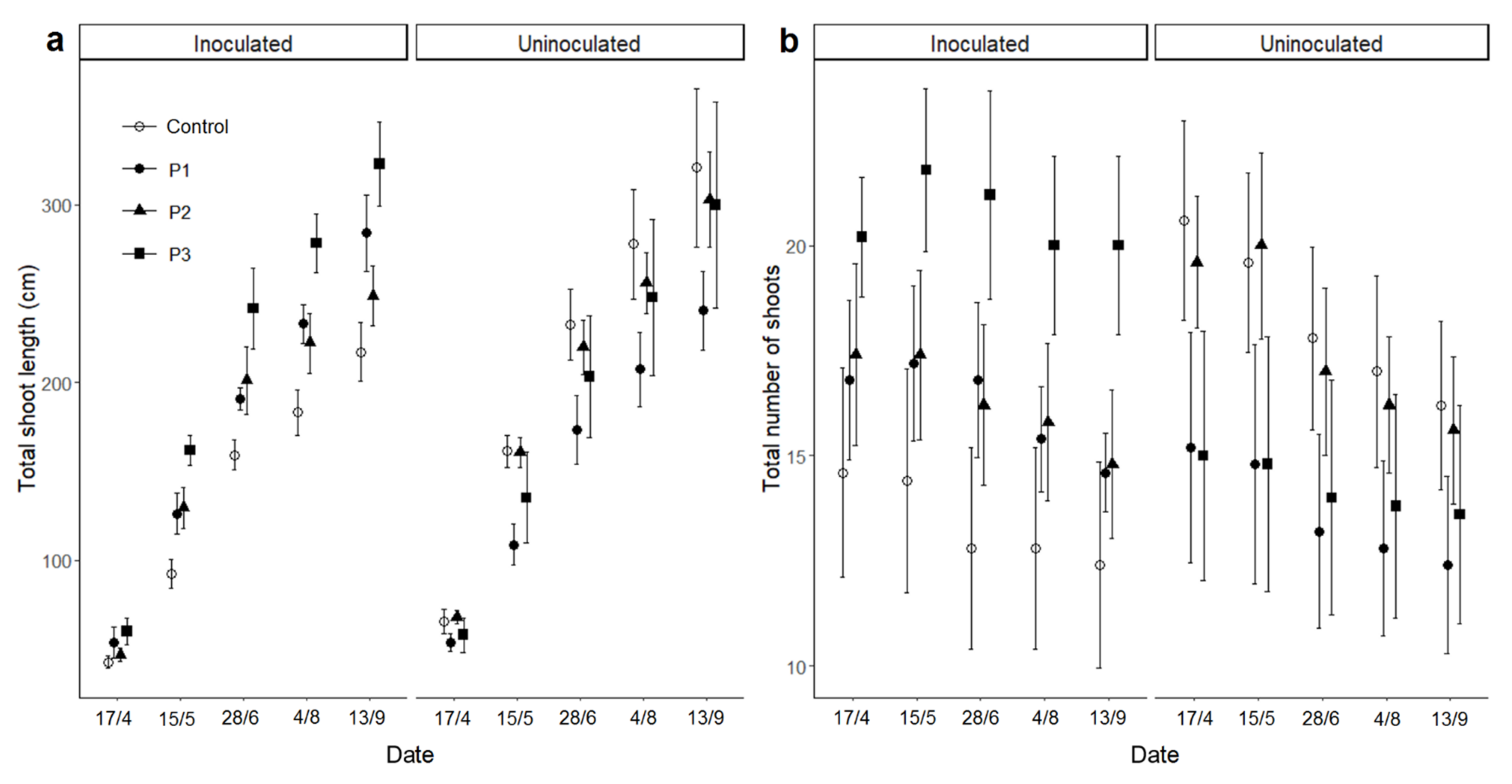
3. Results
3.1. Phosphorus Available to Plants
3.2. Mycorrhizal Colonization
3.3. Growth and Flowering
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Quilambo, O.A. The vesicular-arbuscular mycorrhizal symbiosis. Afr. J. Biotechnol. 2003, 2, 539–546. [Google Scholar]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: San Diego, CA, USA, 2008; p. 803. [Google Scholar]
- Gianinazzi-Pearson, V.; Maldonado-Mendoza, I.; Lopez-Meyer, M.; Weidmann, S.; Harrison, M.J. Arbuscular mycorrhiza. In The Medicago Truncatula Handbook; Mathesius, U., Journet, E.P., Sumner, L.W., Eds.; Noble Research Institute: Ardmore, OK, USA, 2006; Available online: http://www.noble.org/MedicagoHandbook/ (accessed on 27 April 2021).
- Barea, J.-M.; Ferrol, N.; Azcón-Aguilar, C.; Azcón, R. Mycorrhizal symbioses. In The Ecophysiology of Plant-Phosphorus Interactions; White, P.J., Hammond, J.P., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 7, pp. 143–163. [Google Scholar]
- Aggarwal, A.; Kadian, N.; Tanwar, A.; Yadav, A.; Gupta, K. Role of arbuscular mycorrhizal fungi (AMF) in global sustainable development. J. Nat. Appl. Sci. 2011, 3, 340–351. [Google Scholar] [CrossRef]
- Entry, J.A.; Rygiewicz, P.T.; Watrud, L.S.; Donnelly, P.K. Influence of adverse soil conditions on the formation and function of arbuscular mycorrhizas. Adv. Environ. Res. 2002, 7, 123–138. [Google Scholar] [CrossRef]
- Eschen, R.; Müller-Schärer, H.; Schaffner, U. Plant interspecific differences in arbuscular mycorrhizal colonization as a result of soil carbon addition. Mycorrhiza 2013, 23, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thangavelu, M.; Muthuraja, R. Arbuscular mycorrhizal and dark septate endophyte fungal associations in Asparagus. Turk. J. Bot. 2016, 40, 662–675. [Google Scholar] [CrossRef]
- Schüβler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: Phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Weishampel, P.A.; Bedford, B.L. Wetland dicots and monocots differ in colonization by arbuscular mycorrhizal fungi and dark septate endophytes. Mycorrhiza 2006, 16, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Jumpponen, A. Dark septate endophytes—Are they mycorrhizal? Mycorrhiza 2001, 11, 207–211. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Li, L.; Zhao, Z.W. The colonization of plants by dark septate endophytes (DSE) in the valley-type savanna of Yunnan, southwest China. Afr. J. Microbiol. Res. 2011, 5, 5540–5547. [Google Scholar] [CrossRef]
- Zhang, Q.; Gong, M.; Yuan, J.; Hou, Y.; Zhang, H.; Wang, Y.; Hou, X. Dark septate endophyte improves drought tolerance in Sorghum. Int. J. Agric. Biol. 2017, 19, 53–60. [Google Scholar] [CrossRef]
- Zubek, S.; Blaszkowski, J.; Mleczko, P. Arbuscular mycorrhizal and dark septate endophyte associations of medicinal plants. Acta Soc. Bot. Pol. 2011, 80, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Zalewska, M.; Antkowiak, M. Effect of pinching and day length on the growth and flowering of Ajania pacifica (Nakai) Bremer et Humphries in controlled cultivation. Acta Sci. Pol. Hortorum Cult. 2011, 10, 61–73. [Google Scholar]
- Zalewska, M.; Antkowiak, M. Gibberellic acid effect on growth and flowering of Ajania pacifica (Nakai) Bremer et Humphries. Hortic. Res. 2013, 21, 21–27. [Google Scholar] [CrossRef]
- McGraw, A.; Schenck, N. Growth stimulation of citrus, ornamental and vegetable crops by select mycorrhizal fungi. Proc. Fla. State Hortic. Soc. 1980, 93, 201–205. [Google Scholar]
- Sohn, B.K.; Kim, K.Y.; Chung, S.J.; Kim, W.S.; Park, S.M.; Kang, J.G.; Rim, Y.S.; Cho, J.S.; Kim, T.H.; Lee, J.H. Effect of the different timing of AMF inoculation on plant growth and flower quality of chrysanthemum. Sci. Hortic. 2003, 98, 173–183. [Google Scholar] [CrossRef]
- Gaur, A.; Gaur, A.; Adholeya, A. Growth and flowering in Petunia hybrida, Callistephus chinensis and Impatiens balsamina inoculated with mixed AM inocula or chemical fertilizers in a soil of low P fertility. Sci. Hortic. 2000, 84, 151–162. [Google Scholar] [CrossRef]
- Naik, B.H.; Nalawadi, U.; Sreenivasa, M.; Patil, A. Field responses of China aster (Callistephus chinensis (L) Nees.) cv. ‘Ostrich plume’ to the inoculation of vesicular arbuscular mycorrhizal fungi at different phosphorus levels. Sci. Hortic. 1995, 62, 129–133. [Google Scholar] [CrossRef]
- Asrar, A.; Abdel-Fattah, G.M.; Elhindi, K.M.; Abdel-Salam, E.M. The impact of arbuscular mychorrhizal fungi in improving growth, flower yield and tolerance of kalanchoe (Kalanchoe blossfeldiana Poelin) plants grown in NaCl-stress conditions. J. Food Agric. Environ. 2014, 12, 105–112. [Google Scholar]
- Asrar, A.; Abdel-Fattah, G.; Elhindi, K. Improving growth, flower yield, and water relations of snapdragon (Antirhinum majus L.) plants grown under well-watered and water-stress conditions using arbuscular mycorrhizal fungi. Photosynthetica 2012, 50, 305–316. [Google Scholar] [CrossRef]
- Navarro, A.; Elia, A.; Conversa, G.; Campi, P.; Mastrorilli, M. Potted mycorrhizal carnation plants and saline stress: Growth, quality and nutritional plant responses. Sci. Hortic. 2012, 140, 131–139. [Google Scholar] [CrossRef]
- Banuelos, J.; Alarcón, A.; Larsen, J.; Cruz-Sánchez, S.; Trejo, D. Interactions between arbuscular mycorrhizal fungi and Meloidogyne incognita in the ornamental plant Impatiens balsamina. J. Soil Sci. Plant Nutr. 2014, 14, 63–74. [Google Scholar]
- Pagliarini, N.; Jurjević, Ž.; Vinceljak Toplak, M.; Ostojić, Z.; Vršek, I. Sve o Krizantemi; Hrvatsko Agronomsko Drustvo: Zagreb, Croatia, 1997; p. 170. [Google Scholar]
- Egnér, H.; Riehm, H.; Domingo, W. Untersuchungen über die chemische Bodenanalyse als Grundlage für die Beurteilung des Nährstoffzustandes der Böden. II. Chemische Extraktionsmethoden zur Phosphor-und Kaliumbestimmung. Lantbrukshögskolans Ann. 1960, 26, 199–215. [Google Scholar]
- Phillips, J.M.; Hayman, D. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du Taux de Mycorhization VA d’un Système Radiculaire Recherche de Methods D’estimation Ayant une Signification Fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; INRA Publications: Paris, France, 1986; pp. 217–221. [Google Scholar]
- Alarcón, C.; Cuenca, G. Arbuscular mycorrhizas in coastal sand dunes of the Paraguanä Peninsula, Venezuela. Mycorrhiza 2005, 16, 1–9. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing Computer Program, Version 3.6.1; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.r-project.org (accessed on 25 February 2021).
- Jamiołkowska, A.; Księżniak, A.; Gałązka, A.; Hetman, B.; Kopacki, M.; Skwaryło-Bednarz, B. Impact of abiotic factors on development of the community of arbuscular mycorrhizal fungi in the soil: A Review. Int. Agrophys. 2018, 32, 133–140. [Google Scholar] [CrossRef]
- Pongrac, P.; Vogel-Mikuš, K.; Kump, P.; Nečemer, M.; Tolrà, R.; Poschenrieder, C.; Barceló, J.; Regvar, M. Changes in elemental uptake and arbuscular mycorrhizal colonisation during the life cycle of Thlaspi praecox Wulfen. Chemosphere 2007, 69, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- García, I.; Mendoza, R.; Pomar, M.C. Arbuscular mycorrhizal symbiosis and dark septate endophytes under contrasting grazing modes in the Magellanic steppe of Tierra del Fuego. Agric. Ecosyst. Environ. 2012, 155, 194–201. [Google Scholar] [CrossRef]
- Cronk, J.K.; Fennessy, M.S. Wetland Plants, Biology and Ecology; CRC Press: Boca Raton, FL, USA, 2001; pp. 1–462. [Google Scholar]
- Azcón-Aguilar, C.; Barea, J. Applying mycorrhiza biotechnology to horticulture: Significance and potentials. Sci. Hortic. 1997, 68, 1–24. [Google Scholar] [CrossRef]
- Shukla, A.; Kumar, A.; Jha, A.; Rao, D.N. Phosphorus threshold for arbuscular mycorrhizal colonization of crops and tree seedlings. Biol. Fertil. Soils 2012, 48, 109–116. [Google Scholar] [CrossRef]
- Habte, M.; Manjunath, A. Categories of vesicular-arbuscular mycorrhizal dependency of host species. Mycorrhiza 1991, 1, 3–12. [Google Scholar] [CrossRef]
- Kivelä, J.; Chen, L.; Muurinen, S.; Kivijärvi, P.; Hintikainen, V.; Helenius, J. Effects of meat bone meal as fertilizer on yield and quality of sugar beet and carrot. Agric. Food Sci. 2015, 24, 68–83. [Google Scholar] [CrossRef]
- Chen, L.; Kivelä, J.; Helenius, J.; Kangas, A. Meat bone meal as fertiliser for barley and oat. Agric. Food Sci. 2011, 20, 235–244. [Google Scholar] [CrossRef]
- Jeng, A.S.; Haraldsen, T.K.; Grønlund, A.; Pedersen, P.A. Meat and bone meal as nitrogen and phosphorus fertilizer to cereals and rye grass. Nutr. Cycl. Agroecosyst. 2006, 76, 183–191. [Google Scholar] [CrossRef]
- Nogalska, A.; Zalewska, M. The effect of meat and bone meal on phosphorus concentrations in soil and crop plants. Plant Soil Environ. 2013, 59, 575–580. [Google Scholar] [CrossRef] [Green Version]
- Bøen, A.; Haraldsen, T.K. Meat and bone meal and biosolids as slow-release phosphorus fertilizers. Agric. Food Sci. 2013, 22, 235–246. [Google Scholar] [CrossRef]
- Cekic, C.; Yilmaz, E. Effect of arbuscular mycorrhiza and different doses of phosphor on vegetative and generative components of strawberries applied with different phosphor doses in soilless culture. Afr. J. Agric. Res. 2011, 6, 4736–4739. [Google Scholar]
- Deng, Y.; Feng, G.; Chen, X.; Zou, C. Arbuscular mycorrhizal fungal colonization is considerable at optimal Olsen-P levels for maximized yields in an intensive wheat-maize cropping system. Field Crops Res. 2017, 209, 1–9. [Google Scholar] [CrossRef]
- Balzergue, C.; Chabaud, M.; Barker, D.G.; Bécard, G.; Rochange, S.F. High phosphate reduces host ability to develop arbuscular mycorrhizal symbiosis without affecting root calcium spiking responses to the fungus. Front. Plant Sci. 2013, 4, 426. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Tullio, M.; Rivera, C.M.; Rea, E. Alleviation of salt stress by arbuscular mycorrhizal in zucchini plants grown at low and high phosphorus concentration. Biol. Fertil. Soils 2008, 44, 501–509. [Google Scholar] [CrossRef]
- Oh, W.; Kim, J.; Kim, Y.H.; Lee, I.-J.; Kim, K.S. Shoot elongation and gibberellin contents in Cyclamen persicum are influenced by temperature and light intensity. Hortic. Environ. Biotechnol. 2015, 56, 762–768. [Google Scholar] [CrossRef]
- Zhiyu, M.; Shimizu, H.; Moriizumi, S.; Miyata, M.; Douzono, M.; Tazawa, S. Effect of light intensity, quality and photoperiod on stem elongation of chrysanthemum cv. Reagan. Environ. Control. Biol. 2007, 45, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Anderson, N.O. Chrysanthemum (Dendranthema × grandiflora Tzvelv.). In Flower Breeding and Genetics; Anderson, N.O., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 389–437. [Google Scholar]
- Elhindi, K.M.; Al-Mana, F.A.; El-Hendawy, S.; Al-Selwey, W.A.; Elgorban, A.M. Arbuscular mycorrhizal fungi mitigates heavy metal toxicity adverse effects in sewage water contaminated soil on Tagetes erecta L. Soil Sci. Plant Nutr. 2018, 64, 662–668. [Google Scholar] [CrossRef] [Green Version]
- Saini, I.; Aggarwal, A.; Kaushik, P. Inoculation with mycorrhizal fungi and other microbes to improve the morpho-physiological and floral traits of Gazania rigens (L.) Gaertn. Agriculture 2019, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Garmendia, I.; Mangas, V. Application of arbuscular mycorrhizal fungi on the production of cut flower roses under commercial-like conditions. Span. J. Agric. Res. 2012, 1, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Scagel, C.; Schreiner, R.P. Phosphorus supply alters tuber composition, flower production, and mycorrhizal responsiveness of container-grown hybrid Zantedeschia. Plant Soil 2006, 283, 323–337. [Google Scholar] [CrossRef]
- Aboul-Nasr, A. Effects of vesicular-arbuscular mycorrhiza on Tagetes erecta and Zinnia elegans. Mycorrhiza 1995, 6, 61–64. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fertilization Treatment | Superphosphate Fertilizer | P2O5 | 0-0-60 Fertilizer | K2O | KAN-Kutina | Calcium Ammonium Nitrate |
|---|---|---|---|---|---|---|
| Control | 0 | 0 | 20 | 12 | 150 | 40.5 |
| P1 | 20 | 5.2 | 20 | 12 | 150 | 40.5 |
| P2 | 40 | 10.4 | 20 | 12 | 150 | 40.5 |
| P3 | 80 | 20.8 | 20 | 12 | 150 | 40.5 |
| Main Effect | Phosphorus Available to Plants |
|---|---|
| Phosphorus Treatment | |
| Control | 1367 ± 126 c |
| P1 | 1630 ± 135 b |
| P2 | 1852 ± 133 a |
| P3 | 1938 ± 130 a |
| Time | |
| 3 July 2015 | 1732 ± 114 b |
| 30 September 2015 | 2434 ± 93 a |
| 27 July 2016 | 1459 ± 67 c |
| 11 October 2016 | 1163 ± 79 d |
| ANOVA | |
| Phosphorus treatment | *** |
| Time | *** |
| Phosphorus treatment × Time | NS |
| Main Effect | F% | M% | A% | V% | C% | MS% |
|---|---|---|---|---|---|---|
| Time | ||||||
| 2015 | 100 ± 0.0 | 22.9 ± 3.1 b | 0.0 ± 0.0 | 0.1 ± 0.0 b | 0.01 ± 0.0 b | 0.7 ± 0.2 b |
| 2016 | 100 ± 0.0 | 38.1 ± 3.4 a | 0.1 ± 0.0 | 1.0 ± 0.3 a | 2.7 ± 0.4 a | 2.9 ± 0.4 a |
| ANOVA | ||||||
| Phosphorus treatment | NS | NS | NS | NS | NS | NS |
| Time | NS | ** | NS | * | *** | *** |
| Phosphorus treatment × Time | NS | NS | NS | NS | NS | NS |
| Main Effect | Number of Inflorescences | Number of Flowers | Days of Flowering |
|---|---|---|---|
| Mycorrhizal inoculum | |||
| Inoculated plants | 1.0 ± 0.2 a | 29.8 ± 6.5 a | 26.2 ± 5.4 a |
| Uninoculated plants | 0.2 ± 0.1 b | 6.4 ± 2.4 b | 8.0 ± 2.8 b |
| ANOVA | |||
| Inoculum | ** | ** | ** |
| Phosphorus treatment | NS | NS | NS |
| Time | NS | NS | NS |
| Inoculum × Phosphorus treatment | NS | NS | NS |
| Inoculum × Time | NS | NS | NS |
| Phosphorus treatment × Time | NS | NS | NS |
| Inoculum × Phosphorus treatment × Time | NS | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vosnjak, M.; Likar, M.; Osterc, G. The Effect of Mycorrhizal Inoculum and Phosphorus Treatment on Growth and Flowering of Ajania (Ajania pacifica (Nakai) Bremer et Humphries) Plant. Horticulturae 2021, 7, 178. https://doi.org/10.3390/horticulturae7070178
Vosnjak M, Likar M, Osterc G. The Effect of Mycorrhizal Inoculum and Phosphorus Treatment on Growth and Flowering of Ajania (Ajania pacifica (Nakai) Bremer et Humphries) Plant. Horticulturae. 2021; 7(7):178. https://doi.org/10.3390/horticulturae7070178
Chicago/Turabian StyleVosnjak, Matej, Matevz Likar, and Gregor Osterc. 2021. "The Effect of Mycorrhizal Inoculum and Phosphorus Treatment on Growth and Flowering of Ajania (Ajania pacifica (Nakai) Bremer et Humphries) Plant" Horticulturae 7, no. 7: 178. https://doi.org/10.3390/horticulturae7070178
APA StyleVosnjak, M., Likar, M., & Osterc, G. (2021). The Effect of Mycorrhizal Inoculum and Phosphorus Treatment on Growth and Flowering of Ajania (Ajania pacifica (Nakai) Bremer et Humphries) Plant. Horticulturae, 7(7), 178. https://doi.org/10.3390/horticulturae7070178