1. Introduction
The genus
Coffea, a member of the Rubiaceae family, comprises approximately 130 described species, of which only
Coffea arabica and
Coffea canephora are cultivated on a large scale [
1]. Among these,
C. canephora, commonly known as conilon or robusta, accounts for approximately 40% of global coffee production and is widely grown in tropical regions, with Vietnam and Brazil standing out as major producers [
2,
3]. In Brazil, the species is primarily cultivated in the states of Espírito Santo, Rondônia, and Bahia, placing the country as the world’s second-largest producer of
C. canephora [
4].
C. canephora is a diploid species (2n = 2x = 22), perennial, vigorous in growth, and allogamous. Sexual reproduction via self-pollination is not viable due to the presence of a gametophytic self-incompatibility system [
5]. Due to its hardiness and wide phenotypic plasticity,
C. canephora has proven to be essential for meeting market demands, especially under conditions of climatic instability. Recent studies have shown that certain genotypes exhibit high adaptability to diverse environmental conditions, maintaining yield and beverage quality even under abiotic stresses [
6].
Traditionally,
C. canephora is cultivated in low-altitude regions (up to 500 m of altitude), with annual average temperatures ranging between 22 °C and 26 °C and water deficits below 200 mm [
7]. However, future projections indicate that climate change—such as rising temperatures and irregular rainfall patterns—may significantly reduce the land currently suitable for
C. canephora cultivation in Brazil [
8,
9]. In this context, regions above 500 m in elevation, previously considered marginal for conilon cultivation, are now being explored as viable alternatives.
Recent studies, such as those by [
10,
11] suggest that the transition altitude for
C. canephora cultivation lies between 500 and 700 m. The intraspecific genetic variability observed in
C. canephora offers opportunities for the selection and development of cultivars better adapted to the edaphoclimatic conditions of transitional altitude environments [
12,
13]. In addition, irrigated management strategies are essential to ensure the sustainability of coffee production under the new climatic demands [
14,
15].
Climate change impact projections on coffee cultivation indicate significant losses in traditionally suitable areas for C. canephora production in Brazil, particularly in low-altitude, warmer regions. In this scenario, identifying genotypes adaptable to distinct edaphoclimatic conditions—such as those found at transitional altitudes—becomes crucial for maintaining the sustainability and competitiveness of the coffee sector. Nonetheless, systematic studies assessing the agronomic performance and stress tolerance of conilon genotypes in these emerging agricultural frontiers remain scarce. Therefore, expanding knowledge on the agronomic performance of C. canephora genotypes in such environments is essential to provide technical support for crop management and breeding in areas previously considered unsuitable for the species.
2. Materials and Methods
2.1. Local Characterization
The experiment was conducted on farmland in the municipality of Alegre, in the mountainous region of the state of Espírito Santo (Caparaó), in the southeast region of Brazil (20°52′6″ S, 4°28′45″ W), and at ca. 650 m above sea level, which is classified as marginally suitable for the cultivation of
C. canephora, according to the current agricultural climate risk zoning [
15].
The climate of the location is classified as
Cwa, subtropical humid, according to the Koppen climate classification [
16], and is characterized by rainy summers and dry winters. The topography of the location displays a wavy relief, and the soil is classified as typic hapludox [
17].
2.2. Crop Management
The clonal seedlings were planted in 2015 and subjected to bending of orthotropic stems after 50 days. The bending is a technique used to stimulate the growth of shoots, increasing the number of suitable orthotropic stems per plant and aiding in selecting and establishing the planned number of stems per area at the formation of the plantation [
18]. Each plant was cultivated with three orthotropic stems, arranged in a 3.0 × 1.0 m spacing, resulting in a density of 3333 plants/ha and 9999 orthotropic stems/ha. This total population of plants and stems per area is within the recommended density for
C. canephora in the region [
18].
The crop management followed the recommended practices for the cultivation of this species in the state of Espírito Santo [
18]. The nutritional management was based on soil analysis; in order to correct the soil acidity (liming) and to elevate the fertility to match the nutritional demands of the plants, the required fertilizers were added through fertigation and split into monthly parcels. The choice of fertilizers was based on higher solubility and suitability for fertigation (e.g., urea, mono-ammonium phosphate, potassium chloride, magnesium sulfate, calcium nitrate, boric acid, zinc sulfate, copper sulfate). The phytosanitary management aimed at the main problems in the region, such as leaf rust (
Hemileia vastatrix), brown eye spot (
Cercospora coffeicola), phoma leaf spot (
Phoma sp.), leaf miner (
Leucoptera coffeella), berry borer (
Hypothenemus hampei), rosette mealybug (
Planococcus citri), and cicada (
Quesada gigas), used chemical (e.g., thiamethoxam, cyproconazole, benzimidazole, strobilurin and tebuconazole) and biological (e.g.,
Bacillus thuringiensis) methods. The weeds were managed using mechanical (e.g., regular mowing) and chemical (e.g., glyphosate) methods. The plant management was performed along the phenological cycle, removing spontaneous shoots during frutification (ca., three times per cycle), and removing plagiotropic branches (which were previously produced) after harvesting (ca., one time per cycle). The stages of plant management are included in the methodology of programmed cycle pruning for
C. canephora, following the standards for this species in Brazil [
18].
2.3. Experimental Design
The experiment followed a split-plot design, with two hydric managements in the main plots and 27 genotypes in the subplots. The treatments were arranged in a completely randomized block design, with four repetitions and three plants per experimental plot. The values for each parcel corresponded to the mean of the three plants per plot for all evaluated parameters.
2.4. Genetic Material
The tested 27 genotypes are part of three clonal cultivars certified in Brazil by the National Plant Varieties Protection Service (Serviço Nacional de Proteção de Cultivares, SNPC, Brazil). Nine genotypes originate from the cultivar ‘Diamante ES8112’ (SNPC Certification No.: 20140103) and are referred to as 101, 102, 103, 104, 105, 106, 107, 108, and 109. Nine genotypes belong to the cultivar ‘Jequitibá ES8122’ (SNPC Certification No.: 20140104) and are identified as 201, 202, 203, 204, 205, 206, 207, 208, and 209. The remaining nine genotypes are components of the cultivar ‘Centenária ES8132’ (SNPC Certification No.: 20140102) and are referred to as 301, 302, 303, 304, 305, 306, 307, 308, and 309.
These clonal cultivars were developed by the breeding program of the Instituto Capixaba de Pesquisa, Assistência Técnica e Extensão Rural (Incaper, Vitória, Brazil). They result from compatible arrangements characterized by high productivity and the potential for excellent beverage quality. The genotypes from ‘Diamante ES8112’ exhibit an early ripening cycle, while those from ‘Jequitibá ES8122’ have an intermediate ripening cycle, and the genotypes from ‘Centenária ES8132’ exhibit a late ripening cycle [
18].
The genotypes were multiplied by cloning, using sections of stems, and obtained from properly certified nurseries in the national inspection organ (Ministério da Agricultura e Pecuária, MAPA, Brasília, Brazil).
2.5. Hydric Management
The two hydric managements were established starting at the first harvest, carried out in 2017. One management was fully irrigated, managed using the current recommendations for the crop, while the other was minimally irrigated, where water was only added during fertigation.
To establish the water supply for each management, the soil was sampled and subjected to hydro-physical analysis. The soil density was 1.051 g/cm3, and its water retention curve was Y = 0.2889 X−0.121 (R2 = 98.18%), where Y represents the water availability (m3 water/m3 soil) and X represents the water tension (kPa) in the soil. The water availability in the soil at field capacity (10 kPa) was 0.2308 m3/m3 and at the permanent wilting point (1500 kPa), was 0.1561 m3/m3.
The fully irrigated treatment was implemented based on the water retention curve. The soil moisture was monitored using tensiometers, installed to monitor the first 25 cm of depth of soil within the crop and increased to the field capacity level whenever it depleted to nearly 70% of available water (34 kPa), based on the water depletion factor to avoid stresses for coffee trees, allowing for the replenishment of moisture before water deficit damages could occur [
18,
19].
Fertigation was applied on a monthly basis, using 4.29 L of water per plant (0.5 h of irrigation per month) to fully carry the fertilizers, for both managements. This was the only artificial input of water in the minimally irrigated condition. This nutrient supply by fertigation was used in both treatments to standardize the quantity, solubility, and parceling of the fertilizers.
Irrigation, for both managements, was performed using a drip system with self-compensating emitters spaced 40 cm of distance from each other, with a water flow of 3.43 L/h. The meteorological conditions during the period of the experiment were monitored and registered by an automatic weather station (Irriplus, E5000 model) installed adjacent to the experiment.
2.6. Data Collection
The canopy structure and coffee production were evaluated during the third productive cycle (2018–2019). The evaluated traits were selected and quantified following the methods for growth analysis [
20].
During the phenological stage of grain formation, for each plant, one representative orthotropic stem was selected to collect the measurements. The length of the orthotropic stem (LOS; m) was measured from the ground level to the apex of the orthotropic stem. The number of plagiotropic branches (NPBs; unit) was counted. The canopy diameter projection (CDP; m) was measured as the maximum linear distance between two opposite points of the canopy.
During the fruit ripening stage, one plagiotropic branch per plant was selected and collected from the middle portion of the canopy and evaluated to quantify the length of the plagiotropic branches (LPBs; m) using a graduated ruler; the number of nodes per plagiotropic branch (NNP; unit); the number of rosettes (considering a rosette as every reproductive knot with the presence of at least one fruit; NRs; unit); the number of leaves (NLs; unit); the number of fruits per rosette (NFR; unit); the length of plagiotropic internodes (LPIs; cm); the unitary leaf area (ULA; cm
2), estimated using the non-destructive method of the linear dimensions [
21], sampled from the third or fourth pairs of completely expanded leaves (from the apex of the plagiotropic branches); and the total leaf area per plagiotropic branches (LAPs; cm
2). The same plagiotropic branches were dried in a laboratory oven (STF SP-102/2000 CIR), with forced air circulation, at 65 ± 2 °C, until reaching a constant mass, to determine the total dry mass (TDM; g) of the plagiotropic branches.
After the ripening stage, the fruits were harvested, dried to the level of 11.5% moisture (humid base), peeled (Pinhalense, DRC1), and weighed in an electronic precision scale (0.1 mg) in order to quantify the coffee production per plant (PROD; g). A sample of 300 g of the processed coffee was submitted to grain classification through sieves in order to obtain the proportion of large coffee beans (PLBs; %), considering flat and mocha grains retained by sieve No. 17 and above.
2.7. Statistical Analyses
The data were submitted for analyses of variance and, whenever the effects of the various sources were significant, the F-test was used to compare hydric management, and the Scott-Knott criterion was used to compare genotypes, both at a 5% probability level. The correlation coefficients between variables were estimated for both hydric managements, and their significance was tested by t-test at levels of 0.1%, 1%, and 5% probability. Data analyses were performed using “R” software version 4.5.0 [
22].
3. Results
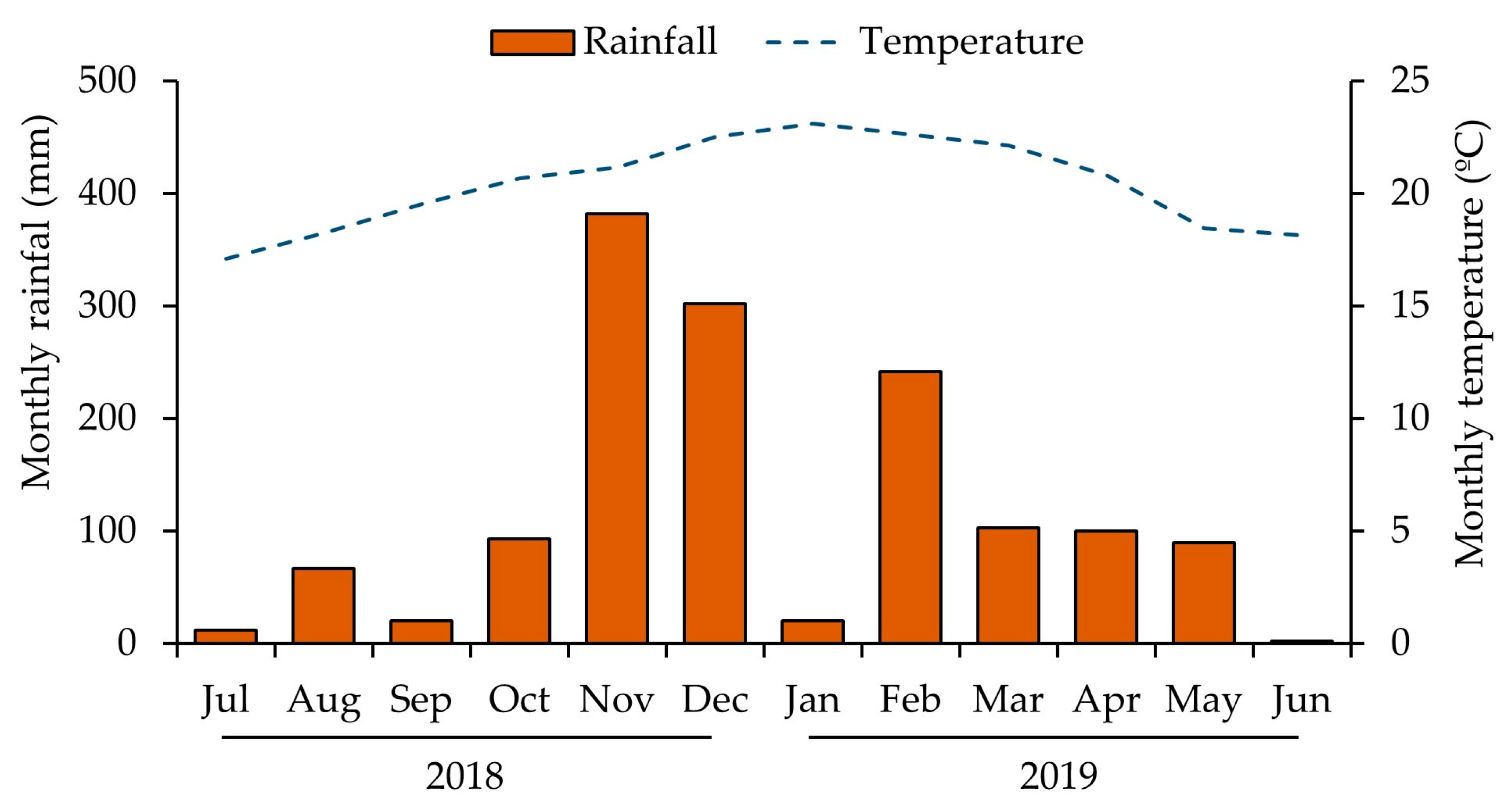
The average air temperature during the experimental period was 20.4 °C; January was the month with the highest average temperature (23.12 °C) and July with the lowest average temperature (17.11 °C). The accumulated annual precipitation was 1433 mm (
Figure 1).
The analysis of variance revealed a significant interaction between genotypes and hydric management for most traits, with the exceptions being LPB, LPI, and NNP (
Table 1); therefore, the interaction was unfolded to study the means for these variables. In the absence of interaction, the individual effects were assessed.
The interaction between the effects of genotypes and hybrid management was unfolded for the 11 traits, for which it was statistically significant.
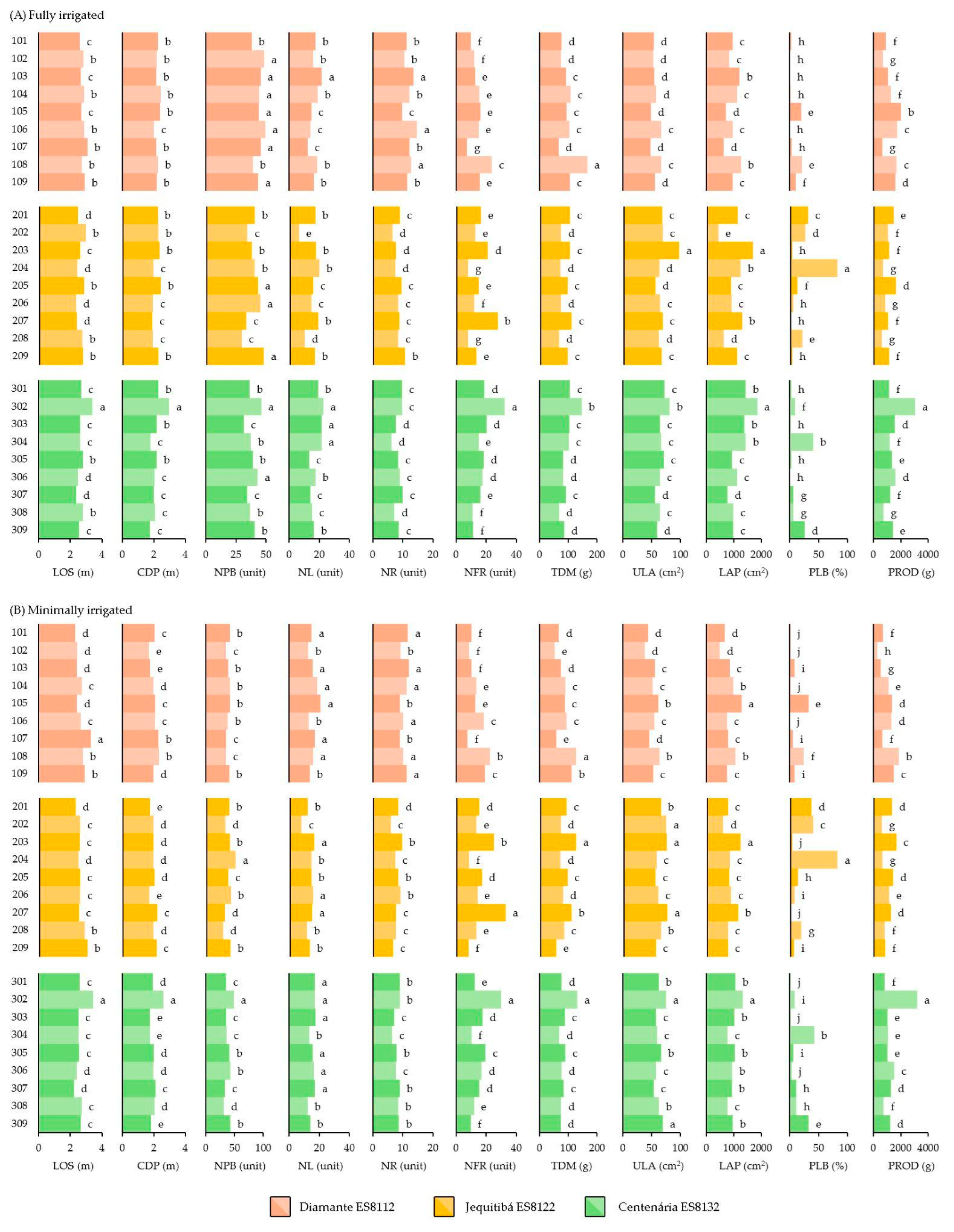
Figure 2 shows the unfolding of the genotypes for each condition of the water supply. Nine homogenous groups were observed for the proportion of large grains (PLBs) in the minimally irrigated and eight in the fully irrigated condition. The same behavior occurred for production (PROD), however, with eight and seven groups in the minimally irrigated and fully irrigated conditions, respectively. For the number of fruits per rosette (NFR), there was an opposite behavior, as seven groups were formed in the fully irrigated and six in the minimally irrigated condition. From the vegetative and biomass-related traits, the leaf area of the plagiotropic branch (LAP) favored the formation of five groups in the fully irrigated and four groups in the minimally irrigated management, while the total biomass of the plagiotropic branches (TDM) allowed for clustering of the genotypes into five groups in the minimally irrigated and four in the fully irrigated condition.
Analyzing the response to the increased irrigation, it is possible to observe relevant gains for several traits. Being cultivated with full irrigation, overall, the plants developed canopies 7 cm higher, with diameters 18 cm larger, and two more plagiotropic branches per plant. Additionally, these branches presented 0.8 more rosettes, with 0.3 more fruits and 1.65 more leaves, with a gain of 164 cm
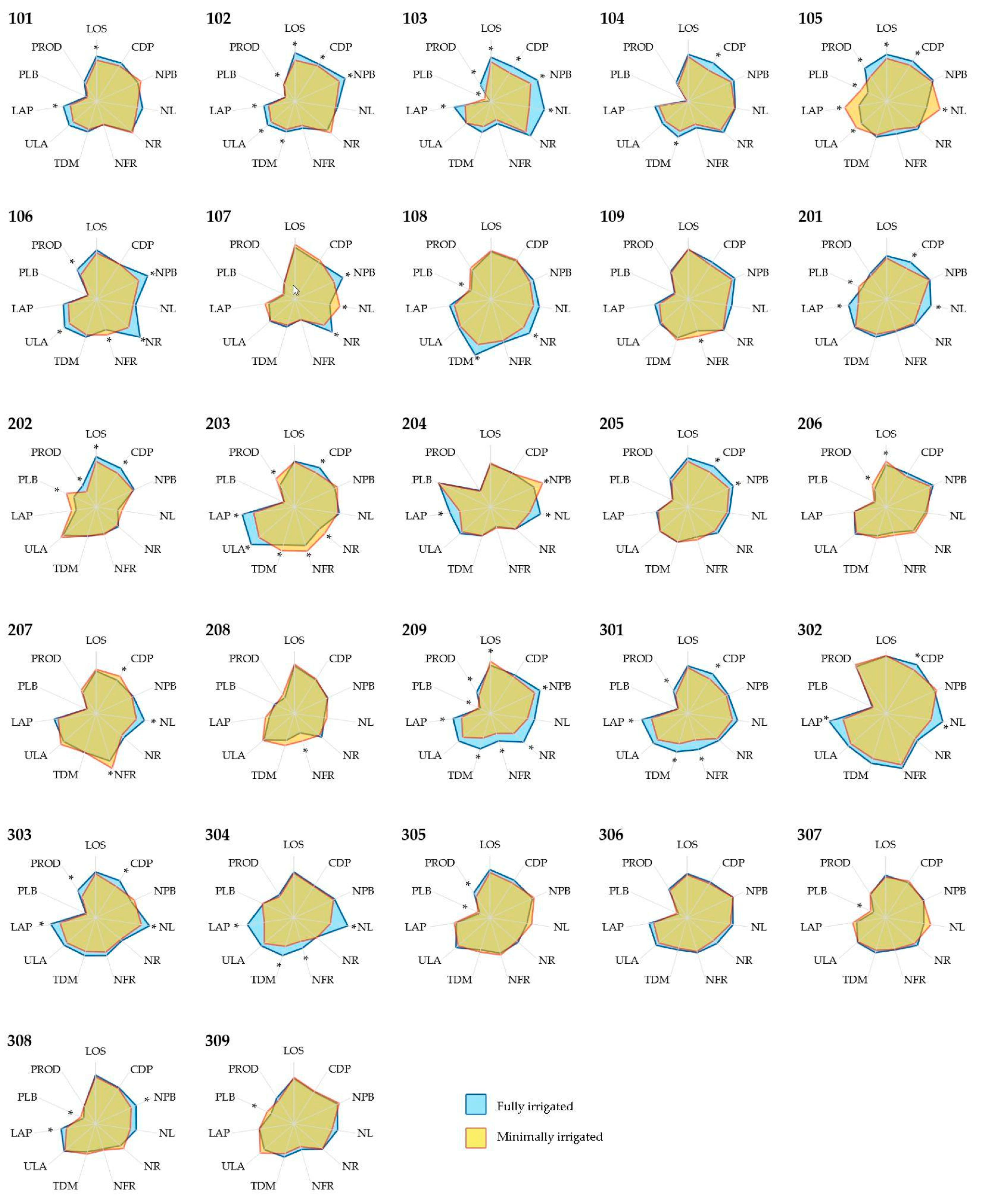
2 of area per branch. This increased growth resulted in 8.5 g more biomass per plagiotropic branch and had an effect on the production, increasing 133 g of processed coffee produced per plant. However, it is important to highlight that some overall losses were also observed, as the fully irrigated management caused a decrease in the proportion of large grains (2.7% less than the proportion in plants from the minimally irrigated condition). However, the genotypes presented different responses to the changes in the hydric management, as shown in
Figure 3.
For the length of the plagiotropic branches (LPBs), length of plagiotropic internodes (LPIs), and number of nodes per plagiotropic branch (NNP), there was no significant interaction between the effects of genotypes and hydric management. Therefore, the isolated effects were assessed, revealing that the genotypes 104, 107, 301, and 302 presented longer branches, the genotypes 103, 105, 204, 206, 209, 306, 307, and 309 presented shorter internodes, and the genotypes 103 and 104 presented more nodes per branch, regardless of the hydric regimen (
Figure 4A). Regarding the response to water management, the length of the plagiotropic branches was promoted with the use of full irrigation, regardless of the genotype (
Figure 4B).
The correlation analyses showed the existence of 33 positive and significant correlations for pairs of traits and 20 negative and significant correlations for the minimally irrigated condition. While in fully irrigated conditions, 47 positive and significant correlations and 9 negative and significant ones were observed (
Figure 5).
4. Discussion
Studies indicate that the optimal annual average temperature for
C. canephora cultivation ranges between 22 °C and 26 °C [
14]. The observed temperature falls slightly below this optimal range, which is attributed to the transitional altitude of the region.
The accumulated annual precipitation was adequate for plant development. Rainfall was particularly significant during the summer months (September to February), reaching 1058.6 mm (
Figure 1). However, despite the substantial precipitation during this period, January presented a water deficit, which coincided with the critical phenological stage of fruit development, when the transition from expansion to grain filling occurs [
23]. In this context, the plants from the fully irrigated management received the necessary supplementation of water during this important period. This approach justifies the need to analyze the precipitation data based on its distribution throughout the phenological phases and not just the annual or seasonal accumulation, since one deficit month alone (e.g., January,
Figure 1) in a critical phenological stage could cause significant productivity losses [
24].
Traditionally,
C. canephora is cultivated at altitudes of up to 500 m, where thermal conditions are more suitable for its development. However, due to the intensification of heat waves resulting from climate change, there is a growing trend of migrating this crop to higher altitudes in search of milder temperatures [
6]. Predictive studies about climate change indicate a reduction in suitable areas for the cultivation of
C. canephora, which reinforces the need to investigate the phenotypic plasticity and to select genotypes better adapted to transitional altitudes, adding alternatives to a more sustainable coffee production [
8,
25,
26,
27].
In this context, [
28] affirms that climate change can have both negative and positive effects, particularly in higher lands. Some advantages are the increases in pollination and increased yield caused by the higher carbon concentration. However, if the gains can be offset, the losses are still an important research topic. These authors also point to a major worry about major coffee-growing regions, most likely to be adversely affected by climate change, which will require shifts in the current ways to cultivate coffee. Valuable alternatives for
C. canephora will probably be the relocation of plantations, agroforestry, and the use of irrigation. Therefore, it is necessary to identify genotypes that already have advantageous behavior with the adoption of these alternatives.
There was significant variation in vegetative and reproductive aspects among the genotypes according to hydric management, which is justified by the high genetic variability of this species [
15,
26,
27]. Aspects related to fruit production were those that allowed the greatest differentiation among groups of genotypes. The performance of the genotype 302 can be highlighted regardless of the hydric management, resulting in plants with higher canopies, a higher number of productive branches, and higher overall coffee production. The genotype 105 was part of a group of relatively low production when minimally irrigated but became, alone, the second highest productive genotype when fully irrigated, which is evidence of its responsiveness. The genotype 203 kept a high investment in leafness when minimally irrigated but presented seemingly a lack of gains of production in the current cycle; this genotype was part of the third highest yielding group when minimally irrigated but composed the sixth when fully irrigated.
Based on the significance of the effect of the hydric management, it is possible to classify the genotypes according to the observed gain for each trait. The genotypes 102, 103, 105, 106, 203, 209, and 301 presented growth and production more modulated by the water management, with influences over several of the evaluated traits. Among these, the growth and production of genotypes 102, 103, 106, 209, and 301 were favored by the fully irrigated condition.
Some genotypes were less influenced by the changes in the water regimen, presenting fewer traits being significantly affected by irrigation. In this context, the genotypes 109, 205, 208, 307, and 309 were stable, regardless of the irrigation, and the genotype 306 can be highlighted as the most stable.
With emphasis on the crop yield, the genotypes 102, 103, 105, 106, 202, 209, 301, 303, and 305 presented significant gains with full irrigation, achieving productions 60% higher than observed with the minimally irrigated condition. The genotypes 203 and 206, however, were capable of achieving higher coffee production (29% higher) in the minimally irrigated condition, which can be related to a different water requirement or a higher tolerance to water stresses. The genotype 203 presented a shift towards vegetative growth in the environment with a higher water supply, mainly towards leafiness. This behavior for coffee production for the genotype 206 could be favored by the significant effect over the length of the orthotropic stem, in addition to combined lesser effects over other traits.
Changes in cultivation practices can act as management strategies to mitigate negative effects caused by climate change (e.g., relocation of plantations to more climatically suitable areas, irrigation, and agroforestry), aiding plant development and productive performance [
28]. In this study, full irrigation increased the magnitude of positive correlations between coffee production and growth and leafiness of genotypes (e.g., PROD with LOS, CDP, NPB, NL, ULA, and LAP), as well as with the number of fruits per rosette and dry biomass of plagiotropic branches (e.g., PROD with NFR and TDM). These effects indicate a balance between vegetative and reproductive growth with increased irrigation, which contributes to mitigating the effects of crop bienniality observed in the species [
29].
Under minimal irrigation, the production was correlated with canopy traits (e.g., PROD with CDP and LPB), leaf area (e.g., PROD with ULA and LAP), fruits per rosette (PROD with NFR), and biomass of plagiotropic branches (PROD with TDM). In this condition, there is a weaker association between canopy growth traits and coffee production, which indicates a slightly more intense competition between reproductive and vegetative growth in the current phenological cycle, which could result in losses to the production capacity of the following cycles.
Balancing vegetative and reproductive growth must be taken into consideration in coffee farming, especially for the ratio between leaves and fruits. The imbalance of these relationships may be the main cause of plant depletion and accentuate the bienniality effect [
30,
31]. Cultural management techniques (e.g., pruning) can be used to modulate the source/sink relationships and to renew the photosynthetic apparatus of coffee plants [
32], and can be strategically used in order to enhance the performance of selected genotypes in higher altitudes and in association with irrigation.
Selecting genotypes that can express larger gains under the mitigative effects of the association between the cultivation at transitional altitudes and the use of irrigation is a strategy that is possible and required in order to better explore the synergy between these technologies, aiding to better use the genetic and water resources and to make the coffee cultivation more sustainable. Further research on this topic is important in order to better understand the tolerance to water stress and to quantify the gains that can be obtained by the association of these agricultural practices, as well as assess the long-term effects on the coffee yield, especially seeking adaptability and stability of genotypes of coffee for future plantations.



 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}