
Mitigation of Drought Damages by Exogenous Chitosan and Yeast Extract with Modulating the Photosynthetic Pigments, Antioxidant Defense System and Improving the Productivity of Garlic Plants

 ,
, 
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site, Plant Materials, and Cultural Practices
- The plants (control) were irrigated eight times to simulate normal conditions of rainfall (100% irrigation).
- The plants were irrigated six times to simulate 75%, or moderate, drought.
- The plants received 75% irrigation and were sprayed with yeast 8 g/L.
- The plants received 75% irrigation and were sprayed with chitosan 300 mM.
- Some plants received 75% irrigation and were sprayed with yeast 8 g/L plus chitosan 300 mM.
- The plants received four irrigations to simulate 50%, or severe, drought.
- The plants received 50% irrigation and were sprayed with yeast 8 g/L.
- The plants received 50% irrigation and were sprayed with chitosan 300 mM.
- The plants received 50% irrigation and were sprayed with yeast 8 g/L plus chitosan 300 mM.
2.2. Morphological Characters
2.3. Physiological and Biochemical Studies
2.3.1. Determination of Chlorophyll A, B Concentration and RWC
2.3.2. Determination of Proline Content
2.3.3. Assay of Electrolyte Leakage (EL%)
2.3.4. Determination of Ascorbic Acid (AsA)
2.3.5. Assay of H2O2, O2− and MDA
2.3.6. Determination of Enzymes Activity
2.4. Yield Characteristics
2.5. Statistical Analysis
3. Results
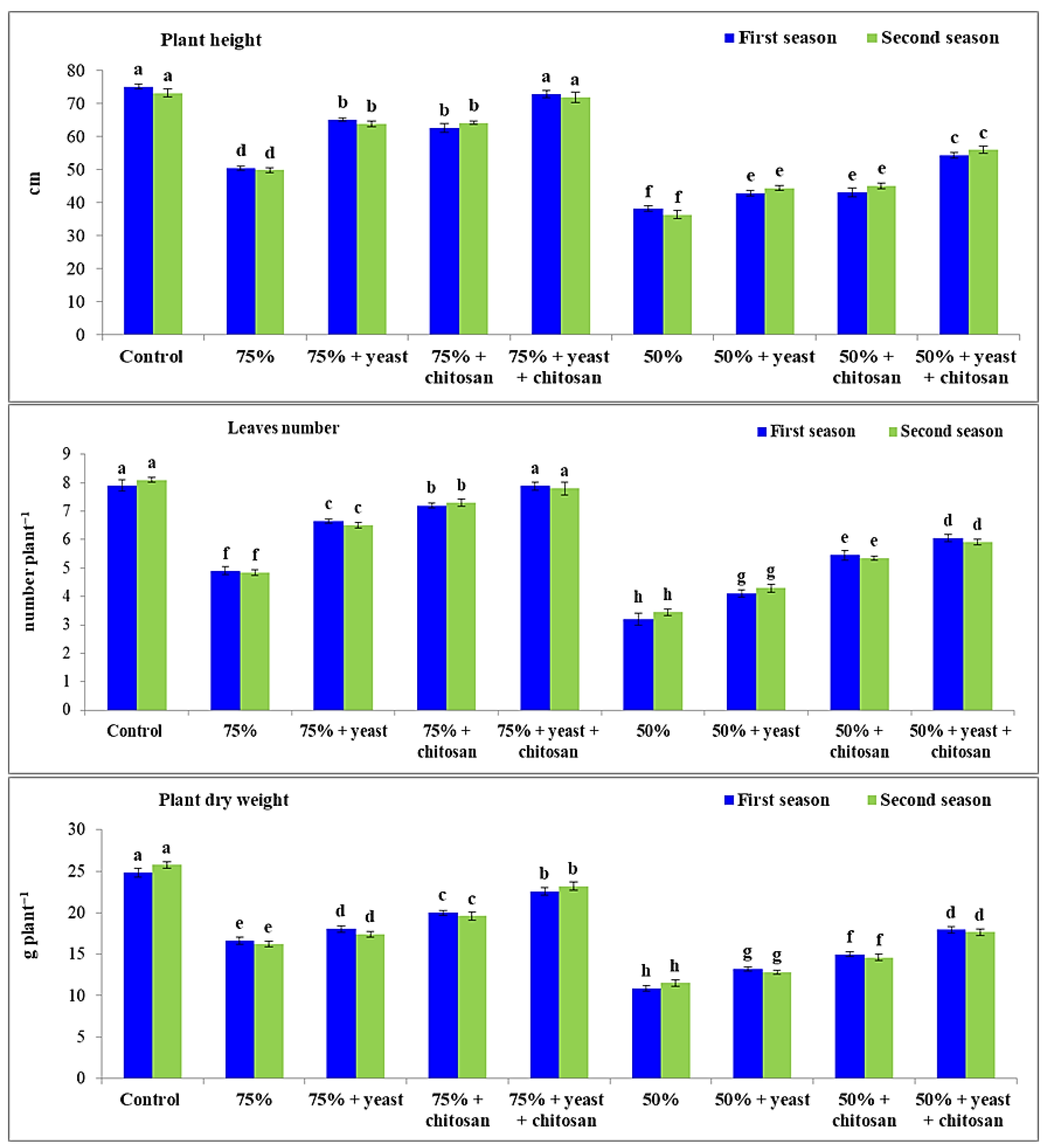
3.1. Effect of Yeast Extract and Chitosan on Plant Height, Number of Leaves per Plant, and Dry Weight of Plant for Garlic Plants in Drought Conditions
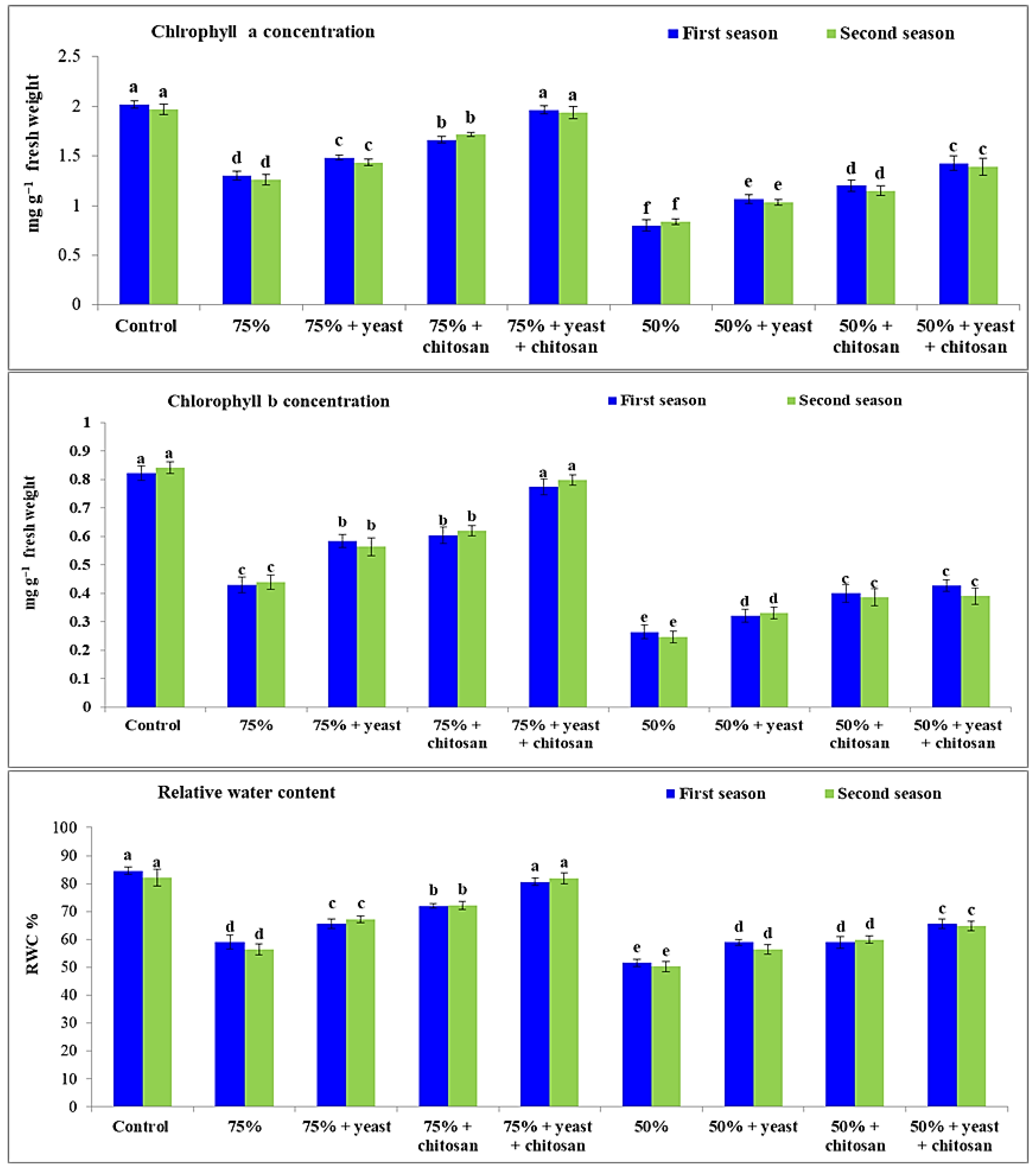
3.2. Effect of Yeast Extract or Chitosan on Concentrations of Chlorophyll A and B and on RWC in Stressed Garlic Plants
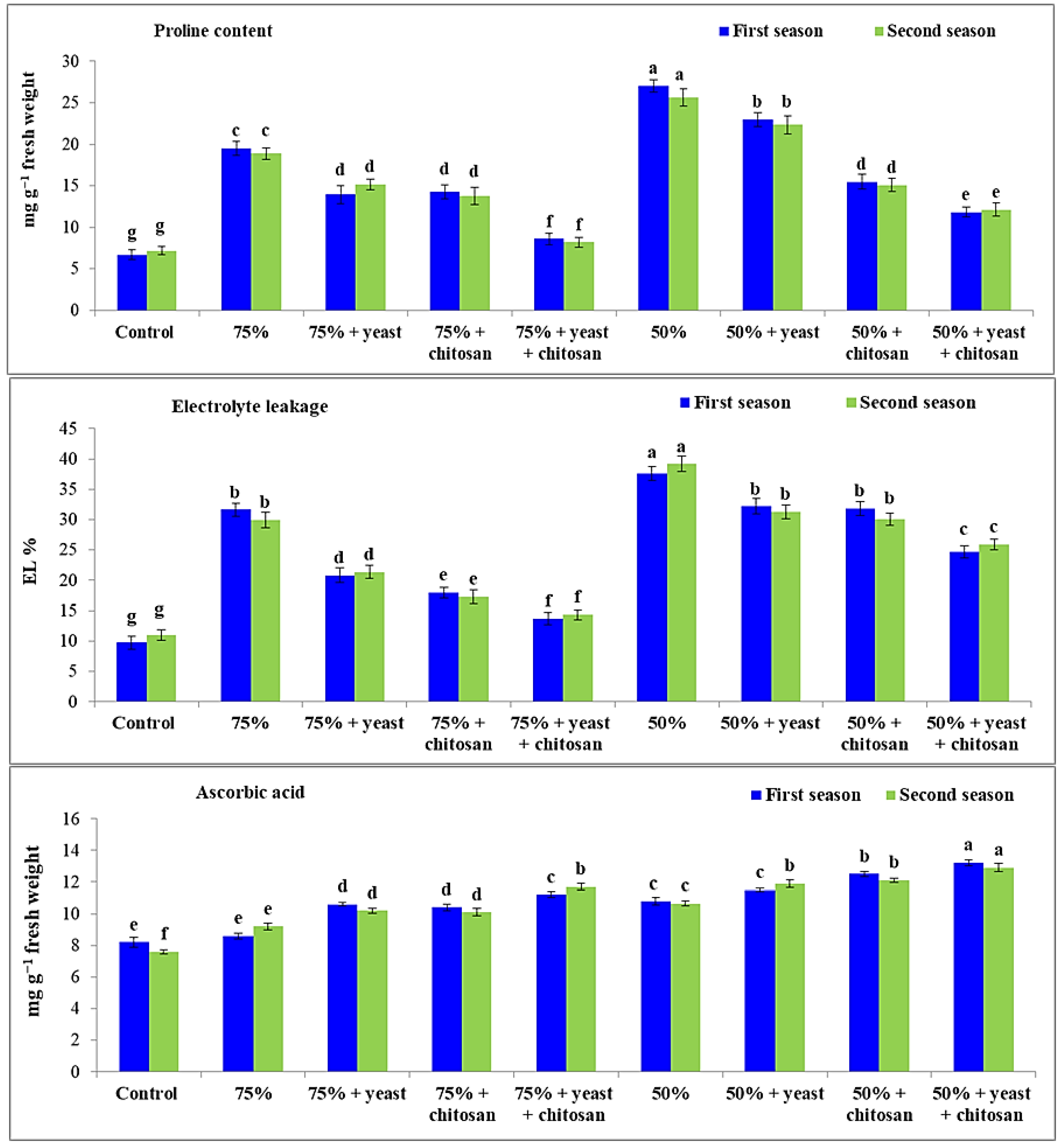
3.3. Effect of Yeast Extract or Chitosan on Proline Levels, Percentage of Electrolyte Leakage and Concentration of Ascorbic Acid in Garlic Plants in Drought Conditions
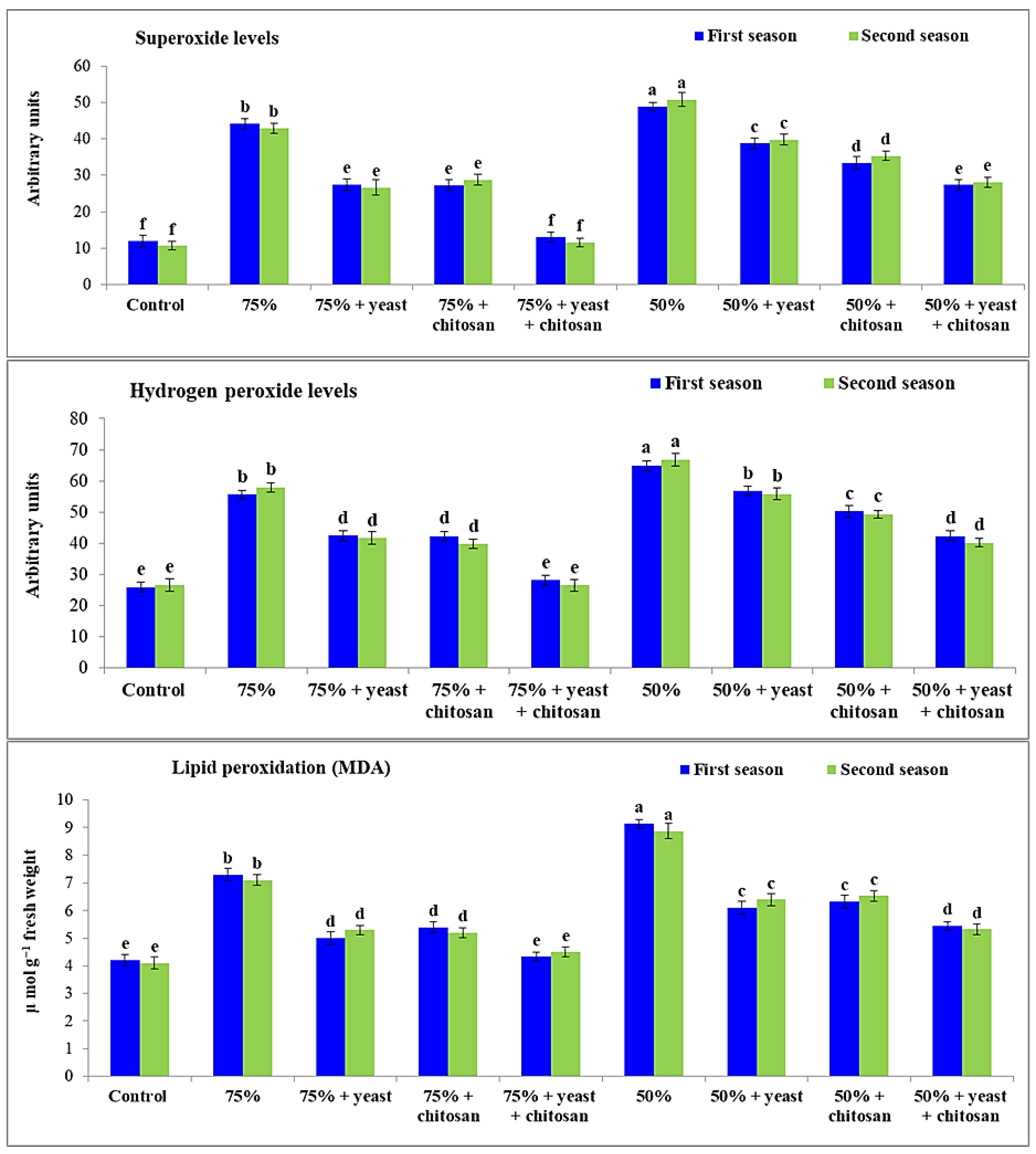
3.4. Effect of Yeast Extract or Chitosan on Levels of Hydrogen Peroxide, Superoxide, and MDA of Garlic Plants in Drought Conditions
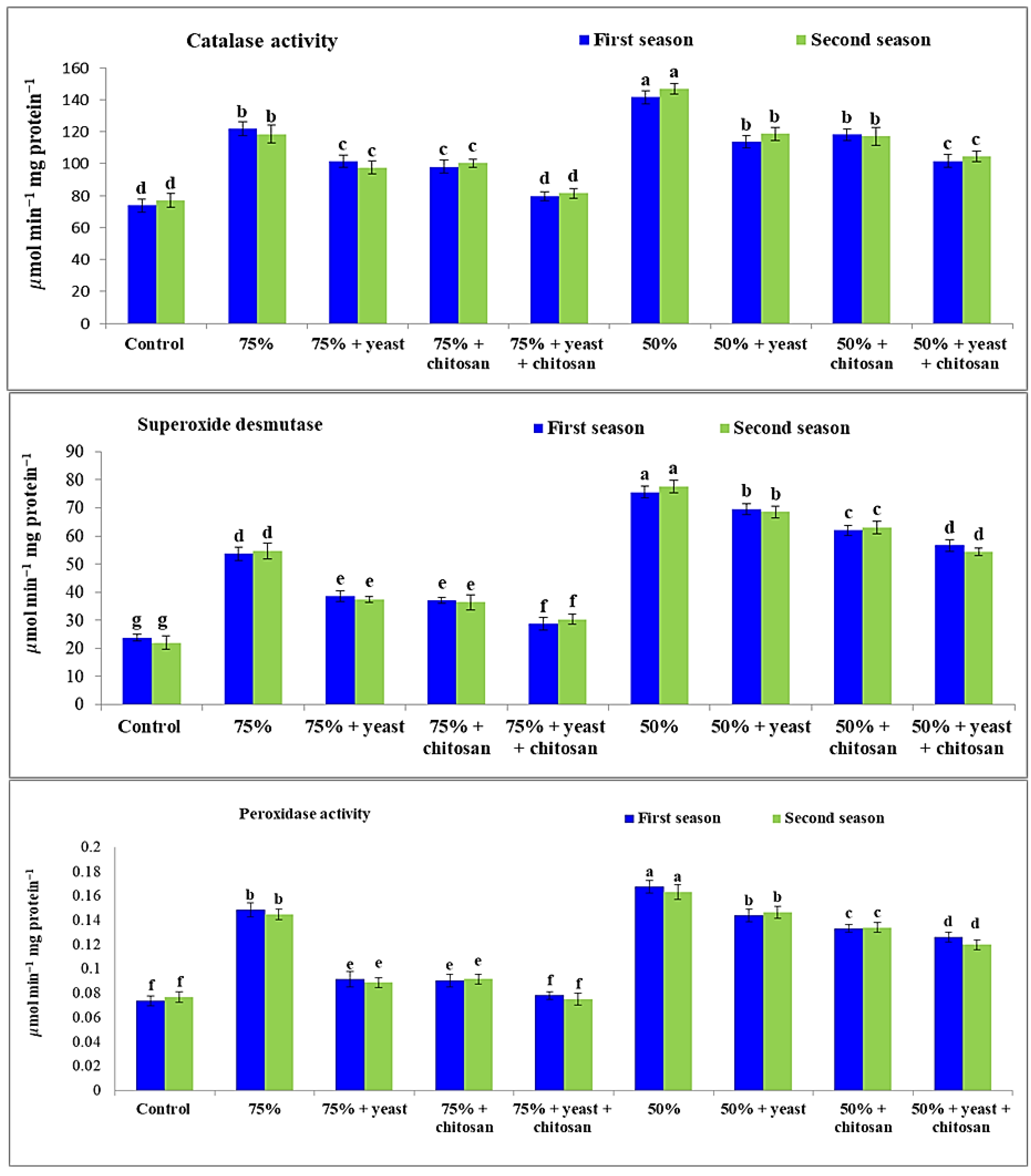
3.5. Effect of Yeast Extract or Chitosan on Catalase, Peroxidase, and Superoxide Dismutase Activity of Garlic in Drought Conditions
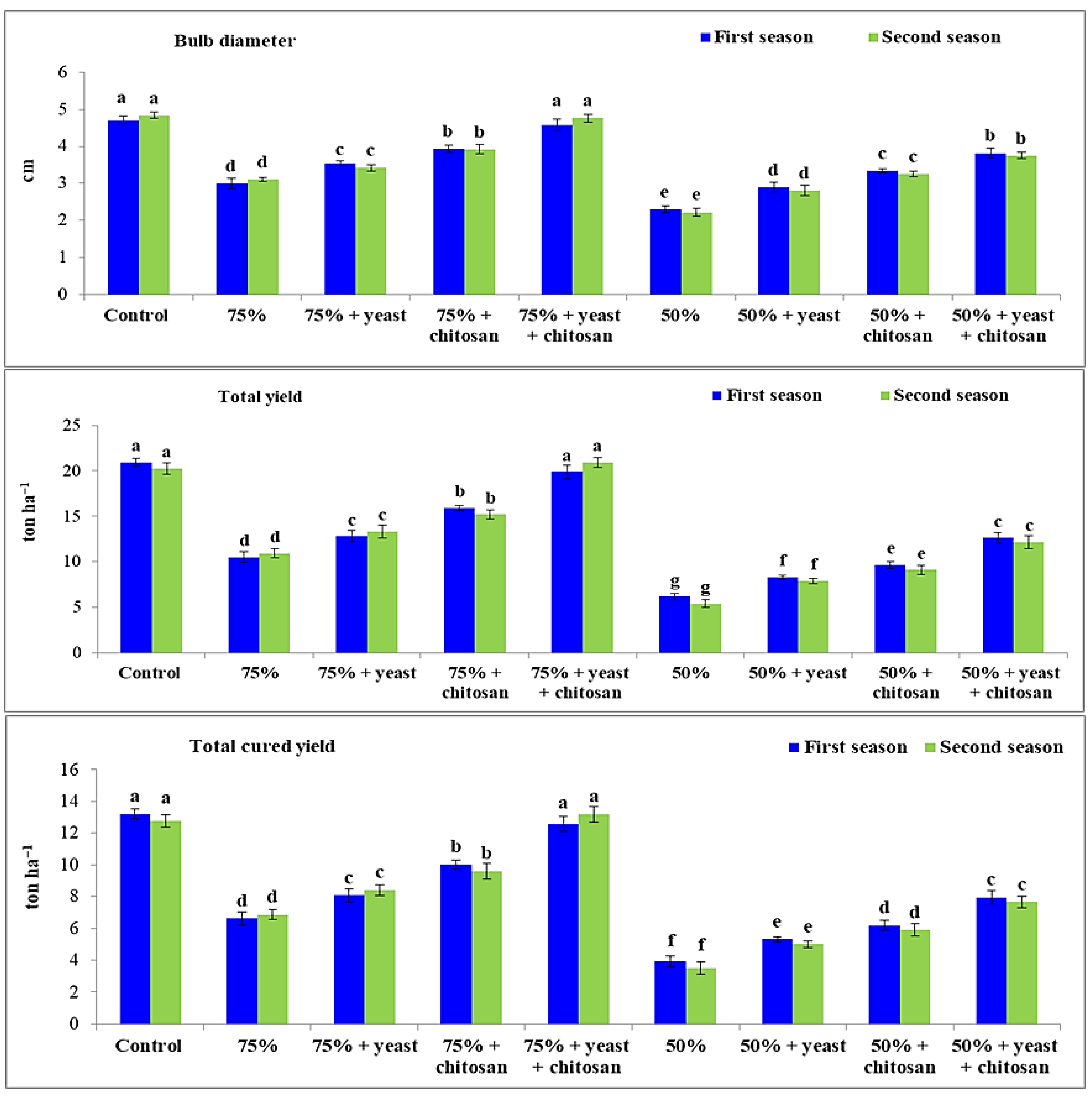
3.6. Effect of Yeast Extract or Chitosan on Bulb Diameter, Total Yield, and Total Cured Yield of Garlic Plants in Drought Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Retrieved December 2020 from the FAOSTAT on the World Wide; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018; Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 25 August 2021).
- Abdelrahman, M.; Hirata, S.; Mukae, T.; Yamada, T.; Sawada, Y.; El-Syaed, M.; Yutaka, Y.; Sato, M.; Hirai, M.; Shigyo, M. Comprehensive Metabolite Profiling in Genetic Resources of Garlic (Allium sativum L.) Collected from Different Geographical Regions. Molecules 2021, 26, 1415. [Google Scholar] [CrossRef]
- Shang, A.; Cao, S.Y.; Xu, X.Y.; Gan, R.Y.; Tang, G.Y.; Corke, H.; Li, H.B. Bioactive compounds and biological functions of garlic (Allium sativum L.). Foods 2019, 8, 246. [Google Scholar] [CrossRef]
- Ismail, M.S.M.; Ghallab, M.M.A.; Soliman, M.F.M.; AboGhalia, A.H. Acaricidal activities of some essential and fixed oils on the two-spotted spider mite, Tetranychus urticae. Egypt. Acad. J. Biol. Sci. 2011, 3, 41–48. [Google Scholar] [CrossRef]
- Sharaby, A.; Montasser, S.A.; Mahmoud, Y.A.; Ibrahim, S.A. Natural plant essential oils for controlling the grasshopper (Heteracris littoralis) and their pathological effects on the alimentary canal. Ecol. Balk. 2012, 4, 39–52. [Google Scholar]
- Omara, R.I.; El-Kot, G.A.; Fadel, F.M.; Abdelaal, K.A.; Saleh, E.M. Efficacy of Certain Bioagents on Patho-Physiological Characters of Wheat Plants under Wheat Leaf Rust Stress. Physiol. Mol. Plant Pathol. 2019, 106, 102–108. [Google Scholar] [CrossRef]
- Hafez, Y.M.; Abdelaal, K.A. Investigation of susceptibility and resistance mechanisms of some Egyptian wheat cultivars (Triticum aestivum L.) inoculated with Blumeria graminis f.sp. tritici using certain biochemical, molecular characterization and SEM. J. Plant Prot. Pathol. Mansoura Univ. 2015, 6, 431–454. [Google Scholar] [CrossRef][Green Version]
- Abdelaal, K.A.; Hafez, Y.M.; Badr, M.M.; Youseef, W.A.; Esmaeil, S.M. Biochemical, histological and molecular changes in susceptible and resistant wheat cultivars inoculated with stripe rust fungus Puccinia striiformis f. sp. tritici. Egypt J. Biol. Pest. Control 2019, 24, 421–429. [Google Scholar]
- Abdelaal, K.A.A.; Essawy, M.; Quraytam, A.; Abdallah, F.; Mostafa, H.; Shoueir, K.; Fouad, H.; Fahmy, A.S.H.; Hafez, Y.M. Toxicity of Essential Oils Nanoemulsion against Aphis Craccivora and Their Inhibitory Activity on Insect Enzymes. Processes 2021, 9, 624. [Google Scholar] [CrossRef]
- El-Flaah, R.F.; El-Said, R.A.R.; Nassar, M.A.; Hassan, M.; Abdelaal, K.A.A. Effect of rhizobium, nano silica and ascorbic acid on morpho-physiological characters and gene expression of POD and PPO in faba bean (Vicia faba L.) under salinity stress conditions. Fresenius Environ. Bull. 2021, 30, 5751–5764. [Google Scholar]
- Abdelaal, K.A.A.; EL-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.I.; El-Banna, M.; El-Esawi, M.; Elkelish, A. Treatment of Sweet Pepper with Stress Tolerance-Inducing Compounds Alleviates Salinity Stress Oxidative Damage by Mediating the Physio-Biochemical Activities and Antioxidant Systems. Agronomy 2020, 10, 26. [Google Scholar] [CrossRef]
- El-Banna, M.F.; Abdelaal, K.A.A. Response of Strawberry Plants Grown in the Hydroponic System to Pretreatment with H2O2 Before Exposure to Salinity Stress. J. Plant Prod. Mansoura Univ. 2018, 9, 989–1001. [Google Scholar] [CrossRef]
- Zhou, X.; Condori-Apfata, J.A.; Liu, X.; Condori-Pacsi, S.J.; Valencia, M.V.; Zhang, C. Transcriptomic Changes Induced by Drought Stress in Hardneck Garlic during the Bolting/Bulbing Stage. Agronomy 2021, 11, 246. [Google Scholar] [CrossRef]
- El-Shawa, G.M.R.; Rashwan, E.M.; Abdelaal, K.A.A. Mitigating salt stress effects by exogenous application of proline and yeast extract on morphophysiological, biochemical and anatomical characters of calendula plants. Sci. J. Flowers Ornam. Plants 2020, 7, 461–482. [Google Scholar] [CrossRef]
- Hasan, M.K.; El Sabagh, A.; Sikdar, M.S.I.; Alam, M.J.; Ratnasekera, D.; Barutcular, C.; Abdelaal, K.A.A.; Islam, M.S. Comparative adaptable agronomic traits of Blackgram and mungbean for saline lands. Plant Arch. 2017, 17, 589–593. [Google Scholar]
- ALKahtani, M.D.F.; Attia, K.A.; Hafez, Y.M.; Khan, N.; Eid, A.M.; Ali, M.A.M.; Abdelaal, K.A.A. Chlorophyll Fluorescence Parameters and Antioxidant Defense System Can Display Salt Tolerance of Salt Acclimated Sweet Pepper Plants Treated with Chitosan and Plant Growth Promoting Rhizobacteria. Agronomy 2020, 10, 1180. [Google Scholar] [CrossRef]
- Hafez, Y.; Elkohby, W.; Mazrou, Y.S.A.; Ghazy, M.; Elgamal, A.; Abdelaal, K.A. Alleviating the detrimental impacts of salt stress on morpho-hpysiological and yield characters of rice plants (Oryza sativa L.) using actosol, Nano-Zn and Nano-Si. Fresenius Environ. Bull. 2020, 29, 6882–6897. [Google Scholar]
- Abdelaal, K.A.A.; AlKahtani, M.D.F.; Attia, K.; Hafez, Y.; Király, L.; Künstler, A. The pivotal role of plant growth promoting bacteria in alleviating the adverse effects of drought and facilitating sustainable agriculture. Biology 2021, 10, 520. [Google Scholar] [CrossRef]
- Marostica, T.F.; Cazarolli1, L.H.; Moura, G.S.; Luz, V.C.D.; Guimarães, E.A.C.M.; Cargnelutti, D. Does Allium sativum L. tolerate water deficit? Sci. Elec. Arch. 2019, 12, 43–51. [Google Scholar] [CrossRef][Green Version]
- Ragab, A.Y.; Geries, L.S.M.; Abdelaal, K.A.A.; Hanna, S.A. Growth and productivity of onion plant (Allium cepa L.) as affected by transplanting method and NPK fertilization. Fresenius Environ. Bull. 2019, 28, 7777–7786. [Google Scholar]
- AlKahtani, M.D.F.; Hafez, Y.M.; Attia, K.; Rashwan, E.; Husnain, L.A.; AlGwaiz, H.I.M.; Abdelaal, K.A.A. Evaluation of Silicon and Proline Application on the Oxidative Machinery in Drought-Stressed Sugar Beet. Antioxidants 2021, 10, 398. [Google Scholar] [CrossRef]
- Rashwan, E.; Alsohim, A.S.; El-Gammaal, A.; Hafez, Y.; Abdelaal, K.A.A. Foliar application of nano zink-oxide can alleviate the harmful effects of water deficit on some flax cultivars under drought conditions. Fresenius Environ. Bull. 2020, 29, 8889–8904. [Google Scholar]
- Abdelaal, K.A.A.; Attia, K.A.; Alamery, S.F.; El-Afry, M.M.; Ghazy, A.I.; Tantawy, D.S.; Al-Doss, A.A.; El-Shawy, E.S.E.; Abu-Elsaoud, A.M.; Hafez, Y.M. Exogenous Application of Proline and Salicylic Acid can Mitigate the Injurious Impacts of Drought Stress on Barley Plants Associated with Physiological and Histological Characters. Sustainability 2020, 12, 1736. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 17672. [Google Scholar] [CrossRef] [PubMed]
- Abdelaal, K.A.A.; Rashed, S.H.; Ragab, A.; Hossian, A.; El Sabagh, A. Yield and quality of two sugar beet (Beta vulgaris L. ssp. vulgaris var. altissima Doll) cultivars are influenced by foliar application of salicylic Acid, irrigation timing, and planting density. Acta Agric. Slov. 2020, 115, 239–248. [Google Scholar] [CrossRef]
- Abdelaal, K.; Elafry, M.; Abdel-Latif, I.; Elshamy, R.; Hassan, M.; Hafez, Y. Pivotal role of yeast and ascorbic acid in improvement the morpho-physiological characters of two wheat cultivars under water deficit stress in calcareous soil. Fresenius Environ. Bull. 2021, 30, 2554–2565. [Google Scholar]
- Hafez, Y.M.; Attia, K.A.; Alamery, S.; Ghazy, A.; Al-Dosse, A.; Ibrahim, E.; Rashwan, E.; El-Maghraby, L.; Awad, A.; Abdelaal, K.A.A. Beneficial Effects of Biochar and Chitosan on Antioxidative Capacity, Osmolytes Accumulation, and Anatomical Characters of Water-Stressed Barley Plants. Agronomy 2020, 10, 630. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; Hafez, Y.M.; El-Afry, M.M.; Tantawy, D.S.; Alshaal, T. Effect of some osmoregulators on photosynthesis, lipid peroxidation, antioxidative capacity and productivity of barley (Hordeum vulgare L.) under water deficit stress. Environ. Sci. Pollut. Res. 2018, 25, 30199–30211. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; Hafez, Y.M.; El Sabagh, A.; Saneok, H. Ameliorative effects of Abscisic acid and yeast on morpho-physiological and yield characteristics of maize plant (Zea mays L.) under water deficit conditions. Fresenius Environ. Bull. 2017, 26, 7372–7383. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Omara, R.I.; Abdelaal, K.A.A. Biochemical, histopathological and genetic analysis associated with leaf rust infection in wheat plants (Triticum aestivum L). Physiol. Mol. Plant. Pathol. 2018, 104, 48–57. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; Mazrou, Y.S.A.; Hafez, Y.M. Silicon Foliar Application Mitigates Salt Stress in Sweet Pepper Plants by Enhancing Water Status, Photosynthesis, Antioxidant Enzyme Activity and Fruit Yield. Plants 2020, 9, 733. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; El-Afry, M.; Metwaly, M.; Zidan, M.; Rashwan, E. Salt tolerance activation in faba bean plants using proline and salicylic acid associated with physio-biochemical and yield characters improvement. Fresenius Environ. Bull. 2021, 30, 3175–3186. [Google Scholar]
- Pongprayoon, W.; Roytrakul, S.; Pichayagkura, R.; Chadchawan, S. The role of hydrogen peroxide in chitosan-induced resistance to osmotic stress in rice (Oryza sativa L.). Plant Growth Regul. 2013, 70, 159–173. [Google Scholar] [CrossRef]
- Monirul, I.M.; Humayun, K.M.; Mamun, A.N.K.; Monirul, I.; Pronabananda, D. Studies on yield and yield attributes in tomato and chilli using foliar application of oligo-chitosan. GSC Biol. Pharm. Sci. 2018, 3, 20–28. [Google Scholar]
- Ahmed, K.B.M.; Khan, M.M.A.; Siddiqui, H.; Jahan, A. Chitosan and its oligosaccharides, a promising option for sustainable crop production—A review. Carbohydr. Polym. 2020, 227, 115331. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Luo, X.; Tu, R. Application of bioactive coatings based on chitosan for soybean seed protection. Int. J. Carbohydr. Chem. 2012, 2012, 104565. [Google Scholar] [CrossRef]
- Bistgani, Z.E.; Siadat, S.A.; Bakhshandeh, A.; Pirbalouti, A.G.; Hashemi, M. Morpho-physiological and phytochemical traits of (Thymus daenensis Celak.) in response to deficit irrigation and chitosan application. Acta Physiol. Plant. 2017, 39, 231. [Google Scholar] [CrossRef]
- Barnett, J.A.; Payne, R.W.; Yarrow, D. (Eds.) Yeasts: Characterisation and Identification, 3rd ed.; Cambridge University Press: Cambridge, MA, USA, 1990. [Google Scholar] [CrossRef]
- Matter, F.M.A.; Abou-Sreea, A.I.B. Influence of application methods of bio-fertilization on morhological growth characters, seed yield and chemical composition of fenugreek plants. Egypt. J. Hort. 2016, 43, 19–33. [Google Scholar]
- Nagodowithana, W.T. Yeast Technology; Van Nostrsnd Reinhold: New York, NY, USA, 1991; 273p. [Google Scholar]
- Haider, I.; Raza, M.; Sammar, A.; Iqbal, R.; Ahmad, S.; Aslam, M.U.; Israr, M.; Riaz, U.; Sarfraz, M.; Abbas, N.; et al. Alleviating the Drought Stress in Wheat (Triticum aestivum L.) by Foliar Application of Amino Acid and Yeast. Pak. J. Agric. Res. 2021, 34, 239–246. [Google Scholar] [CrossRef]
- Sarhan, T.; Abdullah, O.K. Effect of Azotobacter inoculation. Dry bread yeast suspension varying levels of urea on growth of potato Cv. Desiree. In Proceedings of the Tropentag World Food System—A Contribution from Europe, Zurich, Switzerland, 14–16 September 2010. [Google Scholar]
- Shalaby, T.A.; El-Ramady, H. Effect of foliar application of bio-stimulants on growth, yield, components, and storability of garlic (Allium sativum L.). Aust. J. Crop Sci. 2014, 8, 271–275. [Google Scholar]
- Ali, M.A.M. Effect of some Bio-stimulants on Growth, Yield and Bulb Quality of Garlic Grown in Newly Reclaimed Soil, New Valley-Egypt. J. Plant Prod. Mansoura Univ. 2017, 8, 1285–1294. [Google Scholar] [CrossRef][Green Version]
- Association of Official Analytical Chemists (A.O.A.C.). Official Methods of Analysis, 26th ed.; A.O.A.C. International: Washington, DC, USA, 2005. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Sanchez, F.J.; de Andrés, E.F.; Tenorio, J.L.; Ayerbe, L. Growth of epicotyls, turgor maintenance and osmotic adjustment in pea plants (Pisum sativum L.) subjected to water stress. Field Crop. Res. 2004, 86, 81–90. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Szalai, G.; Janda, T.; Padi, E.; Szigeti, Z. Role of light in post-chilling symptoms in maize. J. Plant Physiol. 1996, 148, 378–383. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Yesbergenova, Z.; Yang, G.; Oron, E.; Soffer, D.; Fluhr, R.; Sagi, M. The plant Mo-hydroxylases aldehyde oxidase and xanthine dehy-drogenase have distinct reactive oxygen species signatures and are induced by drought and abscisic acid. Plant J. 2005, 42, 862–876. [Google Scholar] [CrossRef]
- Yu, C.W.; Murphy, T.M.; Lin, C.H. Hydrogen peroxide-induces chilling tolerance in mung beans mediated through ABA independent glutathione accumulation. Funct. Plant Biol. 2003, 30, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplast. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Havir, E.A.; McHale, N.A. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef]
- Spitz, D.R.; Oberley, L.W. An assay for superoxide dismutase activity in mammalian tissue homogenates. Anal. Biochem. 1989, 179, 8–18. [Google Scholar] [CrossRef]
- Castillo, F.I.; Penel, I.; Greppin, H. Peroxidase release induced by ozone in sedum album leaves. Plant Physiol. 1984, 74, 846–851. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; Wiley Inter Science: New York, NY, USA, 1984; 690p. [Google Scholar]
- Duncan, B.D. Multiple ranges and multiple F-test. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Liu, F.; Christian, R.; Shahanzari, J.A.; Andersen, M.N.; Jacobsen, E.E. ABA regulated stomata control and photosynthetic water use efficiency of potato (Solanum tuberosum L.) during progressive soil drying. Plant Sci. 2005, 168, 831–836. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef] [PubMed]
- Abu-Muriefah, S.S. Effect of chitosan on common bean (Phaseolus vulgaris L.) plants grown under water stress conditions. Int. Res. J. Agric. Sci. Soil Sci. 2013, 3, 192–199. [Google Scholar]
- Farouk, S.; Amany, A.R. Improving growth and yield of cowpea by foliar application of chitosan under water stress. Egypt. J. Biol. 2012, 14, 14–16. [Google Scholar] [CrossRef]
- Lai, Q.; Zhi-yi, B.; Zhu-Jun, Z.; Qiong-Qiu, Q.; Bi-Zeng, M. Effects of osmotic stress on antioxidant enzymes activities in leaf discs of PSAG12-IPT modified gerbera. J. Zhejiang Univ. Sci. 2007, 8, 458–464. [Google Scholar] [CrossRef]
- Shinde, B.P.; Thakur, J. Influence of Arbuscular mycorrhizal fungi on chlorophyll, proteins, proline and total carbohydrates content of the pea plant under water stress condition. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 809–821. [Google Scholar]
- Gedam, P.A.; Thangasamy, A.; Shirsa, D.V.; Ghosh, S.; Bhagat, K.P.; Sogam, O.A.; Gupta, A.J.; Mahajan, V.; Soumia, P.S.; Salunkhe, V.N.; et al. Screening of Onion (Allium cepa L.) Genotypes for Drought Tolerance Using Physiological and Yield Based Indices Through Multivariate Analysis. Front. Plant Sci. 2021, 12, 600371. [Google Scholar] [CrossRef]
- Massacci, A.; Nabiev, S.M.; Pietrosanti, L.; Nematov, S.K.; Chernikova, T.N.; Thor, K.; Leipner, J. Response of the photosynthetic apparatus of cotton (Gossypium hirsutum) to the onset of drought stress under field conditions studied by gas-exchange analysis and chlorophyll fluorescence imaging. Plant Physiol. Biochem. 2008, 46, 189–195. [Google Scholar] [CrossRef]
- Nayyar, H.; Gupta, D. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot. 2006, 58, 106–113. [Google Scholar] [CrossRef]
- AL-Quraan, N.A.; Al-Ajlouni, Z.I.; Qawasma, N.F. Physiological and Biochemical Characterization of the GABA Shunt Pathway in Pea (Pisum sativum L.) Seedlings under Drought Stress. Horticulturae 2021, 7, 125. [Google Scholar] [CrossRef]
- Possingham, J.V. Plastid replication and development in the life cycle of higher plants. Annu. Rev. Plant Physiol. 1980, 31, 113–129. [Google Scholar] [CrossRef]
- Khan, W.M.; Prithiviraj, B.; Smiyh, D.L. Effect of foliar application of chitin oligosaccharides on photosynthesis of maize and soybean. Photosynthetica 2002, 40, 87. [Google Scholar] [CrossRef]
- Gupta, A.; Medina-Rico, A.; Delgado-Cano, A. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Ghaffaria, H.; Tadayona, M.R.; Bahadora, M.; Razmjoo, J. Investigation of the proline role in controlling traits related to sugar and root yield of sugar beet under water deficit conditions. Agric. Water Manag. 2021, 243, 106448. [Google Scholar] [CrossRef]
- Omar, A.; Zayed, B.; Abdel Salam, A.; Hafez, Y.M.; Abdelaal, K.A.A. Folic acid as foliar application can improve growth and yield characters of rice plants under irrigation with drainage water. Fresenius Environ. Bull. 2020, 29, 9420–9428. [Google Scholar]
- Shahin, A.; Esmaeil, R.A.; Badr, M.; Abdelaal, K.A.A.; Hassan, F.A.S.; Hafez, Y.M. Phenotypic characterization of race-specific and slow rusting resistance to stem rust disease in promising wheat genotypes. Fresenius Environ. Bull. 2021, 30, 6223–6236. [Google Scholar]
- Esmail, S.M.; Omara, R.I.; Abdelaal, K.A.; Hafez, M. Histological and biochemical aspects of compatible and incompatible wheat-Puccinia striiformis interactions. Physiol. Mol. Plant Pathol. 2019, 106, 120–128. [Google Scholar] [CrossRef]
- Rocychoudhury, A.; Banerjee, A. Endogenous glycine betaine accumulation mediates abiotic stress tolerance in plants. Trop. Plant Res. 2016, 3, 105–111. [Google Scholar]
- Kheradmand, M.A.; Shahmoradzadeh, F.S.; Fatahi, E.; Raoofi, M.M. Effect of water stress on oil yield and some characteristics of Brassica napus. Int. Res. J. Appl. Basic Sci. 2014, 8, 1447–1453. [Google Scholar]
- Yang, F.; Hu, J.; Li, J.; Wu, X.; Qian, Y. Chitosan enhances leaf membrane stability and antioxidant enzyme activities in apple seedlings under drought stress. Plant Growth Regul. 2009, 58, 131–136. [Google Scholar] [CrossRef]
- Jiao, Z.; Li, Y.; Li, J.; Xu, X.; Li, H.; Lu, D.; Wang, J. Effects of exogenous chitosan on physiological characteristics of potato seedlings under drought stress and rehydration. Potato Res. 2012, 55, 293–301. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Rady, M.O.A.; Marey, R.A.; Abd El-Mageed, T.A. Exogenously applied proline enhances growth and productivity of drought stressed onion by improving photosynthetic efficiency, water use efficiency and up-regulating osmoprotectants. Sci. Hortic. 2020, 272, 109580. [Google Scholar] [CrossRef]
- Sun, T.; Xie, W.M.; Xu, P.X. Superoxide anion scavenging activity of graft chitosan derivatives. Carbohydr. Polym. 2004, 58, 379–382. [Google Scholar] [CrossRef]
- Bittelli, M.; Flury, M.; Campbell, G.S.; Nichols, E.J. Reduction of transpiration through foliar application of chitosan. Agric. For. Meteorol. 2001, 107, 167–175. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Zhang, X.; Merewitz, E.; Peng, Y.; Ma, X.; Yan, Y. Metabolic pathways regulated by chitosan contributing to drought resistance in white clover. J. Proteome Res. 2017, 16, 3039–3052. [Google Scholar] [CrossRef]
- Su, Y.; Xia, S.; Zhong, R.; Wang, L. Phytohormonal quantification based on biological principles. Horm. Metab. Signal. Plants 2017, 13, 431–470. [Google Scholar]
- Vasconcelos, A.C.F.D.; Chaves, L.H.G. Biostimulants and their role in improving plant growth under abiotic stresses. In Biostimulants in Plant Science; Intech Open: London, UK, 2019. [Google Scholar] [CrossRef]
- Landi, L.; De Miccoli, R.M.; Pollastro, S.; Feliziani, E.; Faretra, F.; Romanazzi, G. Global transcriptome analysis and identification of differentially expressed genes in strawberry after preharvest application of Benzothiadiazole and chitosan. Front. Plant Sci. 2017, 8, 235. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelaal, K.; Attia, K.A.; Niedbała, G.; Wojciechowski, T.; Hafez, Y.; Alamery, S.; Alateeq, T.K.; Arafa, S.A. Mitigation of Drought Damages by Exogenous Chitosan and Yeast Extract with Modulating the Photosynthetic Pigments, Antioxidant Defense System and Improving the Productivity of Garlic Plants. Horticulturae 2021, 7, 510. https://doi.org/10.3390/horticulturae7110510
Abdelaal K, Attia KA, Niedbała G, Wojciechowski T, Hafez Y, Alamery S, Alateeq TK, Arafa SA. Mitigation of Drought Damages by Exogenous Chitosan and Yeast Extract with Modulating the Photosynthetic Pigments, Antioxidant Defense System and Improving the Productivity of Garlic Plants. Horticulturae. 2021; 7(11):510. https://doi.org/10.3390/horticulturae7110510
Chicago/Turabian StyleAbdelaal, Khaled, Kotb A. Attia, Gniewko Niedbała, Tomasz Wojciechowski, Yaser Hafez, Salman Alamery, Talal K. Alateeq, and Sally A. Arafa. 2021. "Mitigation of Drought Damages by Exogenous Chitosan and Yeast Extract with Modulating the Photosynthetic Pigments, Antioxidant Defense System and Improving the Productivity of Garlic Plants" Horticulturae 7, no. 11: 510. https://doi.org/10.3390/horticulturae7110510
APA StyleAbdelaal, K., Attia, K. A., Niedbała, G., Wojciechowski, T., Hafez, Y., Alamery, S., Alateeq, T. K., & Arafa, S. A. (2021). Mitigation of Drought Damages by Exogenous Chitosan and Yeast Extract with Modulating the Photosynthetic Pigments, Antioxidant Defense System and Improving the Productivity of Garlic Plants. Horticulturae, 7(11), 510. https://doi.org/10.3390/horticulturae7110510