
Impact of Elevated CO2 and Temperature on Growth, Development and Nutrient Uptake of Tomato


 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Details
2.2. Growth Indicating Parameters
2.2.1. Leaf Area (cm2)
2.2.2. Leaf Area Index
2.2.3. Leaf Area Duration (Days)
2.2.4. Crop Growth Rate (g/m2/day)
2.2.5. Radiation Interception (MJ/m2)
2.3. Phenological Observations
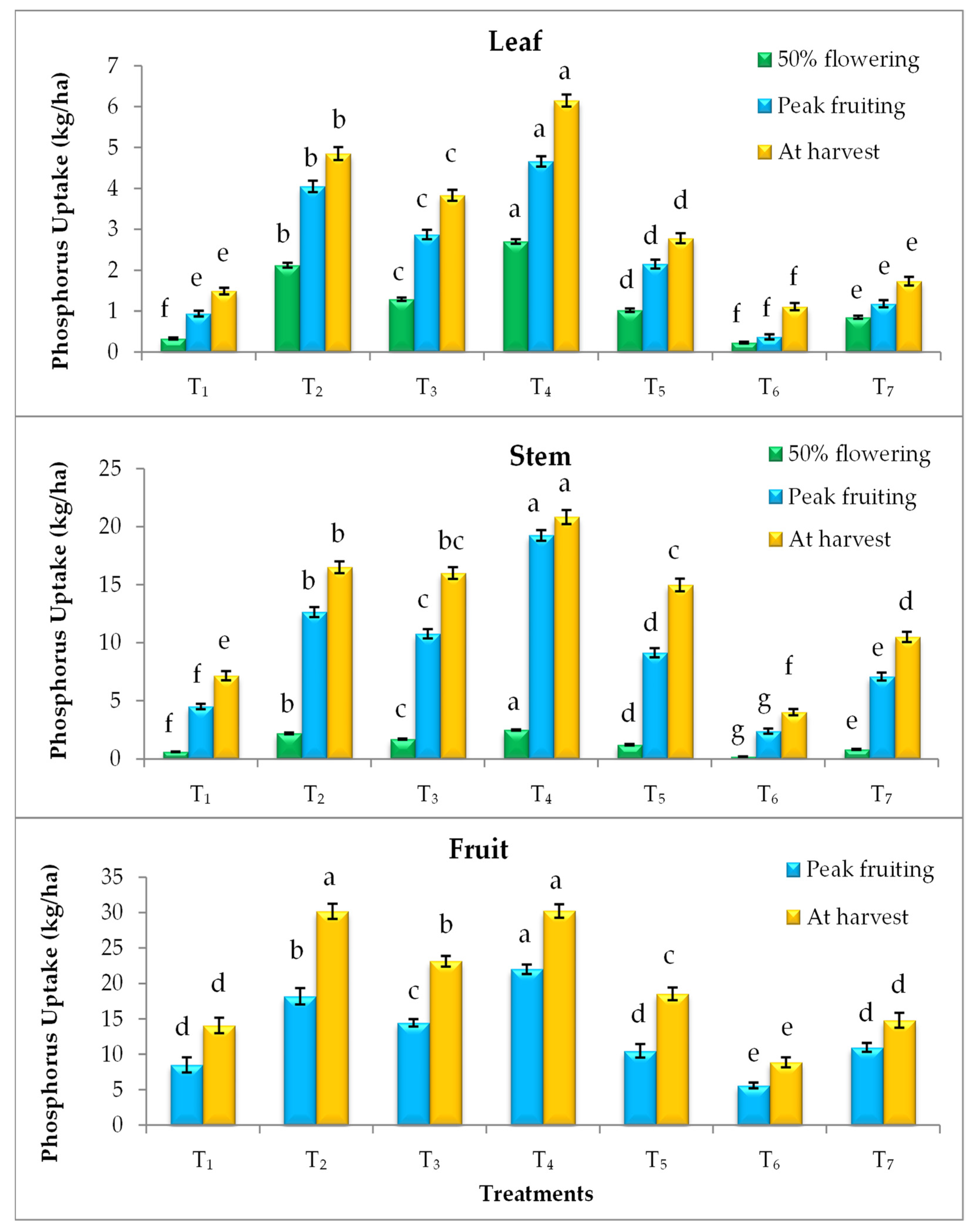
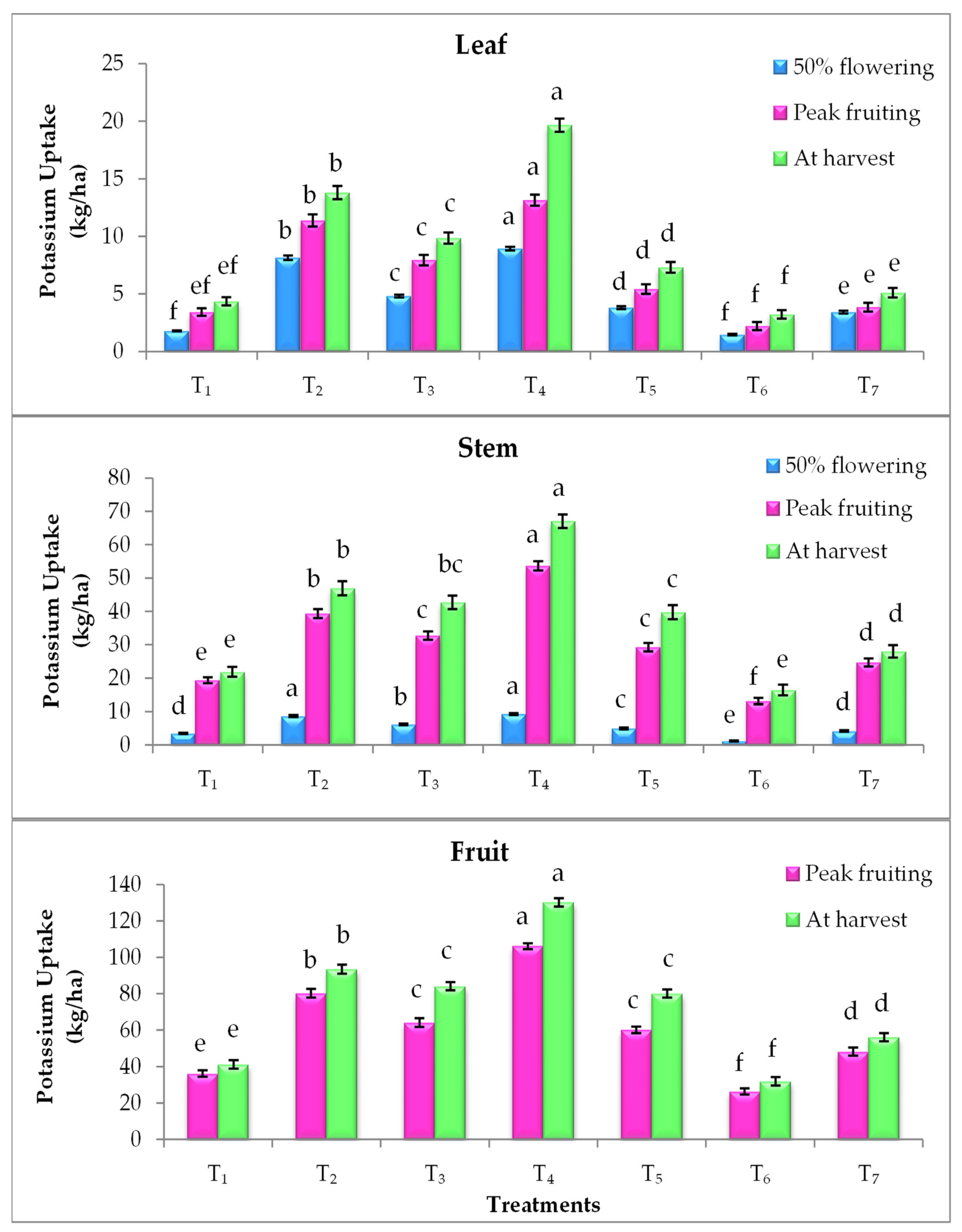
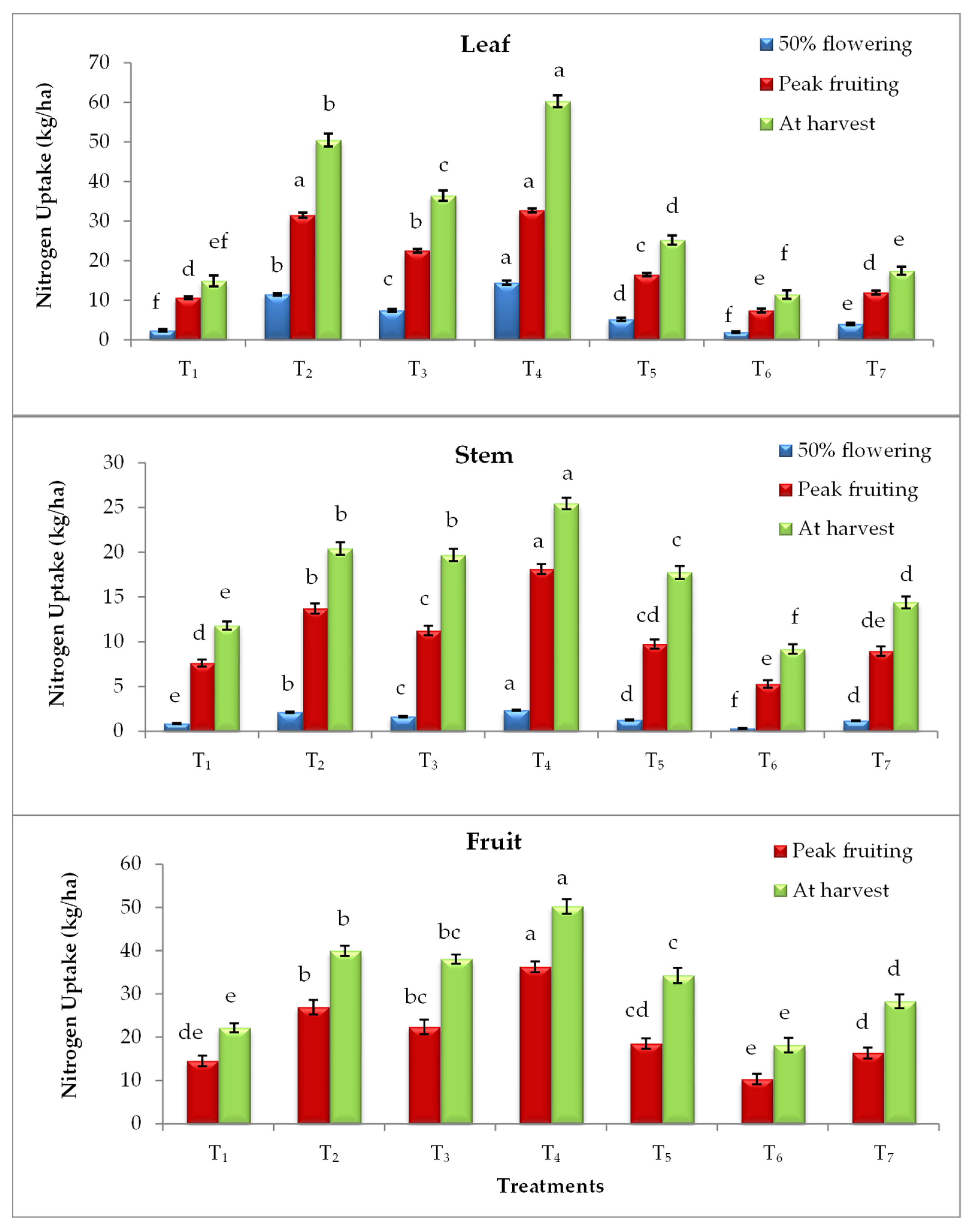
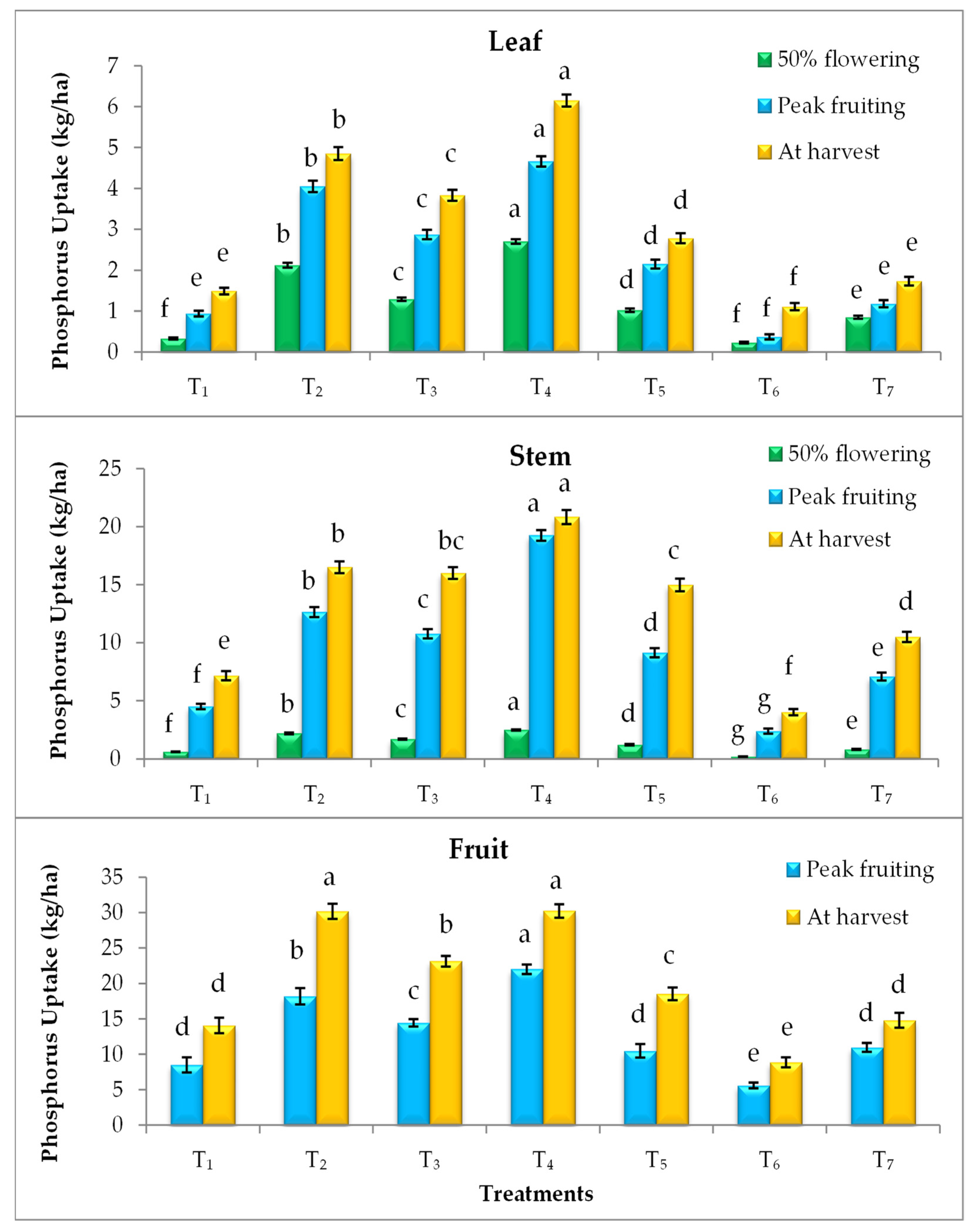
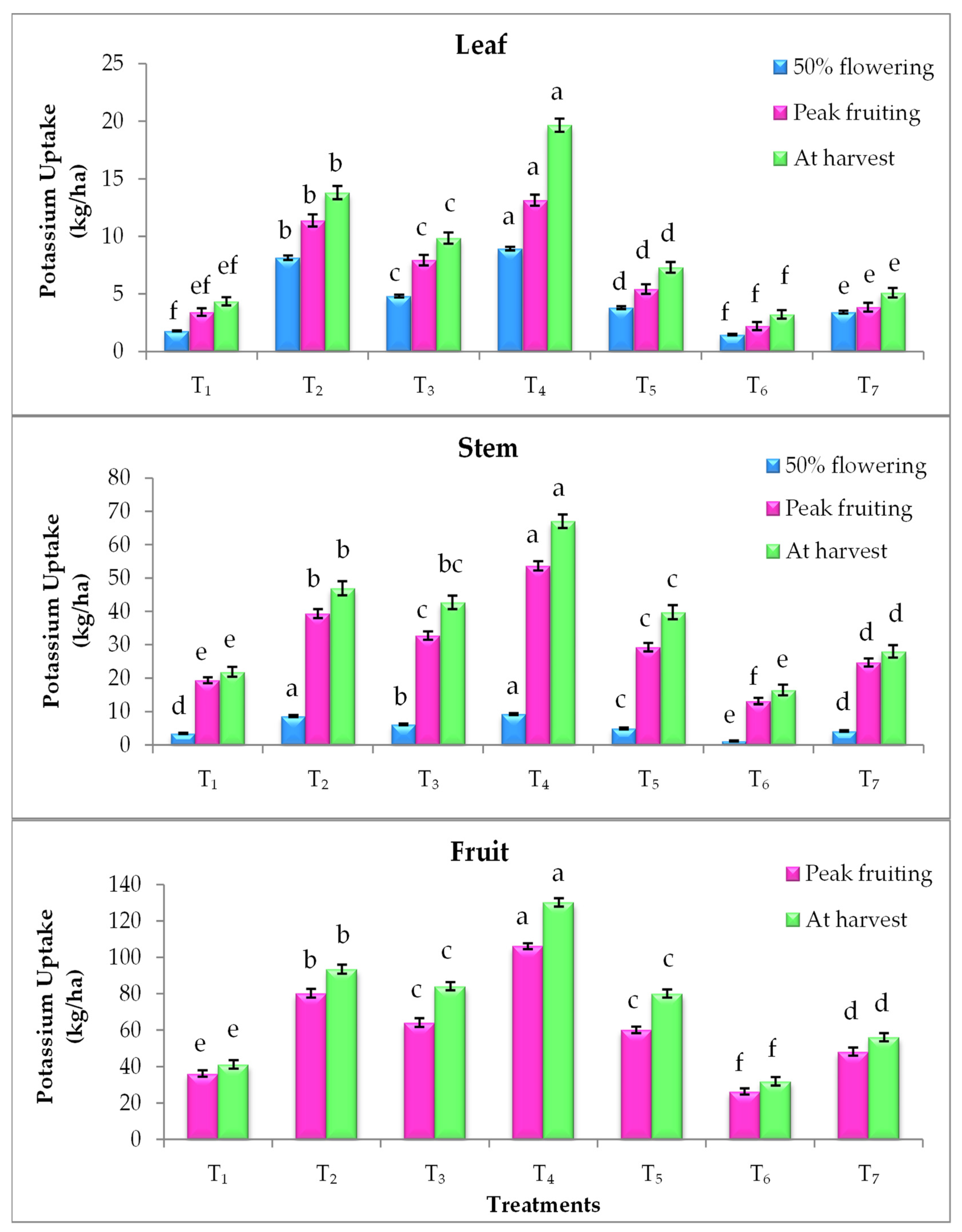
2.4. Nutrient Uptake (kg/ha)
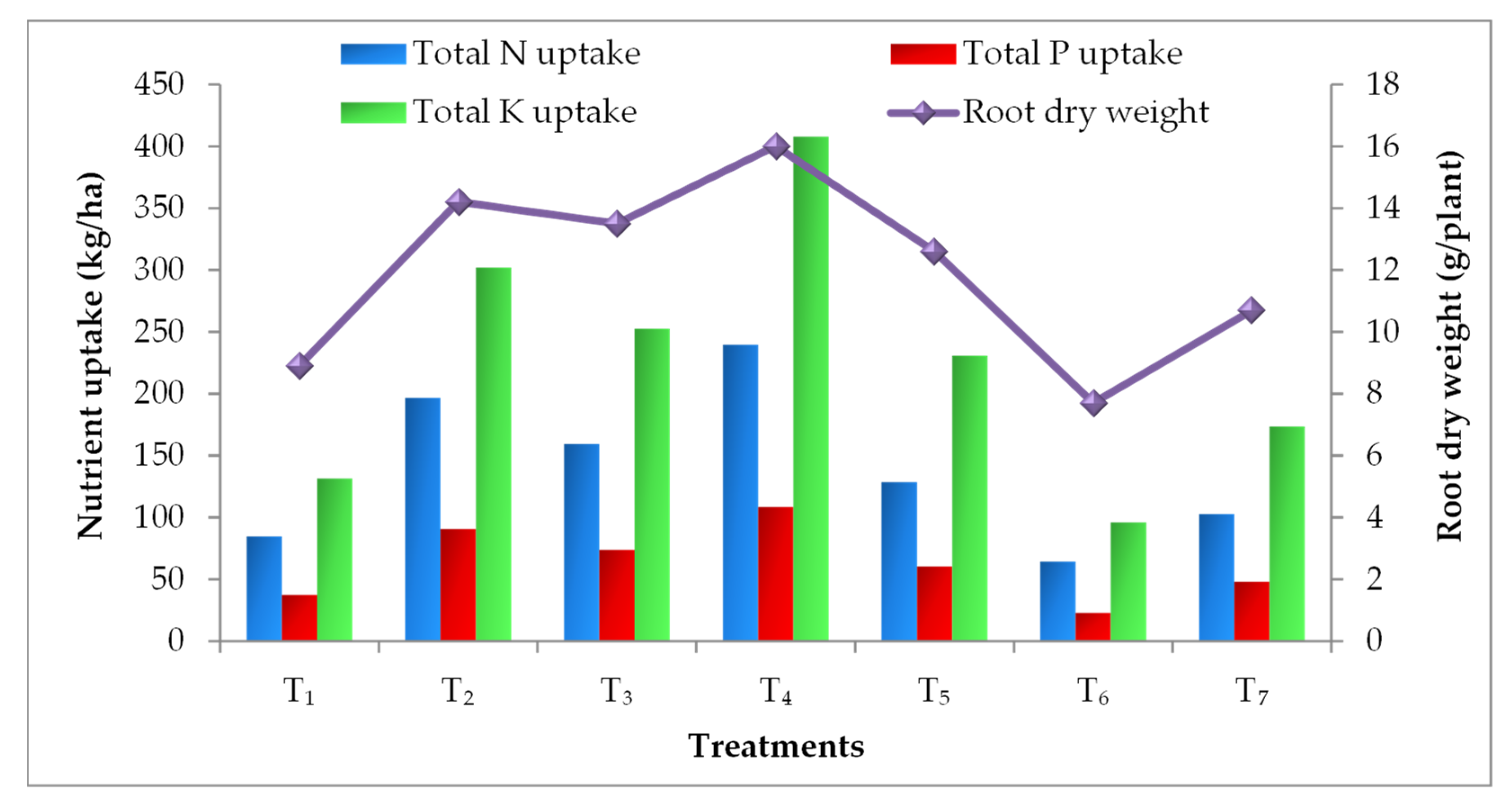
2.5. Root Dry Weight
2.6. Data Analysis
3. Results
3.1. Effect on Growth Indicating Parameters
3.2. Effect on Cumulative Radiation Interception (MJ/m2)
3.3. Effect on Phenological Phases
3.4. Effect on Nutrient Uptake
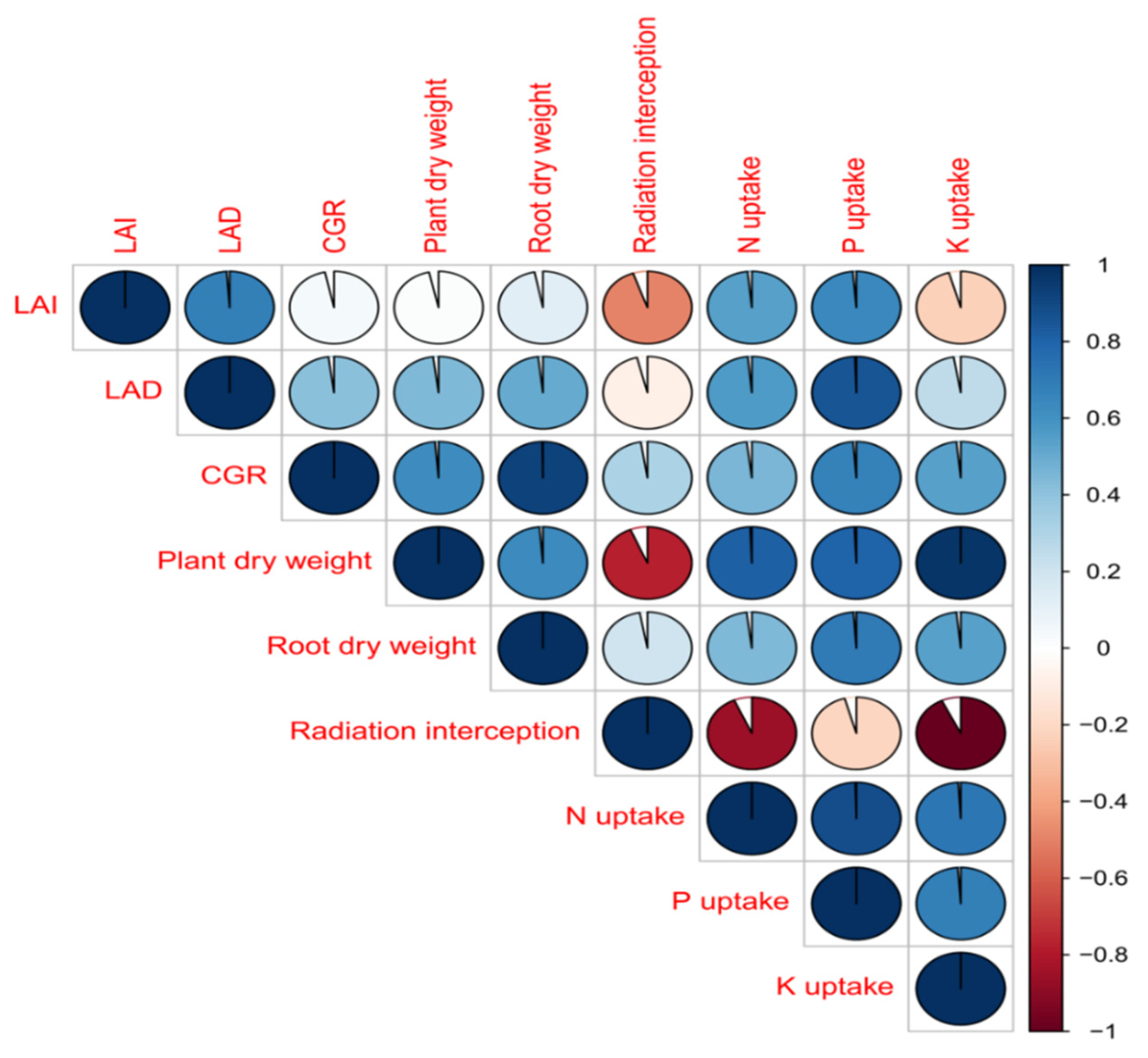
3.5. Correlation Studies
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC Climate Change. Synthesis Report: Contribution of working groups I. II and III to the Fifth Assessment Report. In Proceedings of the Intergovernmental Panel on Climate Change, Copenhagen, Denmark, 1 November 2014; pp. 1–167. [Google Scholar]
- IPCC Climate Change. The Physical Science Basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Reddy, A.R.; Rasineni, G.K.; Raghavendra, A.S. The impact of global elevated CO2 concentration on photosynthesis and plant productivity. Curr. Sci. 2010, 99, 46–57. [Google Scholar]
- Van der Kooi, C.J.; Reich, M.; Low, M.; De Kok, L.J.; Tausz, M. Growth and yield stimulation under elevated CO2 and drought: A meta-analysis on crops. Environ. Exp. Biol. 2016, 122, 150–157. [Google Scholar] [CrossRef]
- Taub, D. Effects of rising atmospheric concentrations of carbon dioxide on plants. Nat. Educ. Knowl. 2010, 3, 21. [Google Scholar]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising (CO2): Mechanisms and environmental interactions. Plant. Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Mamatha, H.; Srinivasa Rao, N.K.; Laxman, R.H.; Shivashankara, K.S.; Bhatt, R.M.; Pavithra, K.C. Impact of elevated CO2 on growth, physiology, yield, and quality of tomato (Lycopersicon esculentum Mill) cv. Arka Ashish. Photosynthetica 2014, 52, 519–528. [Google Scholar] [CrossRef]
- Pan, T.; Ding, J.; Qin, G.; Wang, Y.; Xi, L.; Yang, J.; Li, J.; Zhang, J.; Zou, Z. Interaction of supplementary light and CO2 enrichment improves growth, photosynthesis, yield, and quality of tomato in autumn through spring greenhouse production. HortScience 2019, 54, 246–252. [Google Scholar] [CrossRef] [Green Version]
- Soltani, A.; Gholipoor, M.; Ghassemi-Golezani, K. Analysis of temperature and atmospheric CO2 effects on radiation use efficiency in chickpea (Cicer arietinum L.). J. Plant. Sci. 2007, 2, 89–95. [Google Scholar] [CrossRef]
- Weerakoon, W.M.W.; Ingram, K.T.; Moss, D.N. Atmospheric carbon dioxide and fertilizer nitrogen effects on radiation interception by rice. Plant. Soil 2020, 2000, 99–106. Available online: https://www.jstor.org/stable/42950704 (accessed on 9 October 2021).
- Hangs, R.D.; Van Rees, K.C.J.; Schoenau, J.J.; Guo, X. A simple technique for estimating above-ground biomass in short-rotation willow plantations. Biomass Bioenergy 2011, 35, 2156–2162. [Google Scholar] [CrossRef]
- Hikosaka, K. Leaf canopy as a dynamic system: Ecophysiology and optimality in leaf turnover. Ann. Bot. 2005, 95, 521–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.; Yin, X.; He, S.; Jiang, W.; Si, C.; Struik, P.C.; Luo, W.I.; Li, G.; Xie, Y.; Xiong, Y.; et al. Responses of wheat and rice to factorial combinations of ambient and elevated CO2 and temperature in FACE experiments. Glob. Chang. Biol. 2016, 22, 856–874. [Google Scholar] [CrossRef]
- Craufurd, P.Q.; Wheeler, T.R. Climate change and the flowering time of annual crops. J. Exp. Bot. 2009, 60, 2529–2539. [Google Scholar] [CrossRef] [Green Version]
- Abebe, A.; Pathak, H.; Singh, S.D.; Bhatia, A.; Harit, R.C.; Kumar, V. Growth, yield and quality of maize with elevated atmospheric carbon dioxide and temperature in north–west India. Agric. Ecosyst. Environ. 2016, 218, 66–72. [Google Scholar] [CrossRef]
- Haque, M.S.; Karimi, M.A.; Haque, M.M.; Hamid, A.; Nawata, E. Effect of elevated CO2 concentration on growth, chlorophyll content and yield of mungbean (Vigna radiata L. Wilczek) genotypes. Jpn. J. Trop. Agr. 2005, 49, 189–196. [Google Scholar]
- Tian, Y.; Zheng, C.; Chen, J. Climate warming increases winter wheat yield but reduces grain nitrogen concentration in east China. PLoS ONE 2014, 9, e95108. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.; Li, G.; Yang, H.; Yang, J.; Liu, H.; Struik, P.C.; Luo, W.; Yin, X.; Di, L.; Guo, X.; et al. Do all leaf photosynthesis parameters of rice acclimate to elevated CO2, elevated temperature, and their combination, in FACE environments? Glob. Chang. Biol. 2018, 24, 1685–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johkan, M.; Oda, M.; Maruo, T.; Shinohara, Y. Crop Production and Global Warming Impacts, Case Studies on the Economy, Human Health, and on Urban and Natural Environments. 2011. Available online: http://www.intechopen.com (accessed on 9 October 2021).
- Dusenge, M.E.; Duarte, A.G.; Danielle, A. Way Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Zhuang, Q.; Ciais, P.; Welp, L.; Li, W.; Xin, Q. Elevated atmospheric CO2 negatively impacts photosynthesis through radiative forcing and physiology-mediated climate feedback. Geophysic. Res. Lett. 2017, 44, 1956–1963. [Google Scholar] [CrossRef] [Green Version]
- Raj, M.; Chakrabarti, B.; Pathak, H.; Singh, S.D.; Mina, M.; Purakayastha, T.J. Growth, yield and nutrient uptake in crop grown under elevated carbon dioxide and different doses of nitrogen fertilizer. Indian J. Experiment. Biol. 2019, 57, 181–187. [Google Scholar]
- Veneklaas, E.J.; Lambers, H.; Bragg, J.; Finnegan, P.M.; Lovelock, C.E.; Plaxton, W.C.; Price, C.A.; Scheible, W.; Shane, M.W.; White, P.J.; et al. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef]
- Taub, D.R.; Wang, X.Z. Why are nitrogen concentrations in plant tissues lower under elevated CO2? A critical examination of the hypotheses. J. Integr. Plant. Biol. 2008, 50, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Navas, M.L. Plant growth and competition at elevated CO2: On winners, losers and functional groups. New Phytol. 2003, 157, 175–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, P.; Roy, K.S.; Dash, P.K.; Neogi, S.; Shahid, M.D.; Nayak, A.K.; Raja, R.; Karthikeyan, S.; Balachandar, D.; Rao, K.S. Effect of elevated carbon dioxide and temperature on phosphorus uptake in tropical flooded rice (Oryza sativa L.). Eur. J. Agron. 2014, 53, 28–37. [Google Scholar] [CrossRef]
- Seneweera, S. Effects of elevated CO2 on plant growth and nutrient partitioning of rice (Oryza sativa L.) at rapid tillering and physiological maturity. J. Plant. Interac. 2011, 6, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Moretti, C.L.; Mattos, L.M.; Calbo, A.G.; Sargent, S.A. Climate changes and potential impacts on postharvest quality of fruit and vegetable crops—A review. Food Res. Int. 2010, 43, 1824–1832. [Google Scholar] [CrossRef]
- Bertin, N. Analysis of the Tomato Fruit Growth Response to Temperature and Plant Fruit Load in Relation to Cell Division, Cell Expansion and DNA Endoreduplication. Ann. Bot. 2005, 95, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Ploeg, A.V.D.; Heuvelink, E. Influence of sub-optimal temperature on tomato growth and yield: A review. J. Horticul. Sci. Biotechnol. 2005, 80, 652–659. [Google Scholar] [CrossRef]
- Watson, S.J. The physiological basis for variation in yield. Adv. Agron. 1952, 4, 101–145. [Google Scholar]
- Power, J.E.; Willis, W.C.; Igrones, D.L.; Richmon, B.A. Effect of soil temperature, phosphorus and plant age on growth analysis in barley. Agron. J. 1967, 59, 231–234. [Google Scholar] [CrossRef]
- Singh, P. Influence of water deficits on phenology growth and dry matter allocation in chickpea. Field Crops Res. 1991, 28, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Charles-Edwards, D.A.; Lawn, R.J. Light interception by grain legume row crops. Plant. Cell Environ. 1984, 7, 247–251. [Google Scholar] [CrossRef]
- Meek, D.W.; Hatfield, J.L.; Howell, T.A.; Idso, S.B.; Reginto, R.J. A generalized relationship between photosynthetically active radiation and solar radiation. Agron. J. 1984, 76, 939–945. [Google Scholar] [CrossRef]
- Monteith, J.L. Light distribution and photosynthesis in field crops. Ann. Bot. 1965, 29, 18. Available online: https://www.jstor.org/stable/42908627 (accessed on 9 October 2021). [CrossRef]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall of India Pvt. Ltd.: New Delhi, India, 1973; p. 498. [Google Scholar]
- Drake, B.G.; Gonzàlez-Meler, M.A.; Long, S.P. More efficient plants: A consequence of rising atmospheric CO2? Ann. Rev. Plant. Physiol. Mol. Biol. 1997, 48, 609–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, S.G.; Rogers, H.H.; Prior, S.A.; Peterson, C.M. Elevated CO2 and plant structure: A review. Glob. Chang. Biol. 1999, 5, 807–837. [Google Scholar] [CrossRef] [Green Version]
- Teawkul, P.; Chen, W.; Sripontan, Y.; Hwang, S. Elevated CO2 concentration promotes tomato plant growth but impairs Spodoptera litura performance. Res. Rev. J. Zool. Sci. 2015, 3, 35–42. [Google Scholar]
- Manderscheid, R.; Burkart, S.; Bramm, A.; Weigel, H. Effect of CO2 enrichment on growth and daily radiation use efficiency of wheat in relation to temperature and growth stage. Eur. J. Agron. 2003, 19, 411–425. [Google Scholar] [CrossRef]
- Mina, U.; Kumar, R.; Gogoi, R.; Bhatia, A.; Harit, R.C.; Singh, D.; Kumar, A.; Kumar, A. Effect of elevated temperature and carbon dioxide on maize genotypes health index. Ecol. Indic. 2019, 105, 292–302. [Google Scholar] [CrossRef]
- Vu, J.C.V.; Allen, L.H.; Gesch, R.W. Up-regulation of photosynthesis and sucrose metabolism enzymes in young expanding leaves of sugarcane under elevated growth CO2. Plant. Sci. 2006, 171, 123–131. [Google Scholar] [CrossRef]
- Khanboluki, G.; Hosseini, H.M.; Holford, P.; Zadeh, B.M.; Milham, P.J. Effect of elevated atmospheric CO2 concentration on growth and physiology of wheat and sorghum under cadmium stress. Commun. Soil Sci. Plant. Anal. 2018, 49, 2867–2882. [Google Scholar] [CrossRef]
- Yadav, A.; Bhatia, A.; Yadav, S.; Kumar, V.; Singh, B. The effects of elevated CO2 and elevated O3 exposure on plant growth, yield and quality of grains of two wheat cultivars grown in north India. Heliyon 2019, 5, e02317. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, S.; Jellings, A.; Fuller, M. Positive effects of elevated CO2 and its interaction with nitrogen on safflower physiology and growth. Agron. Sustain. Develop. 2013, 33, 497–505. [Google Scholar] [CrossRef] [Green Version]
- Bray, S.; Reid, D.M. The effect of salinity and CO2 enrichment on the growth and anatomy of the second trifoliate leaf of Phaseolus vulgaris. Can. J. Bot. 2002, 80, 349–359. [Google Scholar] [CrossRef]
- Nasser, R.R.; Fuller, M.P.; Jellings, A.J. Effect of elevated CO2 and nitrogen levels on lentil growth and nodulation. Agron. Sustain. Dev. 2007, 28, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Dubey, S.K.; Tripathi, S.K.; Pranuthi, G. Effect of elevated CO2 on wheat crop: Mechanism and impact. Crit. Rev. Environ. Sci. Technol. 2015, 45, 2283–2304. [Google Scholar] [CrossRef]
- Jin, J.; Li, Y.; Liu, X.; Wang, G.; Tang, C.; Yu, Z.; Wang, X.; Herbert, S.J. Elevated CO2 alters distribution of nodal leaf area and enhances nitrogen uptake contributing to yield increase of soybean cultivars grown in Mollisols. PLoS ONE 2017, 12, e0176688. [Google Scholar] [CrossRef] [Green Version]
- Vanaja, M.; Jyothi, M.; Ratnakumar, P.; Vagheera, P.; Raghuram Reddy, P.; Jyothi Lakshmi, N.; Yadav, S.K.; Maheshwari, M.; Venkateswarlu, B. Growth and yield responses of castor bean (Ricinus communis L.) to two enhanced CO2 levels. Plant. Soil Environ. 2008, 54, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Usuda, H. Effects of elevated CO2 on the capacity for photosynthesis of a single leaf and a whole plant, and on growth in a Radish. Plant. Cell Physiol. 2006, 47, 262–269. [Google Scholar] [CrossRef]
- Aien, A.; Pal, M.; Khetarpal, S.; Pandey, S.K. Impact of elevated atmospheric CO2 concentration on the growth, and yield in two potato cultivars. J. Agri. Sci. Tech. 2014, 16, 1661–1670. Available online: http://jast.modares.ac.ir/article-23-4377-en.html (accessed on 9 October 2021).
- Sujatha, K.B. Characterization of the Response of Rice Cultivars (Oryza sativa L.) to the Interaction of Elevated CO2 and Temperature; Dissertation, Indian Agricultural Research Institute: New Delhi, India, 2005. [Google Scholar]
- Kemanian, A.R.; Stöckle, C.O.; Huggins, D.R. Variability of barley radiation-use efficiency. Crop Sci. 2004, 44, 1662–1672. [Google Scholar] [CrossRef]
- Liu, W.; Yan, J.; Li, J.; Sang, T. Yield potential of Miscanthus energy crops in the Loess Plateau of China. GCB Bioenergy 2012, 4, 545–554. [Google Scholar] [CrossRef]
- Manoj, K.N.; Umesh, M.R.; Ramesh, Y.M.; Anand, S.R.; Angadi, S. Dry matter production and radiation use efficiency of pulses grown under different light conditions. Bangladesh J. Bot. 2019, 48, 9–15. [Google Scholar] [CrossRef]
- Schrader, S.M.; Wise, R.R.; Wacholtz, W.F.; Ort, D.R.; Sharkey, T.D. Thylakoid membrane responses to moderately high leaf temperature in Pima cotton. Plant. Cell Environ. 2004, 27, 725–735. [Google Scholar] [CrossRef]
- Wise, R.R.; Olson, A.J.; Schrader, S.M.; Sharkey, T.D. Electron transport is the functional limitation of photosynthesis in field-grown Pima cotton plants at high temperature. Plant. Cell Environ. 2004, 27, 717–724. [Google Scholar] [CrossRef]
- Morison, J.I.L.; Lawlor, D.W. Interactions between increasing CO2 concentration and temperature on plant growth. Plant. Cell Environ. 1999, 22, 659–682. [Google Scholar] [CrossRef] [Green Version]
- Taub, D.R.; Seemann, J.R.; Coleman, J.S. Growth in elevated CO2 protects photosynthesis against high-temperature damage. Plant. Cell Environ. 2000, 23, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Prasad, P.V.V.; Allen, L.H., Jr.; Boote, K.J. Crop Responses to Elevated Carbon Dioxide and Interaction with Temperature: Grain Legumes. Journal of Crop Improvement, and: Ecological Responses and Adaptations of Crops to Rising Atmospheric Carbon Dioxide (ed: Zoltán Tuba); Food Products Press, an imprint of The Haworth Press, Inc.: Binghamton, NY, USA, 2005; pp. 113–154. [Google Scholar] [CrossRef]
- Dwivedi, S.K.; Kumar, S.; Prakash, V.; Mondal, S.; Mishra, J.S. Influence of rising atmospheric CO2 concentrations and temperature on morpho-physiological traits and yield of rice genotypes in sub-humid climate of Eastern India. AJPS 2015, 6, 2339–2349. [Google Scholar]
- Prasad, P.V.V.; Boote, K.J.; Allen, L.H., Jr.; Thomas, J.M.G. Super-optimal temperatures are detrimental to peanut (Arachis hypogea L.) reproductive processes and yield at both ambient and elevated carbon dioxide. Glob. Chang. Biol. 2003, 9, 1775–1787. [Google Scholar] [CrossRef] [Green Version]
- Kimball, B.A.; Kobayashi, K.; Bindi, M. Responses of agricultural crops to Free-Air CO2 Enrichment. Adv. Agron. 2002, 77, 293–368. [Google Scholar]
- Bahuguna, R.N.; Jagadish, S.V.K. Temperature regulation of plant phenological development. Environ. Exp. Bot. 2015, 111, 83–90. [Google Scholar] [CrossRef]
- Sanchez, B.; Rasmussen, A.; Porter, J. Temperatures and the growth and development of maize and rice: A review. Glob. Chang. Biol. 2014, 20, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Sehgal, V.K.; Chakraborty, D.; Pal, M. Atmospheric carbon dioxide enrichment induced modifications in canopy radiation utilization, growth and yield of chickpea Cicer arietinum L. Agric. Meteorol. 2015, 202, 102–111. [Google Scholar] [CrossRef]
- Singh, R.N.; Mukherjee, J.; Sehgal, V.K.; Krishnan, P.; Das, D.K.; Dhakar, R.K.; Bhatia, A. Interactive effect of elevated tropospheric ozone and carbon dioxide on radiation utilisation, growth and yield of chickpea (Cicer arietinum L.). Int. J. Biometeorol. 2021, 65, 1939–1952. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Su, H.; Liu, W.; Tan, K.; Ren, S. Infrared warming reduced winter wheat yields and some physiological parameters, which were mitigated by irrigation and worsened by delayed sowing. PLoS ONE 2013, 8, e67518. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Chen, J.; Zhang, B.; Tian, Y.; Zhang, W. Responses of biomass growth and grain yield of midseason rice to the anticipated warming with FATI facility in East China. Field Crops Res. 2011, 123, 259–265. [Google Scholar] [CrossRef]
- Wang, X.; Liu, F. Effects of Elevated CO2 and Heat on Wheat Grain Quality. Plants 2021, 10, 1027. [Google Scholar] [CrossRef] [PubMed]
- Pendall, E.; Bridgham, S.; Hanson, P.J.; Hungate, B.; Kicklighter, D.W.; Johnson, D.W.; Law, B.E.; Luo, Y.; Megonigal, J.P.; Olsrud, M.; et al. Below-ground process responses to elevated CO2 and temperature: A discussion of observations, measurement methods and models. New Phytol. 2004, 162, 311–322. [Google Scholar] [CrossRef]
- Satapathy, S.S.; Swain, D.K.; Pasupalak, S.; Bhadoria, P.B.S. Effect of elevated [CO2] and nutrient management on wet and dry season rice production in subtropical India. Crop J. 2015, 3, 468–480. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.Y.; Lieffering, M.; Miura, S.; Kobayashi, K.; Okada, M. Growth and nitrogen uptake of CO2-enriched rice under field conditions. New Phytol. 2001, 150, 223–229. [Google Scholar] [CrossRef]
- Carvalho, J.M.; Barreto, R.F.; Prado, R.D.M.; Habermann, E.; Branco, R.B.F.; Martinez, C.A. Elevated CO2 and warming change the nutrient status and use efficiency of Panicum maximum Jacq. PLoS ONE 2020, 15, e0223937. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Tang, C.; Sale, P. The impact of elevated carbon dioxide on the phosphorus nutrition of plants: A review. Ann. Bot. 2015, 116, 987–999. [Google Scholar] [CrossRef] [Green Version]
- Nie, M.; Lu, M.; Bell, J.; Raut, S.; Pendall, E. Altered root traits due to elevated CO2: A meta-analysis. Glob. Ecol. Biogeogr. 2013, 22, 1095–1105. [Google Scholar] [CrossRef]
- Dong, J.; Hunt, J.; Delhaize, E.; Zheng, S.J.; Jin, C.W.; Tang, C. Impacts of elevated CO2 on plant resistance to nutrient deficiency and toxic ions via root exudates: A review. Sci. Total Environ. 2021, 754, 142434. [Google Scholar] [CrossRef] [PubMed]
- Drigo, B.; Van Veen, J.A.; Kowalchuk, G.A. Specific rhizosphere bacterial and fungal groups respond differently to elevated atmospheric CO2. ISME J. 2009, 3, 1204–1217. [Google Scholar] [CrossRef] [PubMed]
- Krey, T.; Vassilev, N.; Baum, C.; Lobermann, B.E. Effects of long-term phosphorus application and plant-growth promoting rhizobacteria on maize phosphorus nutrition under field conditions. Eur. J. Soil Biol. 2013, 55, 124–130. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; Huang, J.; Zhu, J.; Yang, H.; Liu, G.; Liu, H.; Dong, G.; Hu, J. Seasonal changes in the effects of free-air CO2 enrichment (FACE) on phosphorus uptake and utilization of rice at three levels of nitrogen fertilization. Field Crops Res. 2007, 102, 141–150. [Google Scholar] [CrossRef]
- Roy, K.S.; Bhattacharyya, P.; Neogi, S.; Rao, K.S.; Adhya, T.K. Combined effect of elevated CO2 and temperature on dry matter production, net assimilation rate, C and N allocations in tropical rice (Oryza sativa L.). Field Crops Res. 2012, 139, 71–79. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment No. | Treatment | Description |
|---|---|---|
| T1 | C0T0 | Ambient CO2 and ambient temperature at OTC |
| T2 | C1T0 | Elevated CO2 (550 ± 20 ppm) and ambient temperature |
| T3 | C1T1 | Elevated CO2 (550 ± 20 ppm) and elevated temperature of +2 °C |
| T4 | C2T0 | Elevated CO2 (700 ± 20 ppm) and ambient temperature |
| T5 | C2T1 | Elevated CO2 (700 ± 20 ppm) and elevated temperature +2 °C |
| T6 | C0T1 | Ambient CO2 and elevated temperature +2 °C |
| T7 | C0T0 | Ambient CO2 and ambient temperature at open field |
| Description | Average CO2 Concentration (ppm) | Mean Temperature (°C) |
|---|---|---|
| Ambient CO2 and ambient temperature at OTC | - | 25.8 (1.1) |
| Elevated CO2 (550 ± 20 ppm) and ambient temperature | 554 (29) | - |
| Elevated CO2 (550 ± 20 ppm) and elevated temperature of +2 °C | 552 (33) | 27.2 (1.1) |
| Elevated CO2 (700 ± 20 ppm) and ambient temperature | 701 (32) | - |
| Elevated CO2 (700 ± 20 ppm) and elevated temperature +2 °C | 702 (36) | 27.4 (1.4) |
| Ambient CO2 and elevated temperature +2 °C | - | 26.8 (1.1) |
| Ambient CO2 and ambient temperature at Open field | 414 (25) | 24.9 (1.1) |
| Actual Weather | ||
| Tmean | 24.6 (0.6) | |
| Tmaximum (°C) | 30.7 (1.3) | |
| Tminimum (°C) | 17.6 (1.5) | |
| RHmean (%) | 81.8 (5) | |
| Mean sunshine hours | 5.5 (2.1) | |
| Total Rainfall (mm) | 940.5 (197) |
| Leaf Area (cm2/Plant) | LAI | |||||
|---|---|---|---|---|---|---|
| Treatments | 50% Flowering | Peak Fruiting | at Harvest | 50% Flowering | Peak Fruiting | at Harvest |
| (Mean ± SE) | ||||||
| T1 | 3943.44 ± 60.23 c | 5382.56 ± 82.221 e | 1184.46 ± 18.090 c, d | 0.49 ± 0.009 c | 0.66 ± 0.012 d | 0.15 ± 0.003 c |
| T2 | 4431.26 ± 92.236 b | 8660.76 ± 180.280 b | 4193.75 ± 87.299 a | 0.55 ± 0.012 b | 1.07 ± 0.021 a | 0.52 ± 0.012 a |
| T3 | 4193.89 ± 87.303 b, c | 7488.56 ± 155.887 c | 3524.27 ± 73.358 b | 0.52 ± 0.012 b, c | 0.92 ± 0.018 b | 0.44 ± 0.009 b |
| T4 | 4829.93 ± 73.774 a | 9110.68 ± 139.178 a | 4201.54 ± 64.192 a | 0.60 ± 0.009 a | 1.12 ± 0.019 a | 0.52 ± 0.007 a |
| T5 | 4167.54 ± 104.891 b, c | 6764.23 ± 170.229 d | 1274.69 ± 32.076 c | 0.51 ± 0.012 b, c | 0.84 ± 0.022 c | 0.16 ± 0.003 c |
| T6 | 3572.29 ± 94.510 d | 4149.21 ± 109.772 f | 1112.68 ± 29.438 d | 0.44 ± 0.012 d | 0.51 ± 0.012 e | 0.14 ± 0.003 c |
| T7 | 4002.58 ± 105.898 c | 6398.35 ± 169.293 d | 1231.72 ± 32.591 c, d | 0.49 ± 0.012 c | 0.79 ± 0.021 c | 0.15 ± 0.006 c |
| SEM± | 87.97 | 145.32 | 51.95 | 0.01 | 0.02 | 0.01 |
| LSD (p = 0.05) | 271.07 | 447.77 | 160.08 | 0.03 | 0.06 | 0.02 |
| LAD (Days) | CGR (g/m2/day) | |||||||
|---|---|---|---|---|---|---|---|---|
| Treatments | 0–50% Flowering | 50% Flowering-Peak Fruiting | Peak Fruiting-Harvest | Total LAD | 0–50% Flowering | 50% Flowering-Peak Fruiting | Peak Fruiting-Harvest | Average CGR |
| (Mean ± SE) | ||||||||
| T1 | 10.95 ± 0.165 c | 25.91 ± 0.394 e | 6.08 ± 0.092 d | 42.94 ± 0.657 e | 0.78 ± 0.012 f | 2.13 ± 0.034 e | 1.04 ± 0.015 c | 1.32 ± 0.019 f |
| T2 | 12.31 ± 0.255 b | 36.37 ± 0.756 b | 11.90 ± 0.247 a | 60.58 ± 1.261 b | 1.72 ± 0.035 b | 3.05 ± 0.066 b | 2.26 ± 0.045 a | 2.35 ± 0.048 b |
| T3 | 11.65 ± 0.244 b, c | 32.45 ± 0.675 c | 10.20 ± 0.211 b | 54.30 ± 1.130 c | 1.33 ± 0.027 c | 2.88 ± 0.062 b | 2.24 ± 0.047 a | 2.15 ± 0.047 c |
| T4 | 13.42 ± 0.205 a | 38.72 ± 0.593 a | 12.33 ± 0.190 a | 64.47 ± 0.984 a | 1.85 ± 0.027 a | 3.87 ± 0.058 a | 2.32 ± 0.036 a | 2.68 ± 0.039 a |
| T5 | 11.58 ± 0.290 b, c | 30.37 ± 0.764 d | 7.44 ± 0.188 c | 49.39 ± 1.242 d | 1.12 ± 0.027 d | 2.62 ± 0.067 c | 2.21 ± 0.057 a | 1.99 ± 0.050 d |
| T6 | 9.92 ± 0.261 d | 21.45 ± 0.567 f | 4.87 ± 0.129 e | 36.24 ± 0.959 f | 0.46 ± 0.012 g | 1.66 ± 0.044 f | 1.38 ± 0.038 b | 1.17 ± 0.030 g |
| T7 | 11.12 ± 0.294 c | 28.89 ± 0.764 d | 7.06 ± 0.188 c | 47.07 ± 1.247 d | 1.01 ± 0.026 e | 2.37 ± 0.065 d | 1.46 ± 0.041 b | 1.61 ± 0.044 e |
| SEM± | 0.24 | 0.65 | 0.18 | 1.07 | 0.03 | 0.06 | 0.04 | 0.04 |
| LSD (p = 0.05) | 0.75 | 1.99 | 0.55 | 3.29 | 0.08 | 0.17 | 0.13 | 0.12 |
| Cumulative Radiation Interception (MJ/m2) | Days Taken for First Initiation | ||||||
|---|---|---|---|---|---|---|---|
| Treatments | 50% Flowering | Peak Fruiting | at Harvest | Branch Initiation | Flower Initiation | Fruit Initiation | Fruit Maturation |
| (Mean ± SE) | |||||||
| T1 | 122.75 ± 1.876 c | 333.20 ± 5.089 c, d | 410.91 ± 6.276 b | 23.37 ± 0.387 a, b | 38.00 ± 0.255 a, b | 48.60 ± 0.417 a | 75.20 ± 1.149 a, b |
| T2 | 141.03 ± 2.935 a, b | 365.40 ± 7.607 a | 481.49 ± 10.024 a | 23.00 ± 0.479 b | 36.95 ± 0.540 b | 47.67 ± 0.436 a, b, c | 74.00 ± 0.804 a, b, c |
| T3 | 139.24 ± 2.899 a, b | 355.40 ± 7.399 a, b | 466.71 ± 9.716 a | 22.80 ± 0.348 b, c | 35.93 ± 0.749 c, d | 46.13 ± 0.961 b, c, d | 72.33 ± 1.504 b, c, d |
| T4 | 143.61 ± 2.194 a | 365.77 ± 5.589 a | 479.41 ± 7.324 a | 22.77 ± 0.239 b, c | 35.80 ± 0.547 c, d | 45.47 ± 0.693 c, d | 70.80 ± 1.081 c, d |
| T5 | 137.10 ± 3.448 a, b | 353.07 ± 8.884 a, b, c | 455.96 ± 11.472 a | 21.63 ± 0.282 c | 35.06 ± 0.570 c | 45.20 ± 1.138 d | 69.40 ± 1.746 d |
| T6 | 121.12 ± 3.204 c | 316.40 ± 8.369 d | 393.76 ± 10.419 b | 23.33 ± 0.720 a, b | 37.33 ± 0.673 a, b, c | 48.13 ± 0.955 a, b | 74.33 ± 1.646 a, b, c |
| T7 | 134.54 ± 3.557 b | 342.35 ± 7.428 b, c | 416.18 ± 11.013 b | 24.63 ± 0.676 a | 39.20 ± 0.719 a | 49.27 ± 0.136 a | 77.00 ± 1.406 a |
| SEM± | 2.87 | 7.20 | 9.45 | 0.44 | 0.61 | 0.79 | 1.40 |
| LSD (p = 0.05) | 8.83 | 22.19 | 29.12 | 1.35 | 1.87 | 2.44 | 4.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rangaswamy, T.C.; Sridhara, S.; Manoj, K.N.; Gopakkali, P.; Ramesh, N.; Shokralla, S.; Zin El-Abedin, T.K.; Almutairi, K.F.; Elansary, H.O. Impact of Elevated CO2 and Temperature on Growth, Development and Nutrient Uptake of Tomato. Horticulturae 2021, 7, 509. https://doi.org/10.3390/horticulturae7110509
Rangaswamy TC, Sridhara S, Manoj KN, Gopakkali P, Ramesh N, Shokralla S, Zin El-Abedin TK, Almutairi KF, Elansary HO. Impact of Elevated CO2 and Temperature on Growth, Development and Nutrient Uptake of Tomato. Horticulturae. 2021; 7(11):509. https://doi.org/10.3390/horticulturae7110509
Chicago/Turabian StyleRangaswamy, Tejaswini C., Shankarappa Sridhara, Konapura Nagaraja Manoj, Pradeep Gopakkali, Nandini Ramesh, Shadi Shokralla, Tarek K. Zin El-Abedin, Khalid F. Almutairi, and Hosam O. Elansary. 2021. "Impact of Elevated CO2 and Temperature on Growth, Development and Nutrient Uptake of Tomato" Horticulturae 7, no. 11: 509. https://doi.org/10.3390/horticulturae7110509
APA StyleRangaswamy, T. C., Sridhara, S., Manoj, K. N., Gopakkali, P., Ramesh, N., Shokralla, S., Zin El-Abedin, T. K., Almutairi, K. F., & Elansary, H. O. (2021). Impact of Elevated CO2 and Temperature on Growth, Development and Nutrient Uptake of Tomato. Horticulturae, 7(11), 509. https://doi.org/10.3390/horticulturae7110509