
A Point Mutation of the Alpha-Tubulin Gene ClTUA Causes Dominant Dwarf Phenotype in Watermelon (Citrullus lanatus)

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Mapping Population
2.2. Cytological Observation of the Internodes
2.3. DNA Extraction and Resequencing Analysis
2.4. Fine Mapping Through KASP Markers
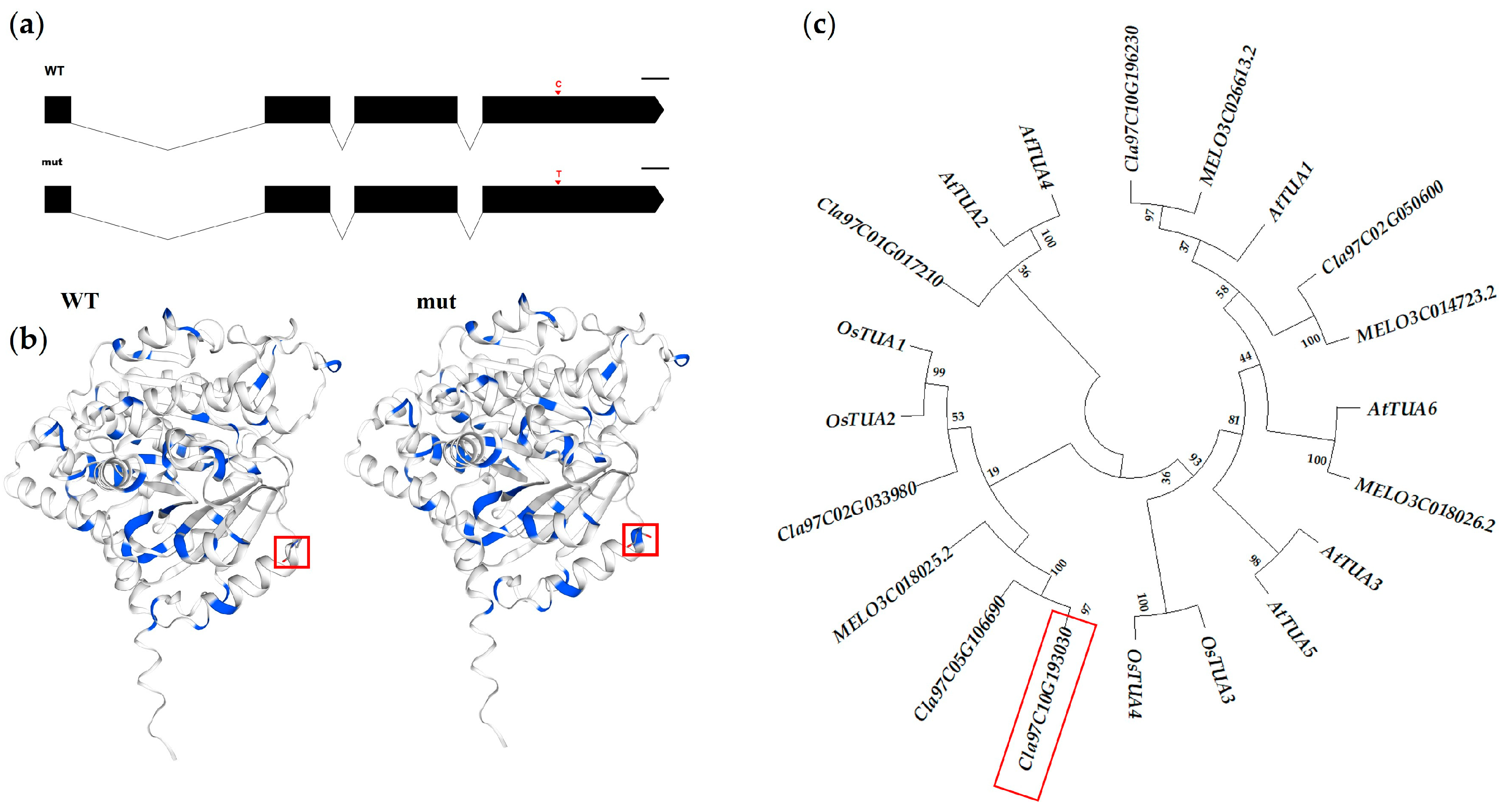
2.5. Cloning, Sequencing Analysis, and Phylogenetic Analysis of Candidate Gene
2.6. Quantitative RT-PCR and Gene Expression Analysis
2.7. Detection of Auxin Content
2.8. Application of Exogenous IAA
2.9. RNA-Seq Analysis
2.10. Statistical Analysis
3. Results
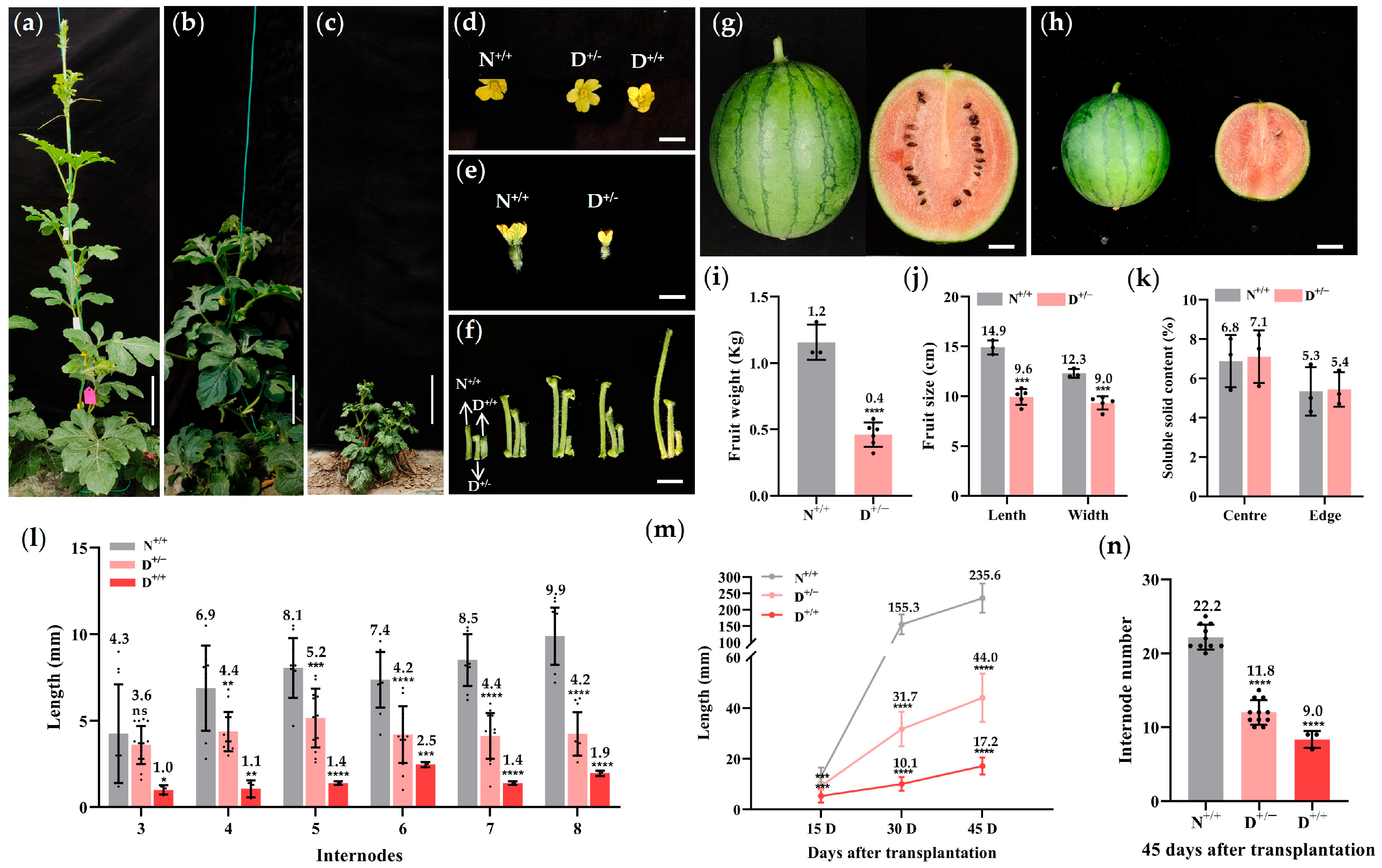
3.1. The Mutant Plants Exhibited Shortened Internodes and Dwarfism
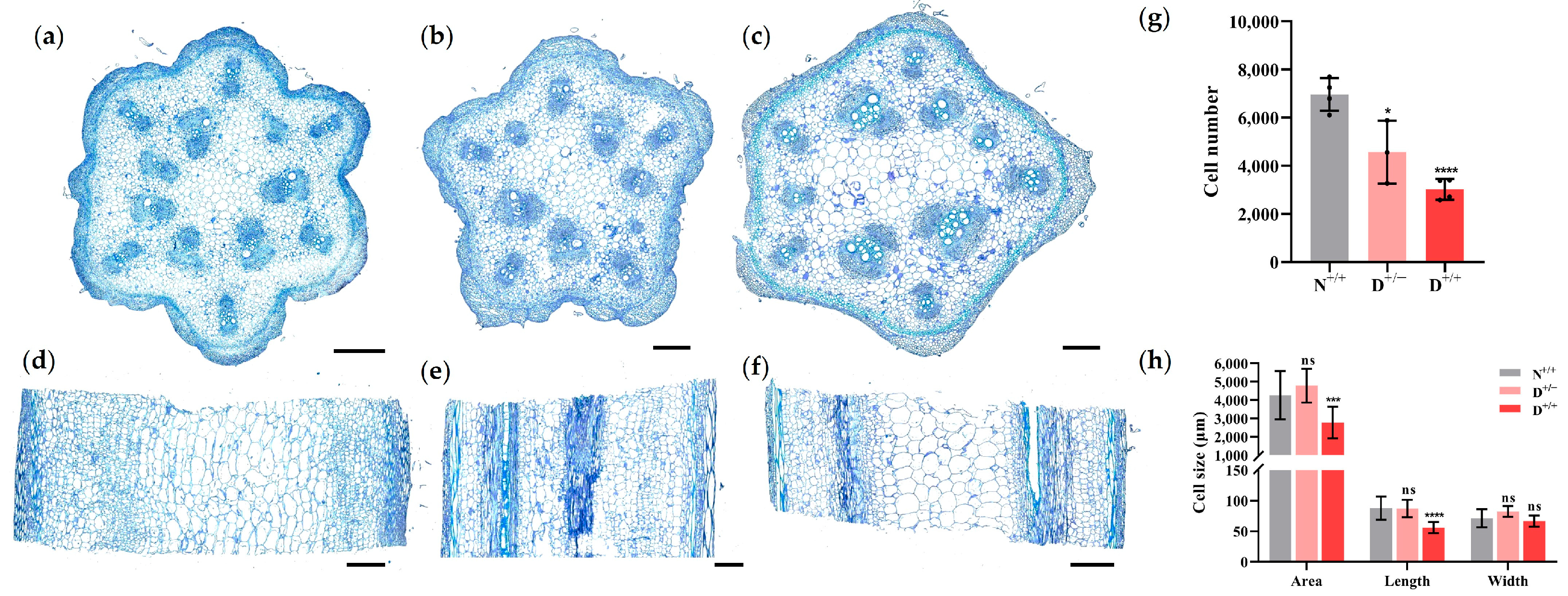
3.2. The Number of Cells Decreases in the Dwarf Plants
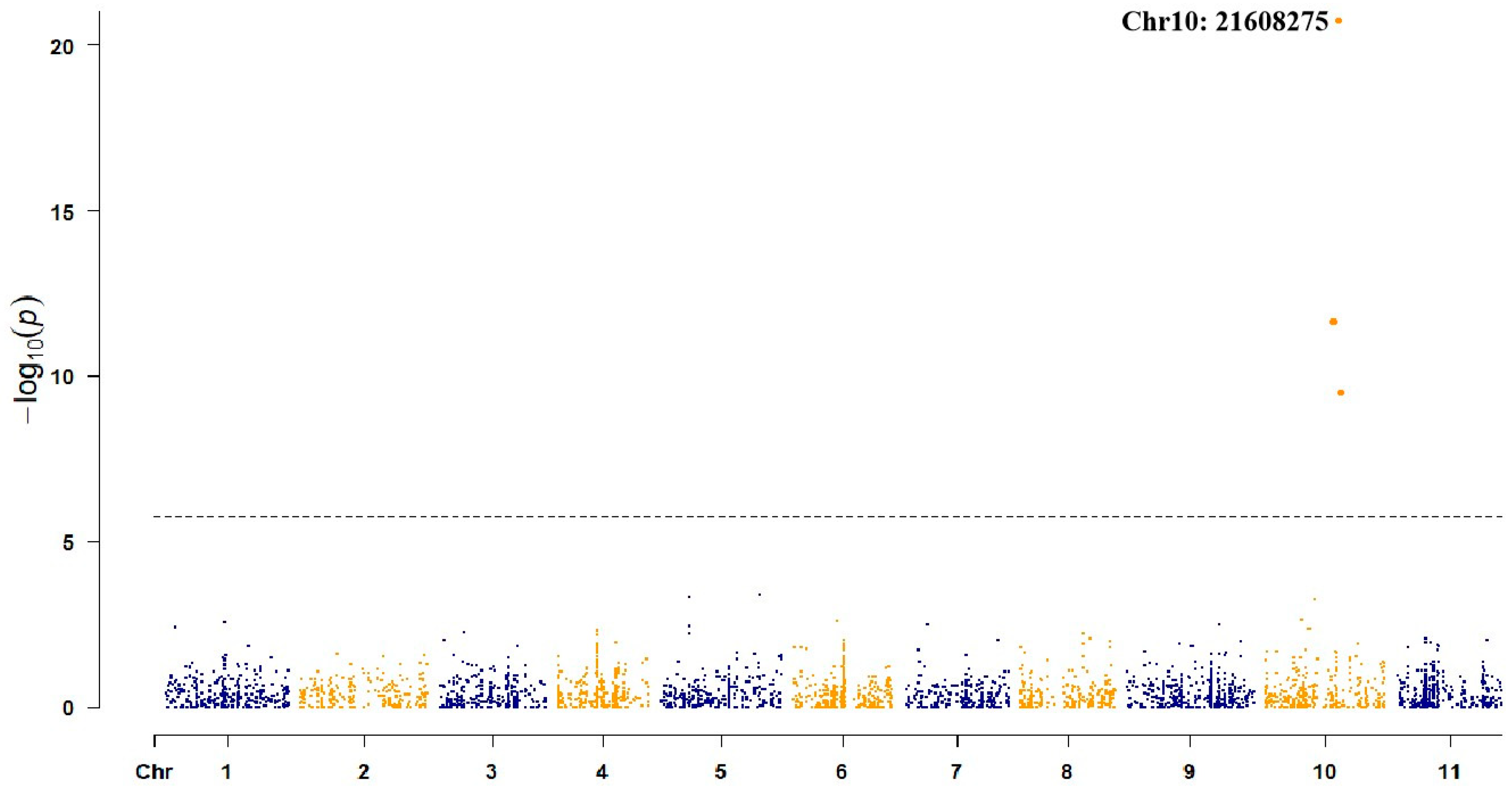
3.3. Localization of Candidate Gene
3.4. ClTUA May Respond to the Auxin Pathway to Regulate the Vine Length
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| KASP | Kompetitive Allele Specific PCR |
| IAA | Indole-3-acetic acid |
| MEIAA | Methyl indole-3-acetate |
| IAN | 3-Indoleacetonitrile |
| IAA-Asp | Indole-3-acetyl-L-aspartic acid |
| ICAld | Indole-3-carboxaldehyde |
| GO | Gene Ontology |
| BP | Biological Process |
| EMS | Ethyl methanesulfonate |
| LC-MS/MS | Liquid Chromatography–Tandem Mass Spectrometry |
| BSA-seq | Bulked Segregant Analysis by Sequencing |
| CTAB | Cetyltrimethylammonium Bromide |
References
- Dane, F.; Liu, J. Diversity and origin of cultivated and citron type watermelon (Citrullus lanatus). Genet. Resour. Crop Evol. 2006, 54, 1255–1265. [Google Scholar] [CrossRef]
- Khush, G.S. Green revolution: The way forward. Nat. Rev. Genet. 2001, 1, 815–822. [Google Scholar] [CrossRef]
- Salamini, F. Hormones and the Green Revolution. Science 2003, 302, 71–72. [Google Scholar] [CrossRef]
- Sakamoto, T.; Miura, K.; Itoh, H.; Tatsumi, T.; Ueguchi-Tanaka, M.; Ishiyama, K.; Kobayashi, M.; Agrawal, G.K.; Takeda, S.; Abe, K.; et al. An overview of gibberellin metabolism enzyme genes and their related mutants in rice. Plant Physiol. 2004, 34, 1642–1653. [Google Scholar] [CrossRef]
- Sato-Izawa, K.; Nakaba, S.; Tamura, K.; Yamagishi, Y.; Nakano, Y.; Nishikubo, N.; Kawai, S.; Kajita, S.; Ashikari, M.; Funada, R.; et al. DWARF50 (D50), a rice (Oryza sativa L.) gene encoding inositol polyphosphate 5-phosphatase, is required for proper development of intercalary meristem. Plant Cell Environ. 2012, 35, 2031–2044. [Google Scholar] [CrossRef]
- Li, W.Q.; Yoshida, A.; Takahashi, M.; Maekawa, M.; Kojima, M.; Sakakibara, H.; Kyozuka, J. SAD1, an RNA polymerase I subunit A34.5 of rice, interacts with mediator and controls various aspects of plant development. Plant J. 2015, 81, 282–291. [Google Scholar] [CrossRef]
- Ren, D.Y.; Rao, Y.C.; Wu, L.W.; Xu, Q.K.; Li, Z.Z.; Yu, H.P.; Zhang, Y.; Leng, Y.J.; Hu, J.; Zhu, L.; et al. The pleiotropic ABNORMAL FLOWER AND DWARF1 affects plant height, floral development and grain yield in rice. J. Integr. Plant Biol. 2016, 58, 529–539. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, X.; Xiong, G.S.; Liu, H.H.; Chen, F.L.; Wang, L.; Xiangbing Meng, X.B.; Liu, G.F.; Yu, H.; Yuan, Y.D.; et al. DWARF53 acts as a repressor of strigolactone signalling in rice. Nature 2013, 504, 401–405. [Google Scholar] [CrossRef]
- Zhou, M.; Li, Y.K.; Cheng, Z.W.; Zheng, X.Y. Important factors controlling gibberellin homeostasis in plant height regulation. J. Agric. Food Chem. 2023, 71, 15895–15907. [Google Scholar] [CrossRef]
- Wang, Y.J.; Deng, D.X.; Ding, H.D.; Xu, X.M.; Zhang, R.; Wang, S.X.; Bian, Y.L.; Yin, Z.T.; Chen, Y. Gibberellin biosynthetic deficiency is responsible for maize dominant dwarf11 (D11) mutant phenotype: Physiological and transcriptomic evidence. PLoS ONE 2013, 8, e66466. [Google Scholar] [CrossRef]
- Cassani, E.; Bertolini, E.; Badone, F.C.; Landoni, M. Characterization of the first dominant dwarf maize mutant carrying a single amino acid insertion in the VHYNP domain of the dwarf 8 gene. Mol. Breed. 2009, 24, 375–385. [Google Scholar] [CrossRef]
- Mohr, H.C. Mode of inheritance of bushy growth characteristic in watermelon. Proc. Assoc. South Agric. Work. 1956, 53, 174. [Google Scholar]
- Dyutin, K.E.; Afanas’eva, E.A. Interaction of short stem in watermelon. Tsitologiya I Genet. 1987, 21, 227–229. [Google Scholar]
- Robinson, R.W.; Munger, H.M.; Whitake, T.W. Genes of Cucubitaceae. Hortscience 1976, 11, 554–568. [Google Scholar] [CrossRef]
- Huang, H.X.; Zhang, X.Q.; Wei, Z.C.; Li, Q.H.; Li, X. Inheritance of male-sterility and dwarfism in watermelon [Citrullus lanatus (Thunb.) Matsum. and Nakai]. Sci. Hortic. 1998, 74, 175–181. [Google Scholar]
- Li, Y.G.; Xu, A.J.; Dong, W.; LI, Z.; Li, G.S. Genetic analysis of a dwarf vine and small fruit watermelon mutant. Hortic. Plant J. 2016, 2, 224–228. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, J.; Sun, H.; Salse, J.; Lucas, W.J.; Zhang, H.; Zheng, Y.; Mao, L.; Ren, Y.; Wang, Z.; et al. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nat. Genet. 2013, 45, 51–58. [Google Scholar] [CrossRef]
- Deng, Y.; Liu, S.C.; Zhang, Y.L.; Tan, J.S.; Li, X.P.; Chu, X.; Xu, B.H.; Tian, Y.; Sun, Y.D.; Li, B.S.; et al. A telomere-to-telomere gap-free reference genome of watermelon and its mutation library provide important resources for gene discovery and breeding. Mol. Plant 2022, 15, 1268–1284. [Google Scholar] [CrossRef]
- Wei, C.H.; Zhu, C.Y.; Yang, L.P.; Zhao, W.; Ma, R.X.; Li, H.; Zhang, Y.; Ma, J.X.; Yang, J.Q.; Zhang, X. A point mutation resulting in a 13 bp deletion in the coding sequence of Cldf leads to a GA-deficient dwarf phenotype in watermelon. Hortic. Res. 2019, 6, 132. [Google Scholar] [CrossRef]
- Gebremeskel, H.; Dou, J.L.; Li, B.B.; Zhao, S.J.; Muhammad, U.; Lu, X.Q.; He, N.; Liu, W.G. Molecular Mapping and Candidate Gene Analysis for GA3 Responsive Short Internode in Watermelon (Citrullus lanatus). Int. J. Mol. Sci. 2020, 21, 290. [Google Scholar] [CrossRef]
- Jang, Y.J.; Yun, H.S.; Rhee, S.J.; Seo, M.; Kim, Y.J.; Lee, G.P. Exploring molecular markers and candidate genes responsible for watermelon dwarfism. Hortic. Environ. Biote. 2020, 61, 173–182. [Google Scholar] [CrossRef]
- Dong, W.; Wu, D.; Li, G.; Wu, D.W.; Wang, Z.C. Next-generation sequencing from bulked segregant analysis identifies a dwarfism gene in watermelon. Sci. Rep. 2018, 8, 2908. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, M.; Sun, S.; Yang, S.; Li, J.X.; Li, H.; Yang, H.H.; Zhang, K.G.; Hu, J.B.; Liu, D.M.; et al. A single nucleotide deletion in an ABC transporter gene leads to a dwarf phenotype in watermelon. Front. Plant Sci. 2019, 10, 1399. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Niu, Q.; Zong, Y.; Qian, M.; Yang, F.; Teng, Y. Simultaneous quantitative determination of major plant hormones in pear flowers and fruit by UPLC/ESI-MS/MS. Anal. Methods 2014, 6, 1766. [Google Scholar] [CrossRef]
- Shen, X.P.; Xu, L.A.; Liu, Y.H.; Dong, H.; Zhou, D.; Zhang, Y.Z.; Lin, S.; Cao, J.S.; Huang, L. Comparative transcriptome analysis and ChIP-sequencing reveals stage-specific gene expression and regulation profiles associated with pollen wall formation in Brassica rapa. BMC Genom. 2019, 20, 264. [Google Scholar] [CrossRef]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids. Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L.; et al. WEGO: A web tool for plotting GO annotations. Nucleic Acids. Res. 2006, 34, W293–W297. [Google Scholar] [CrossRef]
- Liu, P.B.W.; Loy, J.B. Inheritance and morphology of two dwarf mutants in watermelon. Am. Soc. Hortic. Sci. 1972, 97, 745–748. [Google Scholar] [CrossRef]
- Navarro-Quiles, C.; Daniel, L.S.; Muñoz-Nortes, T.; Candela, H.; Micol, J.L. The genetic and molecular basis of haploinsufficiency in flowering plants. Trends Plant Sci. 2024, 29, 72–85. [Google Scholar] [CrossRef]
- Wilkie, A. Themolecular basis of genetic dominance. J. Med. Genet. 1994, 31, 89–98. [Google Scholar] [CrossRef]
- Cleary, J.M.; Hancock, W.O. Molecular mechanisms underlying microtubule growth dynamics. Curr. Biol. 2021, 31, R560–R573. [Google Scholar] [CrossRef]
- Wasteneys, G.O. Progress in understanding the role of microtubules in plant cells. Curr. Opin. Plant Biol. 2004, 7, 651–660. [Google Scholar] [CrossRef]
- Su, B.H.; Guo, Y.; Li, Z.F.; Gao, H.w.; Wei, Z.Y.; Qiu, L.J. Ideal type 1 is caused by a point mutation in the a-tubulin gene that affects microtubule arrangement in soybean. Crop J. 2023, 11, 713–722. [Google Scholar] [CrossRef]
- Xu, C.S.; Chen, B.T.; Huang, S.J.; Deng, Z.Y.; Wang, T. A point mutation in the rice alpha-tubulin gene OsTUBA3 causes grain notching. New Phytol. 2023, 240, 1052–1065. [Google Scholar] [CrossRef]
- Sun, T.P. Gibberellin-GID1-DELLA: A pivotal regulatory module for plant growth and development. Plant Physiol. 2010, 154, 567–570. [Google Scholar] [CrossRef]
- Castorina, G.; Consonni, G. The role of brassinosteroids in controlling plant height in poaceae: A genetic perspective. Int. J. Mol. Sci. 2020, 21, 1191. [Google Scholar] [CrossRef]
- Yamamuro, C.; Ihara, Y.; Wu, X.; Noguchi, T.; Fujioka, S.; Takatsuto, S.; Ashikari, M.; Kitano, H.; Matsuoka, M. Loss of function of a rice brassinosteroid insensitive1 homolog prevents internode elongation and bending of the lamina joint. Plant Cell 2000, 12, 1591–1606. [Google Scholar] [CrossRef]
- Chono, M.; Honda, I.; Zeniya, H.; Yoneyama, K.; Saisho, D.; Akeda, K.; Takatsuto, T.; Hoshino, T.; Watanabe, Y. A semidwarf phenotype of barley uzu results from a nucleotide substitution in the gene encoding a putative brassinosteroid receptor. Plant Physiol. 2003, 133, 1209–1219. [Google Scholar] [CrossRef]
- Mano, Y.; Nemoto, K. The pathway of auxin biosynthesis in plants. J. Exp. Bot. 2012, 63, 2853–2872. [Google Scholar] [CrossRef]
- Chen, X.; Grandont, L.; Li, H.; Hauschild, R.; Paque, S.; Abuzeineh, A.; Rakusová, H.; Benkova, E.; Perrot-Rechenmann, C.; Friml, J. Inhibition of cell expansion by rapid ABP1-mediated auxin effect on microtubules. Nature 2014, 516, 90–93. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Z.; Zhang, L.; Shi, J.; Ying, Q.; Zhang, H.; Zhang, X.; Deng, Y.; Wang, Y. A Point Mutation of the Alpha-Tubulin Gene ClTUA Causes Dominant Dwarf Phenotype in Watermelon (Citrullus lanatus). Horticulturae 2025, 11, 562. https://doi.org/10.3390/horticulturae11060562
Hu Z, Zhang L, Shi J, Ying Q, Zhang H, Zhang X, Deng Y, Wang Y. A Point Mutation of the Alpha-Tubulin Gene ClTUA Causes Dominant Dwarf Phenotype in Watermelon (Citrullus lanatus). Horticulturae. 2025; 11(6):562. https://doi.org/10.3390/horticulturae11060562
Chicago/Turabian StyleHu, Ziwei, Leichen Zhang, Jun Shi, Quansheng Ying, Huafeng Zhang, Xingping Zhang, Yun Deng, and Yuhong Wang. 2025. "A Point Mutation of the Alpha-Tubulin Gene ClTUA Causes Dominant Dwarf Phenotype in Watermelon (Citrullus lanatus)" Horticulturae 11, no. 6: 562. https://doi.org/10.3390/horticulturae11060562
APA StyleHu, Z., Zhang, L., Shi, J., Ying, Q., Zhang, H., Zhang, X., Deng, Y., & Wang, Y. (2025). A Point Mutation of the Alpha-Tubulin Gene ClTUA Causes Dominant Dwarf Phenotype in Watermelon (Citrullus lanatus). Horticulturae, 11(6), 562. https://doi.org/10.3390/horticulturae11060562