Improved Propagation and Growing Techniques for Oleander Nursery Production



Abstract
:1. Introduction
2. Materials and Methods
2.1. Propagation Treatments
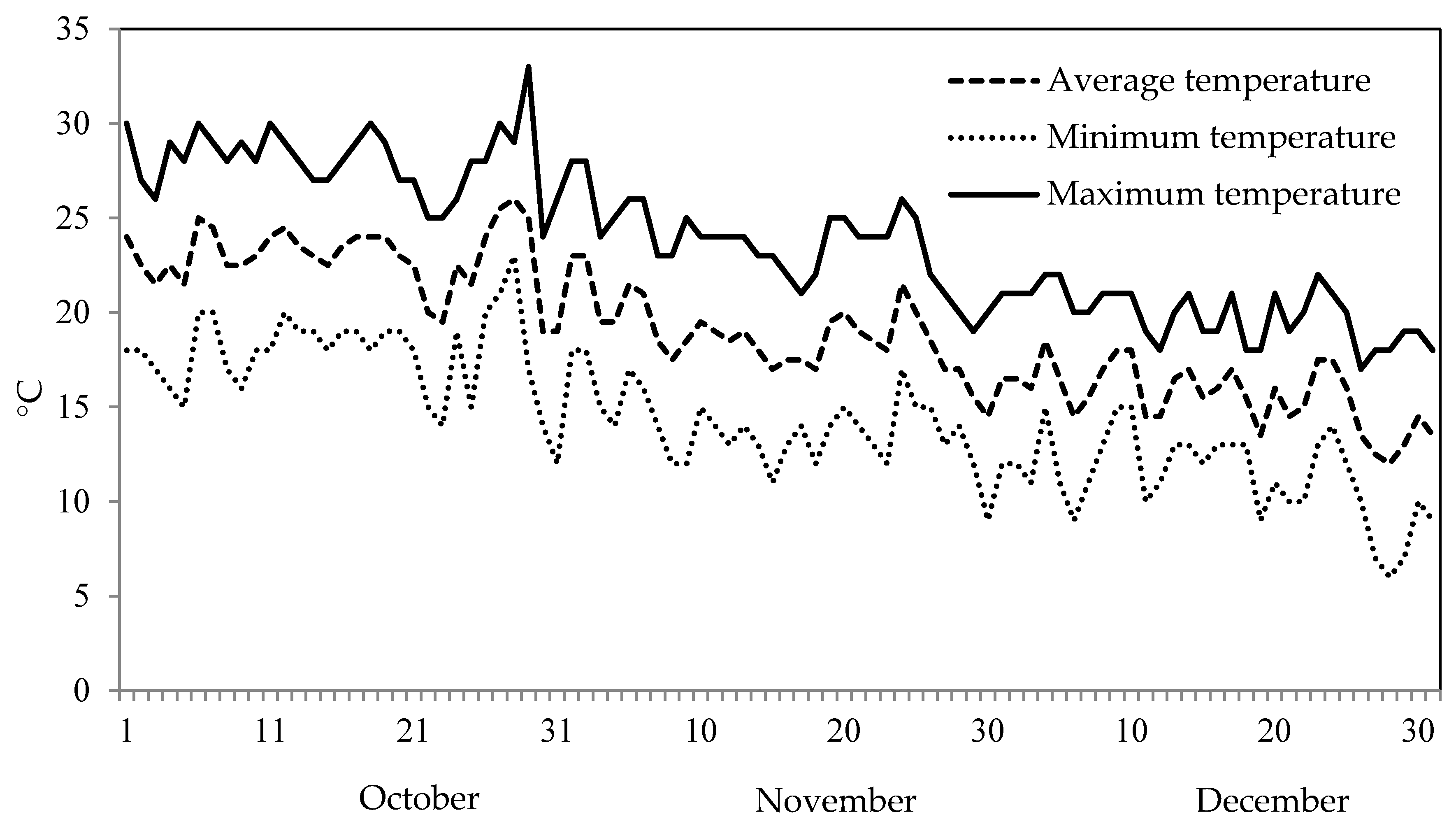
2.2. Forcing and Shading Treatments for Plant Nursery Production
2.3. Statistical Analysis
3. Results
3.1. Propagation Treatments
3.2. Forcing and Shading Treatments
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Franco, J.A.; Martínez-Sánchez, J.J.; Fernández, J.A.; Bañón, S. Selection and nursery production of ornamental plants for landscaping and xerogardening in semi-arid environments. J. Hortic. Sci. Biotechnol. 2006, 81, 3–17. [Google Scholar] [CrossRef]
- Eggenberger, R.; Eggenberger, M.E. The Handbook of Oleanders; Tropical Plant Specialists: Cleveland, GA, USA, 1996. [Google Scholar]
- Lenzi, A.; Pittas, L.; Martnelli, T.; Lombardi, P.; Tesi, R. Response to water stress of some oleander cultivars suitable forr pot plant production. Sci. Hortic. 2009, 122, 426–431. [Google Scholar] [CrossRef]
- Pilon, P. Perennial Solutions: A Grower’s Guide to Perennial Production, 1st ed.; Ball Publishing: Batavia, IL, USA, 2005; p. 546. [Google Scholar]
- Hartmann, H.D.; Kester, D.E.; Davies, F.J.; Geneve, R.L. Plant Propagation: Principles and Practices; Prentice Hall: Upper Saddle River, NJ, USA, 2003; p. 772. [Google Scholar]
- Toogood, A.R. Plant Propagation; DK Publishing Inc.: New York, NY, USA, 1999. [Google Scholar]
- Dirr, M.A.; Heuser, C.W. The Reference Manual of Woody Plant Propagation; Varsity Press: Athens, Greece, 2006. [Google Scholar]
- Ochoa, J.; Bañón, S.; Fernández, J.A.; Gonzalez, A.; Franco, J.A. Influence of cutting position and rooting media on rhizogenesis in oleander Cuttings. Acta Hort. 2003, 608, 101–106. [Google Scholar] [CrossRef]
- Ochoa, J.; Bañón, S.; Fernández, J.A.; Franco, J.A.; Martínez-Sánchez, J.J. Rooting Medium Temperature and Carbohydrates Affected Oleander Rooting. Acta Hort. 2004, 659, 239–244. [Google Scholar] [CrossRef]
- Smalley, T.J.; Dirr, M.A. Effect of cutting size on rooting and subsequent growth of Acer rubrum ‘Red Sunset’ cuttings. J. Environ. Hortic. 1987, 5, 122–124. [Google Scholar]
- Henry, F.A.; Blazich, P.H.; Hinesley, L.E. Vegetative propagation of Eastern redcedar by stem cuttings. HortScience 1992, 27, 1272–1274. [Google Scholar] [CrossRef]
- Hinesley, L.E.; Blazich, F.A.; Snelling, L.K. Propagation of Atlantic white cedar by stem cuttings. HortScience 1994, 29, 217–219. [Google Scholar] [CrossRef]
- Caruso, S.; Iapichino, G. Basal heat improves adventitious root quality in Plumeria (Plumeriarubra L.) stem cuttings of different sizes. J. Appl. Hortic. 2014, 17, 22–25. [Google Scholar]
- Beyl, C.A.; Ghale, G.; Zhang, L. Characteristics of hardwood cuttings influence rooting of Actinidia arguta (Siebold and Zucc.) Planch. HortScience 1995, 30, 973–976. [Google Scholar] [CrossRef]
- Blazich, F.A. Chemicals and formulations used to promote adventitious rooting. In Adventitious Root Formation in Cuttings; Davis, T.D., Haissig, B.E., Sankhla, N., Eds.; Dioscorides Press: Portland, OR, USA, 1988; pp. 132–149. [Google Scholar]
- Iapichino, G. Micropropagation of Globe Artichoke (Cynara cardunculus L. var. scolymus). In Protocols for Micropropagation of Selected Economically-Important Horticultural Plants, Methods in Molecular Biology; Lambardi, M., Ozudogru, E.A., Jain, S.M., Eds.; Springer: New York, NY, USA, 2013; Volume 11013, pp. 329–339. [Google Scholar]
- Iapichino, G.; Arnone, C.; Bertolino, M.; AmicoRoxas, U. Propagation of three Thymus by stem cuttings. Acta Hort. 2006, 723, 411–413. [Google Scholar] [CrossRef]
- Kieffer, M.; Fuller, M.P. In vitro propagation of cauliflower using curd microexplants. In Protocols for Micropropagation of Selected Economically-Important Horticultural Plants, Methods in Molecular Biology; Lambardi, M., Ozudogru, E.A., Jain, S.M., Eds.; Springer: New York, NY, USA, 2013; Volume 11013, pp. 369–380. [Google Scholar]
- Carpenter, W.J.; Cornell, J.A. Auxin application duration and concentration govern rooting of Hibiscus stem cuttings. J. Am. Soc. Hort. Sci. 1992, 117, 68–74. [Google Scholar] [CrossRef]
- Sabatino, L.; D’anna, F.; Iapichino, G. Cutting type and IBA treatment duration affect Teucrium fruticans adventitious root quality. Not. Bot. Horti Agrobot. Cluj Napoca 2014, 42, 478–481. [Google Scholar] [CrossRef]
- Kashefi, M.; Zarei, H.; Bahadori, F. The Effect of Indole Butyric Acid and the Time of Stem Cutting Preparation on Propagation of Damask Rose Ornamental Shrub. JOP 2014, 4, 237–243. [Google Scholar]
- Kaviani, B.; Gholami, S. Improvement of Rooting in Forsythia×intermedia Cuttings by Plant Growth Regulators. JOP 2016, 6, 125–131. [Google Scholar]
- Pacholczak, A. The effect of the auxin application methods on rooting of Physocarpus opulifolius Maxim. cuttings. Propag. Ornam. Plants 2015, 15, 147–153. [Google Scholar]
- Zhang, L.; Wang, S.; Guo, W.; Zhang, Y.; Shan, W.; Wang, K. Effect of Indole-3-Butyric Acid and rooting substrates on rooting response of hardwood cuttings of Rhododendron fortune Lindl. Propag. Ornam. Plants 2015, 15, 79–86. [Google Scholar]
- Cristofori, V.; Rouphael, Y.; Rugini, E. Collection time, cutting age, IBA and putrescine effects on root formation in Corylus avellana L. cuttings. Sci. Hortic. 2010, 124, 189–194. [Google Scholar] [CrossRef]
- Sebastiani, L.; Tognetti, R. Growing season and hydrogen peroxide effects on root induction and development in Olea europaea L. (cvs ‘frantoio’ and ‘gentile di larino’) cuttings. Sci. Hortic. 2004, 100, 75–82. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Treatments | Rooting (%) | No. of Roots Cutting−1 | Root Length (cm) | Shoot Length (cm) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| at 30 d | at 45 d | at 30 d | at 45 d | at 30 d | at 60 d | |||||||||
| Node Number (N) | ||||||||||||||
| 2 | 87.7 | b z | 7.8 | 15.9 | b | 11.8 | 12.0 | 6.8 | 8.5 | |||||
| 3 | 94.1 | a | 8.8 | 22.3 | a | 10.4 | 12.2 | 7.0 | 8.9 | |||||
| 4 | 91.9 | a | 8.9 | 21.1 | a | 12.2 | 14.0 | 8.2 | 8.6 | |||||
| IBA | ||||||||||||||
| 0 ppm | 83.5 | b | 6.9 | b | 16.4 | b | 10.4 | 13.1 | 5.9 | b | 8.4 | |||
| 3000 ppm | 93.0 | a | 10.0 | a | 23.2 | a | 12.5 | 12.4 | 8.1 | a | 8.8 | |||
| Significance | ||||||||||||||
| N | **y | NS | ** | NS | NS | NS | NS | |||||||
| IBA | * | ** | *** | NS | NS | *** | NS | |||||||
| N × IBA | NS | NS | NS | NS | NS | NS | NS | |||||||
| Treatments | Rooting (%) | No. of Roots Cutting−1 | Root Length (cm) | Shoot Length (cm) | No. of Leaves Cutting−1 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| at 30 d | at 45 d | at 30 d | at 45 d | at 30 d | at 60 d | at 30 d | at 60 d | |||||||||||
| Clones | ||||||||||||||||||
| Clone 1 | 93.8 | a z | 8.8 | ab | 22.3 | a | 10.4 | b | 12.1 | 7.0 | 8.8 | b | 16.3 | b | 28.9 | a | ||
| Clone 2 | 90.1 | b | 10.2 | a | 14.2 | bc | 14.7 | a | 13.5 | 8.2 | 10.4 | a | 19.0 | ab | 22.4 | b | ||
| Clone 3 | 87.5 | b | 10.2 | a | 16.5 | b | 11.6 | ab | 12.4 | 9.5 | 11.0 | a | 22.1 | a | 23.0 | b | ||
| Clone 4 | 52.1 | d | 6.3 | b | 7.3 | d | 8.4 | b | 13.3 | 7.4 | 8.8 | b | 15.8 | b | 23.7 | b | ||
| Clone 5 | 71.8 | c | 7.4 | ab | 9.3 | cd | 11.4 | ab | 10.8 | 8.9 | 9.8 | ab | 19.0 | ab | 22.9 | b | ||
| IBA | ||||||||||||||||||
| 0 ppm | 78.3 | 7.3 | b | 12.9 | b | 11.8 | 12.3 | 7.6 | 9.4 | b | 19.5 | 26.3 | a | |||||
| 3000 ppm | 79.8 | 9.9 | a | 15.3 | a | 10.8 | 12.5 | 8.8 | 10 | a | 17.4 | 22.0 | b | |||||
| Significance | ||||||||||||||||||
| Clones | ***y | * | *** | ** | NS | NS | *** | ** | *** | |||||||||
| IBA | NS | ** | * | NS | NS | NS | ** | NS | *** | |||||||||
| Clones × IBA | ** | NS | NS | NS | NS | NS | NS | * | ** | |||||||||
| Treatments | Rooting (%) | No. of Leaves Cutting−1 at 30 Days | No. of Leaves Cutting−1 at 60 Days | |||
|---|---|---|---|---|---|---|
| Clone 1 × 0 ppm | 87.5 | c z | 14.5 | c | 28.6 | a |
| Clone 1 × 3000 ppm | 95.8 | a | 18.0 | ab | 29.2 | a |
| Clone 2 × 0 ppm | 91.7 | b | 21.8 | ab | 24.9 | b |
| Clone 2 × 3000 ppm | 92.7 | b | 16.3 | b | 19.9 | c |
| Clone 3 × 0 ppm | 85.4 | c | 23.5 | a | 24.2 | b |
| Clone 3 × 3000 ppm | 89.6 | bc | 20.8 | ab | 21.8 | bc |
| Clone 4 × 0 ppm | 56.3 | e | 19.0 | ab | 27.6 | a |
| Clone 4 × 3000 ppm | 47.9 | f | 12.5 | c | 19.8 | c |
| Clone 5 × 0 ppm | 70.8 | d | 18.5 | b | 26.3 | b |
| Clone 5 × 3000 ppm | 72.9 | d | 19.5 | b | 19.5 | c |
| Treatments | Plant Height (cm) | Beginning of Flowering (Days from 1 January) | Full Flowering (Days from 1 January) | |||
|---|---|---|---|---|---|---|
| Not-forced × U | 82.3 | d z | 122.7 | a | 142.0 | a |
| October forcing × U | 73.0 | e | 69.0 | e | 85.0 | g |
| November forcing × U | 95.0 | c | 78.3 | d | 112.7 | e |
| December forcing × U | 97.7 | c | 111.7 | b | 130.0 | c |
| Not-forced × S | 98.0 | c | 120.0 | a | 136.0 | b |
| October forcing × S | 116.3 | a | 71.0 | e | 96.7 | f |
| November forcing × S | 103.0 | b | 79.3 | d | 95.0 | f |
| December forcing × S | 105.0 | b | 99.0 | c | 117.0 | d |
| Significance | ||||||
| Forcing (F) | ***y | *** | *** | |||
| Shading (S) | ** | * | NS | |||
| F × S | *** | * | *** | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabatino, L.; D’Anna, F.; Iapichino, G. Improved Propagation and Growing Techniques for Oleander Nursery Production. Horticulturae 2019, 5, 55. https://doi.org/10.3390/horticulturae5030055
Sabatino L, D’Anna F, Iapichino G. Improved Propagation and Growing Techniques for Oleander Nursery Production. Horticulturae. 2019; 5(3):55. https://doi.org/10.3390/horticulturae5030055
Chicago/Turabian StyleSabatino, Leo, Fabio D’Anna, and Giovanni Iapichino. 2019. "Improved Propagation and Growing Techniques for Oleander Nursery Production" Horticulturae 5, no. 3: 55. https://doi.org/10.3390/horticulturae5030055
APA StyleSabatino, L., D’Anna, F., & Iapichino, G. (2019). Improved Propagation and Growing Techniques for Oleander Nursery Production. Horticulturae, 5(3), 55. https://doi.org/10.3390/horticulturae5030055