


Foliar Application of a Mixture of Amino Acid-Based Growth Promoters Enhances Tomato Seedling Production

 ,
, 
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Growth Conditions and Tomato Seeds
2.2. Experimental Design
2.3. Amino Acid Treatments and Composition
2.4. Determination of Growth Parameters
2.5. Data Analysis
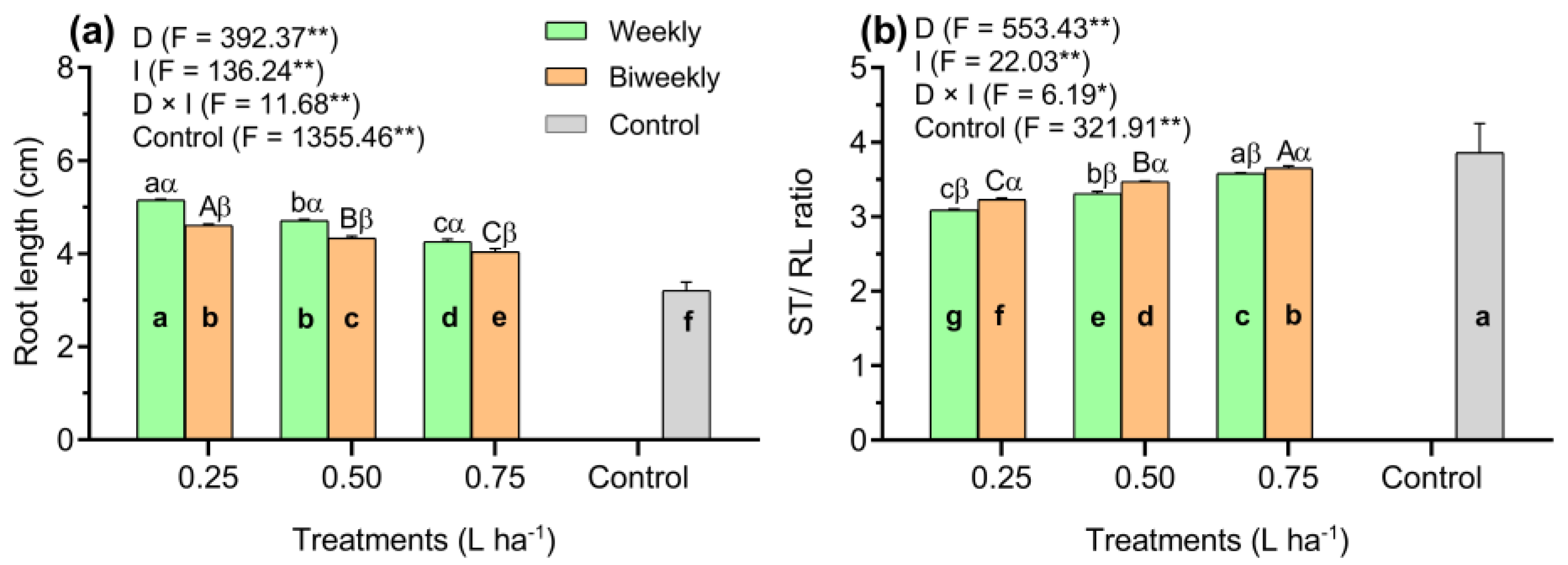
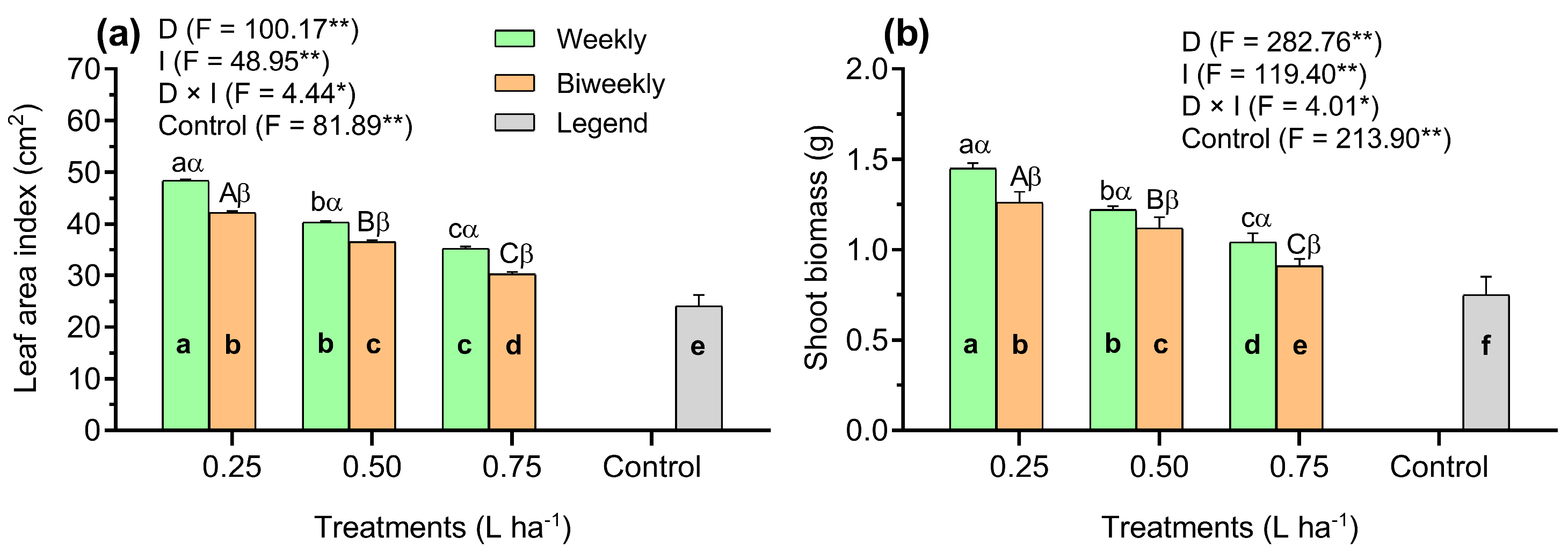
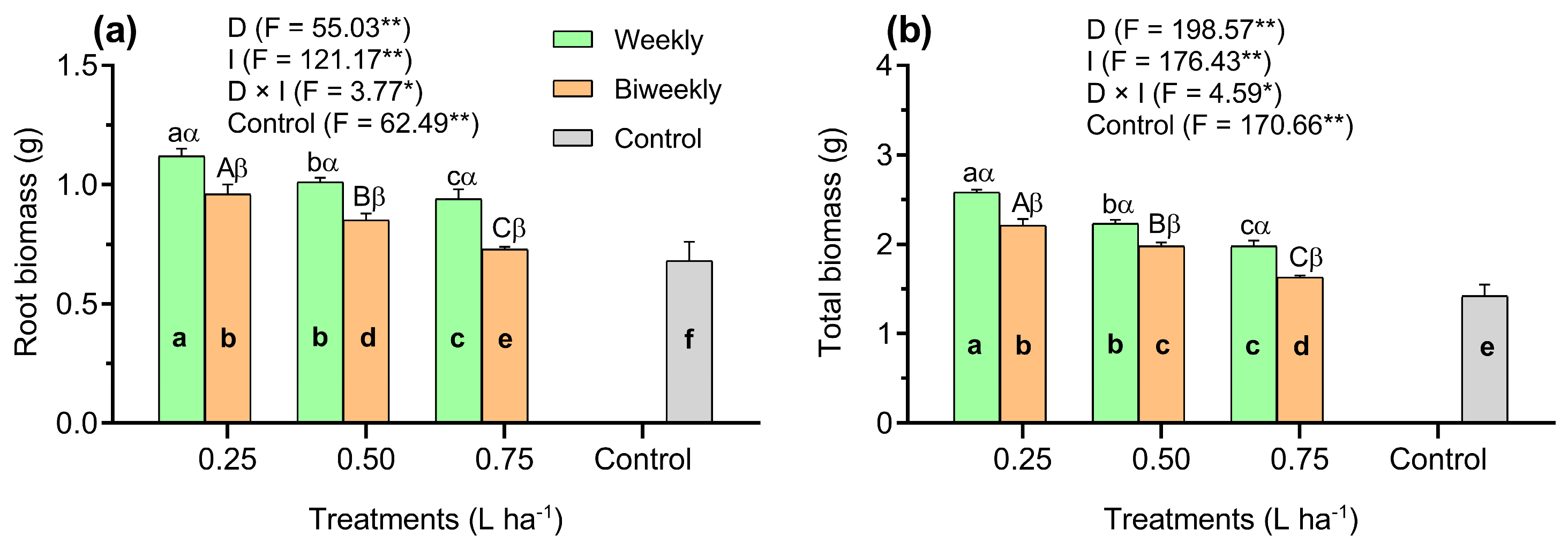
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SI | Stem length |
| ST | Stem thickness |
| RL | Root length |
| RB | Root biomass |
| SB | Shoot biomass |
| TB | Total biomass |
| LAI | Leaf area index |
| DAS | Days after seeding |
| D1W1 | Applying 0.25 L ha−1 of AAM weekly |
| D2W1 | Applying 0.50 L ha−1 of AAM weekly |
| D3W1 | Applying 0.75 L ha−1 of AAM weekly |
| D1W2 | Applying 0.25 L ha−1 of AAM biweekly |
| D2W2 | Applying 0.50 L ha−1 of AAM biweekly |
| D3W2 | Applying 0.75 L ha−1 of AAM biweekly |
| D | Doses of a mixture of amino acids |
| I | Application interval of a mixture of amino acids |
| D × I | D–I interaction |
| F | Fischer values |
References
- Abbass, K.; Qasim, M.Z.; Song, H.; Murshed, M.; Mahmood, H.; Younis, I. A Review of the Global Climate Change Impacts, Adaptation, and Sustainable Mitigation Measures. Environ. Sci. Pollut. Res. 2022, 29, 42539–42559. [Google Scholar] [CrossRef] [PubMed]
- Malhi, G.S.; Kaur, M.; Kaushik, P. Impact of Climate Change on Agriculture and Its Mitigation Strategies: A Review. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Janni, M.; Maestri, E.; Gullì, M.; Marmiroli, M.; Marmiroli, N. Plant Responses to Climate Change, How Global Warming May Impact on Food Security: A Critical Review. Front. Plant Sci. 2024, 14, 1297569. [Google Scholar] [CrossRef] [PubMed]
- Nimma, D.; Devi, O.R.; Laishram, B.; Ramesh, J.V.N.; Boddupalli, S.; Ayyasamy, R.; Tirth, V.; Arabil, A. Implications of Climate Change on Freshwater Ecosystems and Their Biodiversity. Desalination Water Treat. 2025, 321, 100889. [Google Scholar] [CrossRef]
- Rodríguez, D.J.; Ramírez-Pérez, C.; Ramírez-Rodríguez, H.; Villarreal-Quintanilla, J.Á.; Hernández-Pérez, A.; Díaz-Jimenez, M.L.V.; Peña-Ramos, F.M. Bioestimulantes de plantas del semidesierto en el crecimiento radicular y aéreo de plántulas de tomate. Ecosistemas Y Recur. Agropecu. 2024, 11, e4145. [Google Scholar] [CrossRef]
- Domínguez, I.; del Río, J.L.; Ortiz-Somovilla, V.; Cantos-Villar, E. Technological Innovations for Reducing Tomato Loss in the Agri-Food Industry. Food Res. Int. 2025, 203, 115798. [Google Scholar] [CrossRef]
- Goh, Y.S.; Hum, Y.C.; Lee, Y.L.; Lai, K.W.; Yap, W.-S.; Tee, Y.K. A Meta-Analysis: Food Production and Vegetable Crop Yields of Hydroponics. Sci. Hortic. 2023, 321, 112339. [Google Scholar] [CrossRef]
- Lizazo, I.C.; Hurtado, A.C.; Hernández, M.G.R.; Casas, A.P.; Balmori, D.M.; Díaz, Y.P. Potentialities of two biostimulants on germination and growth of tomato seedlings. Cienc. Tecnol. Agropecu. 2022, 23, e2343. [Google Scholar] [CrossRef]
- Calero, A.; Quintero, E.; Pérez, Y.; Olivera, D.; Peña, K.; Castro, I.; Jiménez, J. Evaluation of efficient microorganisms in the tomato seedling production (Solanum lycopersicum L.). Rev. De Cienc. Agrícolas 2019, 36, 67–78. [Google Scholar] [CrossRef]
- Gedeon, S.; Ioannou, A.; Balestrini, R.; Fotopoulos, V.; Antoniou, C. Application of Biostimulants in Tomato Plants (Solanum lycopersicum) to Enhance Plant Growth and Salt Stress Tolerance. Plants 2022, 11, 3082. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, J.; Jeong, S.J.; Masabni, J.; Niu, G. Biostimulants Applied in Seedling Stage Can Improve Onion Early Bulb Growth: Cultivar- and Fertilizer-Type-Specific Positive Effects. Horticulturae 2025, 11, 402. [Google Scholar] [CrossRef]
- Bell, J.C.; Bound, S.A.; Buntain, M. Biostimulants in Agricultural and Horticultural Production. In Horticultural Reviews; John Wiley and Sons, Ltd.: Hoboken, NJ, USA, 2022; pp. 35–95. ISBN 978-1-119-85198-1. [Google Scholar]
- du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Kisvarga, S.; Farkas, D.; Boronkay, G.; Neményi, A.; Orlóci, L. Effects of Biostimulants in Horticulture, with Emphasis on Ornamental Plant Production. Agronomy 2022, 12, 1043. [Google Scholar] [CrossRef]
- Peña Calzada, K.; Olivera Viciedo, D.; Habermann, E.; Calero Hurtado, A.; Lupino Gratão, P.; De Mello Prado, R.; Lata-Tenesaca, L.F.; Martinez, C.A.; Ajila Celi, G.E.; Rodríguez, J.C. Exogenous Application of Amino Acids Mitigates the Deleterious Effects of Salt Stress on Soybean Plants. Agronomy 2022, 12, 2014. [Google Scholar] [CrossRef]
- Peña-Calzada, K.; Calero-Hurtado, A.; Meléndrez-Rodríguez, J.F.; Rodríguez-Fernández, J.C.; Gutiérrez-Cádenas, O.G.; García-González, M.T.; Madrigal-Carmona, L.; Jiménez-Medina, A. Impacts of the Biostimulant VIUSID® Agro on Growth, Productivity, and Tolerance to Salt Stress in Crops: A Systematic Review. Horticulturae 2025, 11, 407. [Google Scholar] [CrossRef]
- Calero Hurtado, A.; Peña Calzada, K.; Fasoli, J.V.B.; Jiménez, J.; Sánchez López, L. Synergic Effects of the Microbial Consortium and Amino Acid-Based Growth Promoter in Sunflower Productivity Under Water-Deficit Conditions. Water 2025, 17, 1365. [Google Scholar] [CrossRef]
- Díaz, Y.P.; Hurtado, A.C.; Calzada, K.P.; Díaz, J.L.G.; González, V.R. Plant densities and foliar application of amino acids increasing sesame yield. Temas Agrar. 2024, 29, 100–112. [Google Scholar] [CrossRef]
- Almutairi, K.F.; Saleh, A.A.; Ali, M.M.; Sas-Paszt, L.; Abada, H.S.; Mosa, W.F.A. Growth Performance of Guava Trees after the Exogenous Application of Amino Acids Glutamic Acid, Arginine, and Glycine. Horticulturae 2022, 8, 1110. [Google Scholar] [CrossRef]
- Ban, Y.J.; Song, Y.H.; Kim, J.Y.; Cha, J.Y.; Ali, I.; Baiseitova, A.; Shah, A.B.; Kim, W.-Y.; Park, K.H. A Significant Change in Free Amino Acids of Soybean (Glycine max L. Merr) through Ethylene Application. Molecules 2021, 26, 1128. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Yi, J.Q.; Piao, X.S.; Li, P.F.; Zeng, Z.K.; Wang, D.; Liu, L.; Wang, G.Q.; Han, X. The Metabolizable Energy Value, Standardized Ileal Digestibility of Amino Acids in Soybean Meal, Soy Protein Concentrate and Fermented Soybean Meal, and the Application of These Products in Early-Weaned Piglets. Asian-Australas J. Anim. Sci. 2013, 26, 691–699. [Google Scholar] [CrossRef]
- Radkowski, A.; Radkowska, I.; Bocianowski, J.; Sladkovska, T.; Wolski, K. The Effect of Foliar Application of an Amino Acid-Based Biostimulant on Lawn Functional Value. Agronomy 2020, 10, 1656. [Google Scholar] [CrossRef]
- Navarro-León, E.; Borda, E.; Marín, C.; Sierras, N.; Blasco, B.; Ruiz, J.M. Application of an Enzymatic Hydrolysed L-α-Amino Acid Based Biostimulant to Improve Sunflower Tolerance to Imazamox. Plants 2022, 11, 2761. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, A.; Borrego, E.J.; Savka, M.A.; Dobson, R.C.J.; Hudson, A.O. Amino Acid–Derived Defense Metabolites from Plants: A Potential Source to Facilitate Novel Antimicrobial Development. J. Biol. Chem. 2021, 296, 100438. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.S.; Ryu, K.W.; Thompson, C.B. Arginine: At the Crossroads of Nitrogen Metabolism. EMBO J. 2025, 44, 1275–1293. [Google Scholar] [CrossRef]
- Lei, S.; Rossi, S.; Huang, B. Metabolic and Physiological Regulation of Aspartic Acid-Mediated Enhancement of Heat Stress Tolerance in Perennial Ryegrass. Plants 2022, 11, 199. [Google Scholar] [CrossRef]
- Oogai, Y.; Yamaguchi, M.; Kawada-Matsuo, M.; Sumitomo, T.; Kawabata, S.; Komatsuzawa, H. Lysine and Threonine Biosynthesis from Aspartate Contributes to Staphylococcus Aureus Growth in Calf Serum. Appl. Environ. Microbiol. 2016, 82, 6150–6157. [Google Scholar] [CrossRef]
- Han, R.; Khalid, M.; Juan, J.; Huang, D. Exogenous Glycine Inhibits Root Elongation and Reduces Nitrate-N Uptake in Pak Choi (Brassica campestris ssp. Chinensis L.). PLoS ONE 2018, 13, e0204488. [Google Scholar] [CrossRef]
- Kawade, K.; Tabeta, H.; Ferjani, A.; Hirai, M.Y. The Roles of Functional Amino Acids in Plant Growth and Development. Plant Cell Physiol. 2023, 64, 1482–1493. [Google Scholar] [CrossRef]
- Hurtado, A.C.; Chiconato, D.A.; Junior, G.d.S.S.; Prado, R.d.M.; Peña Calzada, K.; Olivera Viciedo, D. Silicon Induces Salt Stress Amelioration in Sunflower Plants by Improving Photosynthetic Pigments and Mineral Status. Stresses 2024, 4, 860–869. [Google Scholar] [CrossRef]
- Calzada, K.P.; Hurtado, A.C.; Peistrup, V.; Mühlmann, I.; Miranda, D.R.; Coca, L.I.R.; González, M.R.; Fernández, J.C.R. Physiological and productive responses of sugar beet plants treated with amino acid solution. Temas Agrar. 2024, 29, 113–125. [Google Scholar] [CrossRef]
- Kocira, S.; Szparaga, A.; Hara, P.; Treder, K.; Findura, P.; Bartoš, P.; Filip, M. Biochemical and Economical Effect of Application Biostimulants Containing Seaweed Extracts and Amino Acids as an Element of Agroecological Management of Bean Cultivation. Sci. Rep. 2020, 10, 17759. [Google Scholar] [CrossRef] [PubMed]
- Jacomassi, L.M.; Pacola, M.; Momesso, L.; Viveiros, J.; Júnior, O.A.; de Siqueira, G.F.; de Campos, M.; Crusciol, C.A.C. Foliar Application of Amino Acids and Nutrients as a Tool to Mitigate Water Stress and Stabilize Sugarcane Yield and Bioenergy Generation. Plants 2024, 13, 461. [Google Scholar] [CrossRef] [PubMed]
- Calzada, K.P.; Fernández, J.C.R.; Meléndrez, J.F. EL VIUSID agro una alternativa en el incremento de la producción. Rev. Caribeña De Cienc. Soc. 2016, 15, 1–6. [Google Scholar]
- Kemp, C.D. Methods of Estimating the Leaf Area of Grasses from Linear Measurements. Ann. Bot. 1960, 24, 491–499. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024. [Google Scholar]
- Zhang, C.; Zhang, J.; Liu, W.; Ji, J.; Zhang, K.; Li, H.; Feng, Y.; Xue, J.; Ji, C.; Zhang, L.; et al. Mechanisms of Branched Chain Amino Acids Promoting Growth and Lipid Accumulation in Camelina sativa Seedlings under Drought and Salt Stress. Sustain. Energy Technol. Assess. 2025, 75, 104201. [Google Scholar] [CrossRef]
- Peña, K.; Calero Hurtado, A.; Viciedo, D.; Rodríguez, J.; Fernandes, T.; García, R.; Ajila, G. Agroproductive Response of Zea Mayz L. with the Foliar Application of VIUSID Agro®. Rev. De La Fac. De Agron. Univ. Del Zulia 2021, 38, 573–584. [Google Scholar] [CrossRef]
- Calzada, K.P.; Rodríguez, J.C.; Viciedo, D.O.; Hurtado, A.C.; Meléndrez, J.F.; Valdez, R.G. VIUSID Agro® dose effect on the morpho-physiological and productive behavior of radish (Raphanus sativus L.). Rev. De La Fac. De Agron. Univ. Del Zulia 2018, 35, 293–317. [Google Scholar]
- González, Y.A.; Díaz, Y.P.; Hurtado, A.C.; Calzada, K.P. Plant densities and fertilizers improving sustainable peanut production. Rev. De La Fac. De Cienc. 2025, 14, 23–38. [Google Scholar] [CrossRef]
- Khan, S.; Yu, H.; Li, Q.; Gao, Y.; Sallam, B.N.; Wang, H.; Liu, P.; Jiang, W. Exogenous Application of Amino Acids Improves the Growth and Yield of Lettuce by Enhancing Photosynthetic Assimilation and Nutrient Availability. Agronomy 2019, 9, 266. [Google Scholar] [CrossRef]
- Lea, P.J.; Miflin, B.J. Glutamate Synthase and the Synthesis of Glutamate in Plants. Plant Physiol. Biochem. 2003, 41, 555–564. [Google Scholar] [CrossRef]
- Alcázar, R.; Marco, F.; Cuevas, J.C.; Patron, M.; Ferrando, A.; Carrasco, P.; Tiburcio, A.F.; Altabella, T. Involvement of Polyamines in Plant Response to Abiotic Stress. Biotechnol. Lett. 2006, 28, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Zhang, C.; Suglo, P.; Sun, S.; Wang, M.; Su, T. L-Aspartate: An Essential Metabolite for Plant Growth and Stress Acclimation. Molecules 2021, 26, 1887. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhuang, S.; Zhang, W. Advances in Plant Auxin Biology: Synthesis, Metabolism, Signaling, Interaction with Other Hormones, and Roles under Abiotic Stress. Plants 2024, 13, 2523. [Google Scholar] [CrossRef] [PubMed]
- Sadak, M.S.; Sekara, A.; Al-ashkar, I.; Habib-ur-Rahman, M.; Skalicky, M.; Brestic, M.; Kumar, A.; Sabagh, A.E.; Abdelhamid, M.T. Exogenous Aspartic Acid Alleviates Salt Stress-Induced Decline in Growth by Enhancing Antioxidants and Compatible Solutes While Reducing Reactive Oxygen Species in Wheat. Front. Plant Sci. 2022, 13, 987641. [Google Scholar] [CrossRef]
- Malekzadeh, P.; Hatamnia, A.A.; Tiznado-Hernández, M.E. Arginine Catabolism Induced by Exogenous Arginine Treatment Reduces the Loss of Green Color Rate in Broccoli Florets. Physiol. Mol. Plant Pathol. 2023, 124, 101973. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Cheng, Q.; Sun, W. The Effects of Amino Acids, Phenols and Protein Hydrolysates as Biostimulants on Sustainable Crop Production and Alleviated Stress. Recent Pat. Biotechnol. 2022, 16, 319–328. [Google Scholar] [CrossRef]
- Kheir, A.M.S.; Ding, Z.; Gawish, M.S.; Abou El Ghit, H.M.; Hashim, T.A.; Ali, E.F.; Eissa, M.A.; Zhou, Z.; Al-Harbi, M.S.; El-Gioushy, S.F. The Exogenous Application of Micro-Nutrient Elements and Amino Acids Improved the Yield, Nutritional Status and Quality of Mango in Arid Regions. Plants 2021, 10, 2057. [Google Scholar] [CrossRef]
- Fusco, G.M.; Burato, A.; Pentangelo, A.; Cardarelli, M.; Nicastro, R.; Carillo, P.; Parisi, M. Can Microbial Consortium Applications Affect Yield and Quality of Conventionally Managed Processing Tomato? Plants 2023, 12, 14. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calero Hurtado, A.; Meléndrez Rodríguez, J.F.; Peña Calzada, K.; Pérez Díaz, Y.; Jiménez Medina, A. Foliar Application of a Mixture of Amino Acid-Based Growth Promoters Enhances Tomato Seedling Production. Horticulturae 2025, 11, 582. https://doi.org/10.3390/horticulturae11060582
Calero Hurtado A, Meléndrez Rodríguez JF, Peña Calzada K, Pérez Díaz Y, Jiménez Medina A. Foliar Application of a Mixture of Amino Acid-Based Growth Promoters Enhances Tomato Seedling Production. Horticulturae. 2025; 11(6):582. https://doi.org/10.3390/horticulturae11060582
Chicago/Turabian StyleCalero Hurtado, Alexander, Jorge Félix Meléndrez Rodríguez, Kolima Peña Calzada, Yanery Pérez Díaz, and Alay Jiménez Medina. 2025. "Foliar Application of a Mixture of Amino Acid-Based Growth Promoters Enhances Tomato Seedling Production" Horticulturae 11, no. 6: 582. https://doi.org/10.3390/horticulturae11060582
APA StyleCalero Hurtado, A., Meléndrez Rodríguez, J. F., Peña Calzada, K., Pérez Díaz, Y., & Jiménez Medina, A. (2025). Foliar Application of a Mixture of Amino Acid-Based Growth Promoters Enhances Tomato Seedling Production. Horticulturae, 11(6), 582. https://doi.org/10.3390/horticulturae11060582