

Integrated Transcriptomic and Metabolomic Provide Insights into Strawberry Bioactive Compounds and Antioxidant Activities Under Varying Ecological Conditions

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strawberry Varieties, Environmental Data Collection, and Cultivation Conditions
2.2. Strawberry Metabolomic Extraction and Analysis
2.3. Metabolic Operating System Condition
2.3.1. Electrospray Ionization-Mass Spectrometry (ESI-MS/MS) Techniques
2.3.2. Multiple Reactions Monitoring System
2.4. RNA Sequencing
2.5. Hydroxyl Radical Scavenging Assay
2.6. Alkaloid Content and Antioxidant Properties
2.7. Statistical Analysis
3. Results
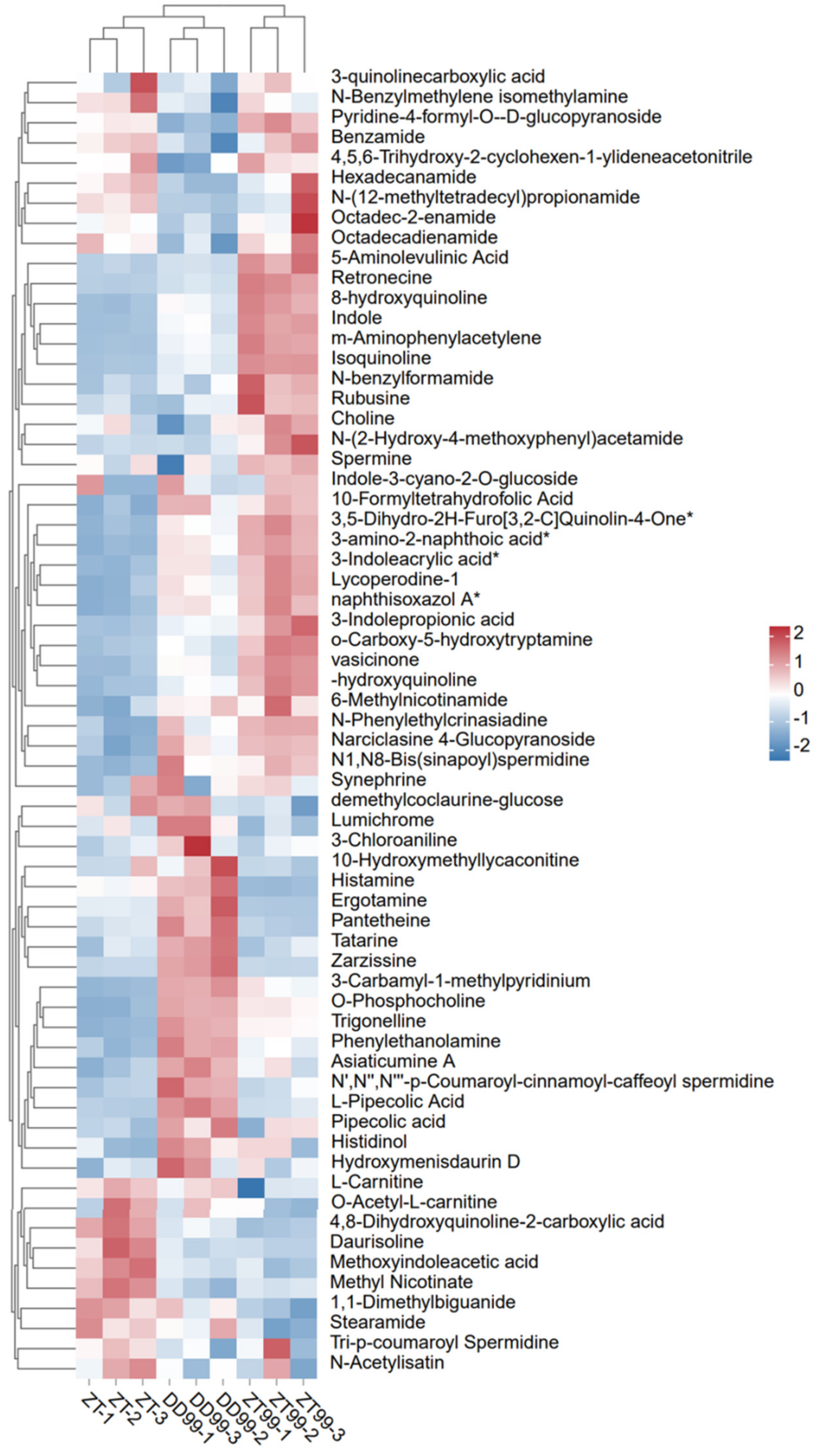
3.1. Clustering Analysis of Bioactive Compounds from Strawberry Fruit
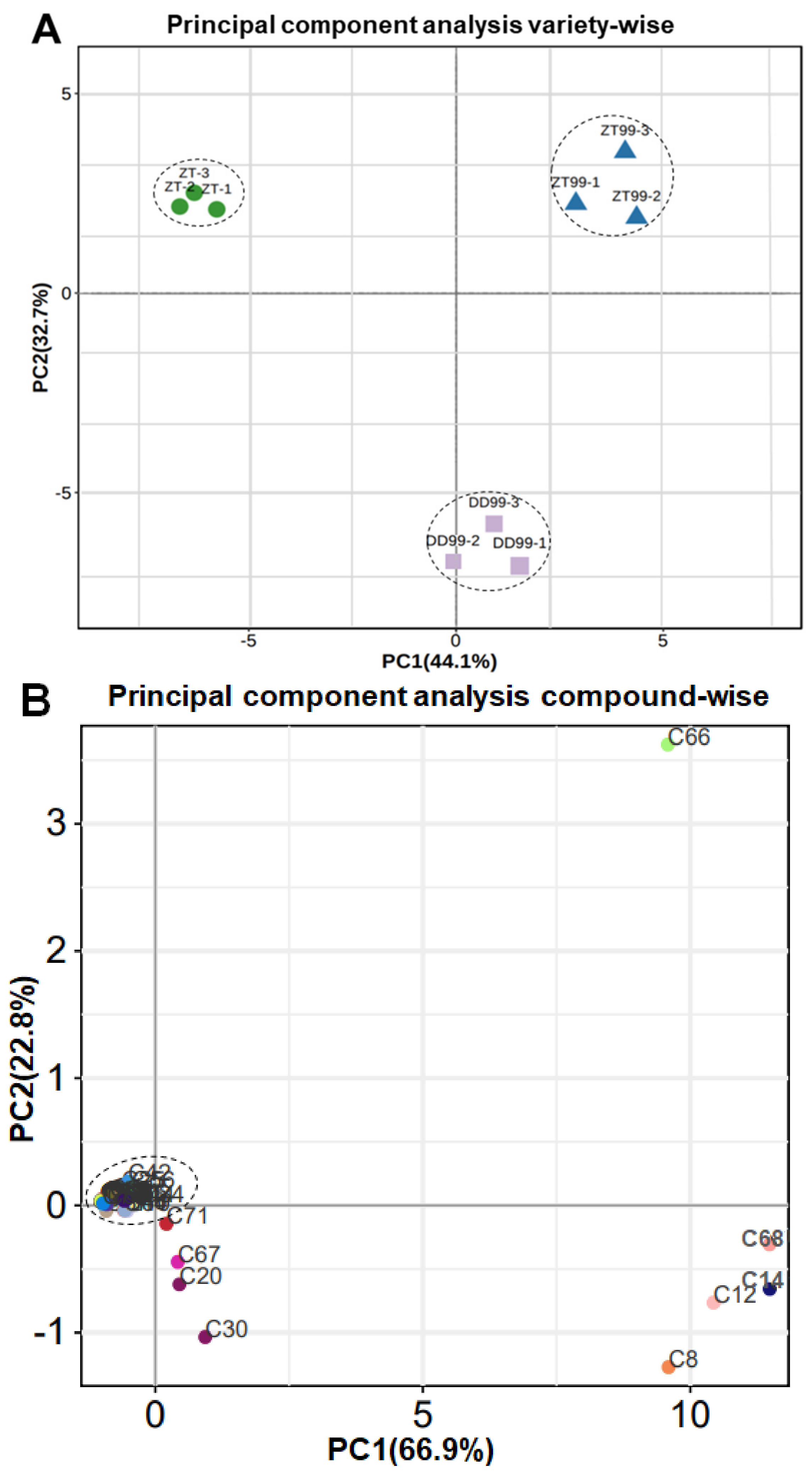
3.2. Principal Component Analysis of Bioactive Compounds from Strawberry Fruit
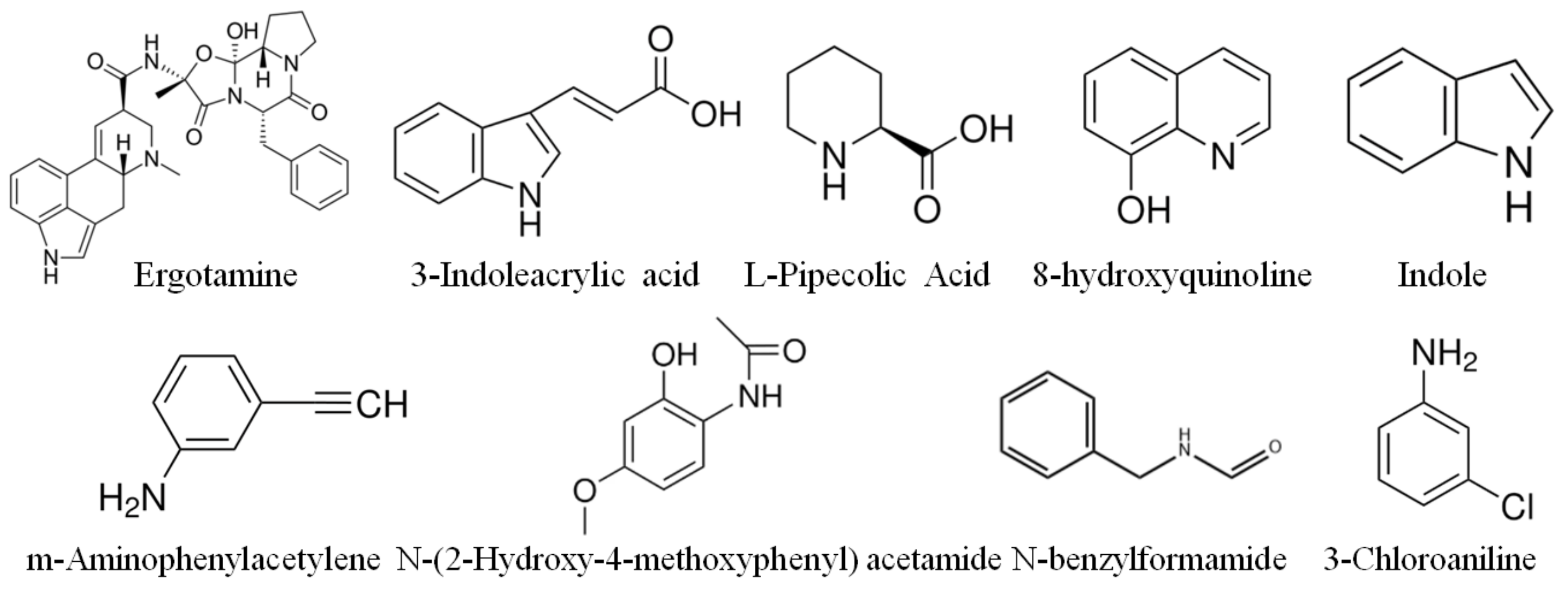
3.3. Chemical Structure of Major Strawberry Alkaloids
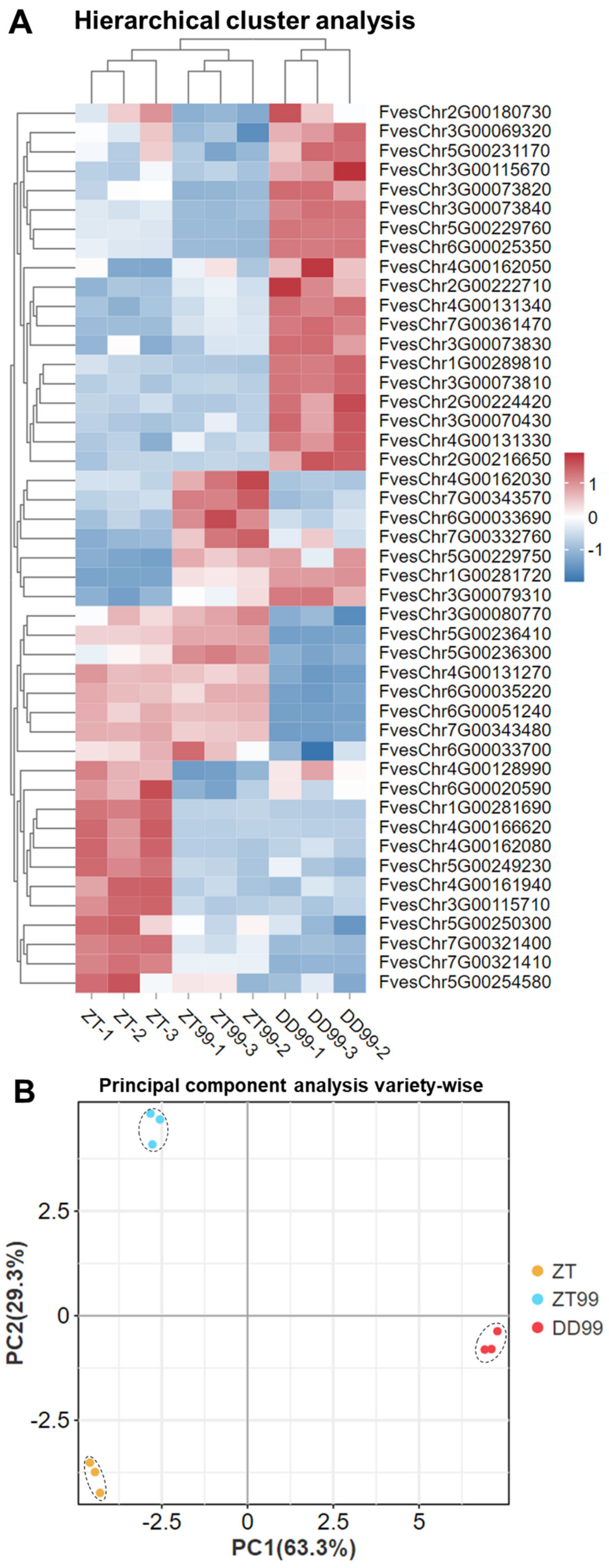
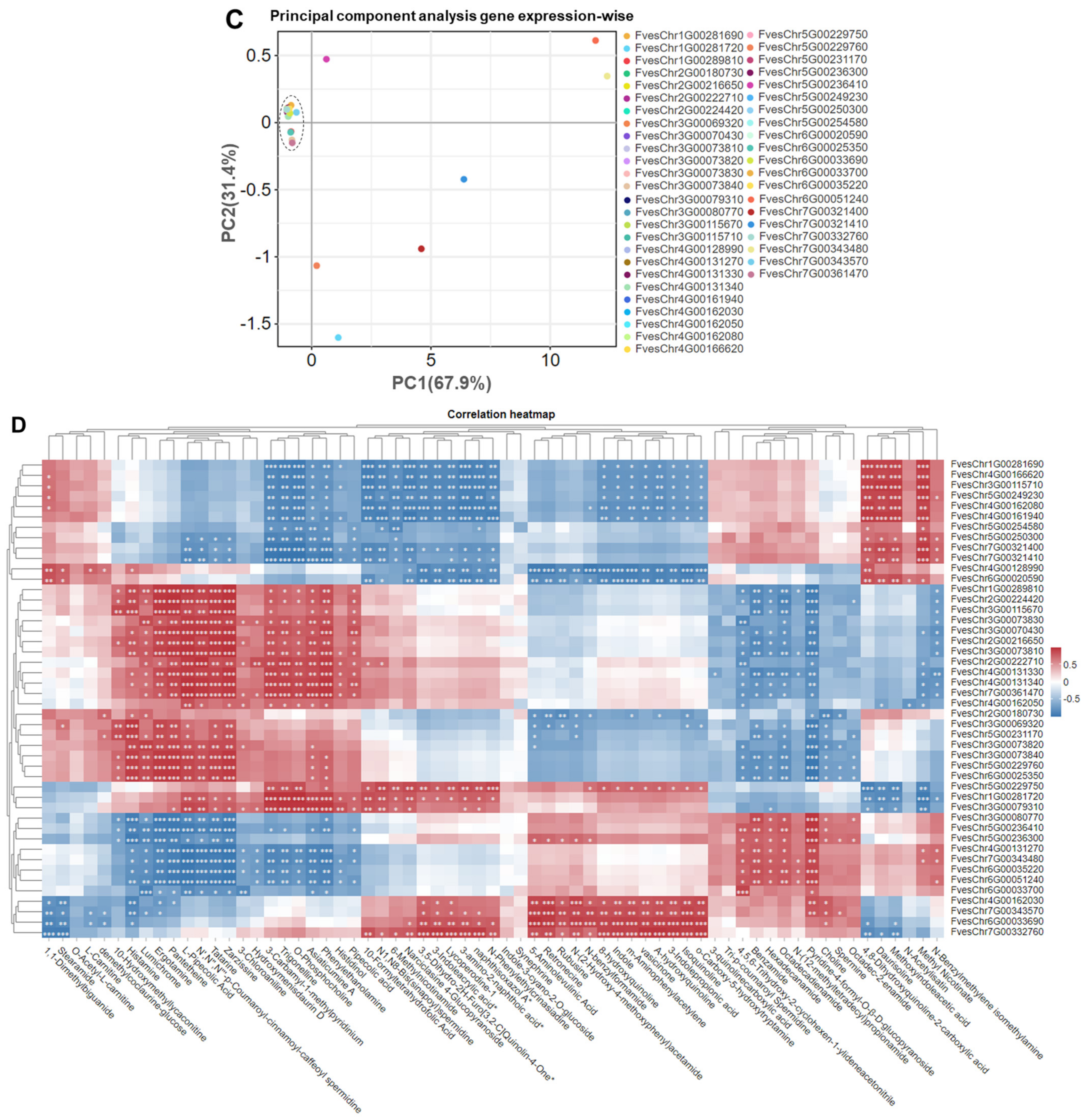
3.4. Clustering and PCA Analysis of Gene Expressions from Strawberry Fruits
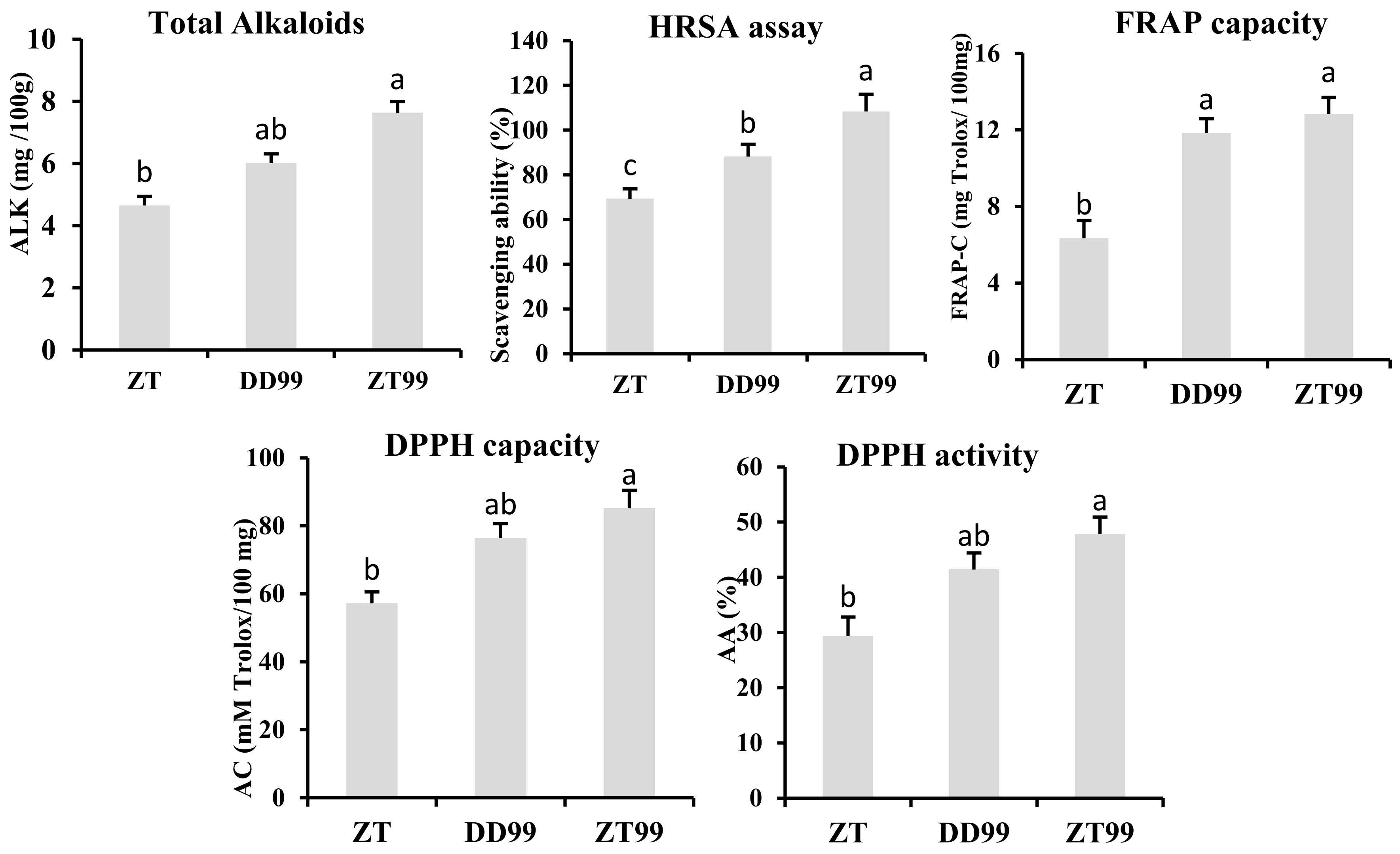
3.5. Total Alkaloids and Bioactivity in Strawberry Fruits
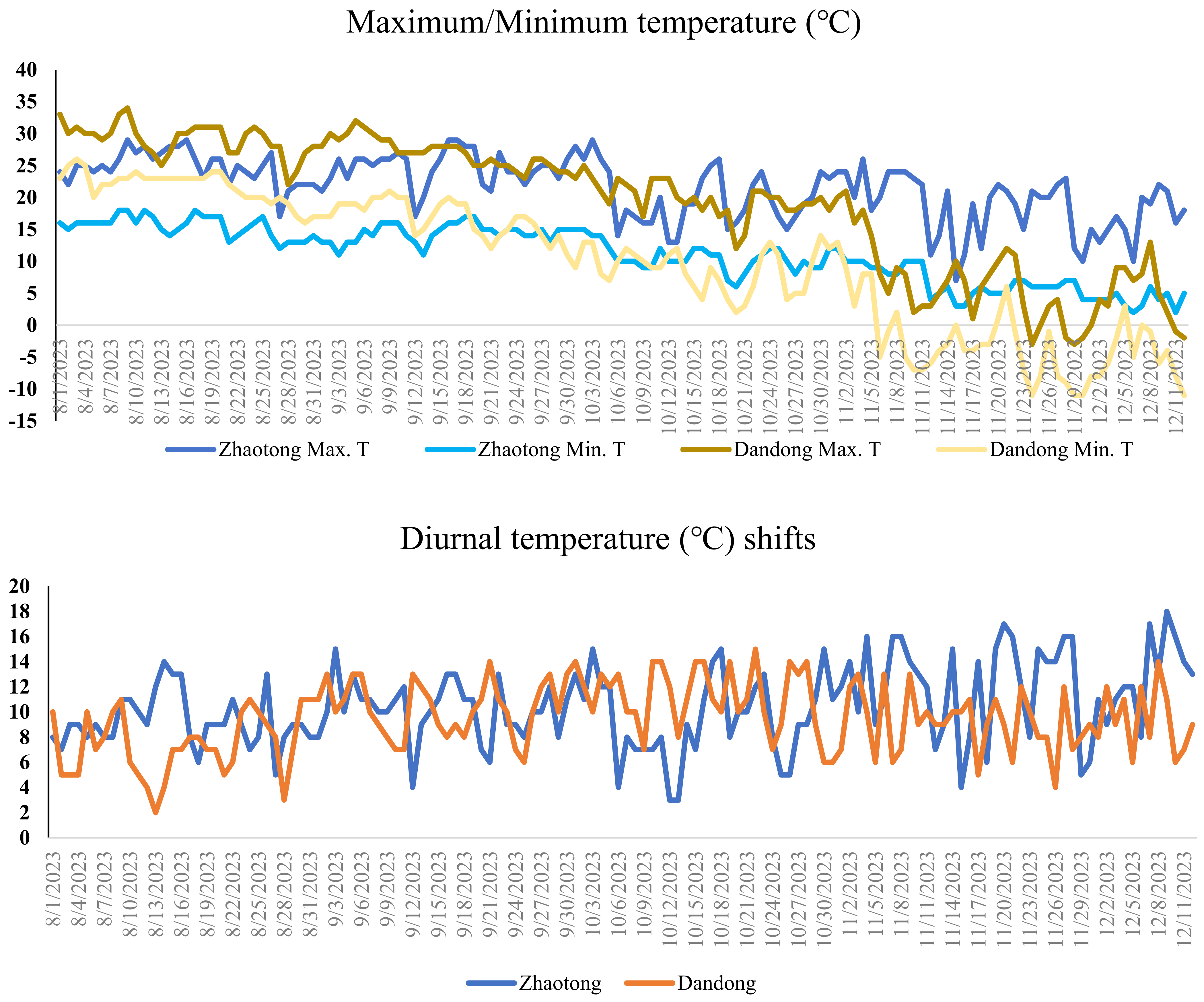
3.6. Environmental Conditions During Strawberry Cultivation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simpson, D. The Economic Importance of Strawberry Crops. In The Genomes of Rosaceous Berries and Their Wild Relatives; Springer: Cham, Switzerland, 2018; pp. 1–7. [Google Scholar]
- Arias, F.; Appelmann, T.; Giraldo, L.F.G. Import and Export of Strawberries in a Global Market. In Ciencias económicas y contables, desafíos y retos para la competitividad; Sepúlveda, J.A., Ed.; Corporación Universitaria Americana (Coruniamericana): Medellín, Colombia, 2020; pp. 280–291. [Google Scholar]
- Rao, M.J.; Zheng, B. The Role of Polyphenols in Abiotic Stress Tolerance and Their Antioxidant Properties to Scavenge Reactive Oxygen Species and Free Radicals. Antioxidants 2025, 14, 74. [Google Scholar] [CrossRef] [PubMed]
- Ordidge, M.; García-Macías, P.; Battey, N.H.; Gordon, M.H.; Hadley, P.; John, P.; Lovegrove, J.A.; Vysini, E.; Wagstaffe, A. Phenolic Contents of Lettuce, Strawberry, Raspberry, and Blueberry Crops Cultivated under Plastic Films Varying in Ultraviolet Transparency. Food Chem. 2010, 119, 1224–1227. [Google Scholar] [CrossRef]
- Panico, A.M.; Garufi, F.; Nitto, S.; Di Mauro, R.; Longhitano, R.C.; Magrì, G.; Catalfo, A.; Serrentino, M.E.; De Guidi, G. Antioxidant Activity and Phenolic Content of Strawberry Genotypes from Fragaria x Ananassa. Pharm. Biol. 2009, 47, 203–208. [Google Scholar] [CrossRef]
- Figuerola, F.E. Berry Jams and Jellies. In Food Science and Technology; Marcel Dekker: New York, NY, USA, 2007; Volume 168, p. 367. [Google Scholar]
- Aguilera, J.M. Berries as Foods: Processing, Products, and Health Implications. Annu. Rev. Food Sci. Technol. 2024, 15, 1–26. [Google Scholar] [CrossRef]
- Hannum, S.M. Potential Impact of Strawberries on Human Health: A Review of the Science. Crit. Rev. Food Sci. Nutr. 2004, 44, 1–17. [Google Scholar] [CrossRef]
- Giampieri, F.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Alvarez-Suarez, J.M.; Afrin, S.; Bompadre, S.; Quiles, J.L.; Mezzetti, B.; Battino, M. Strawberry as a Health Promoter: An Evidence Based Review. Food Funct. 2015, 6, 1386–1398. [Google Scholar] [CrossRef]
- Taher, M.; Shukry, N.A.S.B.; Susanti, D.; Saleh, W.M.N.H.W.; Syukri, Y. Citrus Flavonoids in Preventing Cardiovascular Diseases. In Plant-Derived Bioactives; Springer: Singapore, 2020; pp. 495–508. [Google Scholar]
- Devi, K.P.; Rajavel, T.; Nabavi, S.F.; Setzer, W.N.; Ahmadi, A.; Mansouri, K.; Nabavi, S.M. Hesperidin: A Promising Anticancer Agent from Nature. Ind. Crops Prod. 2015, 76, 582–589. [Google Scholar] [CrossRef]
- Ramakrishnan, R. Anticancer Properties of Zingiber Officinale–Ginger: A Review. Int. J. Med. Pharm. Sci. 2013, 3, 11–20. [Google Scholar]
- Benavente-Garcia, O.; Castillo, J. Update on Uses and Properties of Citrus Flavonoids: New Findings in Anticancer, Cardiovascular, and Anti-Inflammatory Activity. J. Agric. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef]
- Lei, J.J.; Jiang, S.; Ma, R.Y.; Xue, L.; Zhao, J.; Dai, H.P. Current Status of Strawberry Industry in China. In Proceedings of the IX International Strawberry Symposium, Rimini, Italy, 1–5 May 2021; Volume 1309, pp. 349–352. [Google Scholar]
- Zhang, Y.; Wang, G.; Chang, L.; Dong, J.; Zhong, C.; Wang, L. Current Status of Strawberry Production and Research in China. Acta Hortic. 2014, 1049, 67–71. [Google Scholar] [CrossRef]
- Kun, Q.; Yuenan, L.; Yingxue, L. Analysis of Dandong 99 Strawberry Market Competitiveness Based on Porter’s Five Forces Model. In E3S Web of Conferences; EDP Sciences: Les Ulis, France, 2021; Volume 253, p. 03058. [Google Scholar]
- Rao, M.J.; Wang, H.; Lei, H.; Zhang, H.; Duan, X.; Bao, L.; Yang, C.; Han, D.; Zhang, Y.; Yang, S.; et al. LC-MS/MS-Based Metabolomic Study Provides Insights into Altitude-Dependent Variations in Flavonoid Profiles of Strawberries. Front. Plant Sci. 2025, 15, 1527212. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Alvarez-Suarez, J.M.; Battino, M. Strawberry and Human Health: Effects beyond Antioxidant Activity. J. Agric. Food Chem. 2014, 62, 3867–3876. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Tulipani, S.; Alvarez-Suarez, J.M.; Quiles, J.L.; Mezzetti, B.; Battino, M. The Strawberry: Composition, Nutritional Quality, and Impact on Human Health. Nutrition 2012, 28, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Batool, A.; Yaqub, S.; Iqbal, A.; Kauser, S.; Arif, M.R.; Ali, S.; Gorsi, F.I.; Nisar, R.; Firdous, N. Effects of Spray Drying and Ultrasonic Assisted Extraction on the Phytochemicals, Antioxidant and Antimicrobial Activities of Strawberry Fruit. Food Chem. Adv. 2024, 5, 100755. [Google Scholar] [CrossRef]
- Hanhineva, K.; Rogachev, I.; Kokko, H.; Mintz-Oron, S.; Venger, I.; Kärenlampi, S.; Aharoni, A. Non-Targeted Analysis of Spatial Metabolite Composition in Strawberry (Fragaria × Ananassa) Flowers. Phytochemistry 2008, 69, 2463–2481. [Google Scholar] [CrossRef]
- Rao, M.J.; Duan, M.; Wang, J.; Han, S.; Ma, L.; Mo, X.; Li, M.; Hu, L.; Wang, L. Transcriptomic and Widely Targeted Metabolomic Approach Identified Diverse Group of Bioactive Compounds, Antiradical Activities, and Their Associated Genes in Six Sugarcane Varieties. Antioxidants 2022, 11, 1319. [Google Scholar] [CrossRef]
- Rao, M.J.; Tahir Ul Qamar, M.; Wang, D.; Ali, Q.; Ma, L.; Han, S.; Duan, M.; Hu, L.; Wang, L. A High-Throughput Lipidomics and Transcriptomic Approach Reveals Novel Compounds from Sugarcane Linked with Promising Therapeutic Potential against COVID-19. Front. Nutr. 2022, 9, 988249. [Google Scholar] [CrossRef]
- Rao, M.J.; Duan, M.; Wei, X.; Zuo, H.; Ma, L.; Tahir Ul Qamar, M.; Li, M.; Han, S.; Hu, L.; Wang, L. LC–MS/MS-Based Metabolomics Approach Revealed Novel Phytocompounds from Sugarcane Rind with Promising Pharmacological Value. J. Sci. Food Agric. 2022, 102, 6632–6642. [Google Scholar] [CrossRef]
- Rao, M.J.; Feng, B.; Ahmad, M.H.; Tahir Ul Qamar, M.; Aslam, M.Z.; Khalid, M.F.; Hussain, S.B.; Zhong, R.; Ali, Q.; Xu, Q.; et al. LC-MS/MS-Based Metabolomics Approach Identified Novel Antioxidant Flavonoids Associated with Drought Tolerance in Citrus Species. Front. Plant Sci. 2023, 14, 1150854. [Google Scholar] [CrossRef]
- Rao, M.J.; Duan, M.; Eman, M.; Yuan, H.; Sharma, A.; Zheng, B. Comparative Analysis of Citrus Species’ Flavonoid Metabolism, Gene Expression Profiling, and Their Antioxidant Capacity under Drought Stress. Antioxidants 2024, 13, 1149. [Google Scholar] [CrossRef]
- Rao, M.J.; Duan, M.; Shad, M.A.; Aslam, M.Z.; Wang, J.; Wang, L. Widely Targeted LC-MS/MS Approach Provides Insights into Variations in Bioactive Flavonoid Compounds and Their Antioxidant Activities in Green, Red, and Purple Sugarcane. LWT Food Sci. Technol. 2024, 209, 116792. [Google Scholar] [CrossRef]
- George, V.C.; Dellaire, G.; Rupasinghe, H.P.V. Plant Flavonoids in Cancer Chemoprevention: Role in Genome Stability. J. Nutr. Biochem. 2017, 45, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Perin, E.C.; da Silva Messias, R.; Borowski, J.M.; Crizel, R.L.; Schott, I.B.; Carvalho, I.R.; Rombaldi, C.V.; Galli, V. ABA-Dependent Salt and Drought Stress Improve Strawberry Fruit Quality. Food Chem. 2019, 271, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Pedrozo, P.; Vicente, E.; Moltini, A.I.; Ibáñez, F.; Lado, B.; Fariña, L.; Ares, G.; Lado, J. Strawberry Fruit Quality: Impacts of the Harvest Date with a Breeding Perspective. JSFA Rep. 2023, 3, 597–608. [Google Scholar] [CrossRef]
- Abouelenein, D.; Acquaticci, L.; Alessandroni, L.; Borsetta, G.; Caprioli, G.; Mannozzi, C.; Marconi, R.; Piatti, D.; Santanatoglia, A.; Sagratini, G. Volatile Profile of Strawberry Fruits and Influence of Different Drying Methods on Their Aroma and Flavor: A Review. Molecules 2023, 28, 5810. [Google Scholar] [CrossRef]
- Cordenunsi, B.R.; do Nascimento, J.R.O.; Lajolo, F.M. Physico-Chemical Changes Related to Quality of Five Strawberry Fruit Cultivars during Cool-Storage. Food Chem. 2003, 83, 167–173. [Google Scholar] [CrossRef]
- Xu, Y.; Charles, M.T.; Luo, Z.; Mimee, B.; Veronneau, P.-Y.; Rolland, D.; Roussel, D. Preharvest Ultraviolet C Irradiation Increased the Level of Polyphenol Accumulation and Flavonoid Pathway Gene Expression in Strawberry Fruit. J. Agric. Food Chem. 2017, 65, 9970–9979. [Google Scholar] [CrossRef]
- Ullah, I.; Toor, M.D.; Yerlikaya, B.A.; Mohamed, H.I.; Yerlikaya, S.; Basit, A.; ur Rehman, A. High-Temperature Stress in Strawberry: Understanding Physiological, Biochemical and Molecular Responses. Planta 2024, 260, 118. [Google Scholar] [CrossRef]
- Karlund, A.; Hanhineva, K.; Lehtonen, M.; Karjalainen, R.O.; Sandell, M. Nontargeted Metabolite Profiles and Sensory Properties of Strawberry Cultivars Grown Both Organically and Conventionally. J. Agric. Food Chem. 2015, 63, 1010–1019. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, B.-C.; Ma, J.; Cheng, C.-G.; Wang, K.-Y.; Mao, L.-X.; He, Y.-B.; Qiu, M.-J.; Yang, X.-J. Evaluation of Potential Climatic Production of Apple during the Possible Growing Period at Zhaotong, Yunnan across Cool Highland of Southwest China. Chin. J. Agrometeorol. 2021, 42, 87. [Google Scholar]
- Reganold, J.P.; Andrews, P.K.; Reeve, J.R.; Carpenter-Boggs, L.; Schadt, C.W.; Alldredge, J.R.; Ross, C.F.; Davies, N.M.; Zhou, J. Fruit and Soil Quality of Organic and Conventional Strawberry Agroecosystems. PLoS ONE 2010, 5, e12346. [Google Scholar] [CrossRef]
- Bottoms, T.G.; Bolda, M.P.; Gaskell, M.L.; Hartz, T.K. Determination of Strawberry Nutrient Optimum Ranges through Diagnosis and Recommendation Integrated System Analysis. Horttechnology 2013, 23, 312–318. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A Novel Integrated Method for Large-Scale Detection, Identification, and Quantification of Widely Targeted Metabolites: Application in the Study of Rice Metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.J.; Duan, M.; Yang, M.; Fan, H.; Shen, S.; Hu, L.; Wang, L. Novel Insights into Anthocyanin Metabolism and Molecular Characterization of Associated Genes in Sugarcane Rinds Using the Metabolome and Transcriptome. Int. J. Mol. Sci. 2022, 23, 338. [Google Scholar] [CrossRef]
- Gan, J.; Feng, Y.; He, Z.; Li, X.; Zhang, H. Correlations between Antioxidant Activity and Alkaloids and Phenols of Maca (Lepidium meyenii). J. Food Qual. 2017, 2017, 3185945. [Google Scholar] [CrossRef]
- Damiano, S.; Forino, M.; De, A.; Vitali, L.A.; Lupidi, G.; Taglialatela-Scafati, O. Antioxidant and Antibiofilm Activities of Secondary Metabolites from Ziziphus Jujuba Leaves Used for Infusion Preparation. Food Chem. 2017, 230, 24–29. [Google Scholar] [CrossRef]
- Dudonne, S.; Vitrac, X.; Coutiere, P.; Woillez, M.; Mérillon, J.-M. Comparative Study of Antioxidant Properties and Total Phenolic Content of 30 Plant Extracts of Industrial Interest Using DPPH, ABTS, FRAP, SOD, and ORAC Assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- Jing, H.; Liu, J.; Liu, H.; Xin, H. Histochemical Investigation and Kinds of Alkaloids in Leaves of Different Developmental Stages in Thymus Quinquecostatus. Sci. World J. 2014, 2014, 839548. [Google Scholar] [CrossRef]
- Swamy, M.K. Plant-Derived Bioactives: Chemistry and Mode of Action; Springer Nature: Singapore, 2020; ISBN 9789811523618. [Google Scholar]
- Jan, R.; Asaf, S.; Numan, M.; Lubna; Kim, K.-M. Plant Secondary Metabolite Biosynthesis and Transcriptional Regulation in Response to Biotic and Abiotic Stress Conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Hussain, S.A.; Ahmad, S.; Butt, Z.A.; Rehman, K.U.; Ullah, S.; Khan, S.S. Flavonoids, Alkaloids, and Saponins as Antimicrobial Agents from Fragaria vesca L. Pure Appl. Biol. (PAB) 2021, 10, 761–769. [Google Scholar] [CrossRef]
- Stynoski, J.L.; Torres-Mendoza, Y.; Sasa-Marin, M.; Saporito, R.A. Evidence of Maternal Provisioning of Alkaloid Based Chemical Defenses in the Strawberry Poison Frog Oophaga Pumilio. Ecology 2014, 95, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Ko, M.J.; Jayaramaiah, R.H.; Gupta, R.; Kim, S.W.; An, J.U.; Jziyu, W.; Li, M.; Kang, N.J.; Hong, K.-P.; Kang, J.-S. Evaluation of Bioactive Compounds in Strawberry Fruits by a Targeted Metabolomic Approach. Hortic. Sci. Technol. 2017, 35, 805–819. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, L.; Chen, J.; Zhang, N.; Zhou, W.; Song, Y. Altitudinal Variation of Dragon Fruit Metabolite Profiles as Revealed by UPLC-MS/MS-Based Widely Targeted Metabolomics Analysis. BMC Plant Biol. 2024, 24, 344. [Google Scholar] [CrossRef] [PubMed]
- Naryal, A.; Dolkar, D.; Bhardwaj, A.K.; Kant, A.; Chaurasia, O.P.; Stobdan, T. Effect of Altitude on the Phenology and Fruit Quality Attributes of Apricot (Prunus armeniaca L.) Fruits. Def. Life Sci. J. 2020, 5, 18–24. [Google Scholar] [CrossRef]
- Zeng, Q.; Dong, G.; Tian, L.; Wu, H.; Ren, Y.; Tamir, G.; Huang, W.; Yu, H. High Altitude Is Beneficial for Antioxidant Components and Sweetness Accumulation of Rabbiteye Blueberry. Front. Plant Sci. 2020, 11, 573531. [Google Scholar] [CrossRef]
- Rao, M.J.; Wu, S.; Duan, M.; Wang, L. Antioxidant Metabolites in Primitive, Wild, and Cultivated Citrus and Their Role in Stress Tolerance. Molecules 2021, 26, 5801. [Google Scholar] [CrossRef]
- Liu, Q.; Gao, T.; Liu, W.; Liu, Y.; Zhao, Y.; Liu, Y.; Li, W.; Ding, K.; Ma, F.; Li, C. Functions of Dopamine in Plants: A Review. Plant Signal Behav. 2020, 15, 1827782. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Li, X. Dopamine-Induced Abiotic Stress Tolerance in Horticultural Plants. Sci. Hortic. 2023, 307, 111506. [Google Scholar] [CrossRef]
- Du, J.; Xu, H.; Zhang, D.; Feng, S. Chelation and Nanoparticle Delivery of Monomeric Dopamine to Increase Plant Salt Stress Resistance. Nat. Commun. 2025, 16, 4157. [Google Scholar] [CrossRef]
- Jancewicz, A.L.; Gibbs, N.M.; Masson, P.H. Cadaverine’s Functional Role in Plant Development and Environmental Response. Front. Plant Sci. 2016, 7, 870. [Google Scholar] [CrossRef]
- Choudhary, M.I.; Adhikari, A.; Rasheed, S.; Marasini, B.P.; Hussain, N.; Kaleem, W.A. Cyclopeptide Alkaloids of Ziziphus Oxyphylla Edgw as Novel Inhibitors of α-Glucosidase Enzyme and Protein Glycation. Phytochem. Lett. 2011, 4, 404–406. [Google Scholar] [CrossRef]
- Choi, J.S.; Kim, J.-H.; Ali, M.Y.; Min, B.-S.; Kim, G.-D.; Jung, H.A. Coptis Chinensis Alkaloids Exert Anti-Adipogenic Activity on 3T3-L1 Adipocytes by Downregulating C/EBP-α and PPAR-γ. Fitoterapia 2014, 98, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Bowman, W.C.; Sanghvi, I.S. Pharmacological Actions of Hemlock (Conium Maculatum) Alkaloids. J. Pharm. Pharmacol. 1963, 15, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.J.; Duan, M.; Zhou, C.; Jiao, J.; Cheng, P.; Yang, L.; Wei, W.; Shen, Q.; Ji, P.; Yang, Y.; et al. Antioxidant Defense System in Plants: Reactive Oxygen Species Production, Signaling, and Scavenging During Abiotic Stress-Induced Oxidative Damage. Horticulturae 2025, 11, 477. [Google Scholar] [CrossRef]
- Thurmond, R.L.; Gelfand, E.W.; Dunford, P.J. The Role of Histamine H1 and H4 Receptors in Allergic Inflammation: The Search for New Antihistamines. Nat. Rev. Drug Discov. 2008, 7, 41–53. [Google Scholar] [CrossRef]
- Sharma, N.; Sharma, V.K.; Manikyam, H.K.; Krishna, A.B. Ergot Alkaloids: A Review on Therapeutic Applications. Eur. J. Med. Plants 2016, 14, 1–17. [Google Scholar] [CrossRef]
- Tóth, F.; Cseh, E.K.; Vécsei, L. Natural Molecules and Neuroprotection: Kynurenic Acid, Pantethine and α-Lipoic Acid. Int. J. Mol. Sci. 2021, 22, 403. [Google Scholar] [CrossRef]
- Nguyen, V.; Taine, E.G.; Meng, D.; Cui, T.; Tan, W. Pharmacological Activities, Therapeutic Effects, and Mechanistic Actions of Trigonelline. Int. J. Mol. Sci. 2024, 25, 3385. [Google Scholar] [CrossRef]
- Kim, N.Y.; Dukanya, D.; Sethi, G.; Girimanchanaika, S.S.; Yang, J.; Nagaraja, O.; Swamynayaka, A.; Vishwanath, D.; Venkantesha, K.; Basappa, S. Oxazine Drug-Seed Induces Paraptosis and Apoptosis through Reactive Oxygen Species/JNK Pathway in Human Breast Cancer Cells. Transl. Oncol. 2024, 49, 102101. [Google Scholar] [CrossRef]
- Hartmann, M.; Kim, D.; Bernsdorff, F.; Ajami-Rashidi, Z.; Scholten, N.; Schreiber, S.; Zeier, T.; Schuck, S.; Reichel-Deland, V.; Zeier, J. Biochemical Principles and Functional Aspects of Pipecolic Acid Biosynthesis in Plant Immunity. Plant Physiol. 2017, 174, 124–153. [Google Scholar] [CrossRef]
- Chen, H.; Yu, C.; Wu, H.; Li, G.; Li, C.; Hong, W.; Yang, X.; Wang, H.; You, X. Recent Advances in Histidine Kinase-Targeted Antimicrobial Agents. Front. Chem. 2022, 10, 866392. [Google Scholar] [CrossRef] [PubMed]
- Monti, S.M.; De Simone, G.; D’Ambrosio, K. L-Histidinol Dehydrogenase as a New Target for Old Diseases. Curr. Top. Med. Chem. 2016, 16, 2369–2378. [Google Scholar] [CrossRef] [PubMed]
- Krakowska-Sieprawska, A.; Walczak-Skierska, J.; Pomastowski, P.; Sobolewska, R.; Głogowski, J.; Bernat, C.; Rafińska, K. Advanced Extraction Techniques for Bioactive Compounds from Berry Fruits: Enhancing Functional Food Applications. Foods 2024, 13, 4115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jin, Y.; Wu, Y.; Zhang, C.; Jin, D.; Zheng, Q.; Li, Y. Anti-Hyperglycemic and Anti-Hyperlipidemia Effects of the Alkaloid-Rich Extract from Barks of Litsea Glutinosa in Ob/Ob Mice. Sci. Rep. 2018, 8, 12646. [Google Scholar] [CrossRef]
- Pegg, A.E. The Function of Spermine. IUBMB Life 2014, 66, 8–18. [Google Scholar] [CrossRef]
- Shi, J.; Fu, X.Z.; Peng, T.; Huang, X.S.; Fan, Q.J.; Liu, J.H. Spermine Pretreatment Confers Dehydration Tolerance of Citrus In Vitro Plants via Modulation of Antioxidative Capacity and Stomatal Response. Tree Physiol. 2010, 30, 914–923. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial No | Bioactive Compounds | Q1 (Da) | Q3 (Da) | Molecular Weight | Formula | Ionization Model | Class | CAS | cpd_ID |
|---|---|---|---|---|---|---|---|---|---|
| 1 | A1 | 699.4 | 699.4 | 698.3 | C38H46N6O7 | [M + H]+ | Other compounds | - | - |
| 2 | 1,1-Dimethylbiguanide | 130.1 | 71.1 | 129.1 | C4H11N5 | [M + H]+ | Guanidine compounds | 657-24-9 | C07151 |
| 3 | A2 | 427.3 | 427.3 | 426.3 | C26H36NO4+ | [M + H]+ | Other compounds | - | - |
| 4 | A3 | 221.1 | 175.1 | 220.1 | C11H12N2O3 | [M + H]+ | Quinazolinones | - | - |
| 5 | A4 | 474.2 | 327.1 | 473.2 | C20H23N7O7 | [M + H]+ | tetrahydrofolates | 2800-34-2 | C00234 |
| 6 | A5 | 699.4 | 699.4 | 698.3 | C37H50N2O11 | [M + H]+ | Other alkaloids | - | - |
| 7 | A6 | 438.1 | 306.1 | 439.1 | C19H25N3O7S | [M − H]− | Amino acid | - | - |
| 8 | A7 | 188.1 | 118.1 | 187.1 | C11H9NO2 | [M + H]+ | Quinoline alkaloids | - | - |
| 9 | A8 | 226.2 | 180.2 | 225.2 | C12H23N3O | [M + H]+ | Other compounds | - | - |
| 10 | A9 | 138.1 | 94.1 | 137.1 | C7H9N2O | [M + H]+ | Pyridine alkaloid | 1005-24-9 | C02918 |
| 11 | 3-Chloroaniline | 128.0 | 93.1 | 127.0 | C6H6ClN | [M + H]+ | Aromatic amine | 108-42-9 | - |
| 12 | 3-Indoleacrylic acid * | 188.1 | 118.1 | 187.1 | C11H9NO2 | [M + H]+ | Plumerane | 1204-06-4 | C21283 |
| 13 | 3-Indolepropionic acid | 190.1 | 118.1 | 189.1 | C11H11NO2 | [M + H]+ | Plumerane | 830-96-6 | C22236 |
| 14 | 3-amino-2-naphthoic acid * | 188.1 | 118.1 | 187.1 | C11H9NO2 | [M + H]+ | Naphthoic acid | - | - |
| 15 | 3-quinolinecarboxylic acid | 190.1 | 144.0 | 189.0 | C10H7NO3 | [M + H]+ | Quinoline alkaloids | - | - |
| 16 | A10 | 168.1 | 105.0 | 167.1 | C8H9NO3 | [M + H]+ | Other alkaloids | - | - |
| 17 | A11 | 204.0 | 160.0 | 205.0 | C10H7NO4 | [M − H]− | Quinoline alkaloids | 59-00-7 | C02470 |
| 18 | 5-Aminolevulinic acid | 132.1 | 86.1 | 131.1 | C5H9NO3 | [M + H]+ | Delta-amino acid | 106-60-5 | C00430 |
| 19 | 6-Methylnicotinamide | 137.1 | 65.0 | 136.1 | C7H8N2O | [M + H]+ | Pyridine alkaloid | 6960-22-1 | - |
| 20 | 8-hydroxyquinoline | 146.1 | 91.1 | 145.1 | C9H7NO | [M + H]+ | Quinoline alkaloids | 148-24-3 | C19434 |
| 21 | Asiaticumine A | 284.1 | 152.1 | 283.1 | C16H13NO4 | [M + H]+ | Alkaloids | - | - |
| 22 | Benzamide | 122.1 | 105.0 | 121.1 | C7H7NO | [M + H]+ | Aromatic amides | 55-21-0 | C09815 |
| 23 | Choline | 104.1 | 60.1 | 104.1 | C5H14NO+ | [M]+ | Cholines | 62-49-7 | C00114 |
| 24 | Daurisoline | 609.3 | 417.2 | 610.3 | C37H42N2O6 | [M − H]− | Isoquinoline alkaloid | 70553-76-3 | - |
| 25 | Ergotamine | 580.4 | 506.3 | 581.3 | C33H35N5O5 | [M − H]− | Plumerane | 113-15-5 | C07544 |
| 26 | Hexadecanamide | 256.3 | 88.1 | 255.3 | C16H33NO | [M + H]+ | Fatty amide | 629-54-9 | - |
| 27 | Histamine | 112.1 | 95.1 | 111.1 | C5H9N3 | [M + H]+ | Imidazole | 51-45-6 | C00388 |
| 28 | Histidinol | 142.1 | 124.1 | 141.1 | C6H11N3O | [M + H]+ | Aralkylamine | 501-28-0 | C00860 |
| 29 | Hydroxymenisdaurin D | 332.1 | 152.1 | 331.1 | C14H21NO8 | [M + H]+ | Alkaloid | - | - |
| 30 | Indole | 118.1 | 91.1 | 117.1 | C8H7N | [M + H]+ | Plumerane | 120-72-9 | C00463 |
| 31 | Indole-3-cyano-2-O-glucoside | 349.1 | 187.0 | 350.1 | C16H18N2O5S | [M − H]− | Plumerane | - | - |
| 32 | Isoquinoline | 130.1 | 103.1 | 129.1 | C9H7N | [M + H]+ | Isoquinoline alkaloid | 119-65-3 | C06323 |
| 33 | L-Carnitine | 162.1 | 85.0 | 161.1 | C7H15NO3 | [M + H]+ | Quaternary ammonium | 541-15-1 | C00318 |
| 34 | L-Pipecolic acid | 130.1 | 84.1 | 129.1 | C6H11NO2 | [M + H]+ | Piperidine alkaloid | 3105-95-1 | C00408 |
| 35 | Lumichrome | 243.1 | 145.1 | 242.1 | C12H10N4O2 | [M + H]+ | Flavin | 1086-80-2 | C01727 |
| 36 | Lycoperodine-1 | 217.1 | 144.1 | 216.1 | C12H12N2O2 | [M + H]+ | β-carbolines alkaloid | 42438-90-4 | - |
| 37 | A12 | 206.1 | 145.1 | 205.1 | C11H11NO3 | [M + H]+ | Plumerane | 3471-31-6 | C05660 |
| 38 | A13 | 584.3 | 325.0 | 583.3 | C34H37N3O6 | [M + H]+ | Phenolamine | - | - |
| 39 | A14 | 284.3 | 88.1 | 283.3 | C18H37NO | [M + H]+ | Other compounds | - | - |
| 40 | A15 | 182.1 | 91.1 | 181.1 | C9H11NO3 | [M + H]+ | Other alkaloids | 58469-06-0 | - |
| 41 | N-Acetylisatin | 188.0 | 144.1 | 189.0 | C10H7NO3 | [M − H]− | Plumerane | 574-17-4 | C02172 |
| 42 | A16 | 120.1 | 103.1 | 119.1 | C8H9N | [M + H]+ | Other compounds | - | - |
| 43 | N-Phenylethylcrinasiadine | 344.1 | 147.0 | 343.1 | C22H17NO3 | [M + H]+ | Other compounds | - | - |
| 44 | N-benzylformamide | 136.1 | 91.1 | 135.1 | C8H9NO | [M + H]+ | Formamides | 6343-54-0 | C15561 |
| 45 | A17 | 558.3 | 207.1 | 557.3 | C29H39N3O8 | [M + H]+ | Phenolamine | - | C21170 |
| 46 | A18 | 470.2 | 131.1 | 469.2 | C20H27N3O8S | [M + H]+ | Amino acid derivative | - | - |
| 47 | A19 | 470.1 | 308.1 | 469.1 | C20H23NO12 | [M + H]+ | Piperidine alkaloid | - | - |
| 48 | A20 | 138.1 | 78.0 | 137.0 | C7H7NO2 | [M + H]+ | Pyridine alkaloid | 93-60-7 | - |
| 49 | O-Acetyl-L-carnitine | 204.1 | 85.0 | 203.1 | C9H17NO4 | [M + H]+ | Acyl carnitines | 3040-38-8 | - |
| 50 | O-Phosphocholine | 184.1 | 125.0 | 184.1 | C5H15NO4P+ | [M]+ | Phospholipids | 107-73-3 | C00588 |
| 51 | Octadec-2-enamide | 282.3 | 69.1 | 281.3 | C18H35NO | [M + H]+ | Fatty acids | - | - |
| 52 | Octadecadienamide | 280.3 | 81.1 | 279.3 | C18H33NO | [M + H]+ | Alkylamides | - | - |
| 53 | Pantetheine | 277.1 | 146.1 | 278.1 | C11H22N2O4S | [M − H]− | Amide | 496-65-1 | C00831 |
| 54 | Phenylethanolamine | 138.1 | 120.1 | 137.1 | C8H11NO | [M + H]+ | Monoamine alkaloid | 7568-93-6 | C02735 |
| 55 | Pipecolic acid | 130.1 | 56.1 | 129.1 | C6H11NO2 | [M + H]+ | Piperidine alkaloid | 535-75-1 | - |
| 56 | A21 | 286.1 | 124.0 | 285.1 | C12H15NO7 | [M + H]+ | Pyridine alkaloid | - | - |
| 57 | Retronecine | 156.1 | 110.1 | 155.1 | C8H13NO2 | [M + H]+ | Pyrrole alkaloid | 480-85-3 | C06177 |
| 58 | Rubusine | 190.1 | 128.1 | 189.0 | C10H7NO3 | [M + H]+ | Polyphenol | - | - |
| 59 | Spermine | 203.2 | 84.1 | 202.2 | C10H26N4 | [M + H]+ | Polyamine | 71-44-3 | C00750 |
| 60 | Stearamide | 284.3 | 102.1 | 283.3 | C18H37NO | [M + H]+ | Fatty amide | 124-26-5 | C13846 |
| 61 | Synephrine | 168.1 | 150.1 | 167.1 | C9H13NO2 | [M + H]+ | Phenethylamine alkaloid | 94-07-5 | C04548 |
| 62 | Tatarine | 325.1 | 191.1 | 324.1 | C15H20N2O6 | [M + H]+ | Other alkaloids | - | - |
| 63 | Tri-p-coumaroyl Spermidine | 584.3 | 438.2 | 583.3 | C34H37N3O6 | [M + H]+ | Phenolamine | - | - |
| 64 | Trigonelline | 138.1 | 94.1 | 137.0 | C7H7NO2 | [M + H]+ | Pyridine alkaloid | 535-83-1 | C01004 |
| 65 | Zarzissine | 136.1 | 119.0 | 135.1 | C5H5N5 | [M + H]+ | Guanidine alkaloid | 160568-14-9 | - |
| 66 | demethylcoclaurine-glucose | 434.2 | 272.1 | 433.2 | C22H27NO8 | [M + H]+ | Other alkaloids | - | - |
| 67 | m-Aminophenylacetylene | 118.1 | 91.1 | 117.1 | C8H7N | [M + H]+ | Alkyne | 54060-30-9 | - |
| 68 | naphthisoxazol A * | 188.1 | 118.1 | 187.1 | C11H9NO2 | [M + H]+ | Isoxazole | - | - |
| 69 | A23 | 221.1 | 130.1 | 220.1 | C11H12N2O3 | [M + H]+ | Tryptamine derivative | - | - |
| 70 | vasicinone | 203.1 | 130.1 | 202.1 | C11H10N2O2 | [M + H]+ | Quinazoline alkaloid | 486-64-6 | C10744 |
| 71 | A-hydroxyquinoline | 146.1 | 118.1 | 145.1 | C9H7NO | [M + H]+ | Quinoline alkaloid | - | - |
| Variables | Total Alkaloid | HRSA Assay | FRAP Capacity | DPPH Activity | DPPH Capacity |
|---|---|---|---|---|---|
| Total Alkaloids | 1 | ||||
| HRSA Assay | 0.99 * | 1 | |||
| FRAP capacity | 0.90 * | 0.92 * | 1 | ||
| DPPH activity | 0.97 * | 0.98 * | 0.97 * | 1 | |
| DPPH capacity | 0.96 * | 0.97 * | 0.98 * | 0.99 * | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, M.; Wang, H.; Song, K.; Liu, H.; Jiang, S.; Feng, J.; Jiang, S.; Rao, M.J.; Rauf, A.; Ikram, M.; et al. Integrated Transcriptomic and Metabolomic Provide Insights into Strawberry Bioactive Compounds and Antioxidant Activities Under Varying Ecological Conditions. Horticulturae 2025, 11, 533. https://doi.org/10.3390/horticulturae11050533
Duan M, Wang H, Song K, Liu H, Jiang S, Feng J, Jiang S, Rao MJ, Rauf A, Ikram M, et al. Integrated Transcriptomic and Metabolomic Provide Insights into Strawberry Bioactive Compounds and Antioxidant Activities Under Varying Ecological Conditions. Horticulturae. 2025; 11(5):533. https://doi.org/10.3390/horticulturae11050533
Chicago/Turabian StyleDuan, Mingzheng, Huaizheng Wang, Kangjian Song, Honggao Liu, Shu Jiang, Jieming Feng, Shiping Jiang, Muhammad Junaid Rao, Abdul Rauf, Muhammad Ikram, and et al. 2025. "Integrated Transcriptomic and Metabolomic Provide Insights into Strawberry Bioactive Compounds and Antioxidant Activities Under Varying Ecological Conditions" Horticulturae 11, no. 5: 533. https://doi.org/10.3390/horticulturae11050533
APA StyleDuan, M., Wang, H., Song, K., Liu, H., Jiang, S., Feng, J., Jiang, S., Rao, M. J., Rauf, A., Ikram, M., Khurram, M. F., Tahira, M., Yang, S., & Duan, X. (2025). Integrated Transcriptomic and Metabolomic Provide Insights into Strawberry Bioactive Compounds and Antioxidant Activities Under Varying Ecological Conditions. Horticulturae, 11(5), 533. https://doi.org/10.3390/horticulturae11050533