
Preliminary Clonal Characterization of Malvasia Volcanica and Listan Prieto by Simple Sequence Repeat (SSR) Markers in Free-Phylloxera Volcanic Vineyards (Lanzarote and Fuerteventura (Canary Island, Spain))
 , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Plant Material
2.1.1. Lanzarote Island
2.1.2. Fuerteventura Island
2.2. DNA Extraction and Purification
2.3. Simple Sequence Repeat (SSR) Markers
2.4. DNA Amplification
2.5. Amplified Fragments Analysis
2.6. Sample Identification
3. Results
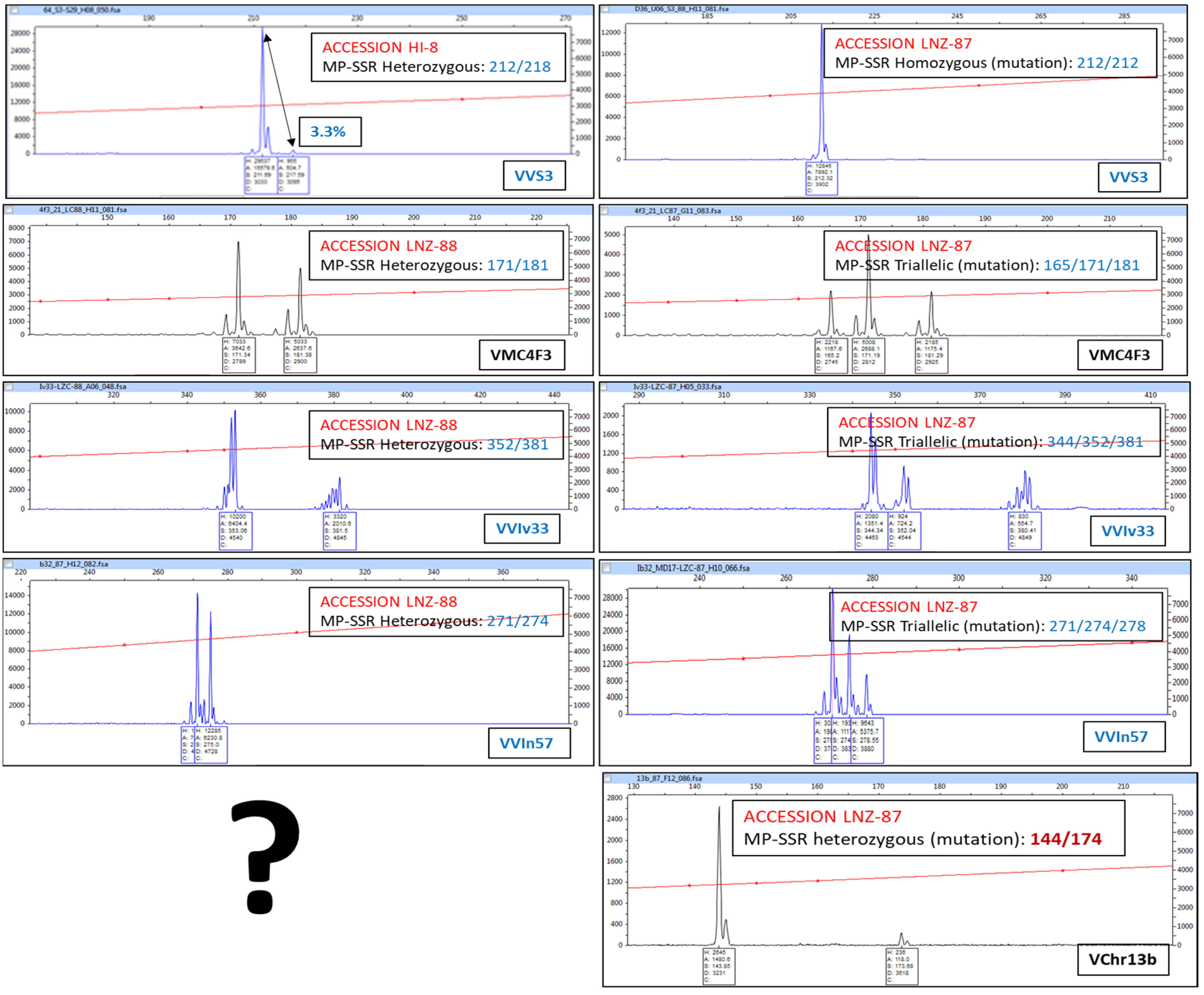
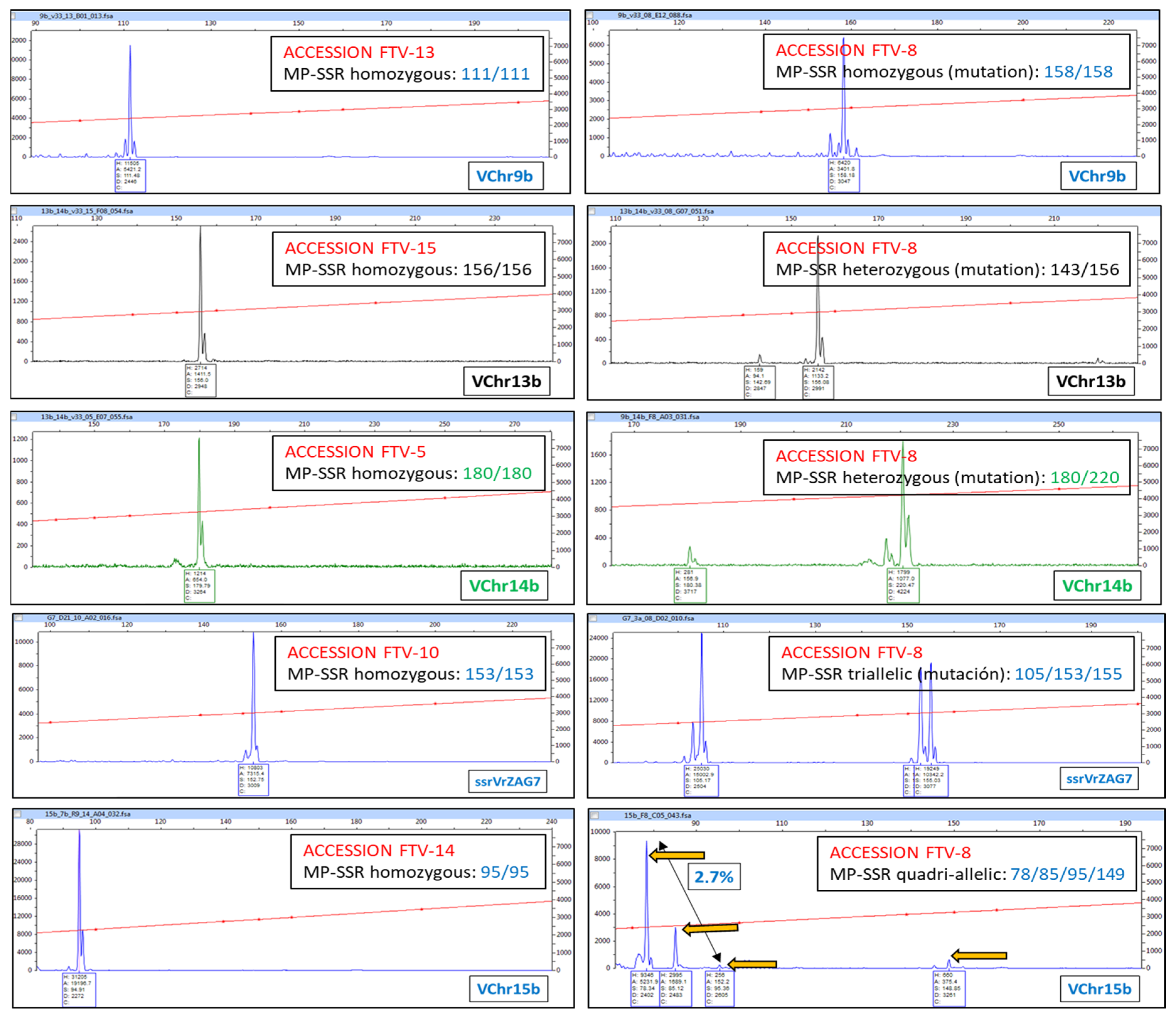
3.1. Inter-Varietal and Intra-Varietal Variability (Clones Identified: Variations Relative to the Most Common Molecular Profile (Mutations))
3.2. SSR Markers Used: Effectiveness and Number of Alleles Computed
3.2.1. Effectiveness in Detecting Molecular Profiles
3.2.2. Variations in Allele Number
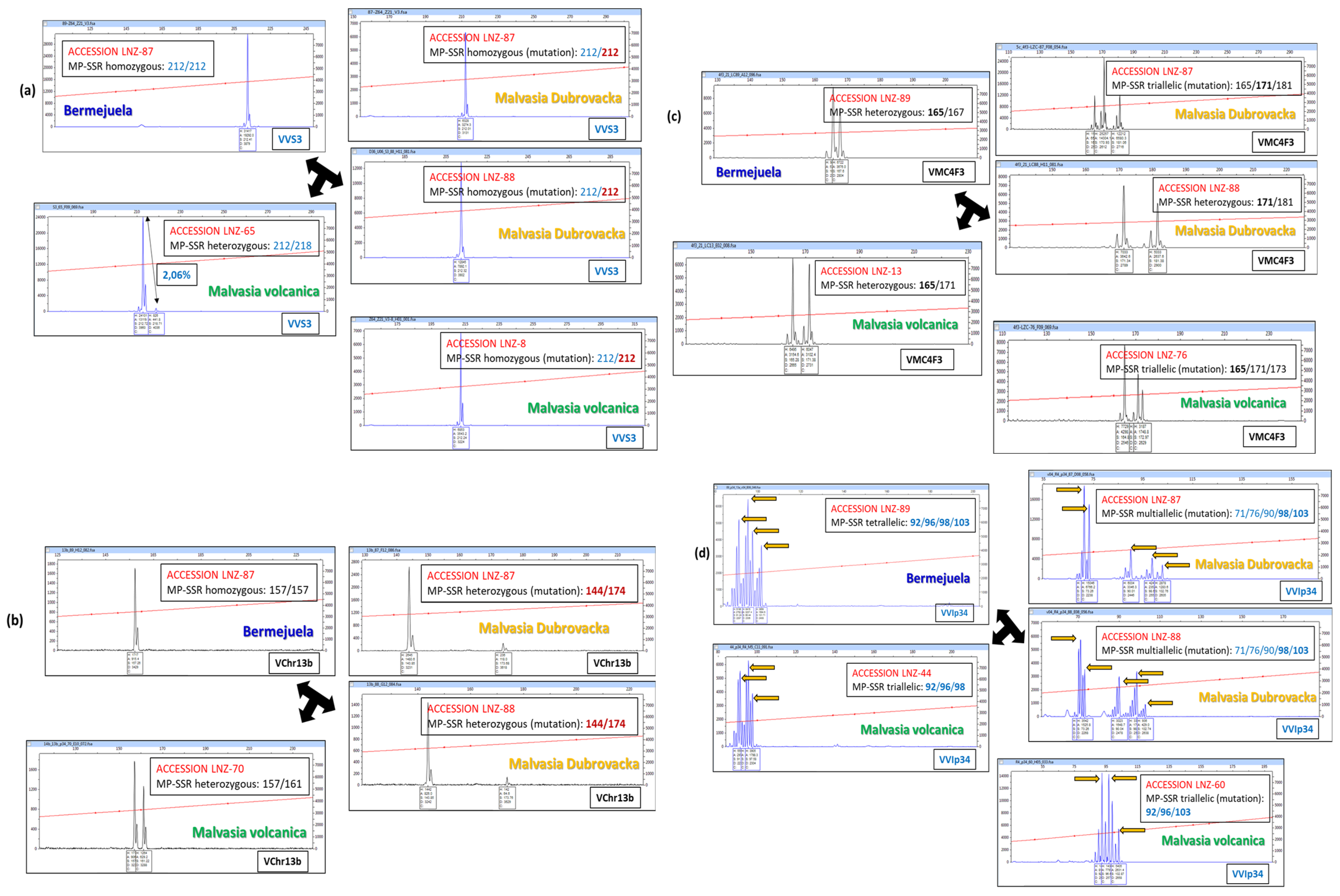
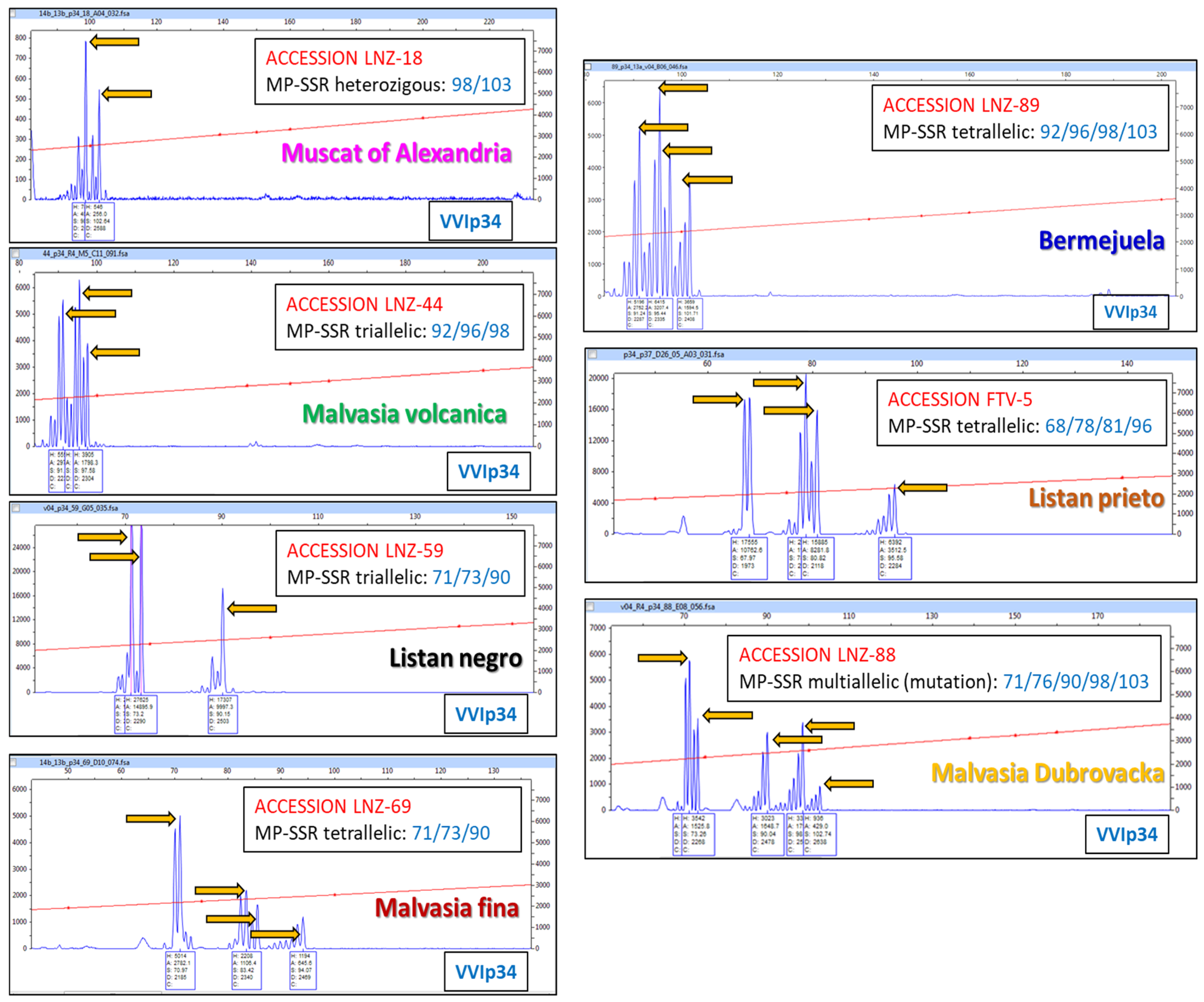
3.3. Pedigree of the Malvasia Volcanica Variety
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myles, S.; Boyko, A.R.; Owens, C.L.; Brown, P.J.; Grassi, F.; Aradhya, M.K.; Prins, B.; Reynolds, A.; Chia, J.-M.; Ware, D.; et al. Genetic structure and domestication history of the grape. Proc. Natl. Acad. Sci. USA 2011, 108, 3457–3462. [Google Scholar] [CrossRef] [PubMed]
- Emanuelli, F.; Lorenzi, S.; Grzeskowiak, L.; Catalano, V.; Stefanini, M.; Troggio, M.; Myles, S.; Martinez-Zapater, J.M.; Zyprian, E.; Moreira, F.M.; et al. Genetic diversity and population structure assessed by SSR and SNP markers in a large germplasm collection of grape. BMC Plant Biol. 2013, 13, 39. [Google Scholar] [CrossRef]
- International Organisation of Vine and Wine. State of the World Vine and Wine Sector, 2023; OIV: Paris, France, 2023; Available online: https://www.oiv.int/sites/default/files/documents/OIV_State_of_the_world_Vine_and_Wine_sector_in_2022_2.pdf (accessed on 15 May 2025).
- Wolkovich, E.M.; García de Cortázar-Atauri, I.; Morales-Castilla, I.; Nicholas, K.A.; Lacombe, T. From Pinot to Xinomavro in the world’s future wine-growing regions. Nat. Clim. Change 2018, 8, 29–37. [Google Scholar] [CrossRef]
- Pelsy, F.; Dumas, V.; Bévilacqua, L.; Hocquigny, S.; Merdinoglu, D. Chromosome replacement and deletion lead to clonal polymorphism of berry color in grapevine. PLoS Genet. 2015, 11, e1005081. [Google Scholar] [CrossRef]
- Jackson, R.S. Wine Science: Principles and Applications, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1–978. [Google Scholar]
- Tympakianakis, S.; Trantas, E.; Avramidou, E.V.; Ververidis, F. Vitis vinifera genotyping toolbox to highlight diversity and germplasm identification. Front. Plant Sci. 2023, 14, 1139647. [Google Scholar] [CrossRef] [PubMed]
- Fort, F.; Lin-Yang, Q.; Valls, C.; Sancho-Galán, P.; Canals, J.M.; Zamora, F. Analysis of the diversity presented by Vitis vinifera L. in the volcanic island of La Gomera (Canary Archipelago, Spain) using simple sequence repeats (SSRs) as molecular markers. Horticulturae 2024, 10, 14. [Google Scholar] [CrossRef]
- Migliaro, D.; De Nardi, B.; Vezzulli, S.; Crespan, M. An upgraded core set of 11 SSR markers for grapevine cultivar identification: The case of berry-color mutants. Am. J. Enol. Vitic. 2017, 68, 496–502. [Google Scholar] [CrossRef]
- Peiró, R.; Soler, J.X.; Crespo, A.; Jiménez, C.; Cabello, F.; Gisbert, C. Genetic variability assessment in ‘Muscat’ grapevines including ‘Muscat of Alexandria’ clones from selection programs. Span. J. Agric. Res. 2018, 16, e0702. [Google Scholar] [CrossRef]
- Carrier, G.; Le Cunff, L.; Dereeper, A.; Legrand, D.; Sabot, F.; Bouchez, O.; Audeguin, L.; Boursiquot, J.M.; This, P. Transposable elements are a major cause of somatic polymorphism in Vitis vinifera L. PLoS ONE 2012, 7, e32973. [Google Scholar] [CrossRef]
- Meneghetti, S.; Calò, A.; Bavaresco, L. A strategy to investigate the intravarietal genetic variability in Vitis vinifera L. for clones and biotypes identification and to correlate molecular profiles with morphological traits or geographic origins. Mol. Biotechnol. 2012, 52, 68–81. [Google Scholar] [CrossRef]
- De Lorenzis, G.; Squadrito, M.; Rossoni, M.; Di Lorenzo, G.S.; Brancadoro, L.; Scienza, A. Study of intra-varietal diversity in biotypes of Aglianico and Muscat of Alexandria (Vitis vinifera L.) cultivars. Aust. J. Grape Wine Res. 2017, 23, 132–142. [Google Scholar] [CrossRef]
- Calderón, L.; Mauri, N.; Muñoz, C.; Carbonell-Bejerano, P.; Bree, L.; Bergamin, D.; Sola, C.; Gomez-Talquenca, S.; Royo, C.; Ibáñez, J.; et al. Whole genome resequencing and custom genotyping unveil clonal lineages in ‘Malbec’ grapevines (Vitis vinifera L.). Sci. Rep. 2021, 11, 7775. [Google Scholar] [CrossRef]
- Gambino, G.; Dal Molin, A.; Boccacci, P.; Minio, A.; Chitarra, W.; Avanzato, C.G.; Tononi, P.; Perrone, I.; Raimondi, S.; Schneider, A.; et al. Whole-genome sequencing and SNV genotyping of ‘Nebbiolo’ (Vitis vinifera L.) clones. Sci. Rep. 2017, 7, 17294. [Google Scholar] [CrossRef] [PubMed]
- Araya-Ortega, D.; Gainza-Cortés, F.; Riadi, G. Exploring genomic differences between a pair of Vitis vinifera clones using WGS data: A preliminary study. Horticulturae 2024, 10, 1026. [Google Scholar] [CrossRef]
- Villano, C.; Procino, S.; Blaiotta, G.; Carputo, D.; D’Agostino, N.; Di Serio, E.; Fanelli, V.; La Notte, P.; Miazzi, M.M.; Montemurro, C.; et al. Genetic diversity and signature of divergence in the genome of grapevine clones of Southern Italy varieties. Front. Plant Sci. 2023, 14, 1201287. [Google Scholar] [CrossRef]
- Procino, S.; Miazzi, M.M.; Savino, V.N.; La Notte, P.; Venerito, P.; D’Agostino, N.; Taranto, F.; Montemurro, C. Genome scan analysis for advancing knowledge and conservation strategies of Primitivo clones (Vitis vinifera L.). Plants 2025, 14, 437. [Google Scholar] [CrossRef]
- Esteras, C.; Peiró, R.; Soler, J.X.; Gisbert, C. Genetic variability in grapevine clones of ‘Muscat of Alexandria’. Acta Hortic. 2019, 1248, 77–80. [Google Scholar] [CrossRef]
- Urra, C.; Sanhueza, D.; Pavez, C.; Tapia, P.; Núñez-Lillo, G.; Minio, A.; Miossec, M.; Blanco-Herrera, F.; Gainza, F.; Castro, A.; et al. Identification of grapevine clones via high-throughput amplicon sequencing: A proof-of-concept study. G3 Genes Genomes Genet. 2023, 13, jkad145. [Google Scholar] [CrossRef]
- Jahnke, G.; Májer, J.; Varga, P.; Szőke, B. Analysis of clones of Pinots grown in Hungary by SSR markers. Sci. Hortic. 2011, 129, 32–37. [Google Scholar] [CrossRef]
- Baltazar, M.; Castro, I.; Gonçalves, B. Adaptation to climate change in viticulture: The role of varietal selection—A review. Plants 2025, 14, 104. [Google Scholar] [CrossRef]
- Marsal, G.; Mendez, J.J.; Mateo-Sanz, J.M.; Ferrer, S.; Canals, J.M.; Zamora, F.; Fort, F. Molecular characterization of Vitis vinifera L. local cultivars from volcanic areas (the Canary Islands and Madeira) using SSR markers. OENO One 2019, 53, 667–680. [Google Scholar] [CrossRef]
- Fort, F.; Lin-Yang, Q.; Suárez-Abreu, L.R.; Sancho-Galán, P.; Canals, J.M.; Zamora, F. Study of molecular biodiversity and population structure of Vitis vinifera L. ssp. vinifera on the volcanic island of El Hierro (Canary Islands, Spain) by using microsatellite markers. Horticulturae 2023, 9, 1297. [Google Scholar] [CrossRef]
- Fort, F.; Marsal, G.; Mateo-Sanz, J.M.; Pena, V.; Canals, J.M.; Zamora, F. Molecular characterisation of the current cultivars of Vitis vinifera L. in Lanzarote (Canary Islands, Spain) reveals nine individuals which correspond to eight new varieties and two new sports. OENO One 2022, 56, 281–295. [Google Scholar] [CrossRef]
- Fort, F.; Lin-Yang, Q.; Valls, C.; Sancho-Galán, P.; Canals, J.M.; Zamora, F. Characterisation and identification of vines from Fuerteventura (Canary Volcanic Archipelago (Spain)) using simple sequence repeat markers. Horticulturae 2023, 9, 1301. [Google Scholar] [CrossRef]
- Mendez, J.J. Acerca del Canary Wine. Compendio de la Vitivinicultura del Archipiélago Canario, 2nd ed.; Asociación de Viticultores y Bodegueros de Canarias AVIBO: Santa Cruz de Tenerife, Spain, 2024. [Google Scholar]
- González Morales, A. El Cultivo y Producción del vino en la isla de Lanzarote. Territoires du vin [Online], 1 March 2011. Available online: https://preo.ube.fr/territoiresduvin/index.php?id=1407 (accessed on 15 May 2025).
- Marsal, G.; Boronat, N.; Canals, J.M.; Zamora, F.; Fort, F. Comparison of the efficiency of some of the most usual DNA extraction methods for woody plants in different tissues of Vitis vinifera L. J. Int. Sci. Vigne Vin 2013, 47, 227–237. [Google Scholar] [CrossRef]
- Marsal, G.; Baiges, I.; Canals, J.M.; Zamora, F.; Fort, F. A fast, efficient method for extracting DNA from leaves, stems, and seeds of Vitis vinifera L. Am. J. Enol. Vitic. 2011, 62, 376–381. [Google Scholar] [CrossRef]
- Fort, F.; Hayoun, L.; Valls, J.; Canals, J.M.; Arola, L.; Zamora, F. A new and simple method for rapid extraction and isolation of high-quality RNA from grape (Vitis vinifera) berries. J. Sci. Food Agric. 2008, 88, 179–184. [Google Scholar] [CrossRef]
- Thomas, M.R.; Scott, N.S. Microsatellite repeats in grapevine reveal DNA polymorphisms when analysed as sequence-tagged sites (STSs). Theor. Appl. Genet. 1993, 86, 985–990. [Google Scholar] [CrossRef]
- Bowers, J.E.; Dangl, G.S.; Vignani, R.; Meredith, C.P. Isolation and characterization of new polymorphic simple sequence repeat loci in grape (Vitis vinifera L.). Genome 1996, 39, 628–633. [Google Scholar] [CrossRef]
- Bowers, J.E.; Dangl, G.S.; Meredith, C.P. Development and characterization of additional microsatellite DNA markers for grape. Am. J. Enol. Vitic. 1999, 50, 243–246. [Google Scholar] [CrossRef]
- Sefc, K.M.; Regner, F.; Turetschek, E.; Glossl, J.; Steinkellner, H. Identification of microsatellite sequences in Vitis riparia and their applicability for genotyping of different Vitis species. Genome 1999, 42, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.D.; Eggler, P.; Seaton, G.; Rosseto, M.; Ablett, E.M.; Lee, L.S.; Henry, R.J. Analysis of SSRs derived from grape ESTs. Theor. Appl. Genet. 2000, 100, 723–726. [Google Scholar] [CrossRef]
- Di Gaspero, G.; Peterlunger, E.; Testolin, R.; Edwards, K.J.; Cipriani, G. Conservation of microsatellite loci within the genus Vitis. Theor. Appl. Genet. 2000, 101, 301–308. [Google Scholar] [CrossRef]
- Lefort, F.; Kyvelos, C.; Zervou, M.; Edwards, K.; Roubelakis-Angelakis, K. Characterization of new microsatellite loci from Vitis vinifera and their conservation in some Vitis species and hybrids. Mol. Ecol. Resour. 2002, 2, 20–21. [Google Scholar] [CrossRef]
- Crespan, M. The parentage of Muscat of Hamburg. Vitis 2003, 42, 193–197. [Google Scholar] [CrossRef]
- Arroyo-García, R.; Martínez-Zapater, J.M. Development and characterization of new microsatellite markers for grape. Vitis 2004, 43, 175–178. [Google Scholar] [CrossRef]
- Merdinoglu, D.; Butterlin, G.; Bevilacqua, L.; Chiquet, V.; Adam-Blondon, A.-F.; Decroocq, S. Development and characterization of a large set of microsatellite markers in grapevine (Vitis vinifera L.) suitable for multiplex PCR. Mol. Breed. 2005, 15, 349–366. [Google Scholar] [CrossRef]
- Doligez, A.; Adam-Blondon, A.F.; Cipriani, G.; Di Gaspero, G.; Laucou, V.; Merdinoglu, D.; Meredith, C.P.; Riaz, S.; Roux, C.; This, P. An integrated SSR map of grapevine based on five mapping populations. Theor. Appl. Genet. 2006, 113, 369–382. [Google Scholar] [CrossRef]
- Regner, F.; Hack, R.; Santiago, J.L. Highly variable Vitis microsatellite loci for the identification of Pinot Noir clones. Vitis 2006, 45, 85–91. [Google Scholar] [CrossRef]
- Cipriani, G.; Marrazzo, M.T.; Di Gaspero, G.; Pfeiffer, A.; Morgante, M.; Testolin, R. A set of microsatellite markers with long core repeat optimized for grape (Vitis spp.) genotyping. BMC Plant Biol. 2008, 8, 127. [Google Scholar] [CrossRef]
- Fournier-Level, A.; Le Cunff, L.; Gomez, C.; Doligez, A.; Ageorges, A.; Roux, C.; Bertrand, Y.; Souquet, J.M.; Cheynier, V.; This, P. Quantitative genetic bases of anthocyanin variation in grape (Vitis vinifera L. ssp. sativa) berry: A quantitative trait locus to quantitative trait nucleotide integrated study. Genetics 2009, 183, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- This, P.; Jung, A.; Boccacci, P.; Borrego, J.; Botta, R.; Costantini, L.; Crespan, M.; Dangl, G.S.; Eisenheld, C.; Ferreira-Monteiro, F.; et al. Development of a standard set of microsatellite reference alleles for identification of grape cultivars. Theor. Appl. Genet. 2004, 109, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Marsal, G.; Mateo, J.M.; Canals, J.M.; Zamora, F.; Fort, F. SSR analysis of 338 accessions planted in Penedès (Spain) reveals 28 unreported molecular profiles of Vitis vinifera L. Am. J. Enol. Vitic. 2016, 67, 466–470. [Google Scholar] [CrossRef]
- Marsal, G.; Bota, J.; Martorell, A.; Canals, J.M.; Zamora, F.; Fort, F. Local cultivars of Vitis vinifera L. in Spanish islands: Balearic Archipelago. Sci. Hortic. 2017, 226, 122–132. [Google Scholar] [CrossRef]
- Maul, E.; Röckel, F. Vitis International Variety Catalogue. Available online: http://www.vivc.de (accessed on 15 May 2025).
- Rodríguez-Torres, I. Variedades de Vid Cultivadas en Canarias. Descriptores Morfológicos. Caracterización Morfológica, Molecular, Agronómica y Enológica; Instituto Canario de Investigaciones Agrarias, Gobierno de Canarias: Santa Cruz de Tenerife, Spain, 2018. [Google Scholar]
- El Oualkadi, A.; Ater, M.; Messaoudi, Z.; Laucou, V.; Boursiquot, J.M.; Lacombe, T.; This, P. Molecular characterization of Moroccan grapevine germplasm using SSR markers for the establishment of a reference collection. OENO One 2009, 43, 135–148. [Google Scholar] [CrossRef]
- Fort, F.; Suárez-Abreu, L.R.; Lin-Yang, Q.; Deis, L.; Canals, J.M.; Zamora, F. Origin and Possible Members of the ‘Malvasia’ Family: The New Fuencaliente de La Palma Hypothesis on the True ‘Malvasia’. Horticulturae 2025, 11, 561. [Google Scholar] [CrossRef]
- Zerolo, J.; Cabello, F.; Espino, A.; Borrego, J.; Ibañez, J.; Rodriguez-Torres, I.; Muñoz-Organero, G.; Rubio, C.; Hernández, M. Variedades de Vid de Cultivo Tradicional en Canarias, 1st ed.; Instituto Canario de Calidad Agroalimentaria, Gobierno de Canarias: Santa Cruz de Tenerife, Spain, 2006; ISBN 978-84-606-3977-0. [Google Scholar]
- Aliquo, G.; Torres, R.; Lacombe, T.; Boursiquot, J.M.; Laucou, V.; Gualpa, J.; Fanzone, M.; Sari, S.; Pérez-Peña, J.; Prieto, J.A. Identity and parentage of some South American grapevine cultivars present in Argentina. Aust. J. Grape Wine Res. 2017, 23, 452–460. [Google Scholar] [CrossRef]
- Galet, P. Dictionnaire Encyclopédique des Cépages, 1st ed.; Hachette: Paris, France, 2000. [Google Scholar]
- Macías, A. El paisaje vitícola de Canarias. Cinco siglos de historia. Ería 2005, 68, 351–364. Available online: https://reunido.uniovi.es/index.php/RCG/article/view/1525 (accessed on 15 May 2025).
- Hidalgo, J.; Hidalgo, L. La Filoxera. In Tratado de Viticultura, 2nd ed.; Mundi-Prensa: Madrid, Spain, 2019; Volume 1. [Google Scholar]
- Jančić, R.S. Morfološka i Molekularna Karakterizacija Potencijalnih Klonova Sorte Vinove loze Vranac [Morphological and Molecular Characterization of Potential Clones of the Grapevine Variety Vranac]. Ph.D. Thesis, University of Novi Sad, Novi Sad, Serbia, 2022. NaRDuS Repository. Available online: https://nardus.mpn.gov.rs/handle/123456789/21475 (accessed on 15 May 2025).
- Dong, Y.; Duan, S.; Xia, Q.; Liang, Z.; Dong, X.; Margaryan, K.; Musayev, M.; Goryslavets, S.; Zdunić, G.; Bert, P.-F.; et al. Dual domestications and origin of traits in grapevine evolution. Science 2023, 379, 892–901. [Google Scholar] [CrossRef]
- This, P.; Lacombe, T.; Thomas, M.R. Historical origins and genetic diversity of wine grapes. Trends Genet. 2006, 22, 511–519. [Google Scholar] [CrossRef]
- Hocquigny, S.; Pelsy, F.; Dumas, V.; Kindt, S.; Heloir, M.-C.; Merdinoglu, D. Diversification within grapevine cultivars goes through chimeric states. Genome 2004, 47, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Massonnet, M.; Sanjak, J.S.; Cantu, D.; Gaut, B.S. Evolutionary genomics of grape (Vitis vinifera ssp. vinifera) domestication. Proc. Natl. Acad. Sci. USA 2019, 116, 3720–3725. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Gschwend, A.R. Vitis labrusca genome assembly reveals diversification between wild and cultivated grapevine genomes. Front. Plant Sci. 2023, 14, 1234130. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.M. Advanced Topics in Forensic DNA Typing: Interpretation; Academic Press: London, UK; Oxford, UK; Boston, MA, USA; New York, NY, USA; San Diego, CA, USA, 2014; ISBN 9780124052130. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SSR No. | 2 | 3 | 4 | 8 | 22 | 23 | 34 | 40 | 47 | 55 | 60 | 69 | 75 | 77 | 80 | 84 | 86 | 87 | 88 | 92 | 97 | 98 | 100 |
| Peak size range/Fluorochrome colour | 214–222 | 168–186 | 215–270 | 210–279 | 278–318 | 156–230 | 239–263 | 217–289 | 106–158 | 114–135 | 350–400 | 250–300 | 102–160 | 90–151 | 150–200 | 106–292 | 250–300 | 222–250 | 250–300 | 176–243 | 185–292 | 50–100 | 145–170 |
| SAMPLE CODE/SSR | VVS3 | VVS29 | VVMD5 | VVMD28 | VVIb01 | VMC4F3 | VRG16 | VVMD32 | ZAG7 | VChr13c | VVIv33 | VVIb09 | VChr9b | VChr15b | VVIv51 | VRG2 | VVIn57 | VVMD14 | VVIt60 | VChr14b | VRG3 | VVIp34 | VChr13b |
| LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 | LNZ-1 |
| LNZ-2 | LNZ-2* | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 | LNZ-2 |
| LNZ-3 | LNZ-3* | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3* | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 | LNZ-3 |
| LNZ-4 | LNZ-4* | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 | LNZ-4 |
| LNZ-5 | LNZ-5* | LNZ-5* | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 | LNZ-5 |
| LNZ-6 | LNZ-6* | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6* | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 | LNZ-6 |
| LNZ-7 | LNZ-7* | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 | LNZ-7 |
| LNZ-8 | LNZ-8* | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8* | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 | LNZ-8 |
| LNZ-9 | LNZ-9* | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 | LNZ-9 |
| LNZ-10 | LNZ-10* | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 | LNZ-10 |
| LNZ-11 | LNZ-11* | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 | LNZ-11 |
| LNZ-12 | LNZ-12* | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 | LNZ-12 |
| LNZ-13 | LNZ-13* | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 | LNZ-13 |
| LNZ-14 | LNZ-14* | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 | LNZ-14 |
| LNZ-15 | LNZ-15* | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 | LNZ-15 |
| LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16 | LNZ-16* | LNZ-16 |
| LNZ-17 | LNZ-17* | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17* | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17* | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17 | LNZ-17 |
| LNZ-19 | LNZ-19* | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19* | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 | LNZ-19 |
| LNZ-20 | LNZ-20* | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 | LNZ-20 |
| LNZ-21 | LNZ-21* | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21* | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 | LNZ-21 |
| LNZ-22 | LNZ-22* | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 | LNZ-22 |
| LNZ-23 | LNZ-23* | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 | LNZ-23 |
| LNZ-24 | LNZ-24 | LNZ-24* | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24* | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 | LNZ-24 |
| LNZ-25 | LNZ-25* | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 | LNZ-25 |
| LNZ-26 | LNZ-26* | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 | LNZ-26 |
| LNZ-27 | LNZ-27* | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 | LNZ-27 |
| LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 | LNZ-28 |
| LNZ-29 | LNZ-29* | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29* | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 | LNZ-29 |
| LNZ-30 | LNZ-30* | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 | LNZ-30 |
| LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31* | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 | LNZ-31 |
| LNZ-32 | LNZ-32* | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32* | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 | LNZ-32 |
| LNZ-33 | LNZ-33* | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 | LNZ-33 |
| LNZ-34 | LNZ-34* | LNZ-34* | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 | LNZ-34 |
| LNZ-35 | LNZ-35* | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35* | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 | LNZ-35 |
| LNZ-36 | LNZ-36* | LNZ-36* | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 | LNZ-36 |
| LNZ-37 | LNZ-37* | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 | LNZ-37 |
| LNZ-38 | LNZ-38* | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 | LNZ-38 |
| LNZ-39 | LNZ-39* | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 | LNZ-39 |
| LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40* | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 | LNZ-40 |
| LNZ-41 | LNZ-41 | LNZ-41* | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 | LNZ-41 |
| LNZ-42 | LNZ-42* | LNZ-42* | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 | LNZ-42 |
| LNZ-43 | LNZ-43* | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 | LNZ-43 |
| LNZ-44 | LNZ-44* | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 | LNZ-44 |
| LNZ-45 | LNZ-45* | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45* | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 | LNZ-45 |
| LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 | LNZ-46 |
| LNZ-47 | LNZ-47* | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47* | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 | LNZ-47 |
| LNZ-48 | LNZ-48* | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48* | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 | LNZ-48 |
| LNZ-49 | LNZ-49* | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 | LNZ-49 |
| LNZ-50 | LNZ-50* | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50* | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 | LNZ-50 |
| LNZ-51 | LNZ-51* | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 | LNZ-51 |
| LNZ-53 | LNZ-53* | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 | LNZ-53 |
| LNZ-54 | LNZ-54* | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54* | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 | LNZ-54 |
| LNZ-55 | LNZ-55* | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55* | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 | LNZ-55 |
| LNZ-56 | LNZ-56* | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56* | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 | LNZ-56 |
| LNZ-57 | LNZ-58 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 | LNZ-57 |
| LNZ-58 | LNZ-58* | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 | LNZ-58 |
| LNZ-60 | LNZ-60* | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60 | LNZ-60* | LNZ-60 |
| LNZ-61 | LNZ-61* | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 | LNZ-61 |
| LNZ-62 | LNZ-62* | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 | LNZ-62 |
| LNZ-63 | LNZ-63* | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 | LNZ-63 |
| LNZ-64 | LNZ-64* | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 | LNZ-64 |
| LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 | LNZ-65 |
| LNZ-66 | LNZ-66* | LNZ-66* | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 | LNZ-66 |
| LNZ-67 | LNZ-67* | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 | LNZ-67 |
| LNZ-68 | LNZ-68* | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68* | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 | LNZ-68 |
| LNZ-70 | LNZ-70* | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 | LNZ-70 |
| LNZ-71 | LNZ-71* | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 | LNZ-71 |
| LNZ-72 | LNZ-72* | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72* | LNZ-72 | LNZ-72* | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 | LNZ-72 |
| LNZ-73 | LNZ-73* | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73* | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 | LNZ-73 |
| LNZ-74 | LNZ-74* | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 | LNZ-74 |
| LNZ-75 | LNZ-75* | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 | LNZ-75 |
| LNZ-76 | LNZ-76* | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76* | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 | LNZ-76 |
| LNZ-77 | LNZ-77* | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 | LNZ-77 |
| LNZ-78 | LNZ-78* | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78* | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 | LNZ-78 |
| LNZ-79 | LNZ-79* | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 | LNZ-79 |
| LNZ-80 | LNZ-80* | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80* | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 | LNZ-80 |
| LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81* | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 | LNZ-81 |
| LNZ-82 | LNZ-82* | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 | LNZ-82 |
| LNZ-83 | LNZ-83* | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83* | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 | LNZ-83 |
| LNZ-84 | LNZ-84* | LNZ-82 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84* | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 | LNZ-84 |
| LNZ-85 | LNZ-85* | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85* | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 | LNZ-85 |
| LNZ-86 | LNZ-86* | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 | LNZ-86 |
| FTV-1 | FTV-1 | FTV-1 | FTV-1 | FTV-1* | FTV-1 | FTV-1 | FTV-1 | FTV-1 | FTV-1 | FTV-1 | FTV-1 | FTV-1 | FTV-1* | FTV-1* | FTV-1 | FTV-1 | FTV-1 | FTV-1 | FTV-1 | FTV-1 | FTV-1 | FTV-1 | FTV-1 |
| FTV-2 | FTV-2* | FTV-2 | FTV-2 | FTV-2* | FTV-2 | FTV-2 | FTV-2 | FTV-2 | FTV-2 | FTV-2 | FTV-2 | FTV-2 | FTV-2 | FTV-2* | FTV-2 | FTV-2 | FTV-2 | FTV-2 | FTV-2 | FTV-2 | FTV-2 | FTV-2 | FTV-2 |
| FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3* | FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3* | FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3 | FTV-3 |
| FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4* | FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4* | FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4 | FTV-4 |
| FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5* | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 | FTV-5 |
| FTV-7 | FTV-7 | FTV-7 | FTV-7 | FTV-7* | FTV-7 | FTV-7 | FTV-7 | FTV-7 | FTV-7 | FTV-7 | FTV-7 | FTV-7 | FTV-7 | FTV-7* | FTV-7 | FTV-7 | FTV-7 | FTV-7 | FTV-7 | FTV-7* | FTV-7 | FTV-7 | FTV-7 |
| FTV-8 | FTV-8 | FTV-8 | FTV-8 | FTV-8 | FTV-8 | FTV-8 | FTV-8 | FTV-8 | FTV-8* | FTV-8 | FTV-8 | FTV-8 | FTV-8* | FTV-8* | FTV-8 | FTV-8 | FTV-8 | FTV-8 | FTV-8 | FTV-8* | FTV-8 | FTV-8 | FTV-8* |
| FTV-9 | FTV-9* | FTV-9 | FTV-9 | FTV-9* | FTV-9 | FTV-9 | FTV-9 | FTV-9 | FTV-9* | FTV-9 | FTV-9 | FTV-9 | FTV-9 | FTV-9 | FTV-9 | FTV-9 | FTV-9 | FTV-9 | FTV-9 | FTV-9 | FTV-9 | FTV-9 | FTV-9 |
| FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10* | FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10* | FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10 | FTV-10 |
| FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11* | FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11* | FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11 | FTV-11 |
| FTV-12 | FTV-12 | FTV-12 | FTV-12 | FTV-12* | FTV-12 | FTV-12 | FTV-12 | FTV-12 | FTV-12 | FTV-12 | FTV-12 | FTV-12 | FTV-12 | FTV-12* | FTV-12 | FTV-12 | FTV-12 | FTV-12 | FTV-12 | FTV-12* | FTV-12 | FTV-12 | FTV-12 |
| FTV-13 | FTV-13* | FTV-13 | FTV-13 | FTV-13* | FTV-13 | FTV-13 | FTV-13 | FTV-13 | FTV-13* | FTV-13 | FTV-13 | FTV-13 | FTV-13 | FTV-13* | FTV-13 | FTV-13 | FTV-13 | FTV-13 | FTV-13 | FTV-13 | FTV-13 | FTV-13 | FTV-13 |
| FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14* | FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14* | FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14 | FTV-14 |
| FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 | FTV-15 |
| FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16* | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16 | FTV-16* | FTV-16 | FTV-16 | FTV-16 |
| FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18* | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 | FTV-18 |
| FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20* | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 | FTV-20 |
| FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32* | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-32 | FTV-31 | FTV-32 | FTV-32 |
| LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18* | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 | LNZ-18 |
| LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 | LNZ-52 |
| LNZ-87 | LNZ-87* | LNZ-87 | LNZ-87 | LNZ-87 | LNZ-87 | LNZ-87* | LNZ-87 | LNZ-87 | LNZ-87 | LNZ-87 | LNZ-87* | LNZ-87 | LNZ-87 | LNZ-87 | LNZ-87 | LNZ-87 | LNZ-87* | LNZ-87 | LNZ-87 | LNZ-87 | LNZ-87 | LNZ-87 | LNZ-87* |
| LNZ-88 | LNZ-88* | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88 | LNZ-88* |
| LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89* | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 | LNZ-89 |
| LNZ-59 | LNZ-59* | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 | LNZ-59 |
| LNZ-69 | LNZ-69* | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 | LNZ-69 |
| SAMPLE CODE/SSR | VVS3* | VVS29* | VVMD5* | VVMD28* | VVIb01 | VMC4F3* | VRG16* | VVMD32* | ZAG7* | VChr13c* | VVIv33* | VVIb09* | VChr9b* | VChr15b* | VVIv51 | VRG2 | VVIn57* | VVMD14 | VVIt60 | VChr14b* | VRG3 | VVIp34* | VChr13b* |
| LNZ-? | MALVASIA VOLCANICA | HETEROZYGOUS INDIVIDUAL (2 PEAK FAMILIES) | |||||||||||||||||||||
| FTV-? | LISTAN PRIETO | HETEROZYGOUS INDIVIDUAL (2 PEAK FAMILIES). NUMERICAL VARIATION | |||||||||||||||||||||
| LNZ-18 | MUSCAT OF ALEXANDRIA | HOMOZYGOUS INDIVIDUAL (1 PEAK FAMILY) | |||||||||||||||||||||
| LNZ-52 | HOMOZYGOUS INDIVIDUAL (1 PEAK FAMILY). NUMERICAL VARIATION | ||||||||||||||||||||||
| LNZ-87 | MALVASIA AROMATICA | TRIALLELIC INDIVIDUAL (3 PEAK FAMILIES) | |||||||||||||||||||||
| LNZ-88 | TRIALLELIC INDIVIDUAL (3 PEAK FAMILIES). NUMERICAL VARIATION | ||||||||||||||||||||||
| LNZ-89 | BERMEJUELA | TETRAALLELIC INDIVIDUAL (4 PEAK FAMILIES) | |||||||||||||||||||||
| LNZ-59 | LISTAN NEGRO | MULTIALLELIC INDIVIDUAL (MORE THAN 4 PEAK FAMILIES) | |||||||||||||||||||||
| LNZ-69 | MALVASIA FINA | * | MUTATED INDIVIDUAL | ||||||||||||||||||||
| LNZ-87 | VVS3 | VMC4F3 | VVIv33 | VVIn57 | VChr13b |
| FTV-8 | ZAG7 | VChr9b | VChr15b | VChr14b | VChr13b |
| FTV-13 | VVS3 | VVMD28 | ZAG7 | VChr15b | |
| LNZ-17 | VVS3 | VVMD32 | VChr9b | ||
| LNZ-72 | VVS3 | VVMD32 | VChr13c | ||
| FTV-1 | VVMD28 | VChr9b | VChr15b | ||
| FTV-2 | VVS3 | VVMD28 | VChr15b | ||
| FTV-7 | VVMD28 | VChr15b | VChr14b | ||
| FTV-9 | VVS3 | VVMD28 | ZAG7 | ||
| FTV-12 | VVMD28 | VChr15b | VChr14b | ||
| LNZ-3 | VVS3 | VChr9b | |||
| LNZ-21 | VVS3 | VChr9b | |||
| LNZ-29 | VVS3 | VChr9b | |||
| LNZ-32 | VVS3 | VChr9b | |||
| LNZ-35 | VVS3 | VChr9b | |||
| LNZ-45 | VVS3 | VChr9b | |||
| LNZ-47 | VVS3 | VChr9b | |||
| LNZ-48 | VVS3 | VChr9b | |||
| LNZ-50 | VVS3 | VChr9b | |||
| LNZ-54 | VVS3 | VChr9b | |||
| LNZ-55 | VVS3 | VChr9b | |||
| LNZ-56 | VVS3 | VChr9b | |||
| LNZ-68 | VVS3 | VChr9b | |||
| LNZ-78 | VVS3 | VChr9b | |||
| LNZ-80 | VVS3 | VChr9b | |||
| LNZ-83 | VVS3 | VChr9b | |||
| LNZ-84 | VVS3 | VChr9b | |||
| LNZ-85 | VVS3 | VChr9b | |||
| LNZ-5 | VVS3 | VVS29 | |||
| LNZ-34 | VVS3 | VVS29 | |||
| LNZ-36 | VVS3 | VVS29 | |||
| LNZ-42 | VVS3 | VVS29 | |||
| LNZ-66 | VVS3 | VVS29 | |||
| LNZ-6 | VVS3 | VVMD32 | |||
| LNZ-8 | VVS3 | VVMD32 | |||
| LNZ-19 | VVS3 | VChr15b | |||
| LNZ-24 | VVS29 | VChr9b | |||
| LNZ-60 | VVS3 | VVIp34 | |||
| LNZ-73 | VVS3 | VVIb09 | |||
| LNZ-76 | VVS3 | VMC4F3 | |||
| FTV-4 | VVMD28 | VChr15b | |||
| FTV-10 | VVMD28 | VChr9b | |||
| FTV-11 | VVMD28 | VChr9b | |||
| FTV-14 | VVMD28 | ZAG7 | |||
| FTV-16 | ZAG7 | VChr14b | |||
| LNZ-88 | VVS3 | VChr13b | |||
| LNZ-2 | VVS3 | ||||
| LNZ-4 | VVS3 | ||||
| LNZ-7 | VVS3 | ||||
| LNZ-9 | VVS3 | ||||
| LNZ-10 | VVS3 | ||||
| LNZ-11 | VVS3 | ||||
| LNZ-12 | VVS3 | ||||
| LNZ-13 | VVS3 | ||||
| LNZ-14 | VVS3 | ||||
| LNZ-15 | VVS3 | ||||
| LNZ-20 | VVS3 | ||||
| LNZ-22 | VVS3 | ||||
| LNZ-23 | VVS3 | ||||
| LNZ-25 | VVS3 | ||||
| LNZ-26 | VVS3 | ||||
| LNZ-27 | VVS3 | ||||
| LNZ-30 | VVS3 | ||||
| LNZ-33 | VVS3 | ||||
| LNZ-37 | VVS3 | ||||
| LNZ-38 | VVS3 | ||||
| LNZ-39 | VVS3 | ||||
| LNZ-43 | VVS3 | ||||
| LNZ-44 | VVS3 | ||||
| LNZ-49 | VVS3 | ||||
| LNZ-51 | VVS3 | ||||
| LNZ-53 | VVS3 | ||||
| LNZ-58 | VVS3 | ||||
| LNZ-61 | VVS3 | ||||
| LNZ-62 | VVS3 | ||||
| LNZ-63 | VVS3 | ||||
| LNZ-64 | VVS3 | ||||
| LNZ-67 | VVS3 | ||||
| LNZ-70 | VVS3 | ||||
| LNZ-71 | VVS3 | ||||
| LNZ-74 | VVS3 | ||||
| LNZ-75 | VVS3 | ||||
| LNZ-77 | VVS3 | ||||
| LNZ-79 | VVS3 | ||||
| LNZ-82 | VVS3 | ||||
| LNZ-86 | VVS3 | ||||
| LNZ-16 | VVIp34 | ||||
| LNZ-31 | VChr9b | ||||
| LNZ-40 | VChr9b | ||||
| LNZ-81 | VChr9b | ||||
| LNZ-41 | VVS29 | ||||
| FTV-3 | VVMD28 | ||||
| FTV-5 | VVMD28 | ||||
| FTV-18 | VVMD28 | ||||
| FTV-20 | VVMD28 | ||||
| FTV-32 | VChr9b | ||||
| LNZ-18 | VVMD5 | ||||
| LNZ-59 | VVS3 | ||||
| LNZ-69 | VVS3 | ||||
| LNZ-89 | VRG16 | ||||
| LNZ-1 | |||||
| LNZ-28 | |||||
| LNZ-46 | |||||
| LNZ-57 | |||||
| LNZ-65 | |||||
| FTV-15 | |||||
| LNZ-52 | |||||
| LNZ-? | MALVASIA VOLCANICA | ||||
| FTV-? | LISTAN PRIETO | ||||
| LNZ-18 | MUSCAT OF ALEXANDRIA | ||||
| LNZ-52 | |||||
| LNZ-87 | MALVASIA AROMATICA | ||||
| LNZ-88 | |||||
| LNZ-89 | BERMEJUELA | ||||
| LNZ-59 | LISTAN NEGRO | ||||
| LNZ-69 | MALVASIA FINA | ||||
| HETEROZYGOUS INDIVIDUAL (2 PEAK FAMILIES) | |||||
| HETEROZYGOUS INDIVIDUAL (2 PEAK FAMILIES). NUMERICAL VARIATION | |||||
| HOMOZYGOUS INDIVIDUAL (1 PEAK FAMILY) | |||||
| HOMOZYGOUS INDIVIDUAL (1 PEAK FAMILY). NUMERICAL VARIATION | |||||
| TRIALLELIC INDIVIDUAL (3 PEAK FAMILIES) | |||||
| TRIALLELIC INDIVIDUAL (3 PEAK FAMILIES). NUMERICAL VARIATION | |||||
| TETRAALLELIC INDIVIDUAL (4 PEAK FAMILIES) | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fort, F.; Suárez-Abreu, L.R.; Lin-Yang, Q.; Deis, L.; Canals, J.M.; Zamora, F. Preliminary Clonal Characterization of Malvasia Volcanica and Listan Prieto by Simple Sequence Repeat (SSR) Markers in Free-Phylloxera Volcanic Vineyards (Lanzarote and Fuerteventura (Canary Island, Spain)). Horticulturae 2025, 11, 823. https://doi.org/10.3390/horticulturae11070823
Fort F, Suárez-Abreu LR, Lin-Yang Q, Deis L, Canals JM, Zamora F. Preliminary Clonal Characterization of Malvasia Volcanica and Listan Prieto by Simple Sequence Repeat (SSR) Markers in Free-Phylloxera Volcanic Vineyards (Lanzarote and Fuerteventura (Canary Island, Spain)). Horticulturae. 2025; 11(7):823. https://doi.org/10.3390/horticulturae11070823
Chicago/Turabian StyleFort, Francesca, Luis Ricardo Suárez-Abreu, Qiying Lin-Yang, Leonor Deis, Joan Miquel Canals, and Fernando Zamora. 2025. "Preliminary Clonal Characterization of Malvasia Volcanica and Listan Prieto by Simple Sequence Repeat (SSR) Markers in Free-Phylloxera Volcanic Vineyards (Lanzarote and Fuerteventura (Canary Island, Spain))" Horticulturae 11, no. 7: 823. https://doi.org/10.3390/horticulturae11070823
APA StyleFort, F., Suárez-Abreu, L. R., Lin-Yang, Q., Deis, L., Canals, J. M., & Zamora, F. (2025). Preliminary Clonal Characterization of Malvasia Volcanica and Listan Prieto by Simple Sequence Repeat (SSR) Markers in Free-Phylloxera Volcanic Vineyards (Lanzarote and Fuerteventura (Canary Island, Spain)). Horticulturae, 11(7), 823. https://doi.org/10.3390/horticulturae11070823