
First Report of Three Tylenchidae Taxa from Southern Alberta, Canada



Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Morphological Studies
2.2. DNA Extraction, PCR, and Sequencing
2.3. Phylogenetic Analyses
3. Results
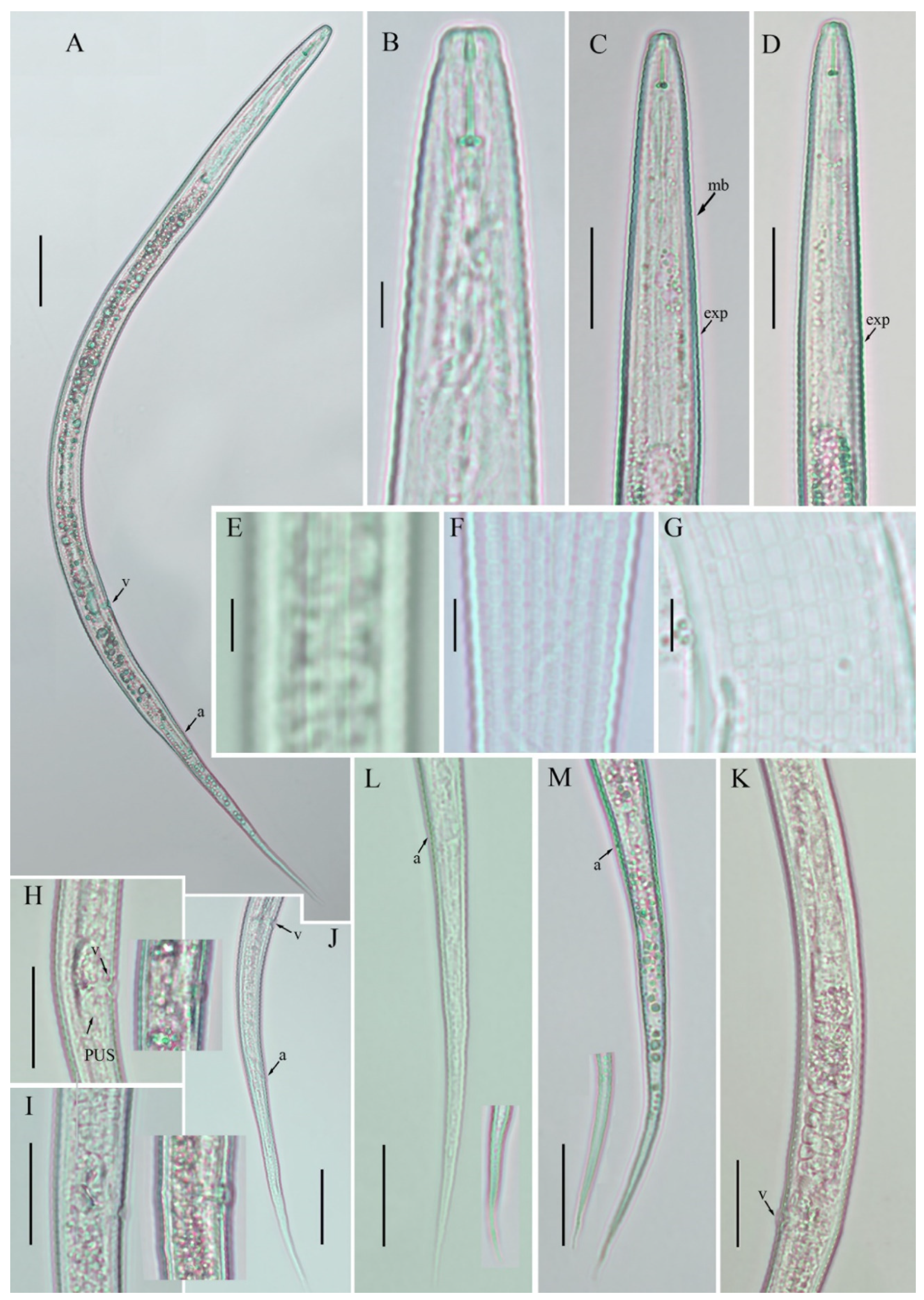
3.1. Description of Basiria bhabi
3.2. Description of Coslenchus acceptus
3.3. Description of Filenchus vulgaris

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brzeski [24] | Raski and Geraert [48] | Karegar and Geraert [49] | ||||
|---|---|---|---|---|---|---|
| Characters | Canadian Population | Poland | Colorado | Belgium | Korea | Iran |
| n * | 15 | 25 | 5 | 51 | 11 | 5 |
| Body length | 779.1 ± 61.8 (677.0–870.0) | 520–810 | 650–690 | 490–750 | 560–740 | 580–730 |
| a | 29.9 ± 2.8 (25.3–36.5) | 29–42 | 36–40 | 24–42.5 | – | 31.6–37.2 |
| b | 6.0 ± 0.3 (5.4–6.5) | 4.7–7.0 | 5.6–5.9 | 4.7–6.4 | 5.1–6.9 | 5.9–6.4 |
| c | 5.3 ± 0.4 (4.7–6.2) | 4.2–5.7 | 4.2–4.9 | 3.4–5.3 | 4.6–5.3 | 3.9–4.5 |
| c’ | 9.7 ± 1.0 (8.2–11.7) | 13 | 12–15 | – | – | 13.4–17.4 |
| V | 61.2 ± 1.4 (59.0–64.0) | 53–65 | 58–61 | 53–64 | 60–65 | 54.6–60.2 |
| MB | 41.2 ± 1.4 (39.7–45.1) | 44 | 40–43 | 40–48 | 39–44.5 | 38.1–45.8 |
| Lip height | 3.2 ± 0.3 (3.0–4.0) | – | – | – | – | – |
| Lip width | 6.5 ± 0.4 (6.0–7.0) | – | – | – | – | – |
| Stylet length | 10.5 ± 1.0 (9.0–12.0) | 10–12 | 10–11 | 9–12 | 9.5–10.5 | 8.1–10.5 |
| Dorsal pharyngeal gland orifice (DGO) | 1.5 ± 0.2 (1.0–2.0) | – | – | – | – | – |
| Median bulb length | 12.4 ± 0.9 (11.0–14.0) | – | – | – | – | – |
| Median bulb width | 7.7 ± 0.7 (7.0–9.0) | – | – | – | – | – |
| Pharynx length | 129.4 ± 5.8 (120.0–141.0) | 114 | 111–123 | 89–123 | 98–124 | 96–115 |
| Anterior end to excretory pore | 98.9 ± 6.8 (87.0–110.0) | 92–102 | 84–94 | – | – | – |
| Maximum body width | 26.3 ± 2.4 (22.0–30.0) | – | – | – | – | – |
| Vulva body width | 24.4 ± 1.9 (21.5–27.5) | – | – | – | – | – |
| Post-vulval uterine sac (PUS) length | 14.1 ± 1.8 (12.0–17.0) | – | – | – | – | – |
| Distance from vulva to anus | 149.3 ± 12.4 (125.0–166.0) | – | – | – | – | 105–122 |
| Anal body width | 15.3 ± 1.2 (13.0–17.0) | – | – | – | – | – |
| Tail length | 146.9 ± 12.0 (133.0–175.0) | 135 | 137–157 | 111–165 | 114–150 | 145–172 |
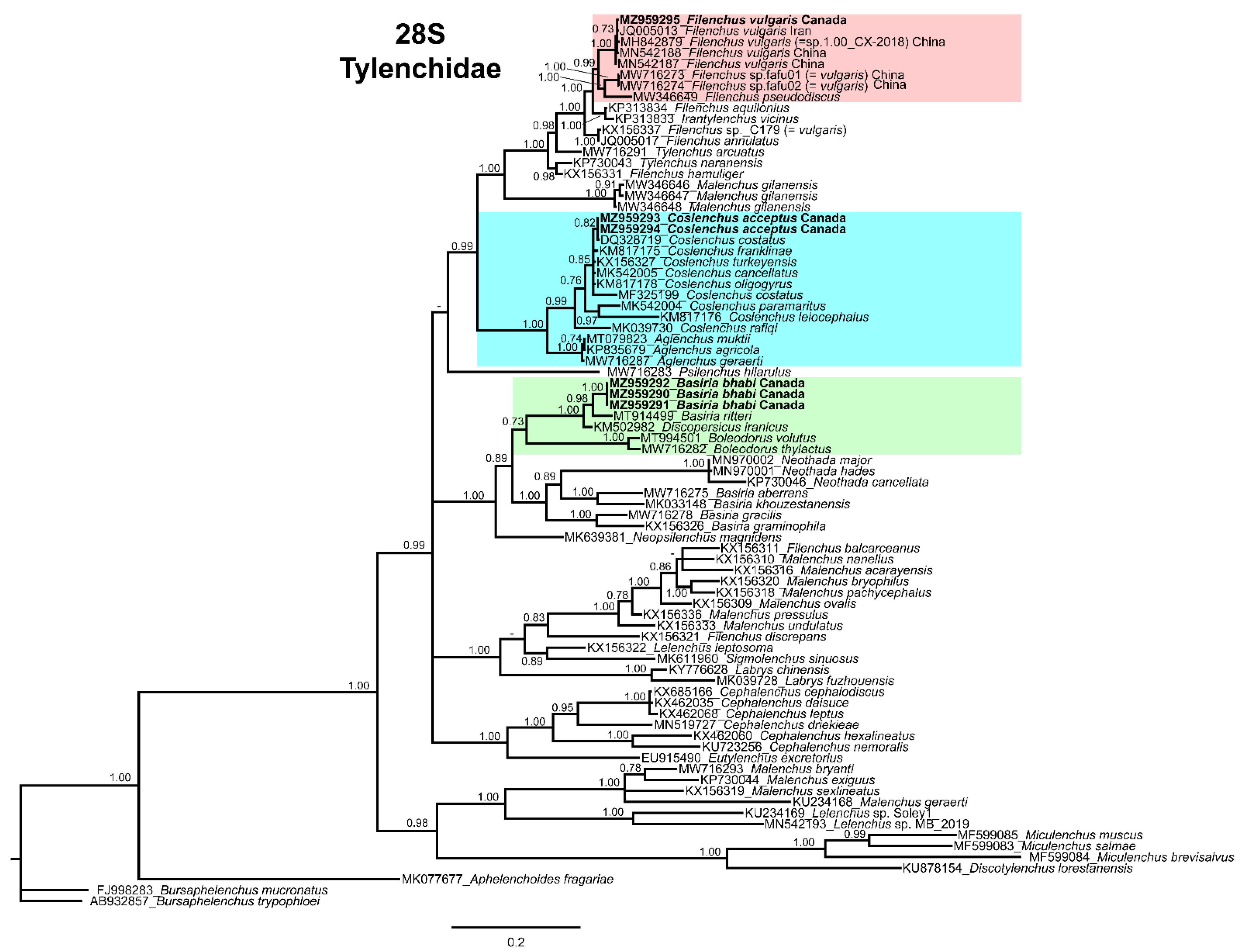
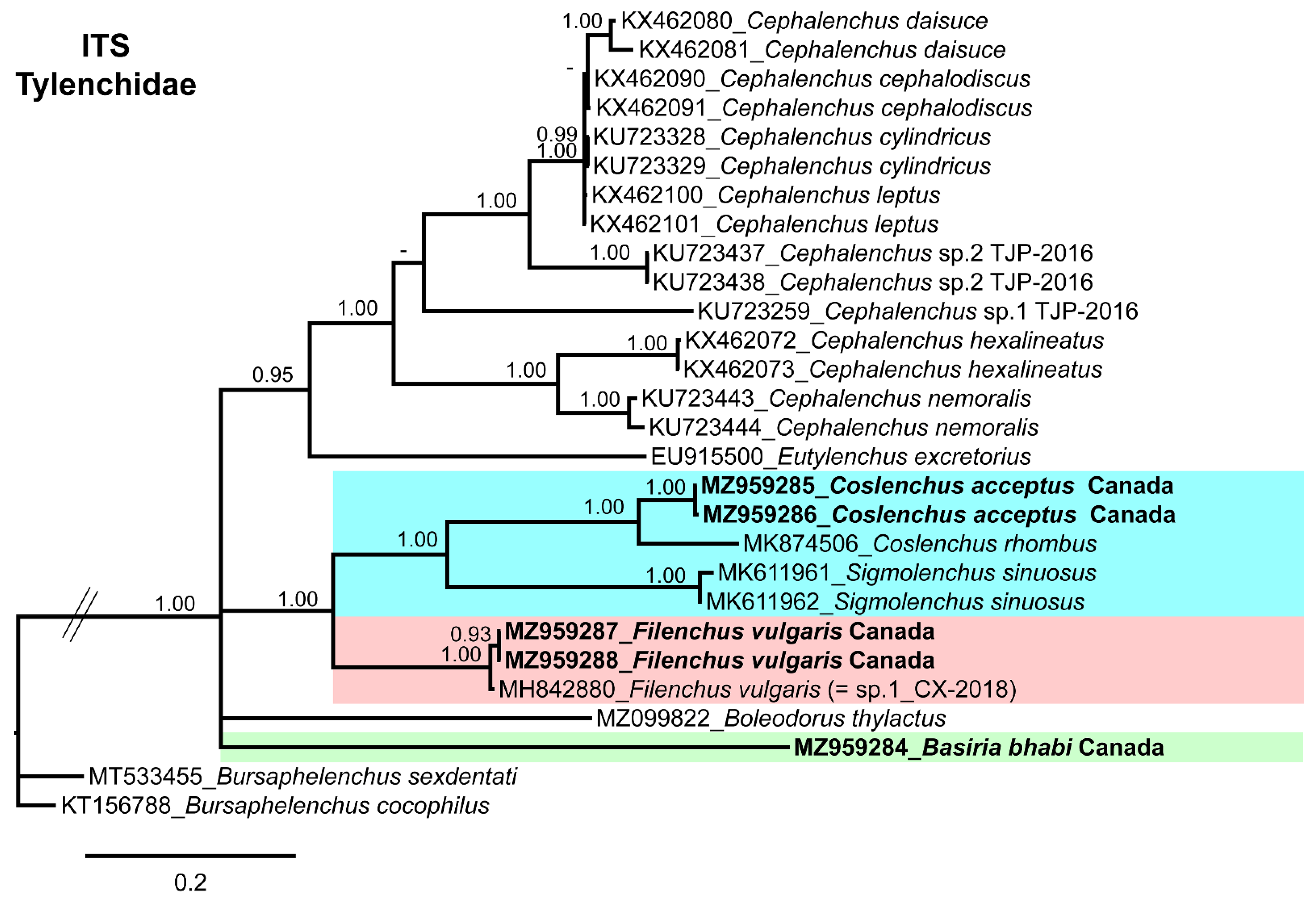
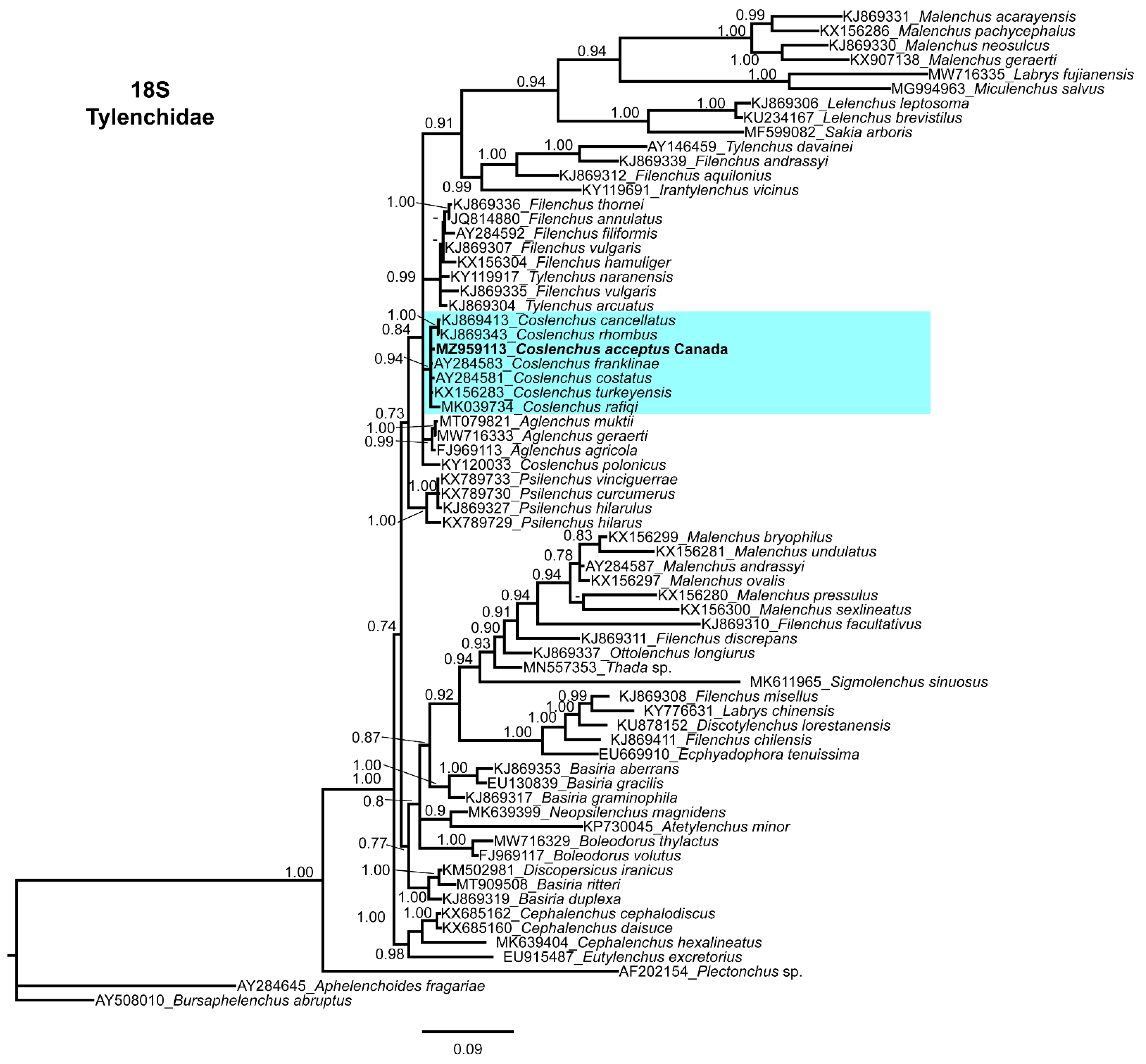
3.4. Molecular Characterization and Phylogenetic Relationship of Basiria bhabi, Coslenchus acceptus, and Filenchus vulgaris with Related Tylenchidae Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Örley, L. Monograph of the Anguillulids. Természetrajzi Füzetek 1880, 4, 16–150. [Google Scholar]
- Geraert, E. Tylenchidae in agricultural soils. In Manual of Agricultural Nematology; CRC Press: Boca Raton, FL, USA, 1991; pp. 795–826. [Google Scholar]
- Siddiqi, M. Tylenchida Parasites of Plants and Insects; CABI Publishing: Wallingford, UK, 2000; p. 833. [Google Scholar]
- Bai, M.; Qing, X.; Qiao, K.; Ning, X.; Xiao, S.; Cheng, X.; Liu, G. Mitochondrial COI gene is valid to delimitate Tylenchidae (Nematoda: Tylenchomorpha) species. J. Nematol. 2020, 52, e2020-38. [Google Scholar] [CrossRef] [Green Version]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Qing, X.; Bert, W. Family Tylenchidae (Nematoda): An overview and perspectives. Org. Divers. Evol. 2019, 19, 391–408. [Google Scholar] [CrossRef]
- Panahandeh, Y.; Pourjam, E.; Pedram, M. Data on some species of the genus Coslenchus Siddiqi, 1978 (Rhabditida, Tylenchidae) from Iran. J. Nematol. 2016, 48, 268–279. [Google Scholar] [CrossRef] [Green Version]
- Okada, H.; Harada, H.; Kadota, I. Fungal-feeding habits of six nematode isolates in the genus Filenchus. Soil Biol. Biochem. 2005, 37, 1113–1120. [Google Scholar] [CrossRef]
- Čerevková, A.; Miklisová, D.; Bobuľská, L.; Renčo, M. Impact of the invasive plant Solidago gigantea on soil nematodes in a semi-natural grassland and a temperate broadleaved mixed forest. J. Helminthol. 2020, 94, e51, 1–14. [Google Scholar] [CrossRef]
- McSorley, R.; Frederick, J. Nematode population fluctuations during decomposition of specific organic amendments. J. Nematol. 1999, 31, 37–44. [Google Scholar] [PubMed]
- Pereira, T.J.; Baldwin, J.G. Contrasting evolutionary patterns of 28S and ITS rRNA genes reveal high intragenomic variation in Cephalenchus (Nematoda): Implications for species delimitation. Mol. Phylogenet. Evol. 2016, 98, 244–260. [Google Scholar] [CrossRef]
- Munawar, M.; Yevtushenko, D.P.; Castillo, P. Integrative taxonomy, distribution, and host associations of Geocenamus brevidens and Quinisulcius capitatus from southern Alberta, Canada. J. Nematol. 2021, 53, e2021-15. [Google Scholar] [CrossRef]
- Munawar, M.; Yevtushenko, D.P.; Palomares-Rius, J.E.E.; Castillo, P. Species diversity of pin nematodes (Paratylenchus spp.) from potato growing regions of southern Alberta, Canada. Plants 2021, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, M.R. Basiria graminophila ng, n. sp. (Nematoda: Tylenchinae) found associated with grassroots in Aligarh India. Nematologica 1959, 4, 217–222. [Google Scholar] [CrossRef]
- Siddiqi, M.R. The unusual position of the phasmids in Coslenchus costatus (de Man, 1921) gen. n., comb. n. and other Tylenchidae (Nematoda: Tylenchida). Nematologica 1978, 24, 449–455. [Google Scholar] [CrossRef]
- Andrássy, I. Revision der Gattung Tylenchus Bastian, 1865 (Tylenchidae, Nematoda). Acta Zool. Hung 1954, 1, 5–42. [Google Scholar]
- Lumactud, R.A. Spatial Distribution of Soil Nematodes in the Sub-Arctic Environment of Churchill, Manitoba. Master’s Thesis, The University of Manitoba, Winnipeg, MB, Canada, 2010. [Google Scholar]
- Pereira, F.G. Survey of plant-parasitic nematodes in pulse crop fields of the Canadian prairies. Master’s Thesis, The University of Manitoba, Winnipeg, MB, Canada, 2018. [Google Scholar]
- Wu, L.-Y. Dactylotylenchus crassacuticulus, a new genus and new species (Tylenchinae: Nematoda). Can. J. Zool. 1968, 46, 831–834. [Google Scholar] [CrossRef]
- Wu, L.-Y. Five new species of Tylenchus Bastian, 1865 (Nematoda: Tylenchidae) from the Canadian high Arctic. Can. J. Zool. 1969, 47, 1005–1010. [Google Scholar] [CrossRef]
- Geraert, E. The Tylenchidae of the World: Identification of the Family Tylenchidae (Nematoda); Academia Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Siddiqi, M. Tylenchida Parasites of Plants and Insects; Commonwealth Agricultural Bureaux Farnham Royal: Slough, UK, 1986; 646p. [Google Scholar]
- Andrássy, I. The genera and species of the family Tylenchidae Örley, 1880 (Nematoda): The genus Coslenchus Siddiqi, 1978. Acta Zool. Acad. Sci. Hung. 1982, 28, 193–232. [Google Scholar]
- Brzeski, M. Tylenchus ditissimus sp. n., a new nematode from Poland (Nematoda, Tylenchidae). Bulletin de l’Academie Polonaise des Sciences. Cl. II Ser. Sci. Biol. 1963, 11, 537–540. [Google Scholar]
- Lownsbery, J.; Lownsbery, B. Plant-parasitic nematodes associated with forest trees in California. Hilgardia 1985, 53, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, W. A rapid centrifugal-flotation technique for separating nematodes from soil. Plant. Dis. Rep. 1964, 48, 692. [Google Scholar]
- Seinhorst, J. A rapid method for the transfer of nematodes from fixative to anhydrous glycerin. Nematologica 1959, 4, 67–69. [Google Scholar] [CrossRef] [Green Version]
- De Grisse, A.T. Redescription ou modifications de quelques technique utilis [a] es dan l’etude des n [a] ematodes phytoparasitaires. Meded. Rijksfakulteit Landbowwetenschappen Gent. 1969, 34, 351–369. [Google Scholar]
- Maria, M.; Powers, T.; Tian, Z.; Zheng, J. Distribution and description of criconematids from Hangzhou, Zhejiang Province, China. J. Nematol. 2018, 50, 183–206. [Google Scholar]
- Holterman, M.; van der Wurff, A.; van den Elsen, S.; van Megen, H.; Bongers, T.; Holovachov, O.; Bakker, J.; Helder, J. Phylum-wide analysis of SSU rDNA reveals deep phylogenetic relationships among nematodes and accelerated evolution toward crown clades. Mol. Biol. Evol. 2006, 23, 1792–1800. [Google Scholar] [CrossRef]
- De Ley, P.; Felix, M.-A.; Frisse, L.; Nadler, S.; Sternberg, P.; Thomas, W.K. Molecular and morphological characterisation of two reproductively isolated species with mirror-image anatomy (Nematoda: Cephalobidae). Nematology 1999, 1, 591–612. [Google Scholar] [CrossRef]
- Ferris, V. Variation in spacer ribosomal DNA in some cyst-forming species of plant parasitic nematodes. Fundam. Appl. Nematol. 1993, 16, 177–184. [Google Scholar]
- Curran, J.; Driver, F.; Ballard, J.; Milner, R. Phylogeny of Metarhizium: Analysis of ribosomal DNA sequence data. Mycol. Res. 1994, 98, 547–552. [Google Scholar] [CrossRef]
- Pereira, T.J.; Qing, X.; Chang, K.F.; Mundo-Ocampo, M.; Cares, J.E.; Ragsdale, E.J.; Nguyen, C.N.; Baldwin, J.G. Phylogeny and biogeography of the genus Cephalenchus (Tylenchomorpha, Nematoda). Zool. Scr. 2017, 46, 506–520. [Google Scholar] [CrossRef]
- Qing, X.; Decraemer, W.; Claeys, M.; Bert, W. Molecular phylogeny of Malenchus and Filenchus (Nematoda: Tylenchidae). Zool. Scr. 2017, 46, 625–636. [Google Scholar] [CrossRef]
- Hosseinvand, M.; Eskandari, A.; Ghaderi, R. Morphological and molecular characterization of Coslenchus paramaritus n. sp. and C. cancellatus (Cobb, 1925) Siddiqi, 1978 (Nematoda: Tylenchidae) from Iran. J. Nematol. 2019, 51, e2019-59. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, P.; Heydari, F.; Abolafia, J.; Castillo, P.; Pedram, M. Morphological and molecular characterization of Filenchus pseudodiscus n. sp. from east Golestan province, north Iran; with an updated phylogeny of Malenchus Andrássy, 1968 (Tylenchomorpha: Tylenchidae). J. Nematol. 2021, 53, e2021-69. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v1. 4.2, a Graphical Viewer of Phylogenetic Trees. 2014. Available online: https://github.com/rambaut/figtree (accessed on 25 August 2021).
- Bajaj, H.; Bhatti, D. Two new species of Basiria Siddiqi, 1959 (Tylenchida) from Haryana, India. Indian J. Nematol. 1979, 8, 95–101. [Google Scholar]
- Chawla, M.; Prasad, S.; Khan, E.; Nand, S. Two new species of the genus Tylenchus Bastian, 1865 (Nematoda: Tylenchidae) from Utter Pradesh, India. Labdev. J. Sci. Technol. 1969, 4, 291–294. [Google Scholar]
- Brzeski, M. Redescription of some species of the genus Filenchus Andrássy, 1954 (Nematoda, Tylenchidae). Miscel Lània Zoològica 1997, 20, 45–64. [Google Scholar]
- Kazachenko, I. New nematode species of the family Tylenchidae Örley, 1880, form litters of coniferous forests. Biol. Pochviennovo Inst. Novaya Ceriya Vladivostok 1975, 26, 178–186. [Google Scholar]
- Raski, D.; Geraert, E. Review of the genus Filenchus Andrássy, 1954 and descriptions of six new species (Nemata: Tylenchidae). Nematologica 1986, 32, 265–311. [Google Scholar]
- Karegar, A.; Geraert, E. The Genus Filenchus Andrássy, 1954 (Nemata: Tylenchidae) from Iran. Belg. J. Zool. 1995, 125, 363–382. [Google Scholar]
- Sturhan, D.; Hohberg, K. Nematodes of the order Tylenchida in Germany–the non-phytoparasitic species. Soil Org. 2016, 88, 19–41. [Google Scholar]
- Khan, E. Boleodorus mirus n. sp. (Tylenchida: Boleodorinae n. subfam.) from Kufri, Simla (HP) India, with a key to the species of the genus Boleodorus Thorne, 1941. Zool. Anz. 1964, 173, 336–341. [Google Scholar]
- Baqri, Q.H.; Jairajpuri, M.S. Two known and three new species of nematodes associated with fibrous crops in India. Ann. Zool. Ecol. Anim. 1969, 1, 327–337. [Google Scholar]
- Bernard, E. Ditylenchus intermedius (De Man) Filipjev (Nematoda: Anguinidae) and Basiria hiberna, n. sp. (Nematoda: Psilenchidae) from Tennessee. Proc. Biol. Soc. Wash. 1980, 93, 303–310. [Google Scholar]
- Ghaemi, R.; Pourjam, E.; Atighi, M.R.; Pedram, M.; Karssen, G. First record of the genus Discotylenchus Siddiqi, 1980 (Nematoda: Tylenchidae) from Iran, with description of one new and data on two known species. Zootaxa 2012, 3493, 72–82. [Google Scholar] [CrossRef]
- Yaghoubi, A.; Pourjam, E.; Alvarez-Ortega, S.; Liebanas, G.; Atighi, M.R.; Pedram, M. Discopersicus n. gen., a new member of the family Tylenchidae Örley, 1880 with detailed SEM study on two known species of the genus Discotylenchus Siddiqi, 1980 (Nematoda; Tylenchidae) from Iran. J. Nematol. 2016, 48, 214–221. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.; Siddiqi, M.R. Boleodorus volutus n. sp. (Nematoda: Nothotylenchinae) found in soil about grass roots in England. Nematologica 1963, 9, 19–23. [Google Scholar] [CrossRef]
- Thorne, G. Some nematodes of the family Tylenchidae which do not possess a valvular median esophageal bulb. Great Basin Nat. 1941, 2, 37–85. [Google Scholar]
- Thorne, G. On the classification of the Tylenchida, new order (Nematoda, Phasmidia). Proc. Helminthol. Soc. Wash. 1949, 16, 37–73. [Google Scholar]
- Siddiqi, M.R. Four new species of the genus Tylenchus Bastian, 1865 (Nematoda) from North India. Z. Für Parasitenkd. 1963, 23, 170–180. [Google Scholar] [CrossRef]
- Eisvand, P.; Nejad, R.F.; Azimi, S. Description of Basiria khouzestanensis n. sp. (Nematoda: Tylenchidae) from Iran and its phylogenetic relationships with other species in the family. Zootaxa 2019, 4563, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Khan, S. On the proposal for Neothada n. gen.(Nematoda: Tylenchinae). Proceedings of the National Academy of Sciences. India Biol. Sci. 1973, 43, 17–18. [Google Scholar]
- Thorne, G.; Malek, R.B. Nematodes of the northern Great Plains. Part I. Tylenchida (Nemata: Secernentea). Tech. Bull. S. Dakota Agric. Exp. Stn. 1968, 31, 1–111. [Google Scholar]
- Skarbilovich, T. On the structure of systematics of nematodes order Tylenchida Thorne, 1949. Acta Parasitol. Pol. 1959, 7, 117–132. [Google Scholar]
- Cobb, N. Biological relationships of the mathematical series 1, 2, 4, etc., with a description of a new nema, Tylenchus Cancellatus. Contrib. A Sci. Nematol. 1925, 15, 371–374. [Google Scholar]
- de Man, J.C. Nouvelles recherches sur les nématodes libres terricoles de la Hollande. Capita Zool. 1921, 1, 3–62. [Google Scholar]
- Siddiqi, M.R. Six new species of Coslenchus Siddiqi, 1978 (Nematoda: Tylenchidae). Nematologica 1980, 26, 432–447. [Google Scholar] [CrossRef]
- Brzeski, M.W. Nematodes of Tylenchina in Poland and Temperate Europe; Muzeum i Instytutu Zoologii, Polska Akademia Nauk (MiIZ PAN): Warsaw, Poland, 1998. [Google Scholar]
- Brzeski, M.W. Taxonomic Notes on Coslenchus Siddiqi, 1978 (Nematoda: Tylenchidae). Ann. Zool. 1987, 40, 417–436. [Google Scholar]
- Siddiqui, A.; Khan, E. Taxonomic studies on tylenchidae (nematoda) of India. II. Descriptions of two new sp. of Cosaglenghus gen. n. alongwith proposition of a new sub-family aglenchinae. Indian J. Nematol. 1982, 12, 303–311. [Google Scholar]
- de Man, J.G. Die, Frei in der Reinen Erde und Im Süssen Wasser Lebenden Nematoden der Niederländischen Fauna: Eine Systematisch-Faunistische Monographie; EJ Brill: Leiden, The Netherlands, 1884; Volume 1, p. 206. [Google Scholar]
- Mizukubo, T. Aglenchus ainakamurae n. sp. and records of Cephalenchus leptus and Lelenchus leptosoma from Japan (Tylenchida: Tylenchidae). Jpn. J. Nematol. 1989, 19, 18–31. [Google Scholar]
- Phukan, P.; Sanwal, K. Two new species of Aglenchus and record of Cephalenchus leptus (Tylenchidae: Nematoda) from Assam. Indian J. Nematol 1980, 10, 28–34. [Google Scholar]
- Siddiqi, M.R. Two new nematode genera, Safianema (Anguinidae and Discotylenchus (Tylenchidae), with descriptions of three new species. Proc. Helminthol. Soc. Wash. 1980, 47, 85–94. [Google Scholar]
- Andrássy, I. Fauna Paraguayensis. 2. Nematoden aus den Galeriewäldern des Acaray-Flusses. Opusc. Zool. Bp. 1968, 8, 167–312. [Google Scholar]
- Andrássy, I. Freilebende nematoden aus Rumänien. Annales Universitatis Scientiarum Budapestinensis de Rolando Eotvos Nominatae. Sect. Biol. 1959, 2, 3–27. [Google Scholar]
- Bastian, H.C., II. Monograph on the Anguillulidæ, or Free Nematoids, Marine, Land, and Freshwater; with Descriptions of 100 New Species. Trans. Linn. Soc. Lond. 1865, 25, 73–184. [Google Scholar] [CrossRef]
- Goodey, J. Tylenchus (Cepha-lenchus) megacephalus n. sbg., n. sp. Nematologica 1962, 7, 331–333. [Google Scholar] [CrossRef]
- Verschoor, B.C.; de Goede, R.G.; de Hoop, J.-W.; de Vries, F.W. Seasonal dynamics and vertical distribution of plant-feeding nematode communities in grasslands. Pedobiologia 2001, 45, 213–233. [Google Scholar] [CrossRef]
- Qing, X.; Bert, W. Redefinition of genus Malenchus Andrássy, 1968 (Tylenchomorpha: Tylenchidae) with additional data on ecology. J. Nematol. 2017, 49, 189–206. [Google Scholar] [CrossRef] [Green Version]
- Azimi, S. Description of Basiria iranica sp. nov. (Nematoda: Tylenchidae) from southwestern Iran and its phylogenetic relationships. Ann. Zool. 2021, 71, 225–233. [Google Scholar] [CrossRef]
- Qiao, K.; Bai, M.; He, Y.; Chen, J.; Xiao, S.; Cheng, X.; Liu, G.; Braun-Miyara, S.; Qing, X. Description of Labrys fuzhouensis sp. n. and first record of Coslenchus rafiqi (Nematoda: Tylenchidae) from China. Nematology 2019, 21, 693–708. [Google Scholar] [CrossRef]
- Qing, X.; Pereira, T.J.; Slos, D.; Couvreur, M.; Bert, W. A new species of Malenchus (Nematoda: Tylenchomorpha) with an updated phylogeny of the Tylenchidae. Nematology 2018, 20, 815–836. [Google Scholar] [CrossRef]
- Forge, T.A.; Larney, F.J.; Kawchuk, L.M.; Pearson, D.C.; Koch, C.; Blackshaw, R.E. Crop rotation effects on Pratylenchus neglectus populations in the root zone of irrigated potatoes in southern Alberta. Can. J. Plant Pathol. 2015, 37, 363–368. [Google Scholar] [CrossRef]


| Characters | Canadian Population | Bajaj and Bhatti [44] |
|---|---|---|
| n * | 15 | 1 |
| Body length | 799.5 ± 58.2 (702.0–933.0) | 760 |
| a | 37.1 ± 3.2 (32.7–41.4) | 36 |
| b | 6.4 ± 0.3 (5.9–6.9) | 7 |
| c | 7.8 ± 0.5 (7.1–9.0) | 8 |
| c’ | 8.1 ± 0.6 (7.2–9.0) | 8 |
| V | 72.9 ± 0.9 (71.3–74.2) | 78 |
| MB | 47.2 ± 2.8 (42.0–52.0) | – |
| Lip height | 3.3 ± 0.3 (3.0–4.0) | 3–4 |
| Lip width | 6.6 ± 0.3 (6.0–7.0) | 6–8 |
| Stylet length | 11.9 ± 0.5 (11.0–13.0) | 12 |
| Pharynx length | 124.2 ± 6.5 (116.0–141.0) | – |
| Anterior end to excretory pore | 100.2 ± 7.7 (87.0–117.0) | 97–100 |
| Maximum body width | 21.7 ± 2.2 (17.0–25.0) | – |
| Vulva body width | 18.8 ± 1.5 (16.5–22.0) | – |
| Post-vulval uterine sac (PUS) length | 11.2 ± 2.0 (9.0–14.0) | – |
| Distance from vulva to anus | 114.2 ± 13.1 (95.0–150.0) | 80–92 |
| Distance from vulva to tail terminus | 216.5 ± 15.4 (191.0–254.0) | – |
| Anal body width | 12.7 ± 0.7 (11.0–14.0) | – |
| Tail length | 102.3 ± 5.8 (95.0–112.0) | 80 |
| Characters | Canadian Population | Andrássy [23] |
|---|---|---|
| n * | 19 | – |
| Body length | 517.3 ± 24.8 (466.0–567.0) | 460–540 |
| a | 31.8 ± 2.1 (27.5–35.0) | 23–29 |
| b | 5.7 ± 0.2 (5.4–6.2) | – |
| c | 4.6 ± 0.2 (4.4–5.0) | 5–6 |
| c’ | 12.2 ± 0.9 (10.1–14.6) | 8–11 |
| V | 64.0 ± 1.0 (62.2–65.9) | 64–67 |
| MB | 46.5 ± 2.0 (42.1–50.5) | 48 |
| G1 | 26.2 ± 3.9 (21.7–30.0) | – |
| Lip height | 3.2 ± 0.2 (3.0–3.5) | – |
| Lip width | 6.2 ± 0.2 (6.0–6.5) | 6–7 |
| Stylet length | 11.7 ± 0.7 (10.0–13.0) | 12 |
| Dorsal pharyngeal gland orifice (DGO) | 2.3 ± 0.4 (2.0–2.5) | – |
| Median bulb length | 11.2 ± 1.3 (10.0–13.5) | – |
| Median bulb width | 7.4 ± 0.6 (6.0–8.0) | |
| Pharynx length | 90.9 ± 2.7 (85.0–95.0) | 92–96 |
| Anterior end to excretory pore | 75.6 ± 2.8 (72.0–82.0) | – |
| Maximum body width | 16.3 ± 1.4 (14.0–19.0) | – |
| Vulva body width | 14.9 ± 1.3 (13.0–18.0) | – |
| Post-vulval uterine sac (PUS) length | 7.7 ± 1.2 (7.0–9.0) | – |
| Distance from vulva to anus | 74.5 ± 4.3 (65.0–81.0) | 79–92 |
| Distance from vulva to tail terminus | 185.9 ± 9.3 (168.0–204.0) | – |
| Anal body width | 9.2 ± 0.6 (8.0–10.0) | – |
| Tail length | 111.5 ± 6.9 (100.0–128.0) | 92–105 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munawar, M.; Yevtushenko, D.P.; Castillo, P. First Report of Three Tylenchidae Taxa from Southern Alberta, Canada. Horticulturae 2021, 7, 449. https://doi.org/10.3390/horticulturae7110449
Munawar M, Yevtushenko DP, Castillo P. First Report of Three Tylenchidae Taxa from Southern Alberta, Canada. Horticulturae. 2021; 7(11):449. https://doi.org/10.3390/horticulturae7110449
Chicago/Turabian StyleMunawar, Maria, Dmytro P. Yevtushenko, and Pablo Castillo. 2021. "First Report of Three Tylenchidae Taxa from Southern Alberta, Canada" Horticulturae 7, no. 11: 449. https://doi.org/10.3390/horticulturae7110449
APA StyleMunawar, M., Yevtushenko, D. P., & Castillo, P. (2021). First Report of Three Tylenchidae Taxa from Southern Alberta, Canada. Horticulturae, 7(11), 449. https://doi.org/10.3390/horticulturae7110449