
Current and Future Geographical Distribution and Potential Habitat of Carpinus fangiana Hu (Betulaceae) Under Different Climate Change Scenarios


Abstract
1. Introduction
2. Materials and Methods
2.1. Acquisition and Processing of Distribution Data
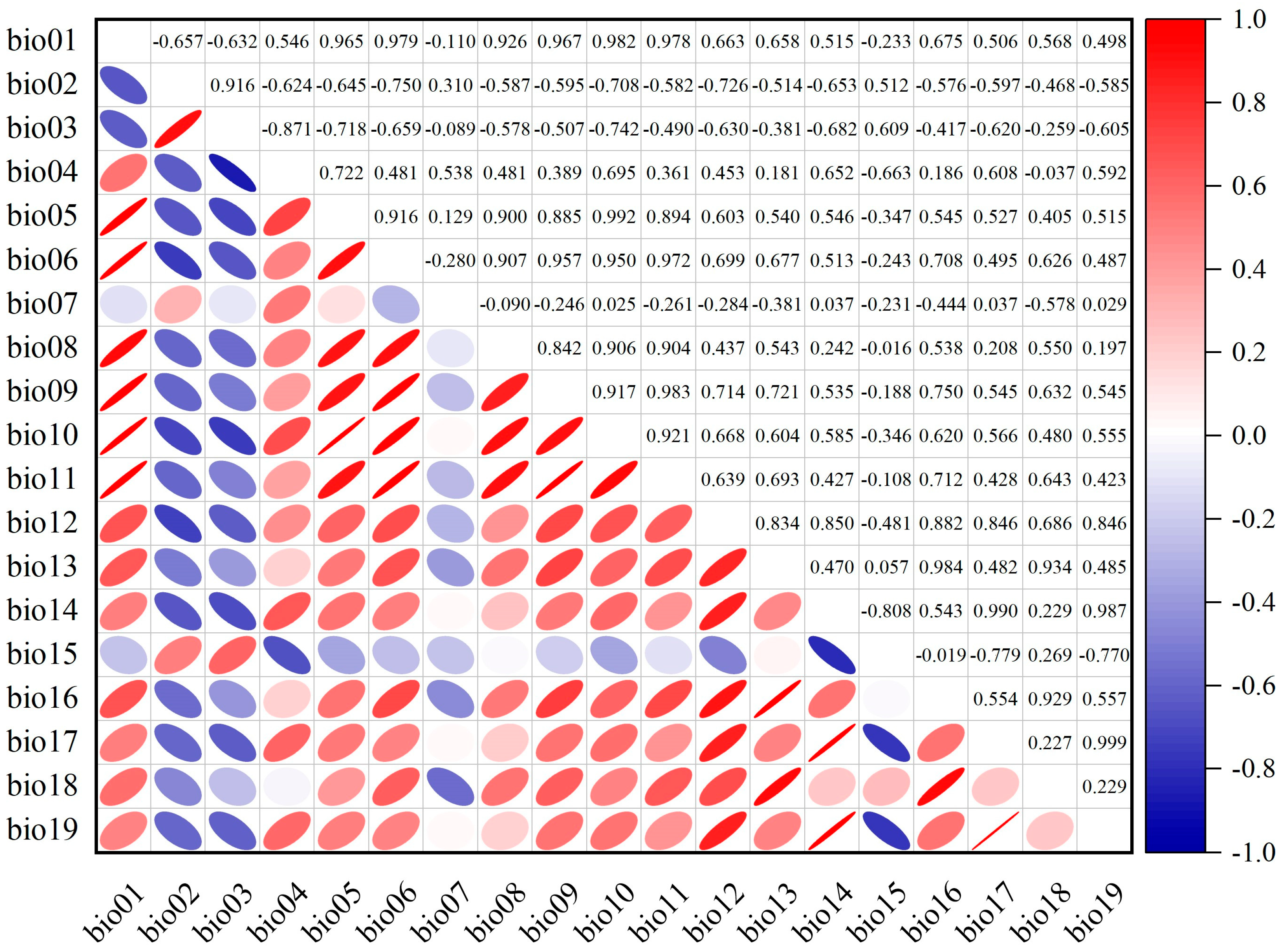
2.2. Acquisition and Processing of Environmental Variables
2.3. Construction and Evaluation of the MaxEnt Model
2.4. Classification of and Dynamic Changes in Suitable Distribution Areas
3. Results
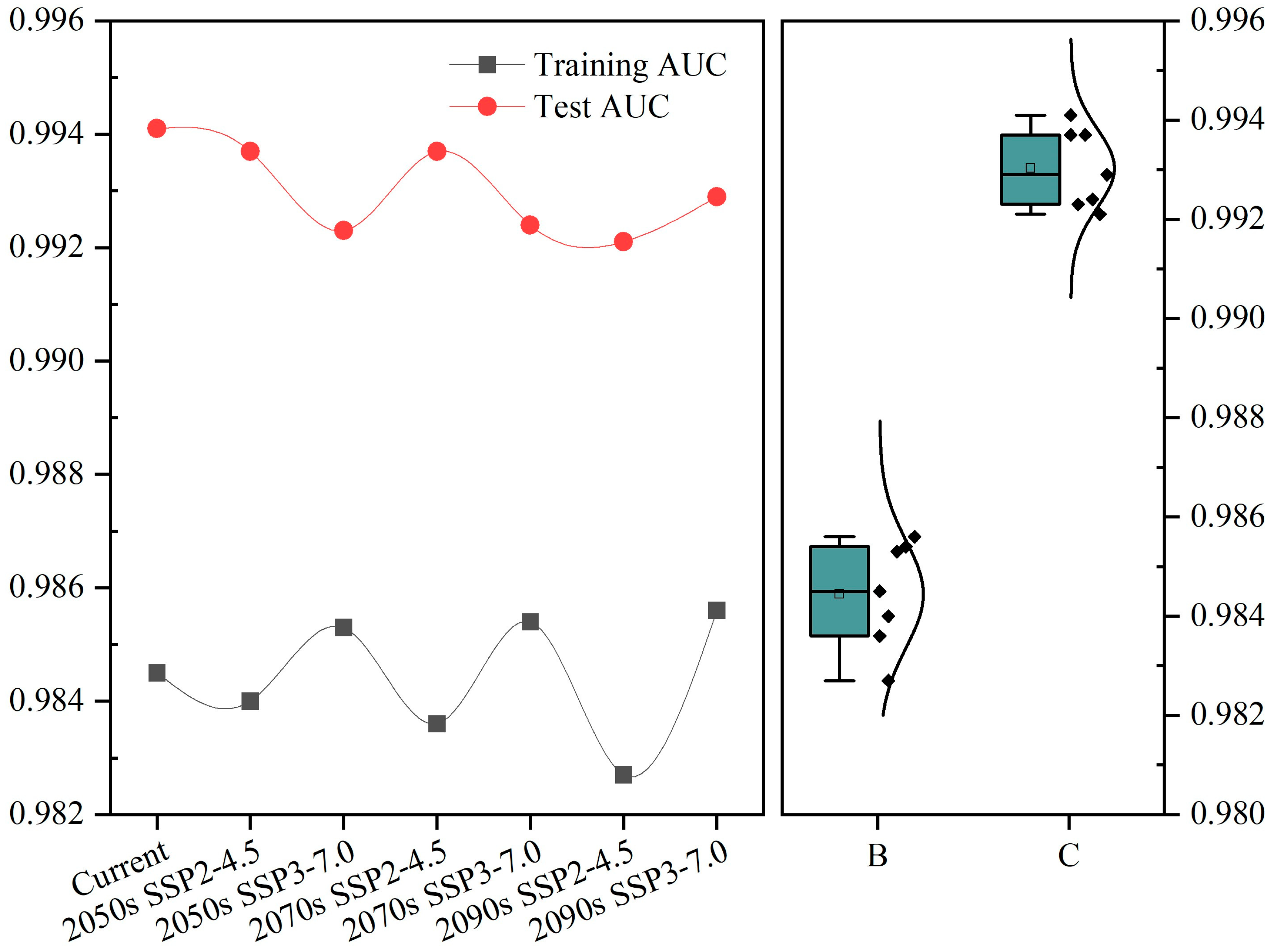
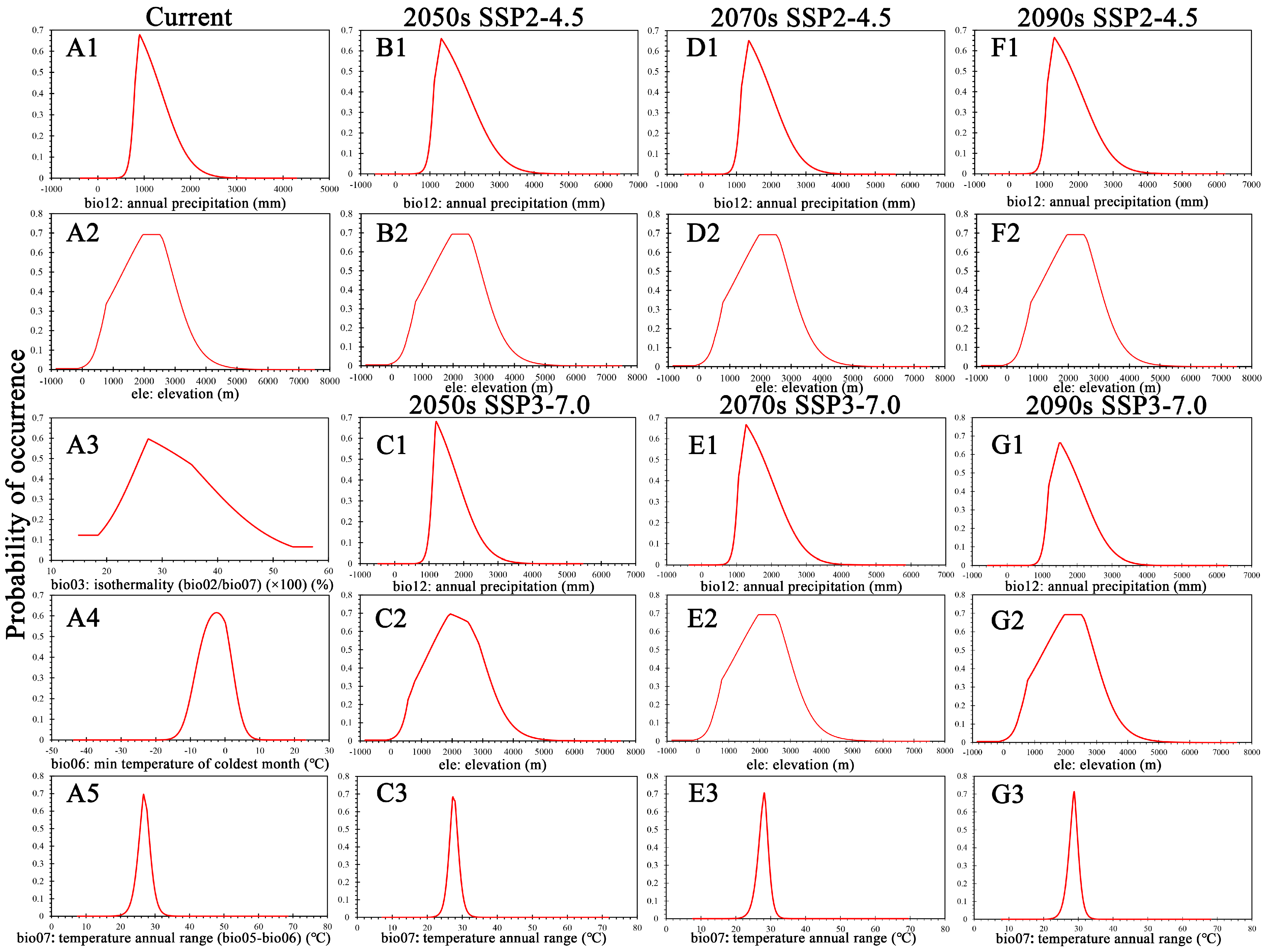
3.1. Model Accuracy and Key Environmental Factors Affecting Distribution
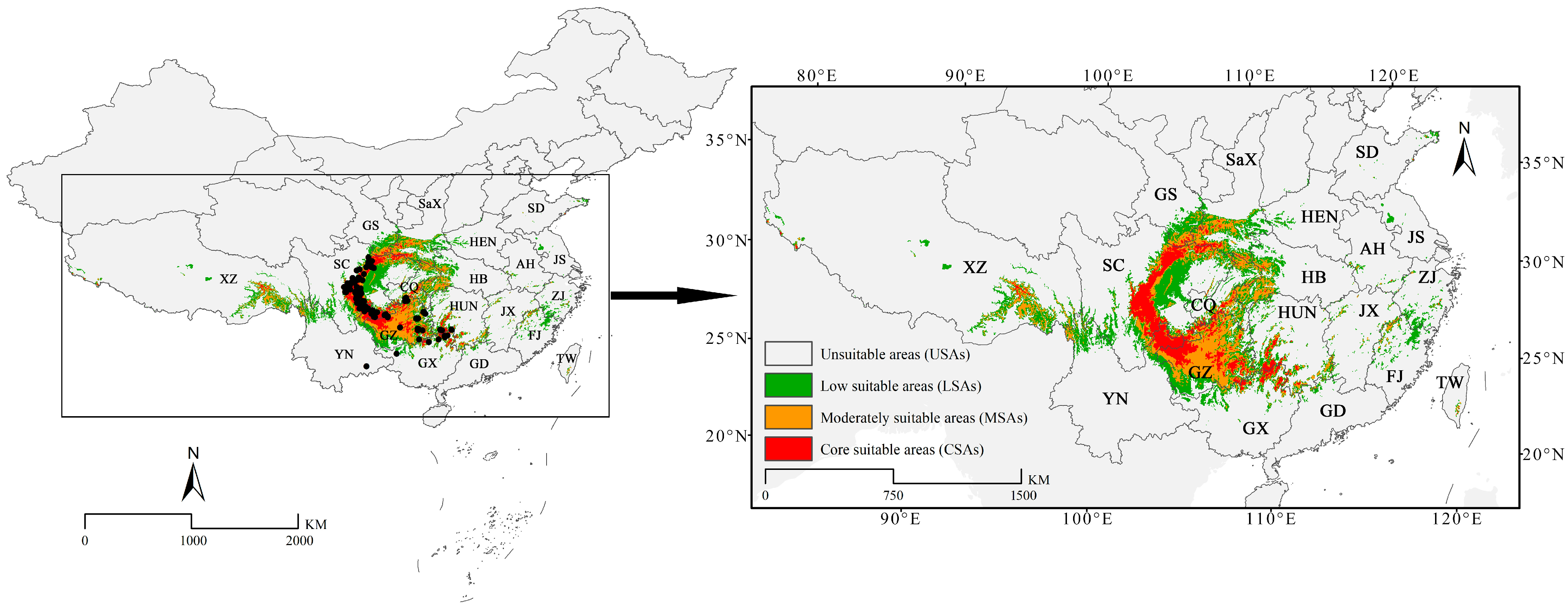
3.2. Current Potential Geographical Distribution
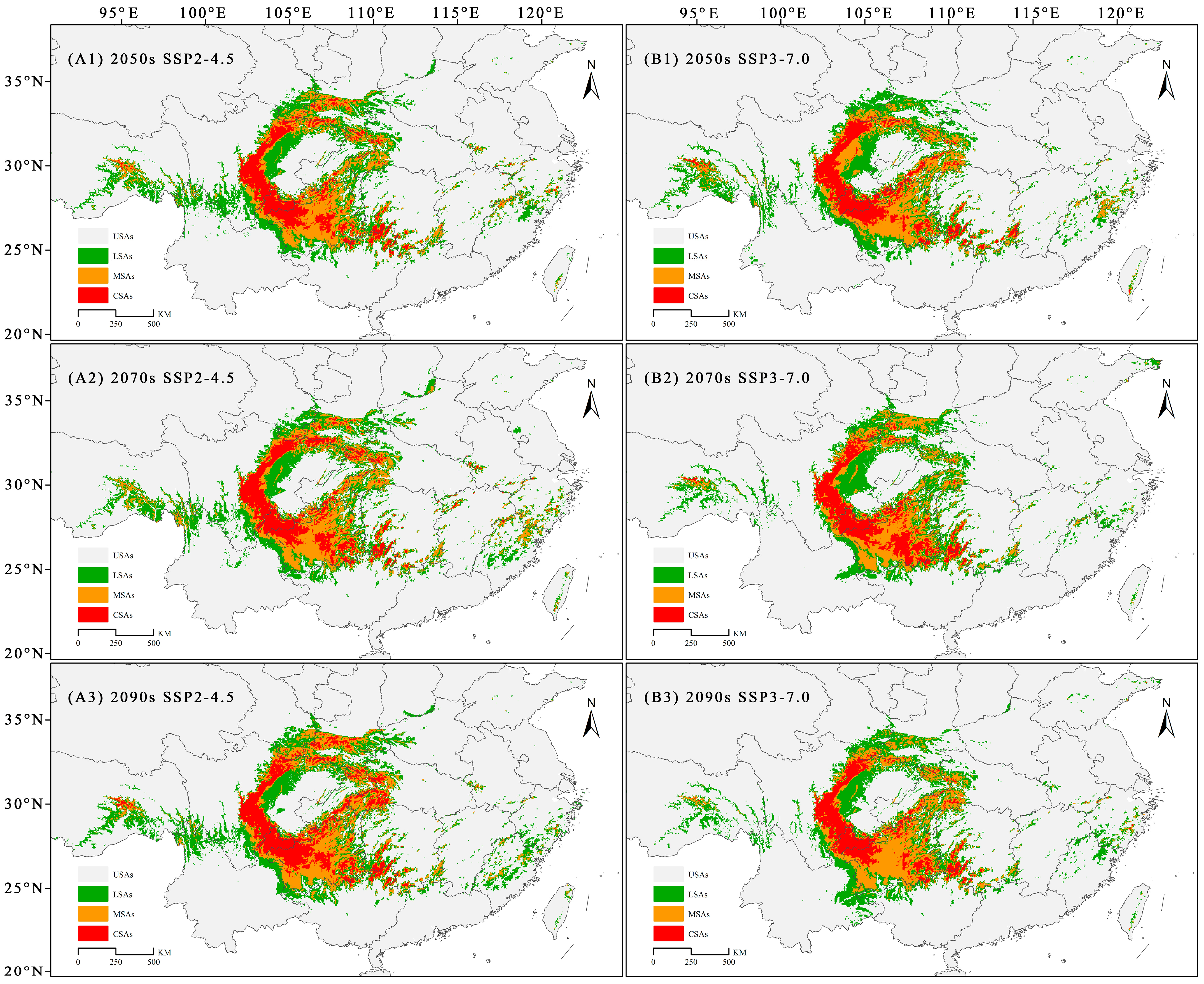
3.3. Future Potential Distribution of C. fangiana
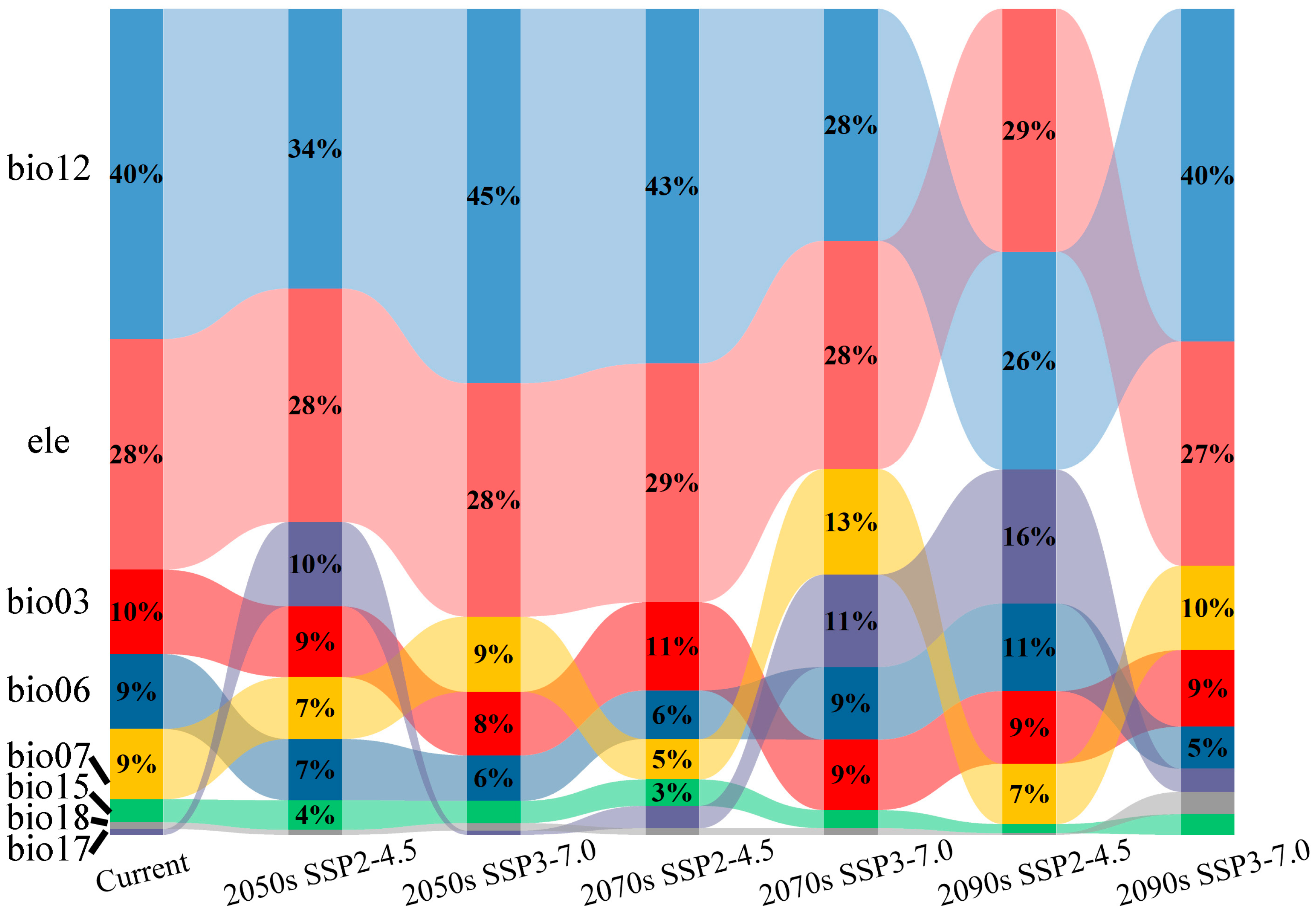
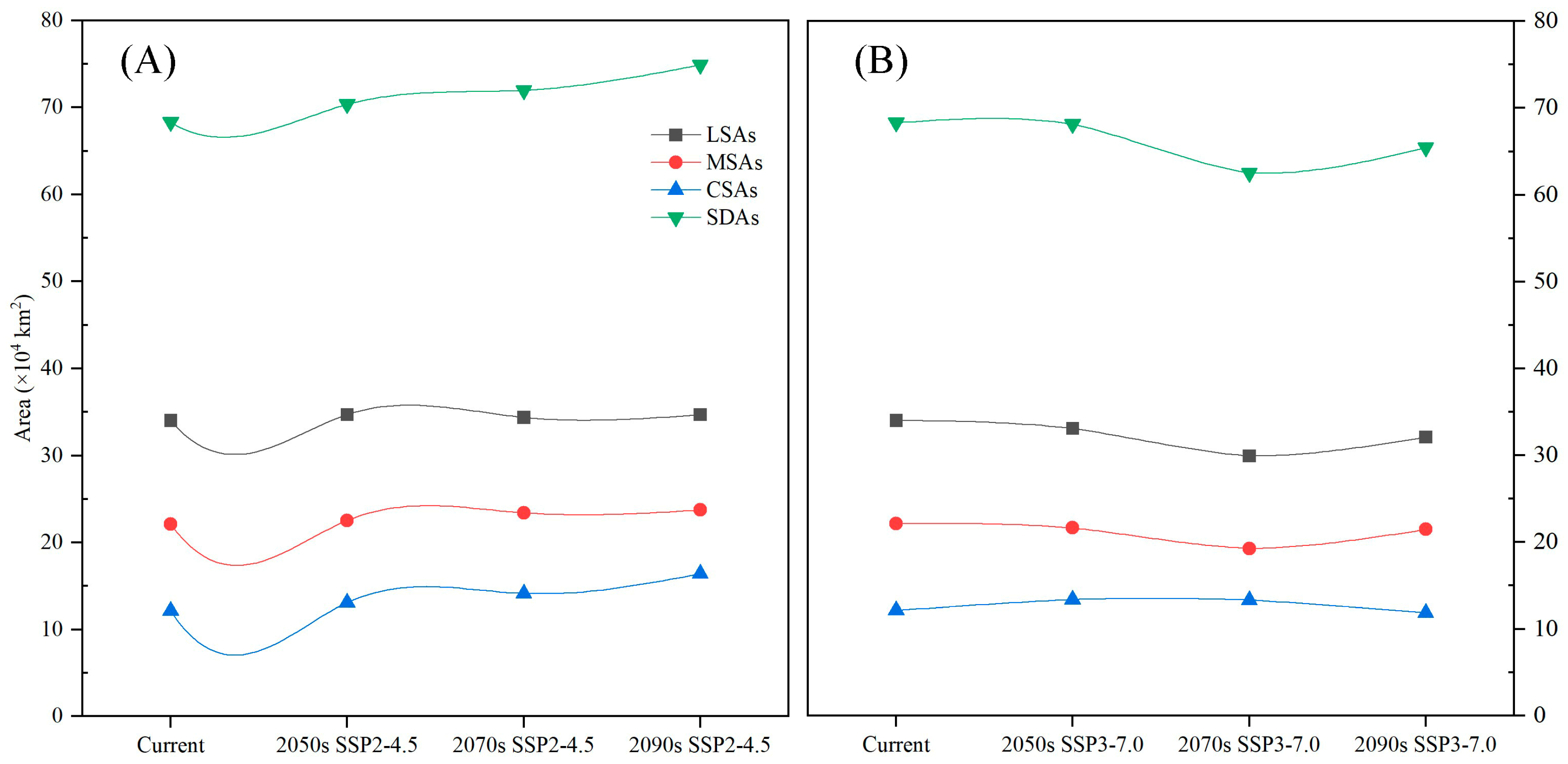
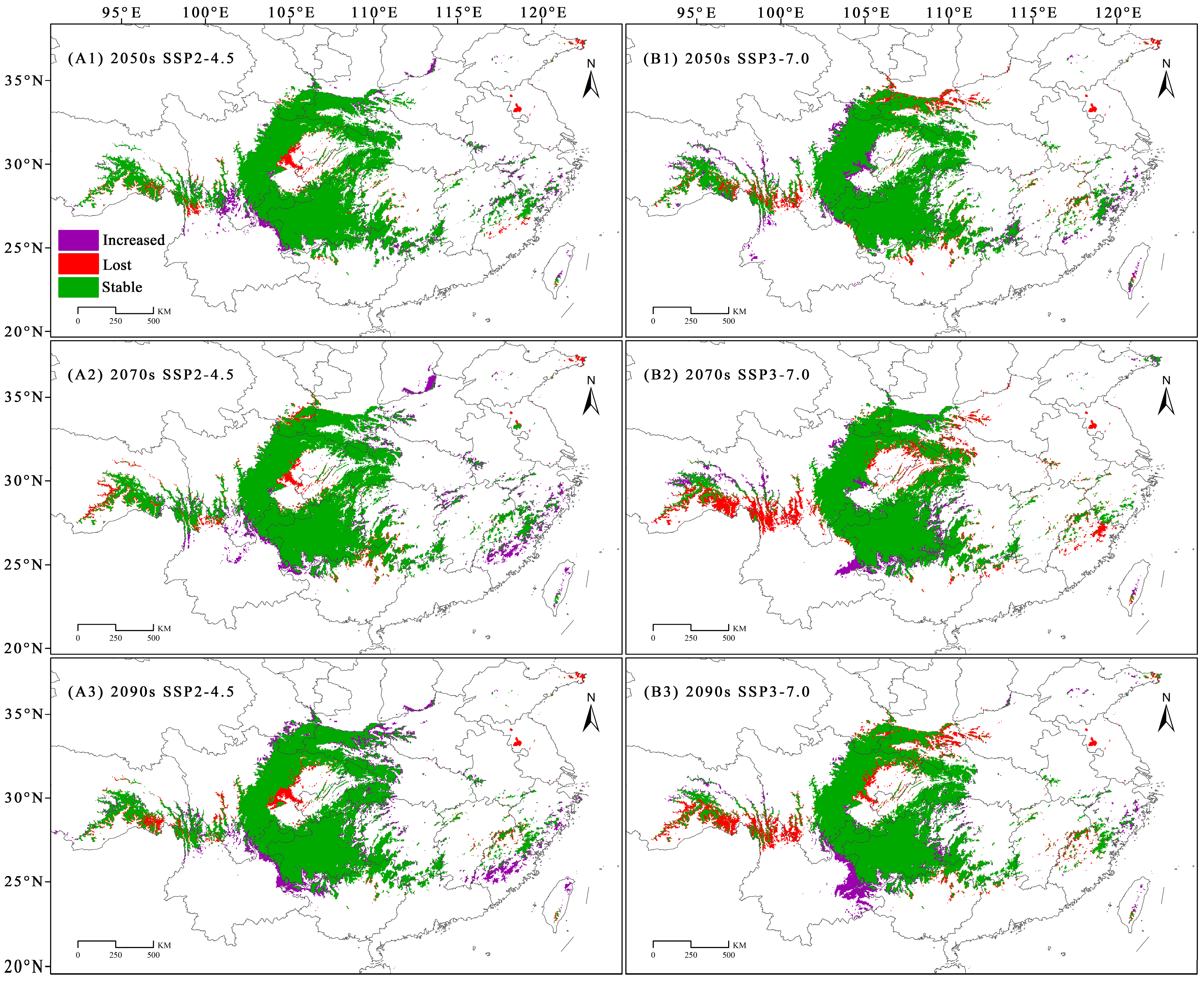
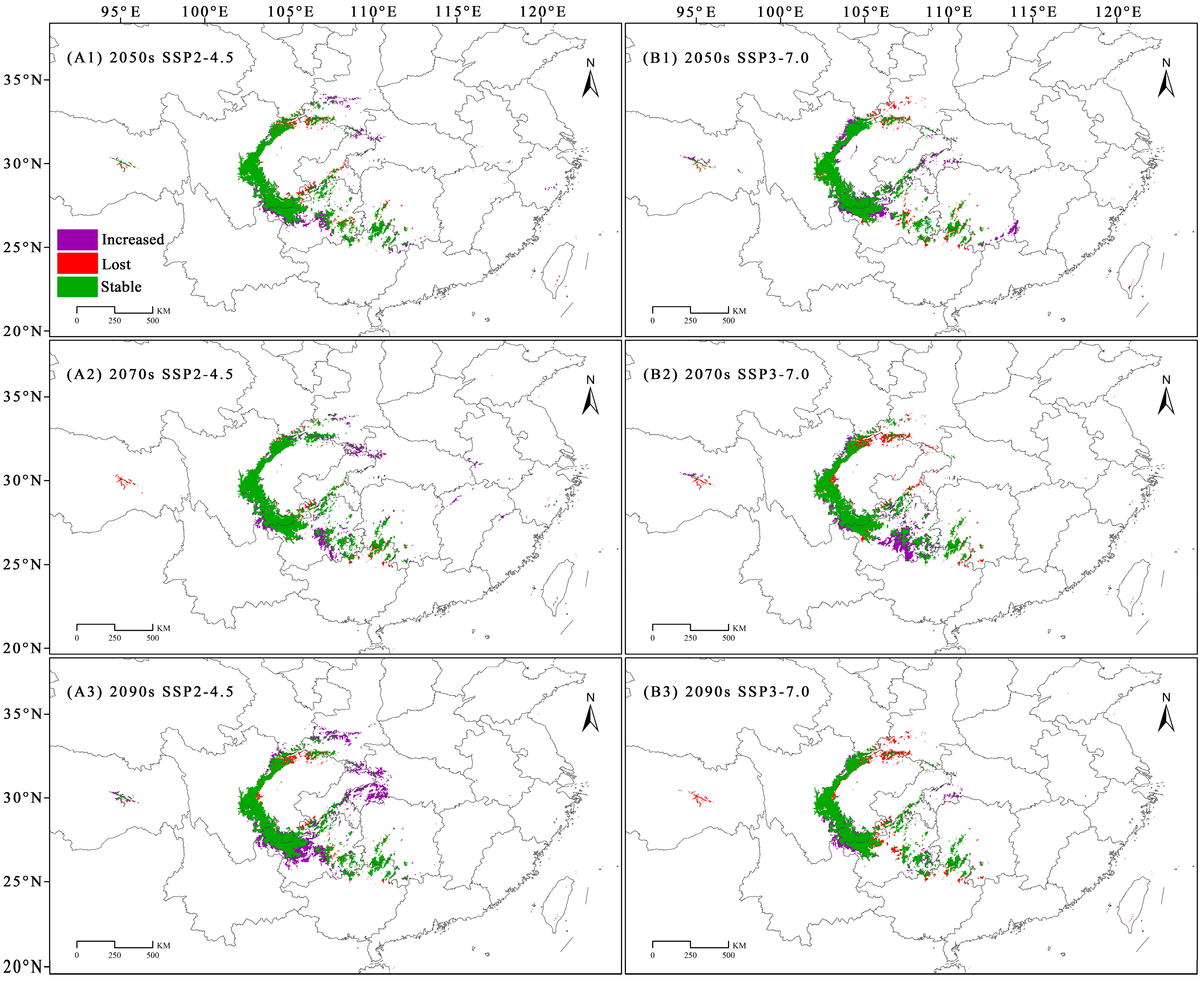
3.4. Dynamic Changes in the Geographical Distribution of C. fangiana Under Climate Change
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AUC | Area under the curve |
| BCC-CSM2-MR | Beijing Climate Center-Climate System Model version 2-Medium Resolution |
| CMIP | Coupled Model Intercomparison Project Phase |
| CSAs | Core suitable areas |
| CVH | Chinese Virtual Herbarium |
| IPCC | Intergovernmental Panel on Climate Change |
| LSAs | Low suitable areas |
| MaxEnt | Maximum Entropy |
| MSAs | Moderately suitable areas |
| RCP | Representative concentration pathway |
| SDAs | Suitable distribution areas |
| SSPs | Shared socioeconomic pathways |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Variable Description | Unit | Percent Contribution |
|---|---|---|---|
| bio12 | Annual Precipitation | mm | 34.4 |
| ele | Elevation | m | 25.3 |
| bio03 | Isothermality (bio02/bio07) (×100) | % | 7.6 |
| bio06 | Min Temperature of Coldest Month | °C | 6.9 |
| bio04 | Temperature Seasonality (standard deviation × 100) | % | 5.6 |
| bio14 | Precipitation of Driest Month | mm | 5.1 |
| bio15 | Precipitation Seasonality (Coefficient of Variation) | % | 4.0 |
| bio09 | Mean Temperature of Driest Quarter | °C | 2.9 |
| bio07 | Temperature Annual Range (bio05-bio6) | °C | 2.5 |
| bio02 | Mean Diurnal Range (Mean of monthly (max temp − min temp)) | °C | 2.0 |
| bio11 | Mean Temperature of Coldest Quarter | °C | 1.7 |
| bio17 | Precipitation of Driest Quarter | mm | 0.8 |
| bio19 | Precipitation of Coldest Quarter | mm | 0.4 |
| bio10 | Mean Temperature of Warmest Quarter | °C | 0.3 |
| bio18 | Precipitation of Warmest Quarter | mm | 0.2 |
| bio05 | Max Temperature of Warmest Month | °C | 0.2 |
| bio13 | Precipitation of Wettest Month | mm | 0.1 |
| bio08 | Mean Temperature of Wettest Quarter | °C | 0.1 |
| bio16 | Precipitation of Wettest Quarter | mm | 0 |
| bio01 | Annual Mean Temperature | °C | 0 |
Appendix B

References
- Kogan, F. The IPCC Reports on Global Warming and Land Changes. In Remote Sensing Land Surface Changes; Springer: Cham, Switzerland, 2022. [Google Scholar] [CrossRef]
- Calvin, K.; Dasgupta, D.; Krinner, G.; Mukherji, A.; Thorne, P.W.; Trisos, C.; Romero, J.; Aldunce, P.; Barrett, K.; Blanco, G.; et al. IPCC, 2023: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Lee, H., Romero, J., Eds.; IPCC: Geneva, Switzerland, 2023. [Google Scholar]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef]
- Malanoski, C.M.; Farnsworth, A.; Lunt, D.J.; Valdes, P.J.; Saupe, E.E. Climate change is an important predictor of extinction risk on macroevolutionary timescales. Science 2024, 383, 1130–1134. [Google Scholar] [CrossRef]
- Zhao, R.N.; Chu, X.J.; He, Q.Q.; Tang, Y.; Song, M.; Zhu, Z.L. Modeling current and future potential geographical distribution of Carpinus tientaiensis, a critically endangered species from China. Forests 2020, 11, 774. [Google Scholar] [CrossRef]
- Briscoe, N.J.; Morris, S.D.; Mathewson, P.D.; Buckley, L.B.; Jusup, M.; Levy, O.; Maclean, I.M.; Pincebourde, S.; Riddell, E.A.; Roberts, J.A.; et al. Mechanistic forecasts of species responses to climate change: The promise of biophysical ecology. Glob. Change Biol. 2023, 29, 1451–1470. [Google Scholar] [CrossRef]
- Franklin, J. Species distribution modelling supports the study of past, present and future biogeographies. J. Biogeogr. 2023, 50, 1533–1545. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference on Machine Learning (ICML ’04), Banff, AB, Canada, 4–8 July 2004; Association for Computing Machinery: New York, NY, USA, 2004. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Ahmadi, M.; Hemami, M.R.; Kaboli, M.; Shabani, F. MaxEnt brings comparable results when the input data are being completed; Model parameterization of four species distribution models. Ecol. Evol. 2023, 13, e9827. [Google Scholar] [CrossRef]
- Mapunda, K.K.; Andrew, S.M. Predicting the distribution of critically endangered tree species Karomia gigas under climate change in Tanzania. Ecol. Eng. 2023, 195, 107065. [Google Scholar] [CrossRef]
- Zhao, R.N.; He, Q.Q.; Chu, X.J.; He, A.G.; Zhang, Y.L.; Zhu, Z.L. Regional environmental differences significantly affect the genetic structure and genetic differentiation of Carpinus tientaiensis Cheng, an endemic and extremely endangered species from China. Front. Plant Sci. 2024, 15, 1277173. [Google Scholar] [CrossRef]
- Li, X.L.; Wu, K.N.; Hao, S.H.; Yue, Z.; Ran, Z.; Ma, J.L. Mapping cropland suitability in China using optimized MaxEnt model. Field Crop. Res. 2023, 302, 109064. [Google Scholar] [CrossRef]
- Yang, Y.; He, J.; Liu, Y.X.; Zeng, J.X.; Zeng, L.Q.; He, R.L.; Guiang, M.M.; Li, Y.Q.; Wu, H. Assessment of Chinese suitable habitats of Zanthoxylum nitidum in different climatic conditions by Maxent model, HPLC, and chemometric methods. Ind. Crops Prod. 2023, 196, 116515. [Google Scholar] [CrossRef]
- Wang, Y.J.; Zhao, R.X.; Zhou, X.Y.; Zhang, X.L.; Zhao, G.H.; Zhang, F.G. Prediction of potential distribution areas and priority protected areas of Agastache rugosa based on Maxent model and Marxan model. Front. Plant Sci. 2023, 14, 1200796. [Google Scholar] [CrossRef]
- Huang, L.L.; Li, S.F.; Huang, W.Y.; Jin, J.H.; Oskolski, A.A. Late Pleistocene glacial expansion of a low-latitude species Magnolia insignis: Megafossil evidence and species distribution modeling. Ecol. Indic. 2024, 158, 111519. [Google Scholar] [CrossRef]
- Cui, L.; Berger, U.; Cao, M.; Zhang, Y.; He, J.; Pan, L.; Jiang, J. Conservation and Restoration of Mangroves in Response to Invasion of Spartina alterniflora Based on the MaxEnt Model: A Case Study in China. Forests 2023, 14, 1220. [Google Scholar] [CrossRef]
- Sorbe, F.; Gränzig, T.; Förster, M. Evaluating sampling bias correction methods for invasive species distribution modeling in Maxent. Ecol. Inform. 2023, 76, 102124. [Google Scholar] [CrossRef]
- Li, P.Q.; Zheng, S.X. Betulaceae. In Flora Republicae Popularis Sinica 21; Kuang, K.R., Li, P.Q., Eds.; Science Press: Beijing, China, 1979; pp. 84–85. [Google Scholar]
- Li, P.C.; Skvortsov, A.K. Betulaceae. In Flora of China 4; Wu, C.Y., Raven, P.H., Eds.; Science Press: Beijing, China, 1999; pp. 289–300. [Google Scholar]
- Holstein, N.; Weigend, M. No taxon left behind? A critical taxonomic checklist of Carpinus and Ostrya (Coryloideae, Betulaceae). Eur. J. Taxon. 2017, 375, 1–52. [Google Scholar] [CrossRef]
- Xue, L.; Jia, L.B.; Nam, G.S.; Huang, Y.J.; Zhang, S.T.; Wang, Y.Q.; Zhou, Z.; Chen, Y.S. Involucre fossils of Carpinus, a northern temperate element, from the Miocene of China and the evolution of its species diversity in East Asia. Plant Divers. 2020, 42, 155–167. [Google Scholar] [CrossRef]
- Dong, C.C.; Lu, Z.Q.; Zhang, H.; Liu, J.Q.; Li, M.J. Delimiting 33 Carpinus (Betulaceae) species with a further phylogenetic inference. AoB Plants 2022, 14, plac006. [Google Scholar] [CrossRef]
- Hu, H.H. Two new species of Carpinus from Szechuan. J. Arnold Arbor. 1929, 10, 154–156. [Google Scholar] [CrossRef]
- Lancaster, R.; Rix, M. 705. Carpinus fangiana. Curtis’s Bot. Mag. 2011, 28, 103–110. [Google Scholar] [CrossRef]
- Yang, X.Y.; Wang, Z.F.; Zhang, L.; Hao, G.Q.; Liu, J.Q.; Yang, Y.Z. A chromosome-level reference genome of the hornbeam, Carpinus fangiana. Sci. Data 2020, 7, 24. [Google Scholar] [CrossRef]
- Wang, Z.F.; Jiang, Y.Z.; Yang, X.Y.; Bi, H.; Li, J.L.; Mao, X.X.; Ma, Y.Z.; Ru, D.F.; Zhang, C.; Hao, G.Q.; et al. Molecular signatures of parallel adaptive divergence causing reproductive isolation and speciation across two genera. Innovation 2022, 3, 100247. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.F.; Kang, M.H.; Li, J.L.; Zhang, Z.Y.; Wang, Y.F.; Chen, C.L.; Yang, Y.L.; Liu, J.Q. Genomic evidence for homoploid hybrid speciation between ancestors of two different genera. Nat. Commun. 2022, 13, 1987. [Google Scholar] [CrossRef]
- Chen, H.P.; Sun, J.Q.; Lin, W.Q.; Xu, H.W. Comparison of CMIP6 and CMIP5 models in simulating climate extremes. Sci. Bull. 2020, 65, 1415–1418. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Lu, Y.; Fang, Y.; Xin, X.; Li, L.; Li, W.; Jie, W.; Zhang, J.; Liu, Y.; Zhang, L.; et al. The Beijing Climate Center Climate System Model (BCC-CSM): The main progress from CMIP5 to CMIP6. Geosci. Model Dev. 2019, 12, 1573–1600. [Google Scholar] [CrossRef]
- Xin, X.; Wu, T.; Zhang, J.; Yao, J.; Fang, Y. Comparison of CMIP6 and CMIP5 simulations of precipitation in China and the East Asian summer monsoon. Int. J. Climatol. 2020, 40, 6423–6440. [Google Scholar] [CrossRef]
- Liang, S.C. A preliminary study on the structure and dynamics of pubescent hornbeam population in karst mountain of Guiyang. Chin. J. Plant Ecol. 1992, 16, 108–117. [Google Scholar]
- Verma, D.; Singh, G.; Ram, N. Carpinus viminea: A pioneer tree species of old landslide regions of Indian Himalaya. Curr. Sci. 2009, 97, 1277–1278. [Google Scholar]
- Gutierrez-Hernandez, O.; Garcia, L.V. Chapter 11-relationship between precipitation and species distribution. In Precipitation; Rodrigo-Comino, J., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2021; pp. 239–259. [Google Scholar] [CrossRef]
- Amissah, L.; Mohren, G.M.J.; Bongers, F.; Hawthorne, W.D.; Poorter, L. Rainfall and temperature affect tree species distribution in Ghana. J. Trop. Ecol. 2014, 30, 435–446. [Google Scholar] [CrossRef]
- Shi, P.J.; Preisler, H.K.; Quinn, B.K.; Zhao, J.; Huang, W.W.; Röll, A.; Cheng, X.F.; Li, H.R.; Hölscher, D. Precipitation is the most crucial factor determining the distribution of moso bamboo in Mainland China. Glob. Ecol. Conserv. 2020, 22, e00924. [Google Scholar] [CrossRef]
- Zhang, L.J.; Zhu, L.Q.; Li, Y.H.; Zhu, W.B.; Chen, Y.Y. Maxent Modelling Predicts a Shift in Suitable Habitats of a Subtropical Evergreen Tree (Cyclobalanopsis glauca (Thunberg) Oersted) under Climate Change Scenarios in China. Forests 2022, 13, 126. [Google Scholar] [CrossRef]
- Zhao, R.N.; He, Q.Q.; Chu, X.J.; Lu, Z.Q.; Zhu, Z.L. Prediction of potential distribution of Carpinus cordata in China under climate change. Chin. J. Appl. Ecol. 2019, 30, 3833–3843. [Google Scholar]
- Oke, O.A.; Thompson, K.A. Distribution models for mountain plant species: The value of elevation. Ecol. Model. 2015, 301, 72–77. [Google Scholar] [CrossRef]
- Wang, J.; Liang, S.; Shi, P. Topography and Landforms. In The Geography of Contemporary China; World Regional Geography Book Series; Springer: Cham, Switzerland, 2022. [Google Scholar]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Spence, A.R.; Tingley, M.W. The challenge of novel abiotic conditions for species undergoing climate-induced range shifts. Ecography 2020, 43, 1571–1590. [Google Scholar] [CrossRef]
- Chen, Z.D. Phylogeny and Phytogeography of the Betulaceae. Acta Phytotaxon. Sin. 1994, 32, 1–31. Available online: https://www.plantsystematics.com/CN/Y1994/V32/I1/1 (accessed on 1 January 2025).
- Chen, Z.D. Phylogeny and Phytogeography of the Betulaceae (Cont.). Acta Phytotaxon. Sin. 1994, 32, 101–153. Available online: https://www.plantsystematics.com/CN/Y1994/V32/I2/101 (accessed on 1 January 2025).
- Liang, S.C. Study on dynamics of pubescent hornbeam population in Karst Mountain of Guiyang. Acta Ecol. Sin. 1992, 12, 53–60. [Google Scholar]
- Manthey, M.; Box, E. Realized climatic niches of deciduous trees: Comparing western Eurasia and eastern North America. J. Biogeogr. 2007, 34, 1028–1040. [Google Scholar] [CrossRef]
- Carotenuto, F.; Di Febbraro, M.; Melchionna, M.; Castiglione, S.; Saggese, F.; Serio, C.; Mondanaro, A.; Passaro, F.; Loy, A.; Raia, P. The influence of climate on species distribution over time and space during the late Quaternary. Quat. Sci. Rev. 2016, 149, 188–199. [Google Scholar] [CrossRef]
- Román-Palacios, C.; Wiens, J.J. Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. USA 2020, 117, 4211–4217. [Google Scholar] [CrossRef] [PubMed]
- Tsiftsis, S.; Štípková, Z.; Rejmánek, M.; Kindlmann, P. Predictions of species distributions based only on models estimating future climate change are not reliable. Sci. Rep. 2024, 14, 25778. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, R.; He, Q.; Chu, X.; Zhu, Z. Current and Future Geographical Distribution and Potential Habitat of Carpinus fangiana Hu (Betulaceae) Under Different Climate Change Scenarios. Horticulturae 2025, 11, 575. https://doi.org/10.3390/horticulturae11060575
Zhao R, He Q, Chu X, Zhu Z. Current and Future Geographical Distribution and Potential Habitat of Carpinus fangiana Hu (Betulaceae) Under Different Climate Change Scenarios. Horticulturae. 2025; 11(6):575. https://doi.org/10.3390/horticulturae11060575
Chicago/Turabian StyleZhao, Runan, Qianqian He, Xiaojie Chu, and Zunling Zhu. 2025. "Current and Future Geographical Distribution and Potential Habitat of Carpinus fangiana Hu (Betulaceae) Under Different Climate Change Scenarios" Horticulturae 11, no. 6: 575. https://doi.org/10.3390/horticulturae11060575
APA StyleZhao, R., He, Q., Chu, X., & Zhu, Z. (2025). Current and Future Geographical Distribution and Potential Habitat of Carpinus fangiana Hu (Betulaceae) Under Different Climate Change Scenarios. Horticulturae, 11(6), 575. https://doi.org/10.3390/horticulturae11060575