
Characterization of the Glutamate Decarboxylase (GAD) Gene and Functional Analysis of DlGAD3 in the Accumulation of γ-Aminobutyric Acid in Longan (Dimocarpus longan Lour.) Pulp

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
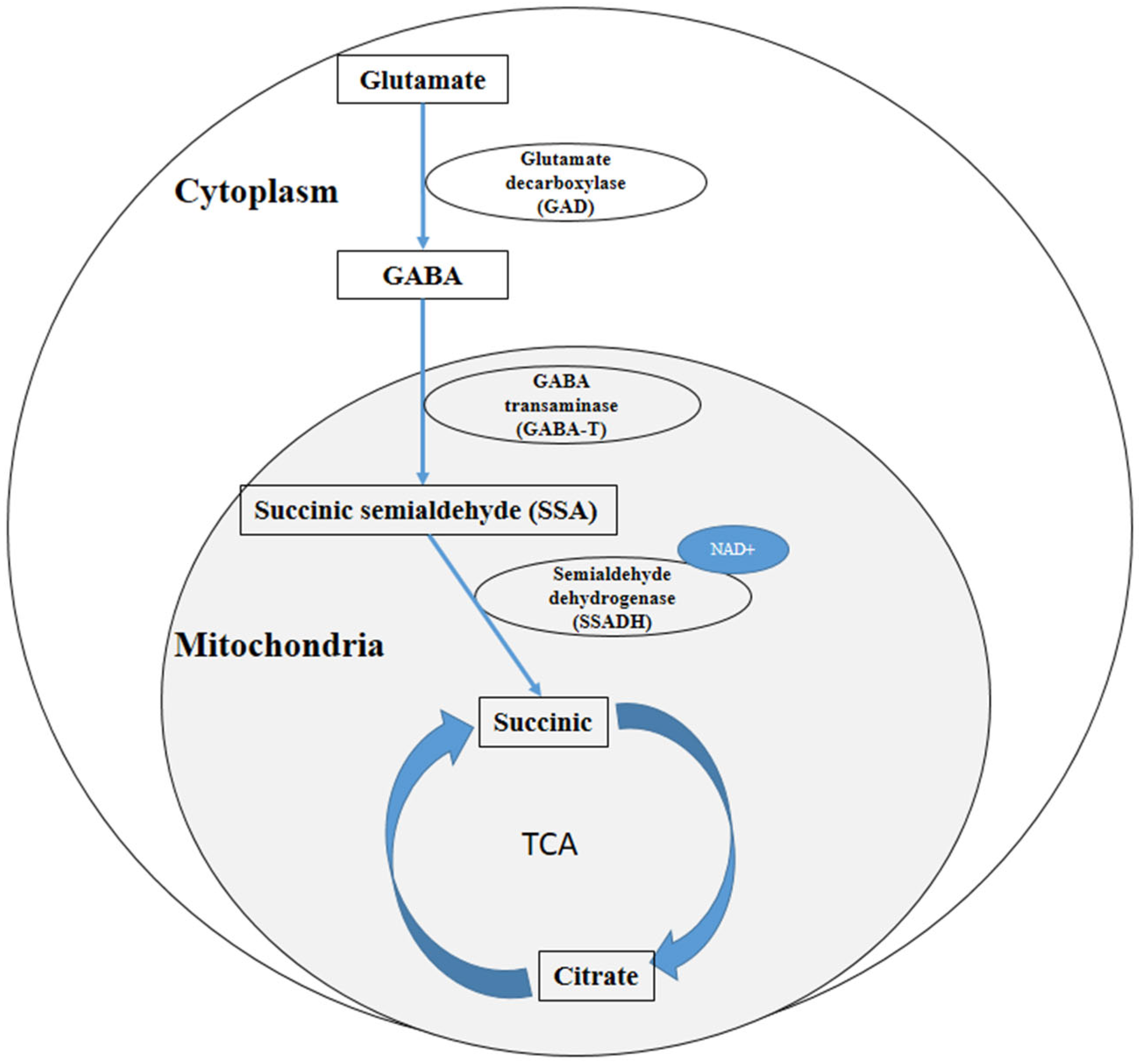
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Samples Preparation of Metabolomics
2.3. Metabolomics Analysis
2.4. Measurement of GABA Content
2.5. Determination of GAD and GABA-T Activities
2.6. RT-qPCR Analysis
2.7. Identification of GAD in Longan and Construction of Phylogenetic Tree
2.8. Transient Expression Analysis and Subcellular Localization
2.9. Statistical Analysis
3. Results
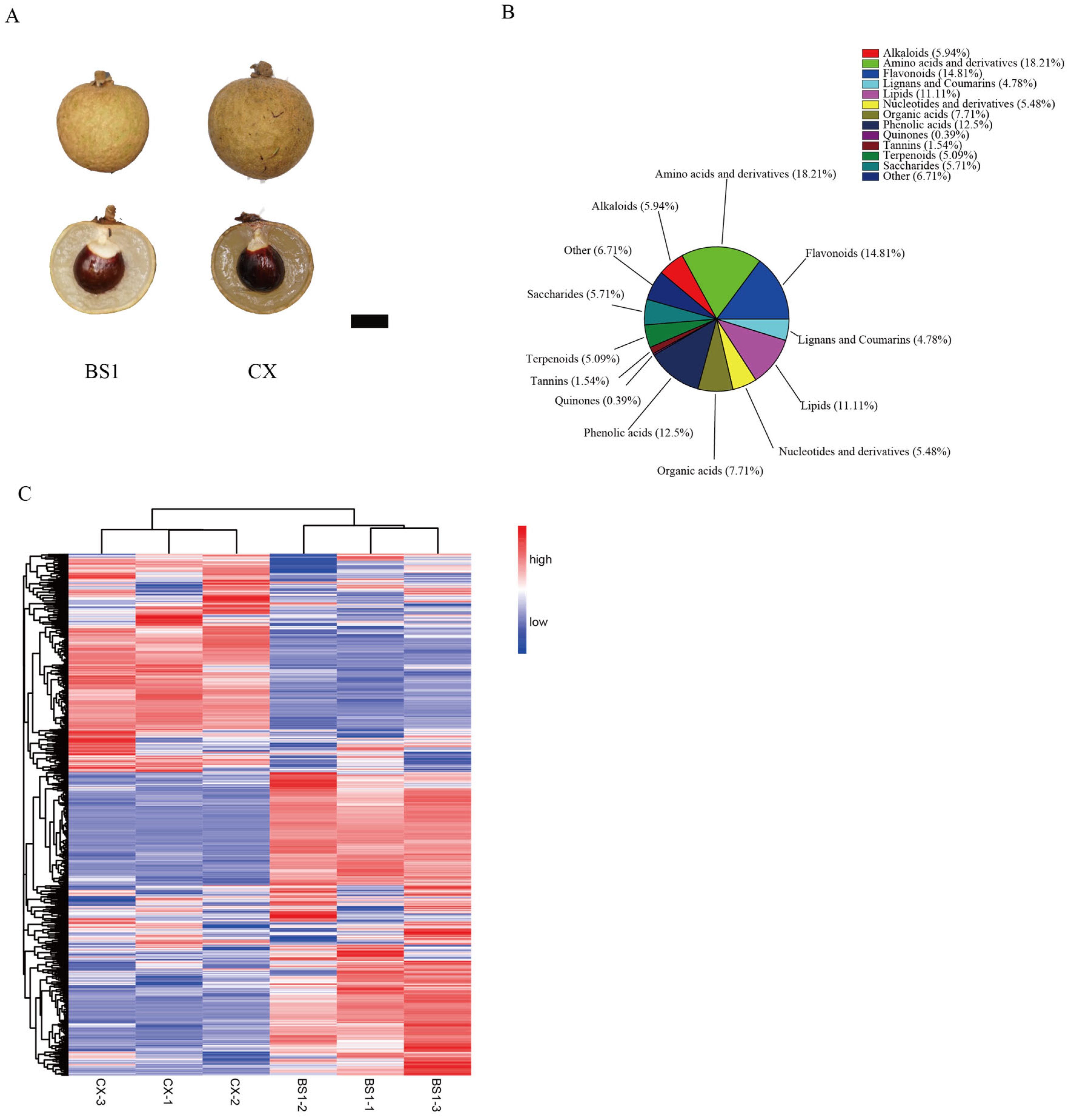
3.1. The Metabolomics Profile of the Two Longan Cultivars
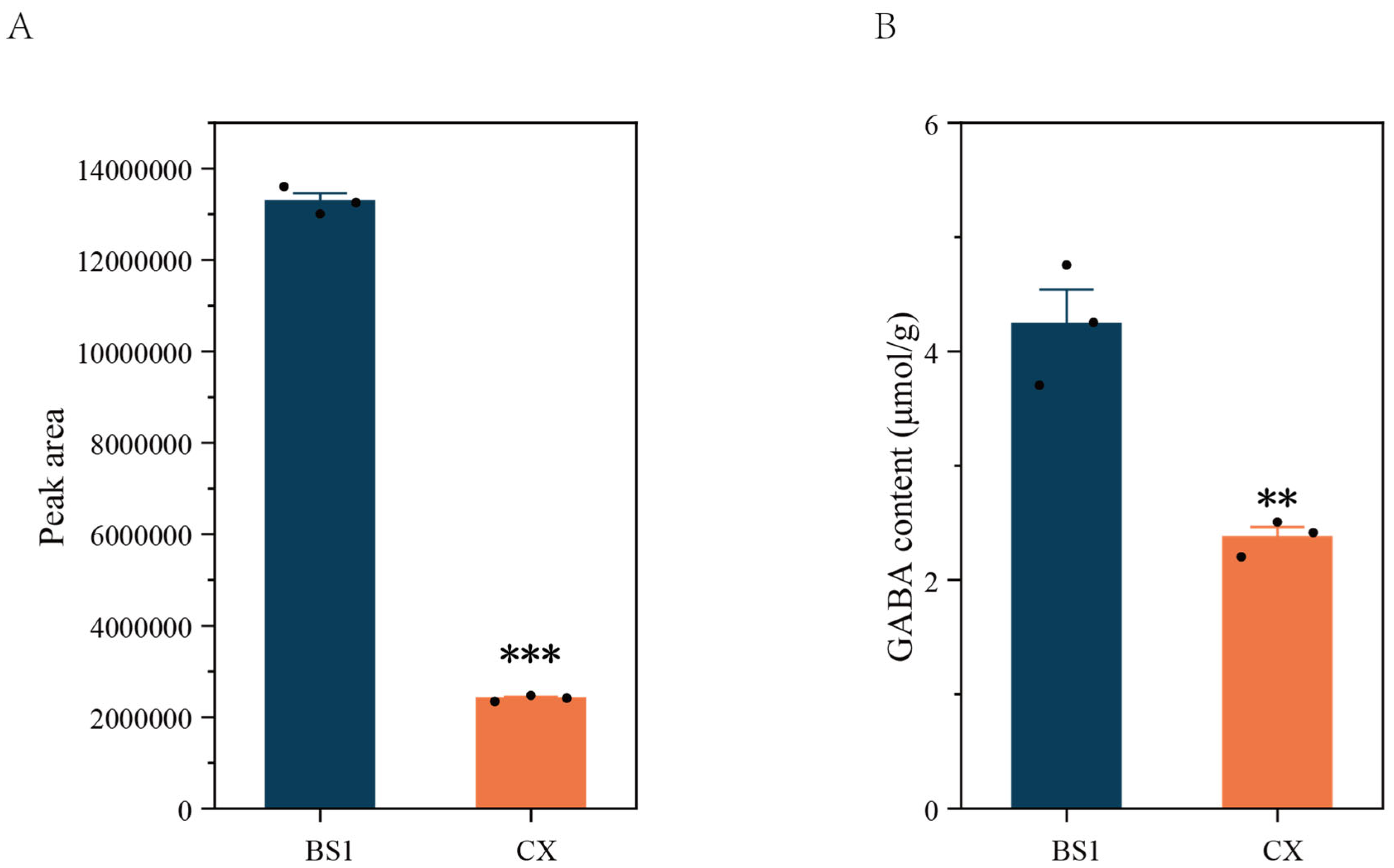
3.2. Differences in GABA Content Between Two Longan Pulps
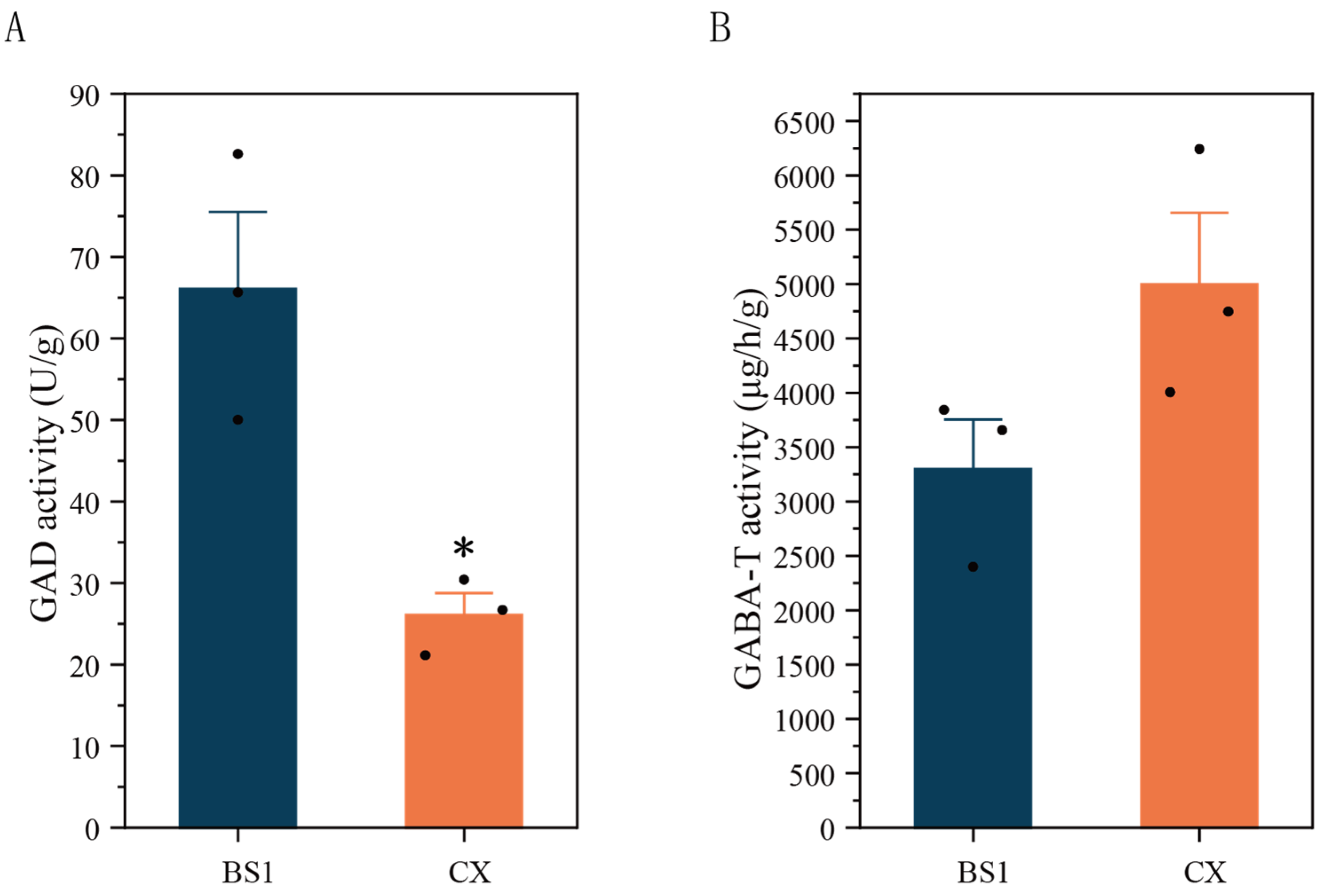
3.3. Differences in GAD and GABA-T Enzyme Activities Between Two Longan Pulps
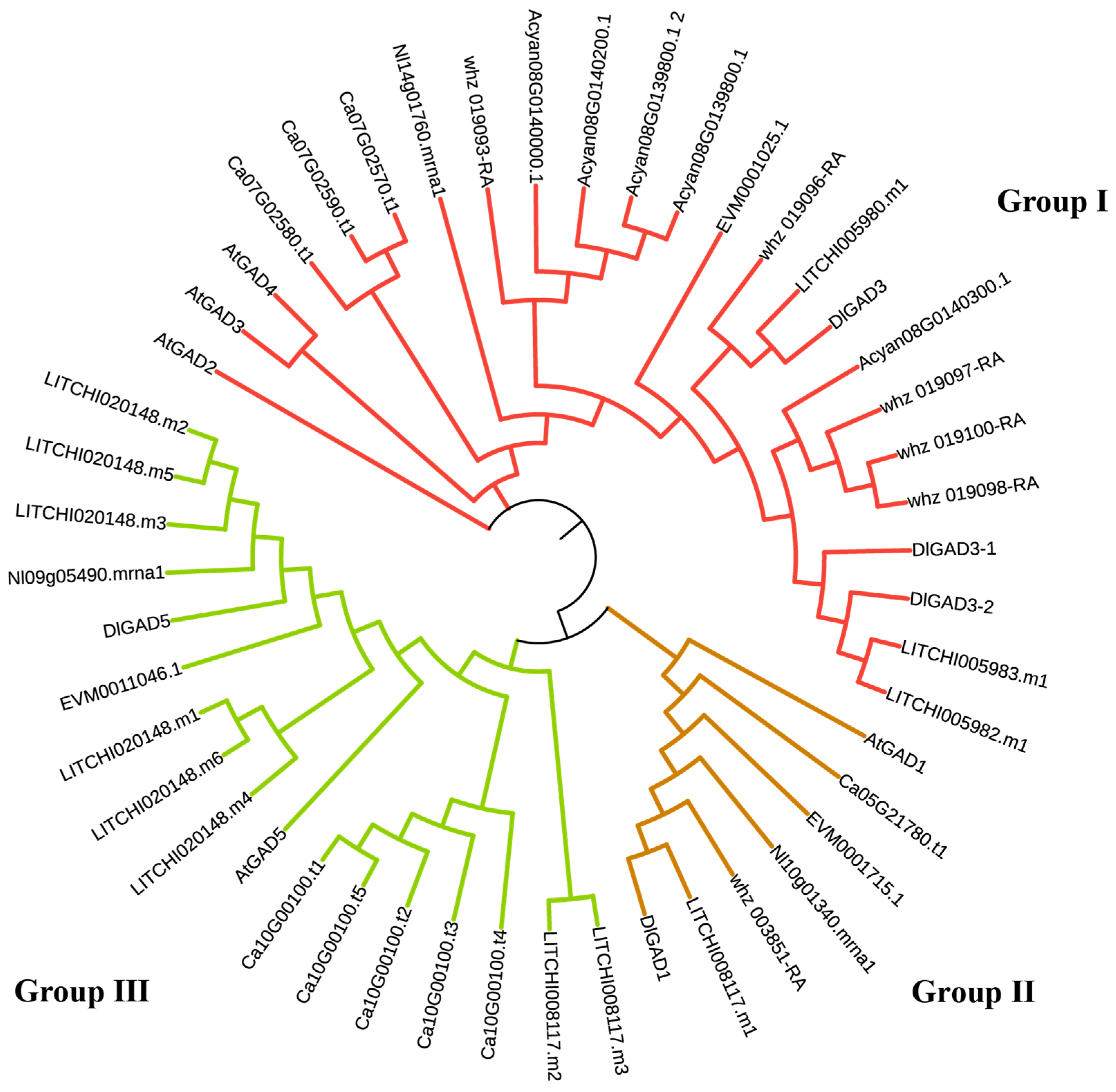
3.4. Identification of GAD and Construction of Phylogenetic Tree for DlGADs
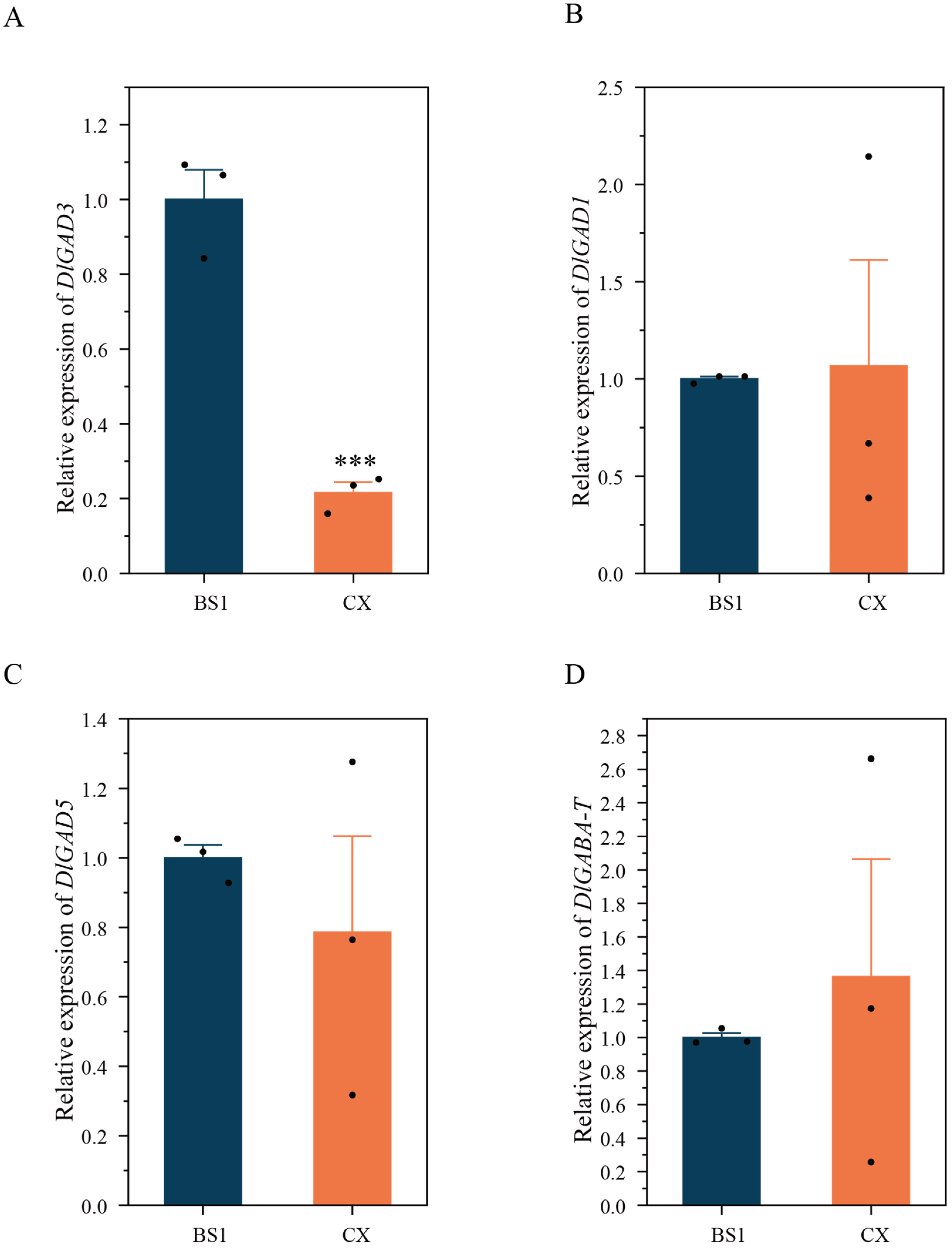
3.5. Gene Expression of GAD and GABA-T in Longan Pulp
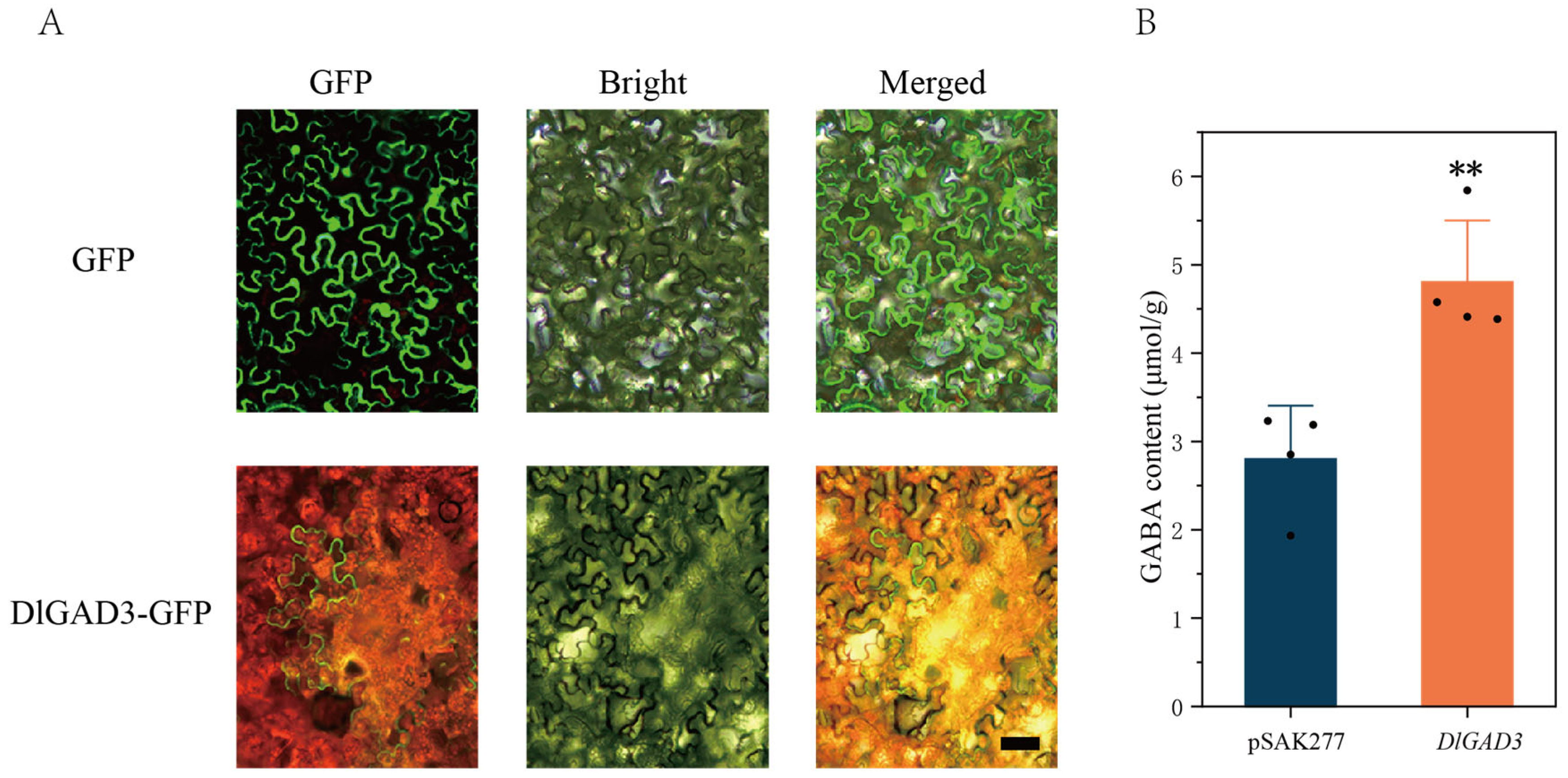
3.6. The Subcellular Localization and Functional Analysis of the DlGAD3 Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| GAD | glutamate decarboxylase |
| BS1 | Baoshi No. 1 |
| CX | Chunxiang |
| GABA | γ-aminobutyric acid |
| TCA | tricarboxylic acid |
| SSA | succinic semialdehyde |
| GABA-T | GABA transaminase |
| SSADH | semialdehyde dehydrogenase |
References
- Xu, X.; Zhang, C.; Lai, C.; Zhang, Z.; Wu, J.; Su, Q.; Gan, Y.; Zhang, Z.; Chen, Y.; Guo, R.; et al. Genome-Wide Identification and Expression Analysis of Bx Involved in Benzoxazinoids Biosynthesis Revealed the Roles of DIMBOA during Early Somatic Embryogenesis in Dimocarpus longan Lour. Plants 2024, 13, 1373. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xie, H.; Tang, J.; Lin, M.; Hung, Y.C.; Lin, H. Effects of acidic electrolyzed water treatment on storability, quality attributes and nutritive properties of longan fruit during storage. Food Chem. 2020, 320, 126641. [Google Scholar] [CrossRef]
- Tan, S.; Ke, Z.; Zhou, C.; Luo, Y.; Ding, X.; Luo, G.; Li, W.; Shi, S. Polyphenol Profile, Antioxidant Activity, and Hypolipidemic Effect of Longan Byproducts. Molecules 2023, 28, 2083. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.Y.; He, X.M.; Sun, J.; Li, C.B.; Li, L.; Sheng, J.F.; Xin, M.; Li, Z.C.; Zheng, F.J.; Liu, G.M.; et al. Polyphenols and Alkaloids in Byproducts of Longan Fruits (Dimocarpus Longan Lour.) and Their Bioactivities. Molecules 2019, 24, 1186. [Google Scholar] [CrossRef]
- Park, S.J.; Park, D.H.; Kim, D.H.; Lee, S.; Yoon, B.H.; Jung, W.Y.; Lee, K.T.; Cheong, J.H.; Ryu, J.H. The memory-enhancing effects of Euphoria longan fruit extract in mice. J. Ethnopharmacol. 2010, 128, 160–165. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, S.; Ho, C.-T.; Bai, N. Phytochemical constituents and biological activities of longan (Dimocarpus longan Lour.) fruit: A review. Food Sci. Hum. Wellness 2020, 9, 95–102. [Google Scholar] [CrossRef]
- Hu, Y.; Huang, X.; Xiao, Q.; Wu, X.; Tian, Q.; Ma, W.; Shoaib, N.; Liu, Y.; Zhao, H.; Feng, Z.; et al. Advances in Plant GABA Research: Biological Functions, Synthesis Mechanisms and Regulatory Pathways. Plants 2024, 13, 2891. [Google Scholar] [CrossRef]
- Li, L.; Dou, N.; Zhang, H.; Wu, C. The versatile GABA in plants. Plant Signal. Behav. 2021, 16, 1862565. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Fariduddin, Q. Deciphering the enigmatic role of gamma-aminobutyric acid (GABA) in plants: Synthesis, transport, regulation, signaling, and biological roles in interaction with growth regulators and abiotic stresses. Plant Physiol. Biochem. 2024, 208, 108502. [Google Scholar] [CrossRef]
- Wu, X.; Huo, R.; Yuan, D.; Zhao, L.; Kang, X.; Gong, B.; Lü, G.; Gao, H. Exogenous GABA improves tomato fruit quality by contributing to regulation of the metabolism of amino acids, organic acids and sugars. Sci. Hortic. 2024, 338, 113750. [Google Scholar] [CrossRef]
- Lorente-Mento, J.M.; Guillén, F.; Martínez-Romero, D.; Carrión-Antoli, A.; Valero, D.; Serrano, M. γ-Aminobutyric acid treatments of pomegranate trees increase crop yield and fruit quality at harvest. Sci. Hortic. 2023, 309, 111633. [Google Scholar] [CrossRef]
- Yang, X.; Liu, Y.; Fu, C.X.; Chu, Y.H.; Chen, Q.; Wang, H.; Wei, D.X.; Yao, Y.F. Selectively Probing the Magnetic Resonance Signals of γ-Aminobutyric Acid in Human Brains In Vivo. J. Magn. Reson. Imaging 2024, 59, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Heli, Z.; Hongyu, C.; Dapeng, B.; Yee Shin, T.; Yejun, Z.; Xi, Z.; Yingying, W. Recent advances of γ-aminobutyric acid: Physiological and immunity function, enrichment, and metabolic pathway. Front. Nutr. 2022, 9, 1076223. [Google Scholar] [CrossRef] [PubMed]
- Arora, I.; Mal, P.; Arora, P.; Paul, A.; Kumar, M. GABAergic implications in anxiety and related disorders. Biochem. Biophys. Res. Commun. 2024, 724, 150218. [Google Scholar] [CrossRef]
- Abdou, A.M.; Higashiguchi, S.; Horie, K.; Kim, M.; Hatta, H.; Yokogoshi, H. Relaxation and immunity enhancement effects of γ-Aminobutyric acid (GABA) administration in humans. Biofactors 2006, 26, 201–208. [Google Scholar] [CrossRef]
- Wang, D.W.; Deng, Y.N.; Zhao, L.; Wang, K.; Wu, D.M.; Hu, Z.Y.; Liu, X.W. GABA and fermented litchi juice enriched with GABA promote the beneficial effects in ameliorating obesity by regulating the gut microbiota in HFD-induced mice. Food Funct. 2023, 14, 8170–8185. [Google Scholar] [CrossRef]
- Shelp, B.J.; Bozzo, G.G.; Trobacher, C.P.; Zarei, A.; Deyman, K.L.; Brikis, C.J. Hypothesis/review: Contribution of putrescine to 4-aminobutyrate (GABA) production in response to abiotic stress. Plant Sci. 2012, 193–194, 130–135. [Google Scholar] [CrossRef]
- Chen, M.; Zhu, C.; Zhang, H.; Chen, S.; Wang, X.; Gan, L. Endogenous γ-Aminobutyric Acid Accumulation Enhances Salinity Tolerance in Rice. Plants 2024, 13, 2750. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, Z.; Mao, L.; Ying, T. Contribution of polyamines metabolism and GABA shunt to chilling tolerance induced by nitric oxide in cold-stored banana fruit. Food Chem. 2016, 197, 333–339. [Google Scholar] [CrossRef]
- Zik, M.; Arazi, T.; Snedden, W.A.; Fromm, H. Two isoforms of glutamate decarboxylase in Arabidopsis are regulated by calcium/calmodulin and differ in organ distribution. Plant Mol. Biol. 1998, 37, 967–975. [Google Scholar] [CrossRef]
- Akihiro, T.; Koike, S.; Tani, R.; Tominaga, T.; Watanabe, S.; Iijima, Y.; Aoki, K.; Shibata, D.; Ashihara, H.; Matsukura, C.; et al. Biochemical mechanism on GABA accumulation during fruit development in tomato. Plant Cell Physiol. 2008, 49, 1378–1389. [Google Scholar] [CrossRef] [PubMed]
- Al-Quraan, N.A.; Sartawe, F.A.-b.; Qaryouti, M.M. Characterization of γ-aminobutyric acid metabolism and oxidative damage in wheat (Triticum aestivum L.) seedlings under salt and osmotic stress. J. Plant Physiol. 2013, 170, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; He, Y.; Cui, A.; Sun, L.; Han, M.; Wang, J.; Rui, C.; Lei, Y.; Liu, X.; Xu, N.; et al. Genome-wide identification of GAD family genes suggests GhGAD6 functionally respond to Cd2+ stress in cotton. Front. Genet. 2022, 13, 965058. [Google Scholar] [CrossRef]
- Scholz, S.S.; Reichelt, M.; Mekonnen, D.W.; Ludewig, F.; Mithöfer, A. Insect Herbivory-Elicited GABA Accumulation in Plants is a Wound-Induced, Direct, Systemic, and Jasmonate-Independent Defense Response. Front. Plant Sci. 2015, 6, 1128. [Google Scholar] [CrossRef] [PubMed]
- Benidickson, K.H.; Raytek, L.M.; Hoover, G.J.; Flaherty, E.J.; Shelp, B.J.; Snedden, W.A.; Plaxton, W.C. Glutamate decarboxylase-1 is essential for efficient acclimation of Arabidopsis thaliana to nutritional phosphorus deprivation. New Phytol. 2023, 240, 2372–2385. [Google Scholar] [CrossRef]
- Bouché, N.; Fait, A.; Zik, M.; Fromm, H. The root-specific glutamate decarboxylase (GAD1) is essential for sustaining GABA levels in Arabidopsis. Plant Mol. Biol. 2004, 55, 315–325. [Google Scholar] [CrossRef]
- Xu, B.; Feng, X.; Piechatzek, A.; Zhang, S.; Konrad, K.R.; Kromdijk, J.; Hedrich, R.; Gilliham, M. The GABA shunt contributes to ROS homeostasis in guard cells of Arabidopsis. New Phytol. 2023, 241, 73–81. [Google Scholar] [CrossRef]
- Xu, B.; Long, Y.; Feng, X.; Zhu, X.; Sai, N.; Chirkova, L.; Betts, A.; Herrmann, J.; Edwards, E.J.; Okamoto, M.; et al. GABA signalling modulates stomatal opening to enhance plant water use efficiency and drought resilience. Nat. Commun. 2021, 12, 1952. [Google Scholar] [CrossRef] [PubMed]
- Akter, N.; Kulsum, U.; Moniruzzaman, M.; Yasuda, N.; Akama, K. Truncation of the calmodulin binding domain in rice glutamate decarboxylase 4 (OsGAD4) leads to accumulation of γ-aminobutyric acid and confers abiotic stress tolerance in rice seedlings. Mol. Breed. 2024, 44, 21. [Google Scholar] [CrossRef]
- Takayama, M.; Koike, S.; Kusano, M.; Matsukura, C.; Saito, K.; Ariizumi, T.; Ezura, H. Tomato Glutamate Decarboxylase GenesSlGAD2andSlGAD3Play Key Roles in Regulating γ-Aminobutyric Acid Levels in Tomato (Solanum lycopersicum). Plant Cell Physiol. 2015, 56, 1533–1545. [Google Scholar] [CrossRef]
- Zeng, S.; Wang, K.; Liu, X.; Hu, Z.; Zhao, L. Potential of longan (Dimocarpus longan Lour.) in functional food: A review of molecular mechanism-directing health benefit properties. Food Chem. 2024, 437, 137812. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Ndeurumio, K.H.; Zhao, L.; Hu, Z. Impact of Precooling and Controlled-Atmosphere Storage on gamma-Aminobutyric Acid (GABA) Accumulation in Longan (Dimocarpus longan Lour.) Fruit. J. Agric. Food Chem. 2016, 64, 6443–6450. [Google Scholar] [CrossRef]
- Zheng, S.Q.; Jiang, F.; Huang, A.P.; Gao, H.Y.; Deng, C.J.; Chen, X.P.; Zheng, J.G. Evaluating of γ-aminobutyric Acid in Different Genotypes from Longan (Dimocarpus longan Lour.) peel. Fujian Fruits 2008, 3, 23–27. [Google Scholar]
- Lan, S.; Zhai, T.; Zhang, X.; Xu, L.; Gao, J.; Lai, C.; Chen, Y.; Lai, Z.; Lin, Y. Genome-wide identification and expression analysis of the GAD family reveal their involved in embryogenesis and hormones responses in Dimocarpus longan Lour. Gene 2024, 927, 148698. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Wu, Y.; Li, C.; Li, X.; Li, K.; Yang, W.; Liang, Y.; Lu, J.; Zhang, L.; Lu, P.; et al. Metabolomics and quantitative descriptive analysis reveal the relationship between metabolites and taste attributes of flowers in two types of albino tea cultivars. LWT 2024, 199, 116074. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Y.; Jiang, X.; Du, B.; Wang, Q.; Ma, Y.; Liu, M.; Mao, Y.; Yang, J.; Li, F.; et al. Uncovering nutritional metabolites and candidate genes involved in flavonoid metabolism in Houttuynia cordata through combined metabolomic and transcriptomic analyses. Plant Physiol. Biochem. 2023, 203, 108059. [Google Scholar] [CrossRef]
- Lin, Y.L.; Lai, Z.X. Reference gene selection for qPCR analysis during somatic embryogenesis in longan tree. Plant Sci. 2010, 178, 359–365. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Priyam, A.; Woodcroft, B.J.; Rai, V.; Moghul, I.; Munagala, A.; Ter, F.; Chowdhary, H.; Pieniak, I.; Maynard, L.J.; Gibbins, M.A.; et al. Sequenceserver: A Modern Graphical User Interface for Custom BLAST Databases. Mol. Biol. Evol. 2019, 36, 2922–2924. [Google Scholar] [CrossRef]
- Huang, G.J.; Wang, B.S.; Lin, W.C.; Huang, S.S.; Lee, C.Y.; Yen, M.T.; Huang, M.H. Antioxidant and Anti-Inflammatory Properties of Longan (Dimocarpus longan Lour.) Pericarp. Evid. Based Complement Altern. Med. 2012, 2012, 709483. [Google Scholar] [CrossRef]
- Yang, M.; Yan, T.; Yu, M.; Kang, J.; Gao, R.; Wang, P.; Zhang, Y.; Zhang, H.; Shi, L. Advances in understanding of health-promoting benefits of medicine and food homology using analysis of gut microbiota and metabolomics. Food Front. 2020, 1, 398–419. [Google Scholar] [CrossRef]
- Lai, T.; Shuai, L.; Han, D.; Lai, Z.; Du, X.; Guo, X.; Hu, W.; Wu, Z.; Luo, T. Comparative metabolomics reveals differences in primary and secondary metabolites between “Shixia” and “Chuliang” longan (Dimocarpus longan Lour.) pulp. Food Sci. Nutr. 2021, 9, 5785–5799. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.; Riechman, S.E.; Fluckey, J.D.; Wu, G. Interorgan Metabolism of Amino Acids in Human Health and Disease. Adv. Exp. Med. Biol. 2021, 1332, 129–149. [Google Scholar] [CrossRef]
- Ling, Z.N.; Jiang, Y.F.; Ru, J.N.; Lu, J.H.; Ding, B.; Wu, J. Amino acid metabolism in health and disease. Signal Transduct. Target. Ther. 2023, 8, 345. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhu, L.; Li, H.; Chen, Q.; Li, N.; Li, J.; Zhao, Z.; Xiao, D.; Tang, T.; Bi, C.; et al. Insights and progress on the biosynthesis, metabolism, and physiological functions of gamma-aminobutyric acid (GABA): A review. PeerJ 2024, 12, e18712. [Google Scholar] [CrossRef]
- Rema Shree, A.B.; Anju, P.; Moothedath, I. Gamma amino butyric acid accumulation in medicinal plants without stress. Anc. Sci. Life 2014, 34, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, U.; Nohmi, T.; Sagane, R.; Hai, J.; Ohbayashi, K.; Miyazaki, M.; Yamatsu, A.; Kim, M.; Iwasaki, Y. Dietary Gamma-Aminobutyric Acid (GABA) Induces Satiation by Enhancing the Postprandial Activation of Vagal Afferent Nerves. Nutrients 2022, 14, 2492. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, Z.; Zhang, N.; Liang, Y.; Gong, Z.; Wang, J.; Ditta, A.; Sang, Z.; Wang, J.; Li, X. Identification and function analysis of GABA branch three gene families in the cotton related to abiotic stresses. BMC Plant Biol. 2024, 24, 57. [Google Scholar] [CrossRef]
- Yan, L.; Zheng, H.H.; Liu, W.; Liu, C.H.; Jin, T.; Liu, S.; Zheng, L. UV-C treatment enhances organic acids and GABA accumulation in tomato fruits during storage. Food Chem. 2021, 338, 128126. [Google Scholar] [CrossRef]
- Deewatthanawong, R.; Nock, J.F.; Watkins, C.B. γ-Aminobutyric acid (GABA) accumulation in four strawberry cultivars in response to elevated CO2 storage. Postharvest. Biol. Technol. 2010, 57, 92–96. [Google Scholar] [CrossRef]
- Li, J.; Zhou, Q.; Zhou, X.; Wei, B.; Zhao, Y.; Ji, S. Calcium Treatment Alleviates Pericarp Browning of ‘Nanguo’ Pears by Regulating the GABA Shunt After Cold Storage. Front. Plant Sci. 2020, 11, 580986. [Google Scholar] [CrossRef] [PubMed]
- Bai, P.; Wang, J.W.; He, Y.M.; Feng, J.F.; Li, J.L.; Shang, X.P.; Wu, Y.; Yu, J.H.; Tang, Z.Q.; Xie, J.M. Exogenous ALA applied on different plant parts promotes tomato fruit quality and GABA synthesis. Front. Nutr. 2025, 11, 1520634. [Google Scholar] [CrossRef] [PubMed]
- Shelp, B.J.; Bown, A.W.; McLean, M.D. Metabolism and functions of gamma-aminobutyric acid. Trends Plant Sci. 1999, 4, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.X.; Xu, X.W.; Liu, Y.; Zhang, Y.; Yang, L.Y.; Zhang, S.Q.; Xu, J. Induction of γ-aminobutyric acid plays a positive role to Arabidopsis resistance against Pseudomonas syringae. J. Integr. Plant Biol. 2020, 62, 1797–1812. [Google Scholar] [CrossRef]
- Jiao, W.H.; Wei, Q.L.; Chen, J.H.; Cao, S.F.; Luo, M.; Qian, Y.A.; Chen, Y.; Wei, Y.Y.; Shao, X.F.; Xu, F. Identification analysis of GAD gene family, and the role of BoGAD 5 in GABA enrichment in broccoli sprouts. Plant Growth Regul. 2024, 104, 1643–1655. [Google Scholar] [CrossRef]
- Liu, X.; Ma, H.; Liu, J.; Liu, D.H.; Wang, C.L. The ?-Aminobutyric Acid (GABA) Synthesis Gene Regulates the Resistance to Water Core-Induced Hypoxia Stress for Pear Fruits. Agronomy 2023, 13, 1062. [Google Scholar] [CrossRef]
- Koike, S.; Matsukura, C.; Takayama, M.; Asamizu, E.; Ezura, H. Suppression of γ-Aminobutyric Acid (GABA) Transaminases Induces Prominent GABA Accumulation, Dwarfism and Infertility in the Tomato (Solanum lycopersicum L.). Plant Cell Physiol. 2013, 54, 793–807. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, W.; Zhang, T.; Chen, Y.; Zhou, Z.; Su, W.; Xu, Q.; Zhang, Y.; Zheng, S.; Jiang, J.; Deng, C. Characterization of the Glutamate Decarboxylase (GAD) Gene and Functional Analysis of DlGAD3 in the Accumulation of γ-Aminobutyric Acid in Longan (Dimocarpus longan Lour.) Pulp. Horticulturae 2025, 11, 686. https://doi.org/10.3390/horticulturae11060686
Wei W, Zhang T, Chen Y, Zhou Z, Su W, Xu Q, Zhang Y, Zheng S, Jiang J, Deng C. Characterization of the Glutamate Decarboxylase (GAD) Gene and Functional Analysis of DlGAD3 in the Accumulation of γ-Aminobutyric Acid in Longan (Dimocarpus longan Lour.) Pulp. Horticulturae. 2025; 11(6):686. https://doi.org/10.3390/horticulturae11060686
Chicago/Turabian StyleWei, Weilin, Tingting Zhang, Yongping Chen, Ziqi Zhou, Wenbing Su, Qizhi Xu, Yaling Zhang, Shaoquan Zheng, Jimou Jiang, and Chaojun Deng. 2025. "Characterization of the Glutamate Decarboxylase (GAD) Gene and Functional Analysis of DlGAD3 in the Accumulation of γ-Aminobutyric Acid in Longan (Dimocarpus longan Lour.) Pulp" Horticulturae 11, no. 6: 686. https://doi.org/10.3390/horticulturae11060686
APA StyleWei, W., Zhang, T., Chen, Y., Zhou, Z., Su, W., Xu, Q., Zhang, Y., Zheng, S., Jiang, J., & Deng, C. (2025). Characterization of the Glutamate Decarboxylase (GAD) Gene and Functional Analysis of DlGAD3 in the Accumulation of γ-Aminobutyric Acid in Longan (Dimocarpus longan Lour.) Pulp. Horticulturae, 11(6), 686. https://doi.org/10.3390/horticulturae11060686