Risk of Human Pathogen Internalization in Leafy Vegetables During Lab-Scale Hydroponic Cultivation

Abstract
1. Introduction
Review Methodology
2. Controlled Environment Agriculture (CEA) and Food Safety
3. Pathogen Internalization in Leafy Vegetables
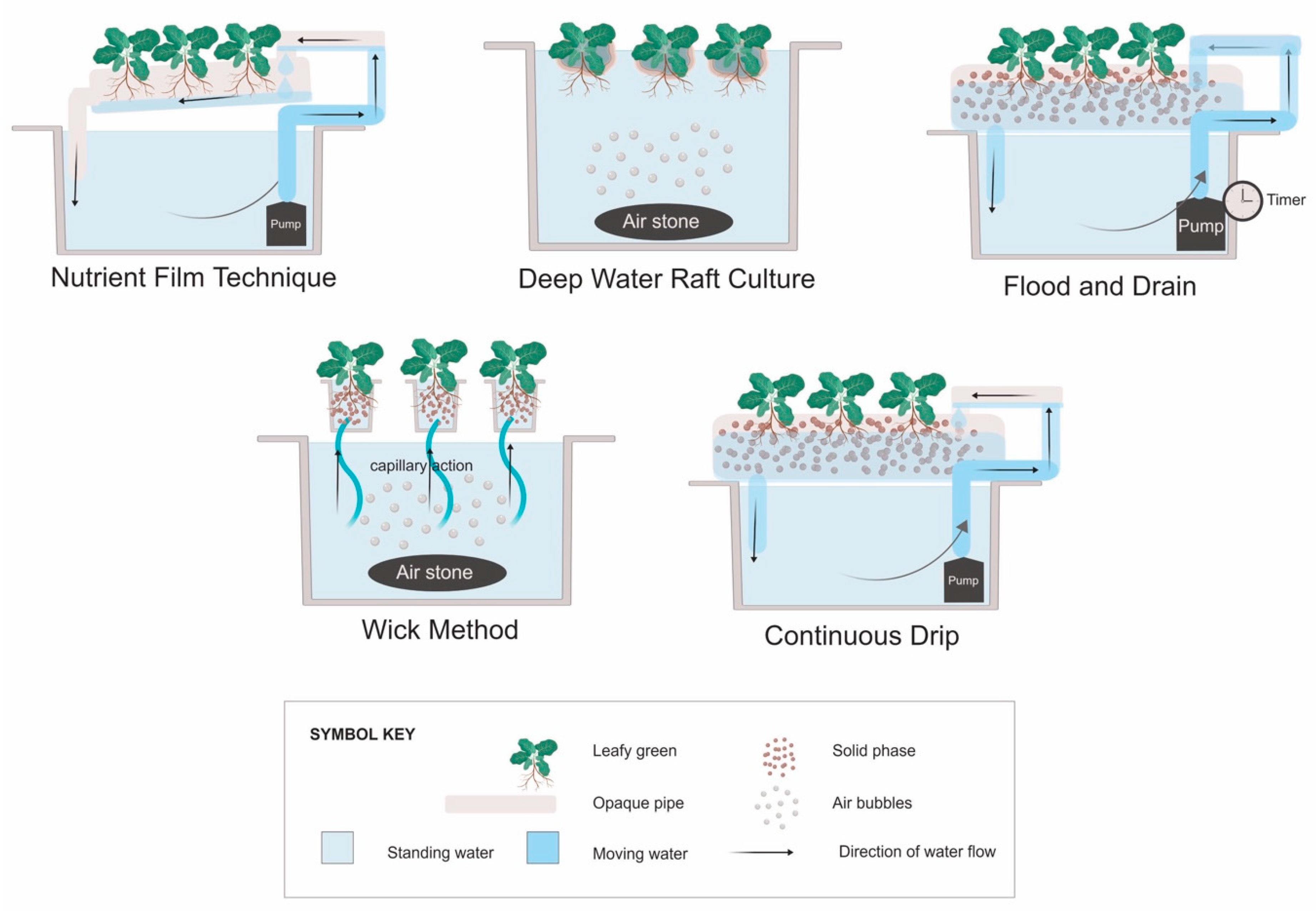
4. Hydroponic System Designs
5. Pathogen Internalization in Hydroponic Systems
5.1. Deep Water Culture
5.2. Nutrient Film Technique
5.3. Other Hydroponic Systems
6. Targeted Preventive Controls in Hydroponic Systems for Leafy Vegetables
6.1. Production Water Quality and Whole System Decontamination
6.1.1. Current Agricultural Water Quality Guidelines for Fresh Produce
6.1.2. Risk of System Contamination
6.1.3. Water Treatment Strategies
6.2. Minimizing Root Damage
6.3. Biological Control
6.4. Plant Cultivar Selection
7. Potential Actual Health Risk from Consumption of Leafy Vegetables with Internalized Pathogens
8. Conclusions
- Development of standard guidelines for lab-scale hydroponic cultivation of leafy vegetables to enable study comparison. This includes seed germination protocols, best practices for water management, and design specifications for each type of hydroponic system.
- Determine appropriate pathogen inoculation concentrations and methods for the research question being addressed. Should there be a range of concentrations considered? How does the inoculation of the seed at germination versus inoculation of the nutrient solution change the interpretation of the results?
- Does the presence of a solid substrate impact colonization efficiency? Is there a differential effect between contamination of the substrate and the contamination of nutrient water flowing through it?
- Standardization of microbial extraction methods from plants to ensure the recovery of truly endophytic microorganisms.
- Selection of microorganisms should be standardized. For instance, surrogate microorganisms should be validated as representative of their human pathogen counterparts. Strains of human pathogens should also be carefully considered and validated for use in hydroponic cultivation systems.
- Given the variation in the susceptibility of plants to pathogen colonization, the selection of plant cultivars should be standardized to represent commercially relevant cultivars, and the validation of cultivars used in hydroponic research is needed.
Author Contributions
Acknowledgements
Conflicts of Interest
References
- California Leafy Green Products Handler Marketing Agreement (LGMA). Farming Leafy Greens. 2019. Available online: https://lgma.ca.gov/about-us/farming-leafy-greens/ (accessed on 9 January 2019).
- Centers for Disease Control and Prevention (CDC). Multistate Outbreak of E. coli O157:H7 Infections Linked to Romaine Lettuce (Final Update)—Case Count Maps. 2018. Available online: https://www.cdc.gov/ecoli/2018/o157h7-04-18/map.html (accessed on 9 January 2019).
- Centers for Disease Control and Prevention (CDC). Multistate Outbreak of Shiga toxin-producing Escherichia coli O157:H7 Infections Linked to Leafy Greens (Final Update)—Case Count Maps. 2018. Available online: https://www.cdc.gov/ecoli/2017/o157h7-12-17/map.html (accessed on 9 January 2019).
- Centers for Disease Control and Prevention (CDC). Outbreak of E. coli Infections Linked to Romaine Lettuce—Timeline of Reported Cases. 2018. Available online: https://www.cdc.gov/ecoli/2018/o157h7-11-18/epi.html (accessed on 9 January 2019).
- Herman, K.M.; Hall, A.J.; Gould, L.H. Outbreaks attributed to fresh leafy vegetables, United States, 1973-2012. Epidemiol. Infect. 2015, 143, 3011–3021. [Google Scholar] [CrossRef] [PubMed]
- Agrylist. State of Indoor Farming. 2017. Available online: https://www.agrilyst.com/stateofindoorfarming2017/ (accessed on 12 March 2019).
- Erickson, M.C. Internalization of fresh produce by foodborne pathogens. Annu. Rev. Food Sci. Technol. 2012, 3, 283–310. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.H. Hydroponics Worldwide. Acta Hortic. 1999, 481, 719–729. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; Saifullah, M.; Mollick, A.K.M.S.R.; Hossain, M.M.; Halim, G.M.A.; Asao, T. Influence of Soilless Culture Substrate on Improvement of Yield and Produce Quality of Horticultural Crops. In Soilless Culture—Use of Substrates for the Production of Quality Horticultural Crops; InTech: London, UK, 2015. [Google Scholar]
- Agrylist. State of Indoor Farming. 2016. Available online: https://www.agrilyst.com/stateofindoorfarming2016/ (accessed on 9 January 2019).
- Harrison, J.A.; Gaskin, J.W.; Harrison, M.A.; Cannon, J.L.; Boyer, R.R.; Zehnder, G.W. Survey of food safety practices on small to medium-sized farms and in farmers markets. J. Food Prot. 2013, 76, 1989–1993. [Google Scholar] [CrossRef] [PubMed]
- Behnke, C.; Seo, S.; Miller, K. Assessing food safety practices in farmers’ markets. Food Prot. Trends 2012, 32, 232–239. [Google Scholar]
- De Keuckelaere, A.; Jacxsens, L.; Amoah, P.; Medema, G.; Mcclure, P.; Jaykus, L.A.; Uyttendaele, M. Zero risk does not exist: Lessons learned from microbial risk assessment related to use of water and safety of fresh produce. Compr. Rev. Food Sci. Food Saf. 2015, 14, 387–410. [Google Scholar] [CrossRef]
- Stamper, C.E.; Hoisington, A.J.; Gomez, O.M.; Halweg-Edwards, A.L.; Smith, D.G.; Bates, K.L.; Kinney, K.A.; Postolache, T.T.; Brenner, L.A.; Rook, G.A.W.; et al. The microbiome of the built environment and human behavior: Implications for emotional health and well-being in postmodern western societies. Int. Rev. Neurobiol. 2016, 131, 289–323. [Google Scholar] [CrossRef]
- Todd, E.C.; Greig, J.D.; Bartleson, C.A.; Michaels, B.S. Outbreaks where food workers have been implicated in the spread of foodborne disease. Part 5. Sources of contamination and pathogen excretion from infected persons. J. Food Prot. 2008, 71, 2582–2595. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.J.; Farber, J.N.; Beuchat, L.R.; Parish, M.E.; Suslow, T.V.; Garrett, E.H.; Busta, F.F. Outbreaks associated with fresh produce: Incidence, growth, and survival of pathogens in fresh and fresh-cut produce. Compr. Rev. Food Sci. 2003, 2, 78–141. [Google Scholar] [CrossRef]
- Heaton, J.C.; Jones, K. Microbial contamination of fruit and vegetables and the behaviour of enteropathogens in the phyllosphere: A review. J. Appl. Microbiol. 2008, 104, 613–626. [Google Scholar] [CrossRef]
- Berger, C.N.; Sodha, S.V.; Shaw, R.K.; Griffin, P.M.; Pink, D.; Hand, P.; Frankel, G. Fresh fruit and vegetables as vehicles for the transmission of human pathogens. Environ. Microbiol. 2010, 12, 2385–2397. [Google Scholar] [CrossRef] [PubMed]
- Wilks, S.A.; Michels, H.; Keevil, C.W. The survival of Escherichia coli O157 on a range of metal surfaces. Int. J. Food Microbiol. 2005, 105, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Lopman, B.; Gastanaduy, P.; Park, G.W.; Hall, A.J.; Parashar, U.D.; Vinjé, J. Environmental transmission of norovirus gastroenteritis. Curr. Opin. Virol. 2012, 2, 96–102. [Google Scholar] [CrossRef]
- Zhu, Q.; Gooneratne, R.; Hussain, M.A. Listeria monocytogenes in fresh produce: Outbreaks, prevalence and contamination levels. Foods 2017, 6, 21. [Google Scholar] [CrossRef]
- Hara-Kudo, Y.; Konuma, H.; Iwaki, M.; Kasuga, F.; Sugita-Konishi, Y.; Ito, Y.; Kumagai, S. Potential hazard of radish sprouts as a vehicle of Escherichia coli O157:H7. J. Food Prot. 1997, 60, 1125–1127. [Google Scholar] [CrossRef]
- Gutierrez, E. Is the Japanese O157:H7 E. coli epidemic over? Lancet 1996, 348, 1371. [Google Scholar] [CrossRef]
- Itoh, Y.; Sugita-Konishi, Y.; Kasuga, F.; Iwaki, M.; Hara-Kudo, Y.; Saito, N.; Noguchi, Y.; Konuma, H.; Kumagai, S. Enterohemorrhagic Escherichia coli O157:H7 present in radish sprouts. Appl. Environ. Microbiol. 1998, 64, 1532–1535. [Google Scholar]
- Seo, K.H.; Frank, J.F. Attachment of Escherichia coli O157:H7 to lettuce leaf surface and bacterial viability in response to chlorine treatment as demonstrated by using confocal scanning laser microscopy. J. Food Prot. 1999, 62, 3–9. [Google Scholar] [CrossRef]
- Solomon, E.B.; Yaron, S.; Matthews, K.R. Transmission of Escherichia coli O157:H7 from contaminated manure and irrigation water to lettuce plant tissue and its subsequent internalization. Appl. Environ. Microbiol. 2002, 68, 397–400. [Google Scholar] [CrossRef]
- Gil, M.I.; Selma, M.V.; López-Gálvez, F.; Allende, A. Fresh-cut product sanitation and wash water disinfection: Problems and solutions. Int. J. Food Microbiol. 2009, 134, 37–45. [Google Scholar] [CrossRef]
- Jablasone, J.; Warriner, K.; Griffiths, M. Interactions of Escherichia coli O157:H7, Salmonella Typhimurium and Listeria monocytogenes plants cultivated in a gnotobiotic system. Int. J. Food Microbiol. 2005, 99, 7–18. [Google Scholar] [CrossRef]
- Bernstein, N.; Sela, S.; Pinto, R.; Ioffe, M. Evidence for internalization of Escherichia coli into the aerial parts of maize via the root system. J. Food Prot. 2007, 70, 471–475. [Google Scholar] [CrossRef]
- Hirneisen, K.A.; Sharma, M.; Kniel, K.E. Human enteric pathogen internalization by root uptake into food crops. Foodborne Pathog. Dis. 2012, 9, 396–405. [Google Scholar] [CrossRef]
- Franz, E.; Visser, A.A.; Van Diepeningen, A.D.; Klerks, M.M.; Termorshuizen, A.J.; van Bruggen, A.H.C. Quantification of contamination of lettuce by GFP-expressing Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium. Food Microbiol. 2007, 24, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Ingram, D.T.; Patel, J.R.; Millner, P.D.; Wang, X.; Hull, A.E.; Donnenberg, M.S. A novel approach to investigate the uptake and internalization of Escherichia coli O157:H7 in spinach cultivated in soil and hydroponic medium. J. Food Prot. 2009, 72, 1513–1520. [Google Scholar] [CrossRef]
- Warriner, K.; Ibrahim, F.; Dickinson, M.; Wright, C.; Waites, W.M. Internalization of human pathogens within growing salad vegetables. Biotechnol. Genet. Eng. Rev. 2003, 20, 117–136. [Google Scholar] [CrossRef] [PubMed]
- Deering, A.J.; Mauer, L.J.; Pruitt, R.E. Internalization of E. coli O157:H7 and Salmonella spp. in plants: A review. Food Res. Int. 2012, 45, 567–575. [Google Scholar] [CrossRef]
- Beuchat, L.R. Vectors and conditions for preharvest contamination of fruits and vegetables with pathogens capable of causing enteric diseases. Br. Food J. 2006, 108, 38–53. [Google Scholar] [CrossRef]
- Resh, H.M. Hydroponic Food Production: A Definitive Guidebook for the Advanced Home Gardener and the Commercial Hydroponic Grower, 7th ed.; CRC Press Taylor and Francis Group: Boca Raton, FL, USA, 2012; ISBN 9781439878675. [Google Scholar]
- Ferrarezi, R.S.; Testezlaf, R. Performance of wick irrigation system using self-compensating troughs with substrates for lettuce production. J. Plant Nutr. 2016, 39, 147–161. [Google Scholar] [CrossRef]
- Hayden, A.L.; Giacomelli, G.A.; Hoffmann, J.J.; Yokelson, T.N. Aeroponics: An alternative production system for high-value root crops. Acta Hortic. 2004, 629, 207–213. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO) of the United Nations. Good Agricultural Practices for Greenhouse Vegetable Crops: Principles for Mediterranean Climate Areas; FAO: Rome, Italy, 2013. [Google Scholar]
- Brechner, M.; Both, A.J. Cornell University Hydroponic Lettuce Handbook; Cornell University Controlled Environment Agriculture: Ithaca, NY, USA, 2013; Available online: http://cea.cals.cornell.edu/attachments/Cornell CEA Lettuce Handbook.pdf (accessed on 9 January 2019).
- University of Kentucky Cooperative Extension Service, New Crop Opportunities Center, 2006. Hydroponic Lettuce. Available online: https://www.hort.vt.edu/ghvegetables/documents/Leafy Greens/GH-grown Lettuce and Greens_UnivKentucky_2006.pdf (accessed on 9 January 2019).
- Nolte, K.D. Winter Lettuce Production: Yuma, Arizona. Powerpoint Presentation. n.d. University of Arizona College of Agriculture and the Life Sciences Available online: https://cals.arizona.edu/fps/sites/cals.arizona.edu.fps/files/Lettuce Production Presentation.pdf (accessed on 9 January 2019).
- Kerns, D.L.; Matheron, M.E.; Palumbo, J.C.; Sanchez, C.A.; Still, D.W.; Tickes, B.R.; Umeda, K.; Wilcox, M.A. Guidelines for Head Lettuce Production in Arizona. IPM Series Number 12. Publication number az1099; Cooperative Extension, College of Agriculture and Life Sciences, University of Arizona: Tucson, AZ, USA, February 1999; Available online: http://cals.arizona.edu/crops/vegetables/cropmgt/az1099.html (accessed on 9 January 2019).
- Smith, R.; Cahn, M.; Daugovish, O.; Koike, S.; Natwick, E.; Smith, H.; Subbarao, K.; Takele, E.; Turini, T. Leaf Lettuce Production in California Publication 7216. University of California Davis. UC Vegetable Resource and Information Center. University of California Agriculture and Natural Resources. 2011. Available online: https://anrcatalog.ucanr.edu/pdf/7216.pdf (accessed on 9 January 2019).
- Kaiser, C.; Ernst, M. Romaine Lettuce. Center for Crop Diversification Crop Profile, CD-CCP-116; University of Kentucky College of Agriculture, Food, and the Environment Cooperative Extension: Lexington, KY, USA, 2017; Available online: http://www.uky.edu/ccd/sites/www.uky.edu.ccd/files/romaine.pdf (accessed on 9 January 2019).
- Kutter, S.; Hartmann, A.; Schmid, M. Colonization of barley (Hordeum vulgare) with Salmonella enterica and Listeria spp. FEMS Microbiol. Ecol. 2006, 56, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Koseki, S.; Mizuno, Y.; Yamamoto, K. Comparison of two possible routes of pathogen contamination of spinach leaves in a hydroponic cultivation system. J. Food Prot. 2011, 74, 1536–1542. [Google Scholar] [CrossRef] [PubMed]
- Macarisin, D.; Patel, J.; Sharma, V.K. Role of curli and plant cultivation conditions on Escherichia coli O157: H7 internalization into spinach grown on hydroponics and in soil. Int. J. Food Microbiol. 2014, 173, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Carducci, A.; Caponi, E.; Ciurli, A.; Verani, M. Possible internalization of an enterovirus in hydroponically grown lettuce. Int. J. Environ. Res. Public Health 2015, 12, 8214–8227. [Google Scholar] [CrossRef] [PubMed]
- Warriner, K.; Ibrahim, F.; Dickinson, M.; Wright, C.; Waites, W.M. Interaction of Escherichia coli with growing salad spinach plants. J. Food Protect. 2003, 66, 1790–1797. [Google Scholar] [CrossRef]
- Wang, Q.; Kniel, K.E. Survival and transfer of murine norovirus within a hydroponic system during kale and mustard microgreen harvesting. Appl. Environ. Microbiol. 2016, 82, 705–713. [Google Scholar] [CrossRef]
- Li, D.; Uyttendaele, M. Potential of human norovirus surrogates and Salmonella enterica contamination of pre-harvest basil (Ocimum basilicum) via leaf surface and plant substrate. Front. Microbiol. 2018, 9, 1728. [Google Scholar] [CrossRef]
- Settanni, L.; Miceli, A.; Francesca, N.; Cruciata, M.; Moschetti, G. Microbiological investigation of Raphanus sativus L. grown hydroponically in nutrient solutions contaminated with spoilage and pathogenic bacteria. Int. J. Food Microbiol. 2013, 160, 344–352. [Google Scholar] [CrossRef]
- Dong, Y.; Iniguez, A.L.; Ahmer, B.M.M.; Triplett, E.W. Kinetics and strain specificity of rhizosphere and endophytic colonization by enteric bacteria on seedlings of Medicago sativa and Medicago truncatula. Appl. Environ. Microbiol. 2003, 69, 1783–1790. [Google Scholar] [CrossRef]
- Klerks, M.M.; Franz, E.; Van Gent-Pelzer, M.; Zijlstra, C.; van Bruggen, A.H.C. Differential interaction of Salmonella enterica serovars with lettuce cultivars and plant-microbe factors influencing the colonization efficiency. ISME J. 2007, 1, 620–631. [Google Scholar] [CrossRef]
- DiCaprio, E.; Ma, Y.; Purgianto, A.; Hughes, J.; Li, J. Internalization and dissemination of human norovirus and animal caliciviruses in hydroponically grown romaine lettuce. Appl. Environ. Microbiol. 2012, 78, 6143–6152. [Google Scholar] [CrossRef]
- Moriarty, M.J.; Semmens, K.; Bissonnette, G.K.; Jaczynski, J. Internalization assessment of E. coli O157:H7 in hydroponically grown lettuce. LWT Food Sci. Technol. 2019, 100, 183–188. [Google Scholar] [CrossRef]
- Moriarty, M.J.; Semmens, K.; Bissonnette, G.K.; Jaczynski, J. Inactivation with UV-radiation and internalization assessment of coliforms and Escherichia coli in aquaponically grown lettuce. LWT Food Sci. Technol. 2018, 89, 624–630. [Google Scholar] [CrossRef]
- Sharma, N.; Acharya, S.; Kumar, K.; Singh, N.; Chaurasia, O.P. Hydroponics as an advanced technique for vegetable production: An overview. J. Soil Water Conserv. 2019, 17, 364. [Google Scholar] [CrossRef]
- Barnhart, M.M.; Chapman, M.R. Curli biogenesis and function. Annu. Rev. Microbiol. 2016, 60, 131–147. [Google Scholar] [CrossRef]
- Barak, J.D.; Gorski, L.; Naraghi-Arani, P.; Charowski, A.O. Salmonella enterica virulence genes are required for bacterial attachment to plant tissue. Appl. Environ. Microbiol. 2005, 7, 5685–5691. [Google Scholar] [CrossRef]
- Higgins, C. (Hort Americas, LLC, Bedford, TX, USA). Personal communication, 17 January 2019.
- Mähönen, A.P.; ten Tusscher, K.; Siligato, R.; Smetana, O.; Díaz-Triviño, S.; Salojärvi, J.; Wachsman, G.; Prasad, K.; Heidstra, R.; Scheres, B. PLETHORA gradient formation mechanism separates auxin responses. Nature 2014, 515, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Allende, A.; Monaghan, J. Irrigation water quality for leaf crops: A perspective of risks and potential solutions. Int. J. Environ. Res. Public Health. 2015, 12, 7457–7477. [Google Scholar] [CrossRef] [PubMed]
- Waechter-Kristensen, B.; Sundin, P.; Gertsson, U.E.; Hultberg, M.; Khalil, S.; Jensén, P.; Berkelmann-Loehnertz, B.; Wohanka, W. Management of microbial factors in the rhizosphere and nutrient solution of hydroponically grown tomato. Acta Hortic. 1997, 450, 335–342. [Google Scholar] [CrossRef]
- Lopez-Galvez, F.; Allende, A.; Pedrero-Salcedo, F.; Alarcon, J.J.; Gil, M.I. Safety assessment of greenhouse hydroponic tomatoes irrigated with reclaimed and surface water. Int. J. Food Microbiol. 2014, 191, 97–102. [Google Scholar] [CrossRef]
- Ohtani, T.; Kaneko, A.; Fukuda, N.; Hagiwara, S.; Sase, S. SW—Soil and water. Development of a membrane disinfection system for closed hydroponics in a greenhouse. J. Agric. Eng. Res. 2000, 77, 227–232. [Google Scholar] [CrossRef]
- Moens, M.; Hendrickx, G. Drain water filtration for the control of nematodes in hydroponic-type systems. Crop Prot. 1992, 11, 69–73. [Google Scholar] [CrossRef]
- Wohanka, W.; Luedtke, H.; Ahlers, H.; Luebke, M. Optimization of slow filtration as a means for disinfecting nutrient solutions. Acta Hortic. 1999, 539–544. [Google Scholar] [CrossRef]
- van Os, E.A.; Postma, J. Prevention of root diseases in closed soilless growing systems by microbial optimisation and slow sand filtration. Acta Hortic. 2000, 97–102. [Google Scholar] [CrossRef]
- van Os, E.A. Design of sustainable hydroponic systems in relation to environment-friendly disinfection methods. Acta Hortic. 2001, 197–206. [Google Scholar] [CrossRef]
- Gratzek, J.B.; Gilbert, J.P.; Lohr, A.L.; Shotts, E.B.; Brown, J. Ultraviolet light control of Ichthyophthirius multifiliis Fouquet in a closed fish culture recirculation system. J. Fish Dis. 1983, 6, 145–153. [Google Scholar] [CrossRef]
- Wohanka, W. Slow sand filtration and UV radiation: Low-cost techniques for disinfection of recirculating nutrient solution or surface water. In Proceedings of the 8th International Congress on Soilless Culture, Rustenburg, Hunter’s Rest, South Africa, 2–9 October 1992; ISOSC: Wageningen, The Netherlands, 1993; pp. 497–511. [Google Scholar]
- Zhang, W.; Tu, J.C. Effect of Ultraviolet disinfection of hydroponic solutions on pythium root rot and non-target bacteria. Eur. J. Plant Pathol. 2000, 106, 415–421. [Google Scholar] [CrossRef]
- Ohashi-Kaneko, K.; Yoshii, M.; Isobe, T.; Park, J.-S.; Kurata, K.; Fujiwara, K. Nutrient solution prepared with ozonated water does not damage early growth of hydroponically grown tomatoes. Ozone Sci. Eng. 2009, 31, 21–27. [Google Scholar] [CrossRef]
- Pagliaccia, D.; Ferrin, D.; Stanghellini, M.E. Chemo-biological suppression of root-infecting zoosporic pathogens in recirculating hydroponic systems. Plant Soil 2007, 299, 163–179. [Google Scholar] [CrossRef]
- Runia, W.T. A review of possibilities for disinfection of recirculation water from soilless cultures. Acta Hortic. 1995, 221–229. [Google Scholar] [CrossRef]
- van Os, E.A.; Bruins, M.; Wohanka, W.; Seidel, R. Slow filtration: A technique to minimise the risks of spreading root-infecting pathogens in closed hydroponic systems. Acta Hortic. 2001, 495–502. [Google Scholar] [CrossRef]
- Wei, C.; Zhang, F.; Hu, Y.; Feng, C.; Wu, H. Ozonation in water treatment: The generation, basic properties of ozone and its practical application. Rev. Chem. Eng. 2017, 33, 49–89. [Google Scholar] [CrossRef]
- Guo, X.; van Iersel, M.W.; Chen, J.; Brackett, R.E.; Beuchat, L.R. Evidence of association of salmonellae with tomato plants grown hydroponically in inoculated nutrient solution. Appl. Environ. Microbiol. 2002, 30, 7–14. [Google Scholar] [CrossRef]
- Lee, S.; Lee, J. Beneficial bacteria and fungi in hydroponic systems: Types and characteristics of hydroponic food production methods. Sci. Hortic. 2015, 195, 206–215. [Google Scholar] [CrossRef]
- Lee, S.; An, R.; Grewal, P.; Yu, Z.; Borherova, Z.; Lee, J. High-performing windowfarm hydroponic system: Transcriptomes of fresh produce and microbial communities in response to beneficial bacterial treatment. Mol. Plant-Microbe Interact. 2016, 29, 965–976. [Google Scholar] [CrossRef]
- Moruzzi, S.; Firrao, G.; Polano, C.; Borselli, S.; Loschi, A.; Ermacora, P.; Loi, N.; Martini, M. Genomic-assisted characterisation of Pseudomonas sp. strain Pf4, a potential biocontrol agent in hydroponics. Biocontrol Sci. Technol. 2017, 27, 969–991. [Google Scholar] [CrossRef]
- Thongkamngam, T.; Jaenaksorn, T. Fusarium oxysporum (F221-B) as biocontrol agent against plant pathogenic fungi in vitro and in hydroponics. Plant Prot. Sci. 2017, 53, 85–95. [Google Scholar] [CrossRef]
- Fujiwara, K.; Iida, Y.; Iwai, T.; Aoyama, C.; Inukai, R.; Ando, A.; Ogawa, J.; Ohnishi, J.; Terami, F.; Takano, M.; et al. The rhizosphere microbial community in a multiple parallel mineralization system suppresses the pathogenic fungus Fusarium oxysporum. Microbiolopen 2013, 2, 997–1009. [Google Scholar] [CrossRef]
- Radzki, W.; Gutierrez Mañero, F.J.; Algar, E.; Lucas García, J.A.; García-Villaraco, A.; Ramos Solano, B. Bacterial siderophores efficiently provide iron to iron-starved tomato plants in hydroponics culture. Antonie van Leeuwenhoek. 2013, 104, 321–330. [Google Scholar] [CrossRef]
- Giurgiu, R.M.; Dumitras, A.; Morar, G.; Scheewe, P.; Schröder, F.G. A study on the biological control of Fusarium oxysporum using Trichoderma app., on soil and rockwool substrates in controlled environment. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2018, 46, 260–269. [Google Scholar] [CrossRef]
- Sirakov, I.; Lutz, M.; Graber, A.; Mathis, A.; Staykov, Y.; Smits, T.H.M.; Junge, R. Potential for combined biocontrol activity against fungal fish and plant pathogens by bacterial isolates from a model aquaponic system. Water 2016, 8, 518. [Google Scholar] [CrossRef]
- Erickson, M.C.; Liao, J.-Y.; Payton, A.S.; Cook, P.W.; Den Bakker, H.C.; Bautista, J.; Pérez, J.C.D. Pre-harvest internalization and surface survival of Salmonella and Escherichia coli O157:H7 sprayed onto different lettuce cultivars under field and growth chamber conditions. Int. J. Food. Microbiol. 2019, 291, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Sapers, G.M. Efficacy of Washing and Sanitizing Methods. Food Technol. Biotechnol. 2001, 39, 305–311. [Google Scholar]
- Hara-Kudo, Y.; Takatori, K. Contamination level and ingestion dose of foodborne pathogens associated with infections. Epidemiol. Infect. 2011, 139, 1505–1510. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Gkogka, E.; Le Guyader, F.S.; Loisy-Hamon, F.; Lee, A.; van Lieshout, L.; Marthi, B.; Myrmel, M.; Sansom, A.; Schultz, A.C.; et al. Foodborne viruses: Detection, risk assessment, and control options in food processing. Int. J. Food Microbiol. 2018, 285, 110–128. [Google Scholar] [CrossRef]
- Franz, E.; Tromp, S.O.; Rijgersberg, H.; Van Der Fels-Klerx, H.J. Quantitative microbial risk assessment for Escherichia coli O157: H7, Salmonella, and Listeria monocytogenes in leafy green vegetables consumed at salad bars. J. Food Prot. 2010, 73, 274–285. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Deep Water Raft Culture | Nutrient Film Technique | Continuous Drip | Wick Method | Flood and Drain | Conventional, Field-based |
|---|---|---|---|---|---|
| Submergence of plant roots in nutrient solution | |||||
| Roots are fully submerged in NS throughout the growing process. | Root tips touch a 1–10-mm film of NS running along the bottom of plastic gutters. | Roots grow through a solid matrix in a grow bed that is filled with NS. | Roots are fully submerged in NS throughout the growing process. | Roots grow through a solid matrix in a grow bed that is mostly filled with NS when flooded, and exposed to air when not flooded. | Roots are fully covered by the soil matrix and exposed to water through irrigation. |
| Water Flow | |||||
| No water flow | NS is actively pumped continuously or intermittently at a low flow rate. | NS is actively pumped continuously at a low flow rate. | No water flow in plant reservoir. NS is passively replenished through capillary action from the tank up through fibrous wicks. | Grow bed is periodically flooded with NS at a higher flow rate than NFT or drip, by active pumping, and then drained. The pump is typically timer-controlled. | Roots grow in soil and are watered by drip irrigation and surface watering. |
| Water recirculation | |||||
| OC | CC | CC | OC | CC | OC |
| Solid phase | |||||
| No | No | Yes | Yes | Yes | Soil, compost, manure |
| Method of root aeration | |||||
| Injection | All but the root tips are exposed to the air inside the gutters. | Agitation from pump | Injection | Exposed to air during drained periods, from agitation by the pump during flood periods. | By ensuring adequate soil drainage |
| System Type | Solid Phase | Pathogen | Plant | Inoculation | Surface Sterilized | Compared with Soil | Internalization Outcome | Ref. |
|---|---|---|---|---|---|---|---|---|
| HA-GB | N/A | E. coli O157:H7, Salmonella Typhimurium, and L. monocytogenes | Carrot, cress, lettuce, radish, spinach and tomato | Seeds soaked in 2 log CFU/mL, and then air-dried on sterile filter paper for 2 h at ~22 °C | Yes | No | Levels of all pathogens increased from 2 log to ~5–6 log CFU during 10-day germination. Counts and SEM showed a plant-specific effect (cress and radish most susceptible), a pathogen-specific effect (L. monocytogenes most abundant), and an age-specific effect (internalization was greater in young plants) | [28] |
| DWC-L-T | No | E. coli TG1 expressing GFP | Corn seedlings (Zea mays) | 7 log CFU/mL added directly to the 4-L tray of nutrient solution | No | No | Internalized E. coli TG1 detected in shoots. Entire root system removed (430 CFU/g), root tips severed (500 CFU/g), undamaged plants (18 CFU/g). | [29] |
| DWC-L-F | No | GFP-expressing E. coli O157:H7 and S. Typhimurium (MAE 110 and 119) | Lettuce (Lactuca sativa cv. Tamburo) | 29 mL of hydroponic nutrient solution with a final concentration of 7 log CFU/mL | Yes | Yes | Hydroponic: S. Typhimurium MAE 119 internalized at 5 log CFU/g. | [31] |
| DWC-L-T | No | GFP-expressing E. coli O157:H7 from a spinach outbreak and a beef outbreak as well as a non-pathogenic clinical E. coli isolate | Spinach | 3 and 7 log CFU/mL or g added directly to the nutrient solution or soil. Group 1: Inoculated hydroponic for 21 d; Group 2: Hydroponic for 21 d, transplanted into sterile soil; Group 3: hydroponic for 21 d, transplanted into inoculated soil | Yes | Yes | At both 4 log and 7 log CFU/mL in hydroponic water, between 2–4 log CFU/shoot internalized pathogen detected at cultivation day 14. Soil recovery was negligible for both high and low inocula and required enrichment to detect. 23/108 soil-grown plants showed E. coli in root tissues, but no internalization in shoots. | [32] |
| DWC-L-F | Sand | S. Typhimurium (LT1 and S1) and L. monocytogenes sv4b, L. ivanovii, L. innocua | Barley (Hordeum vulgare) | 8 log CFU/mL suspension per bacterial species added directly to the surface of the sand 1 to 2 days after planting | Yes | No | Salmonella internalized in roots, stems, and leaves, while Listeria spp. only colonized the root hairs. | [46] |
| DWC-L-C | No | Six strains of E. coli O157:H7, five strains of S. Typhimurium and S. Enteritidis, six strains of L. monocytogenes | Spinach (Brassica rapa var. perviridis) | 3 or 6 log CFU/mL added directly to the hydroponic water solution | No | No | Across all microorganisms, the 3 log CFU/mL had an average recovery of <1.7 log CFU/leaf in 7/72 samples. The 6 log CFU/mL inoculum resulted in better recovery (50/76 samples) in a range of 1.7 to 4.4 log CFU/leaf. | [47] |
| DWC-L-T | No | E. coli O157:H7 | Spinach cultivars Space and Waitiki | 5 or 7 log CFU/mL added directly to the Hoagland medium. Hoagland medium was re-inoculated as needed to maintain initial bacterial levels. | Yes | Yes | E. coli O157:H7 internalized in 15/54 samples at 7 days after inoculation with 7 log CFU/mL. Neither curli or spinach cultivar had an impact on the internalization rate. | [48] |
| DWC-L-J | Vermiculite | Coxsackievirus B2 | Lettuce (L. sativa) | 7.62–9.62 log genomic copies/L in water solution | Unknown | No | Virus detected in leaves on the first day at all inoculation levels; however, decreased to below LOD over the next 3 days. | [49] |
| NFT | Rockwool plugs | E. coli P36 (fluorescence labeled) | Spinach (Spinacia oleracea L. cv. Sharan) | 2 to 3 log CFU/mL E. coli added to the nutrient solution in the holding tank. 2 log CFU/g was added to soil. | Yes | Yes | For hydroponic: total surface (7.17 ± 1.39 log CFU/g), internal (4.03 ± 0.95 log CFU/g). For soil: total surface (6.30± 0.64 log CFU/g), internal (2.91± 0.81 log CFU/g) | [50] |
| NFT | No | MNV | Kale microgreens (Brassica napus) and mustard microgreens (Brassica juncea) | Nutrient solution containing ~3.5 log PFU/mL on day 8 of growth | Unknown | No | MNV was internalized into roots and edible tissues of both microgreens within 2 h of nutrient solution inoculation in all samples at 1.98 to 3.47 log PFU/sample. After 12 days, MNV remained internalized and detectable in 27/36 samples at 1.42 to 1.61 log PFU/sample. | [51] |
| DS | Peat pellets/clay pebbles | MNV (type 1), S. Thompson (FMFP 899) | Basil (Ocimum basilicum) | MNV (8.46 log-PFU/mL) or S. Thompson (8.60 log-CFU/mL) via soaking the germinating discs for 1 h | No | No | MNV was internalized into edible parts of basil via the roots with 400 to 580 PFU/g detected at day 1 p.i., and the LOD was reached by day 6. Samples were positive for S. Thompson on days 3 and 6 post-enrichment. | [52] |
| DWC | No | Citrobacter freundii PSS60, Enterobacter spp. PSS11, E. coli PSS2, Klebsiella oxytoca PSS82, Serratia grimesii PSS72, Pseudomonas putida PSS21, Stenotrophomonas maltophilia PSS52, L. monocytogenes ATCC 19114 | Radish (R. sativus L.) microgreens | Final concentration of 7 log CFU/mL for each bacterium added directly to the nutrient solution | Yes | No | C. freundii PSS60, Enterobacter spp. PSS11, K. oxytoca PSS82 were suspected to have internalized in hypocotyls. These three strains were detected with and without the surface sterilization of plant samples. | [53] |
| HA-TT | N/A | Klebsiella pneumoniae 342, Salmonella Cubana, Infantis, 8137, and Typhimurium; E. coli K-12, E. coli O157:H7 | Alfalfa (M. sativa) and Barrelclover (M. truncatula) | 1 to 7 log CFU/mL added directly to the growth medium at the seedling root area after 1 day of germination. | Yes | No | K. pneumoniae 342 colonized root tissue at low inoculation levels. S. Cubana H7976 colonized at high inoculation levels. No difference between Salmonella serovars | [54] |
| HA-TT | N/A | S. Dublin, Typhimurium, Enteritidis, Newport, Montevideo | Lettuce (Lactuca sativa cv. Tamburo, Nelly, Cancan) | 10 µL of a 7 log CFU/mL suspension per serovar added directly to the 0.5% Hoagland’s water agar containing two-week old seedlings | Yes | Yes | Hydroponic: S. Dublin, Typhimurium, Enteritidis, Newport, and Montevideo internalized in L. sativa Tamburo at 4.6 CFU/g, 4.27 CFU/g, 3.93 CFU/g, ~3 CFU/g, and ~4 log CFU/g, respectively | [55] |
| DWC | No | hNoV GII.4 isolate 5 M, MNV, and TV | Romaine lettuce (Lactuca sativa) | TV and MNV (6 log PFU/mL), and hNoV (6.46 log RNA copies/mL) added directly to the nutrient solution | Yes | No | TV, MNV, and hNoV detected in leaves within 1 day. At day 14, recovery levels were TV: 5.8 log PFU/g, MNV: 5.5 log PFU/g, and hNoV: 4 log RNA copies/g were recovered | [56] |
| DWC | Vermiculite | E. coli O157:H7 | Red sails lettuce (Lactuca sativa) | Started with 7 log CFU/mL and maintained in water at 5 log CFU/mL | Yes | No | E. coli O157:H7 internalized in contaminated lettuce of cut and uncut roots. Mean uncut: 2.4 ± 0.7; Mean 2 cuts: 4.0 ± 1.9; Mean 3 cuts: 3.3 ± 1.3. No significant difference was found between two and three cuts. | [57] |
| DWC-(AP) | Vermiculite | Total coliforms | Red sails lettuce (Lactuca sativa) | No inoculation. Detected 2 to 4 log CFU/mL natural concentration of coliform bacteria in a pilot system downstream of a cattle pasture | Yes | No | UV light at 96.6% transmittance and a flow rate of 48.3 L/min reduced total coliforms by 3 log CFU/mL in water. Internalized coliform was not recovered from either samples or control lettuce. | [58] |
| Method | Advantages | Disadvantages |
|---|---|---|
| Membrane filtration | Precise filtration, can choose pore size to suit needs | Reduced flow rate, easy clogging |
| Slow sand filtration | Most common, inexpensive, a variety of substrate choices. | May not effectively remove pathogens on its own |
| UV light treatment | Can be combined with slow sand filtration for high efficiency | Water needs high clarity, so must be combined with sediment filter to ensure maximum light penetration |
| Chlorination | Inexpensive, standard recommendation | Storage issues, toxic to humans |
| Iodine | Less toxic than chlorine | Need high doses to be effective, costly |
| Hydrogen peroxide | Less toxic than chlorine, weak oxidizer | Need high doses to be effective, costly |
| Ozonation | Non-toxic to humans, no residues left behind | Strong oxidizer may cause hydroponic mineral nutrients to precipitate, reducing bioavailability |
| Biological control agents | Takes advantage of natural features of the system to suppress pathogens without addition of harsh chemicals | Inconsistent, difficult to maintain microbial numbers to sufficiently suppress pathogens, manipulation of microbiome for this purpose still a poorly understood research area. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riggio, G.M.; Jones, S.L.; Gibson, K.E. Risk of Human Pathogen Internalization in Leafy Vegetables During Lab-Scale Hydroponic Cultivation. Horticulturae 2019, 5, 25. https://doi.org/10.3390/horticulturae5010025
Riggio GM, Jones SL, Gibson KE. Risk of Human Pathogen Internalization in Leafy Vegetables During Lab-Scale Hydroponic Cultivation. Horticulturae. 2019; 5(1):25. https://doi.org/10.3390/horticulturae5010025
Chicago/Turabian StyleRiggio, Gina M., Sarah L. Jones, and Kristen E. Gibson. 2019. "Risk of Human Pathogen Internalization in Leafy Vegetables During Lab-Scale Hydroponic Cultivation" Horticulturae 5, no. 1: 25. https://doi.org/10.3390/horticulturae5010025
APA StyleRiggio, G. M., Jones, S. L., & Gibson, K. E. (2019). Risk of Human Pathogen Internalization in Leafy Vegetables During Lab-Scale Hydroponic Cultivation. Horticulturae, 5(1), 25. https://doi.org/10.3390/horticulturae5010025