
Optimizing Storage and Regeneration of Clonal Propagules of Salix tetrasperma Through Double-Layered Encapsulation


Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Explant
2.2. Single-Layer Encapsulation
2.3. Double Layer Encapsulation
2.4. Growth Media and Culture Conditions
2.5. Rooting and Acclimatization
2.6. Effect of Low-Temperature Storage on Encapsulated and Non-Encapsulated Synthetic Seeds
2.7. Biochemical Studies
2.8. Superoxide Dismutase (SOD)
2.9. Catalase (CAT)
2.10. Estimation of Proline
2.11. SCoT Analysis
2.12. Statistical Analysis
3. Results and Discussion
3.1. Effect of Sodium Alginate and Calcium Chloride Concentrations on Bead Formation
3.2. Effect of Complexing Time on Bead Formation
3.3. Effect of Media Composition on Conversion of SLSs and DLSs into Plantlets
3.4. Low-Temperature Storage of Encapsulated and Non-Encapsulated Beads
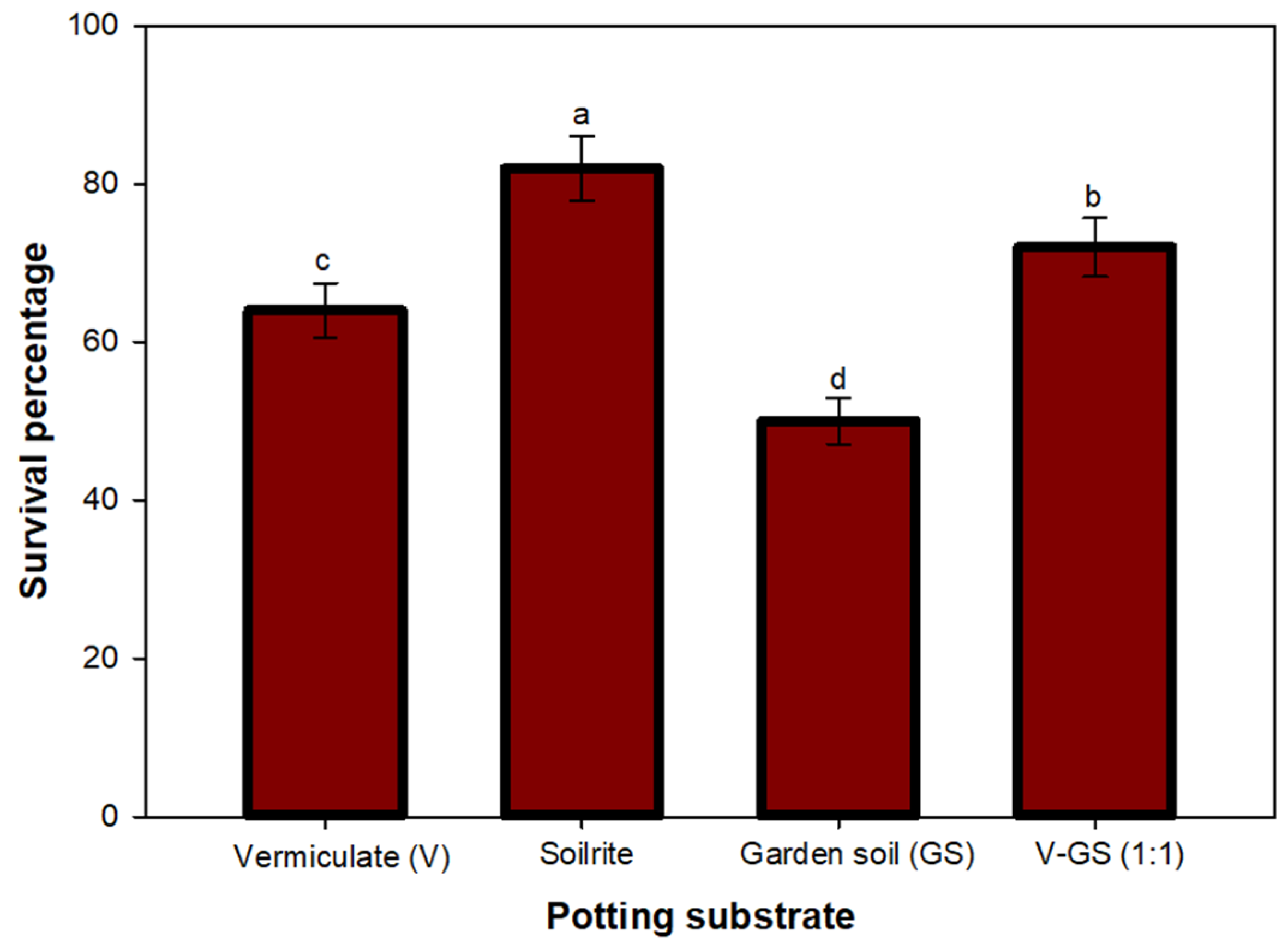
3.5. Acclimatization
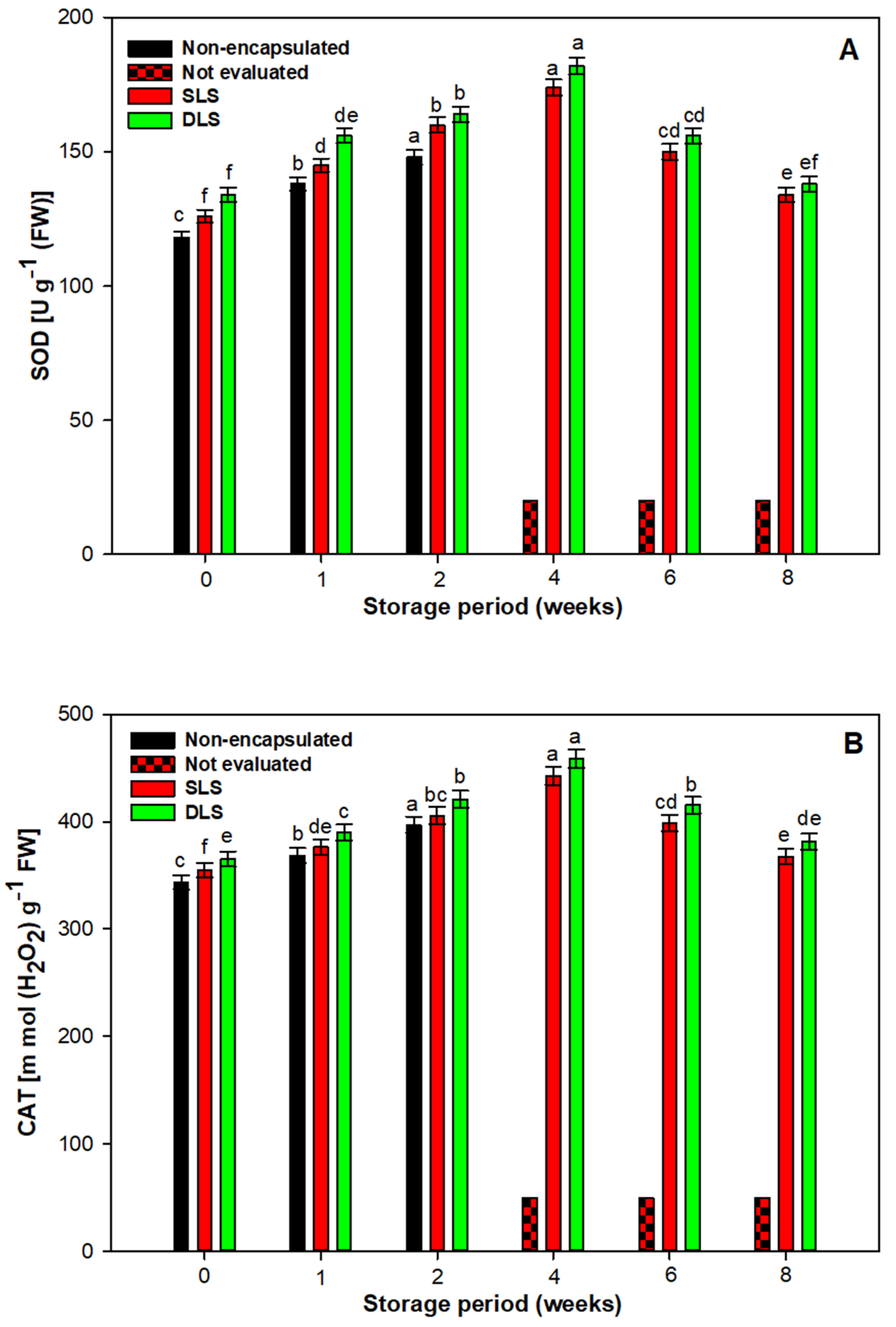
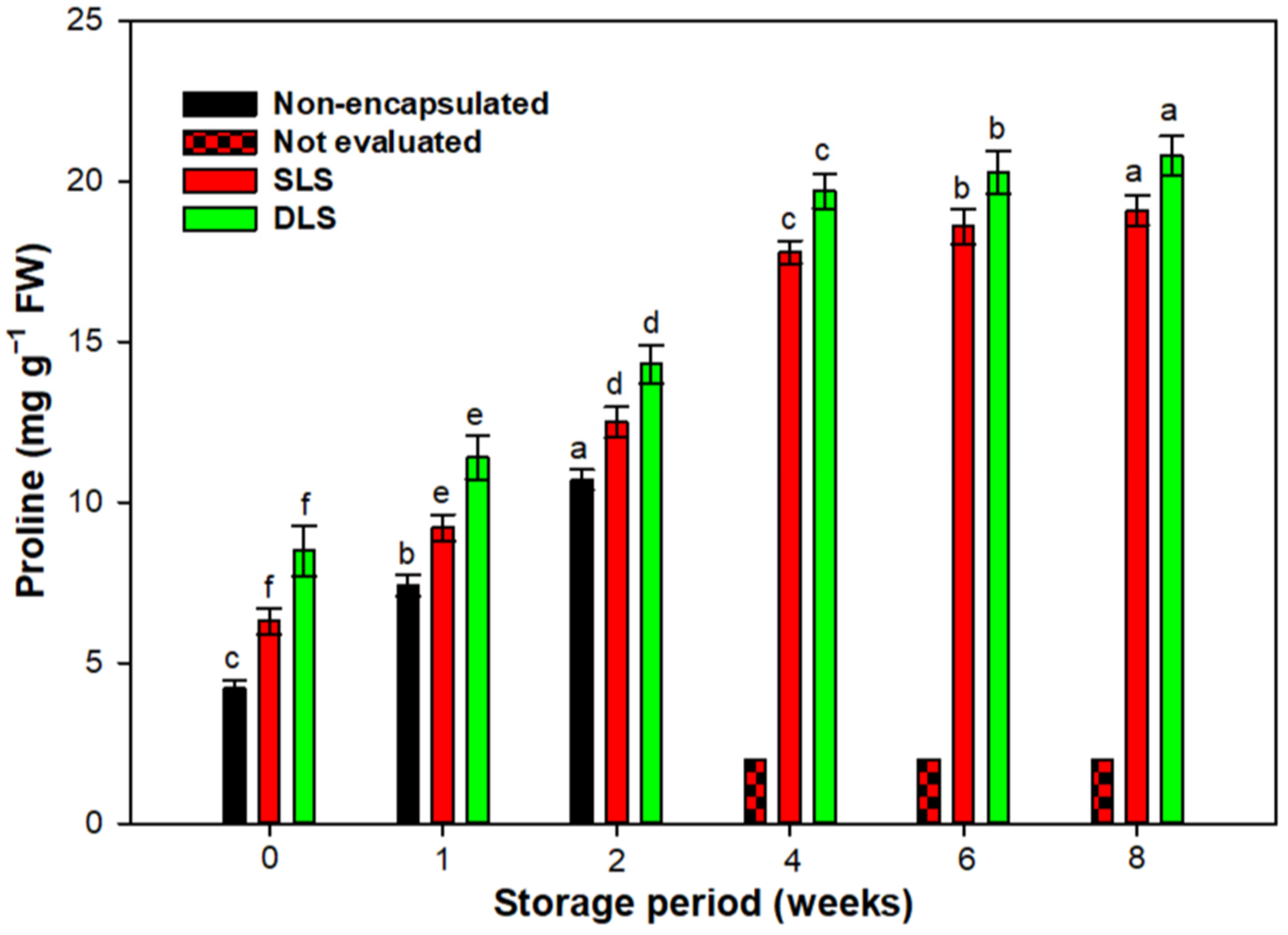
3.6. Biochemical Studies
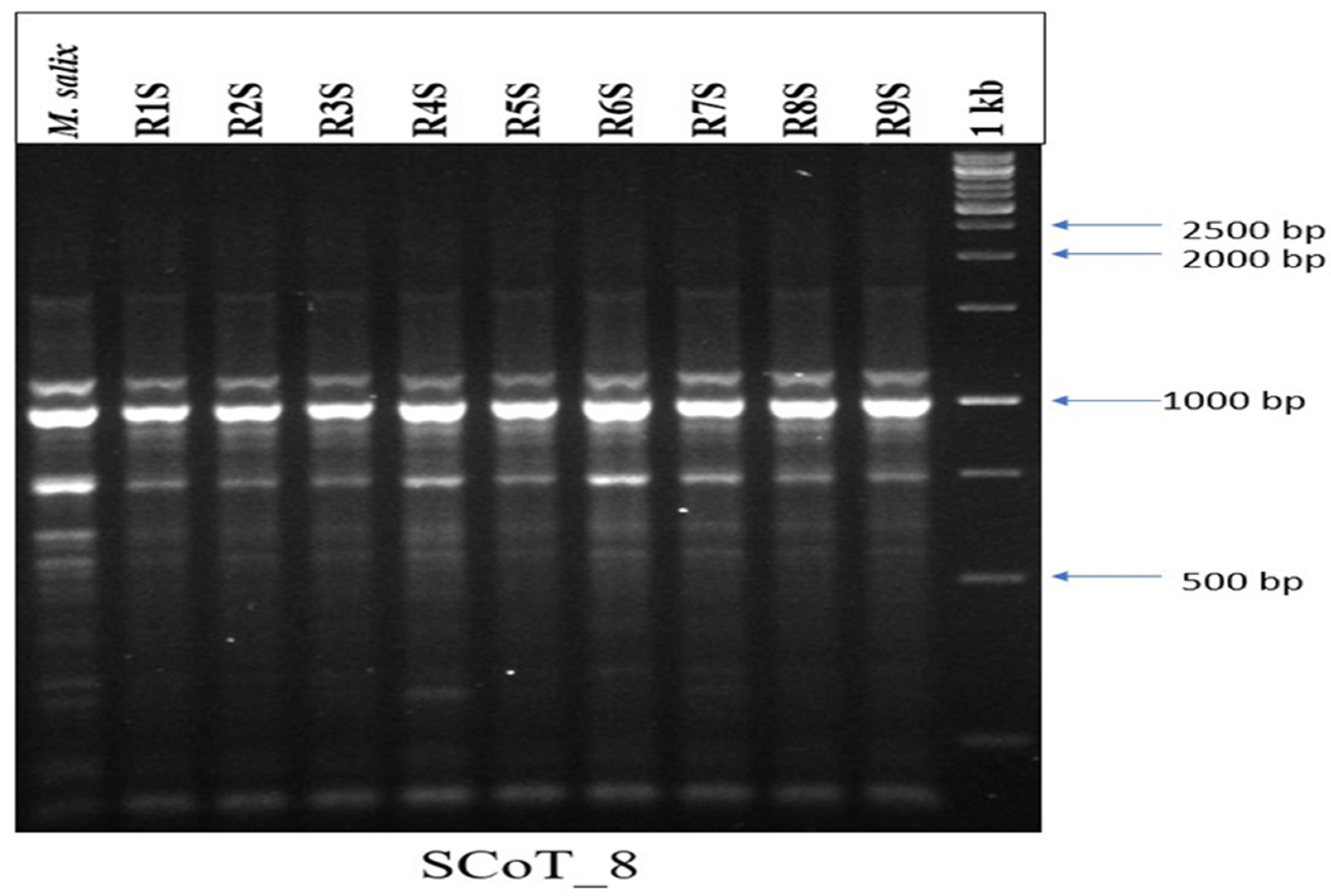
3.7. Evaluation of Genetic Stability Using SCoT Molecular Markers
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tienaho, J.; Reshamwala, D.; Sarjala, T.; Kilpeläinen, P.; Liimatainen, J.; Dou, J.; Viherä-Aarnio, A.; Linnakoski, R.; Marjomäki, V.; Jyske, T. Salix spp. Bark Hot Water Extracts Show Antiviral, Antibacterial, and Antioxidant Activities—The Bioactive Properties of 16 Clones. Front. Bioeng. Biotechnol. 2021, 9, 797939. [Google Scholar] [CrossRef] [PubMed]
- El-Shazly, A.; El-Sayed, A.; Fikrey, E. Bioactive Secondary Metabolites from Salix tetrasperma Roxb. Z. Fur Naturforschung. C 2012, 67, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Virupaksha, J.H.; Nadendla, R.R.; Kumar, S.M.; Kavya, S. Effect of Salix tetrasperma Roxburgh Leaf Extracts on Central Nervous System Activities. Res. J. Pharm. Biol. Chem. Sci. 2017, 7, 2060–2064. [Google Scholar]
- Prashith Kekuda, T.; Vinayaka, K.; Raghavendra, H. Ethnobotanical Uses, Phytochemistry, and Biological Activities of Salix tetrasperma Roxb. (Salicaceae)—A Review. J. Med. Plants 2017, 5, 201–206. [Google Scholar]
- Tawfeek, N.; Mahmoud, M.F.; Hamdan, D.I.; Sobeh, M.; Farrag, N.; Wink, M.; El-Shazly, A.M. Phytochemistry, Pharmacology, and Medicinal Uses of Plants of the Genus Salix: An Updated Review. Front. Pharmacol. 2021, 12, 593856. [Google Scholar] [CrossRef] [PubMed]
- Gligorić, E.; Igić, R.; Čonić, B.S.; Kladar, N.; Teofilović, B.; Grujić, N. Chemical Profiling and Biological Activities of “Green” Extracts of Willow Species (Salix L., Salicaceae): Experimental and Chemometric Approaches. Sustain. Chem. Pharm. 2023, 32, 100981. [Google Scholar] [CrossRef]
- Khan, M.I.; Anis, M. Modulation of In Vitro Morphogenesis in Nodal Segments of Salix tetrasperma Roxb. through the Use of TDZ, Different Media Types, and Culture Regimes. Agrofor. Syst. 2012, 86, 95–103. [Google Scholar] [CrossRef]
- Kordrostami, M.; Mafakheri, M.; Al-Khayri, J.M. Date Palm (Phoenix dactylifera L.) Genetic Improvement via Biotechnological Approaches. Tree Genet. Genomes 2022, 18, 26. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A. Production of Synthetic Seeds for Hybrid Cymbidium Using Protocorm-Like Bodies. J. Fruit Ornam. Plant Res. 2012, 20, 135–146. [Google Scholar] [CrossRef]
- Standardi, A.; Micheli, M. Encapsulation of In Vitro-Derived Explants: An Innovative Tool for Nurseries. Methods Mol. Biol. 2013, 11013, 397–418. [Google Scholar] [CrossRef] [PubMed]
- Nongdam, P. Development of Synthetic Seed Technology in Plants and Its Applications: A Review. Int. J. Curr. Sci. 2016, 19, 86–101. [Google Scholar]
- Javed, S.B.; Alatar, A.A.; Anis, M.; Faisal, M. Synthetic Seeds Production and Germination Studies for Short-Term Storage and Long-Distance Transport of Erythrina variegata L.: A Multipurpose Tree Legume. Ind. Crops Prod. 2017, 105, 41–46. [Google Scholar] [CrossRef]
- Khan, S.A.; Verma, P.; Rahman, L.U.; Parasharami, V. Synthetic Seed-Mediated Synchronized Multiplication Under In Vitro Conditions: An Efficient Technique for Conservation, Multiplication, and Storage of Medicinal Plants. In Medicinal and Aromatic Plants; Aftab, T., Hakeem, K.R., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Abbas, M.K.; Alhasan, A.S. Production of Synthetic Seeds in Vegetable Crops: A Review. IOP Conf. Series Earth Environ. Sci. 2022, 1060, 012099. [Google Scholar] [CrossRef]
- Ahmad, A.; Ansari, F.A.; Anis, M.; Khan, A.S. Micropropagation of Pterocarpus marsupium Roxb. Through Synthetic Seeds and Its Novel Antibiofilm Activities Against ESKAPE Pathogens. Ind. Crops Prod. 2023, 198, 116681. [Google Scholar] [CrossRef]
- Gantait, S.; Kundu, S.; Ali, N.; Sahu, N.C. Synthetic Seed Production of Medicinal Plants: A Review on Influence of Explants, Encapsulation Agent, and Matrix. Acta Physiol. Plant. 2015, 37, 98. [Google Scholar] [CrossRef]
- Prakash, A.V.; Nair, D.S.; Alex, S.; Soni, K.B.; Viji, M.M.; Reghunath, B.R. Calcium Alginate Encapsulated Synthetic Seed Production in Plumbago rosea L. for Germplasm Exchange and Distribution. Physiol. Mol. Biol. Plants 2018, 24, 963–971. [Google Scholar] [CrossRef]
- Rodrigues, V.; Kumar, A.; Gokul, S.; Verma, R.S.; Rahman, L.U.; Sundaresan, V. Micropropagation, Encapsulation, and Conservation of Decalepis salicifolia, a Vanillin Isomer Containing Medicinal and Aromatic Plant. In Vitro Cell. Dev. Biol. Plant 2020, 56, 526–537. [Google Scholar] [CrossRef]
- Khanam, M.N.; Javed, S.B.; Ahmad, N.; Anis, M. Encapsulation of Nodal Segments of Allamanda cathartica for Short-Term Storage and Germplasm Exchange. Plant Cell Tissue Organ Cult. 2021, 145, 435–443. [Google Scholar] [CrossRef]
- Asadi, R.; Abdollahi, M.R.; Moosavi, S.S.; Mirzaie-Asl, A. Alginate Encapsulation of Micro-Cuttings in Endangered Satureja khuzistanica Species: A Promising Method for Obtaining Genetically Stable Plants with High Rosmarinic Acid Content. Plant Cell Tissue Organ Cult. 2022, 151, 307–320. [Google Scholar] [CrossRef]
- Qahtan, A.A.; Alatar, A.A.; Faisal, M. In Vitro Regeneration, Phytochemical Profiling, and Antioxidant Activity in Ruta chalepensis Plants Established from Alginate Encapsulated Synthetic Seeds. S. Afr. J. Bot. 2023, 161, 575–585. [Google Scholar] [CrossRef]
- Rai, M.K.; Asthana, P.; Singh, S.K.; Jaiswal, V.S.; Jaiswal, U. The Encapsulation Technology in Fruit Plants—A Review. Biotechnol. Adv. 2009, 27, 671–679. [Google Scholar] [CrossRef]
- Sharma, S.; Shahzad, A.; Parveen, S. Synthetic Seed Technology for Short-Term Storage and Propagation of Vitex Species: A Review. In Vitro Cell. Dev. Biol. Plant 2013, 49, 507–515. [Google Scholar] [CrossRef]
- Xu, Y.; Yan, X.; Zheng, H.; Li, J.; Wu, X.; Xu, J.; Zhen, Z.; Du, C. The Application of Encapsulation Technology in the Food Industry: Classifications, Recent Advances, and Perspectives. Food Chem. X 2024, 13, 101240. [Google Scholar] [CrossRef] [PubMed]
- Szopa, D.; Mielczarek, M.; Skrzypczak, D.; Izydorczyk, G.; Mikula, K.; Chojnacka, K.; Witek-Krowiak, A. Encapsulation Efficiency and Survival of Plant Growth-Promoting Microorganisms in an Alginate-Based Matrix—A Systematic Review and Protocol for a Practical Approach. Ind. Crops Prod. 2022, 181, 114846. [Google Scholar] [CrossRef]
- Ma, Y. Seed Coating with Beneficial Microorganisms for Precision Agriculture. Biotechnol. Adv. 2019, 37, 107423. [Google Scholar] [CrossRef]
- Hu, D.; Ma, G.; Wang, Q.; Yao, J.; Wang, Y.; Pritchard, H.W.; Wang, X. Spatial and Temporal Nature of Reactive Oxygen Species Production and Programmed Cell Death in Elm (Ulmus pumila L.) Seeds during Controlled Deterioration. Plant Cell Environ. 2012, 35, 2045–2059. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Poór, P.; Ördög, A.; Czékus, Z.; Borbély, P.; Takács, Z.; Kovács, J.; Tari, I. Regulation of the Key Antioxidant Enzymes by Developmental Processes and Environmental Stresses in the Dark. Biol. Plant. 2018, 62, 201–210. [Google Scholar] [CrossRef]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic Stress in Crop Species: Improving Tolerance by Applying Plant Metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Dhinsa, P.S.; Plumb-Dhinsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Aebi, H. Catalase In Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, J.D. Rapid Determination of Proline for Water Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of Plant DNA from Fresh Tissue. Focus 1990, 12, 39–40. [Google Scholar]
- Collard, B.C.Y.; Mackill, D.J. Start Codon Targeted (SCoT) Polymorphism: A Simple, Novel DNA Marker Technique for Generating Gene-Targeted Markers in Plants. Plant Mol. Biol. Rep. 2009, 27, 86–93. [Google Scholar] [CrossRef]
- Behera, S.; Rout, K.K.; Panda, P.C.; Naik, S.K. Production of Non-Embryogenic Synthetic Seeds for Propagation and Germplasm Transfer of Hedychium coronarium J Koenig. J. Appl. Res. Med. Aromat. Plants 2020, 19, 100271. [Google Scholar] [CrossRef]
- Siddique, I.; Bukhari, N.A.W. Synthetic Seed Production by Encapsulating Nodal Segment of Capparis decidua (Forsk.), In Vitro Regrowth of Plantlets and Their Physio-Biochemical Studies. Agrofor. Syst. 2018, 92, 1711–1719. [Google Scholar] [CrossRef]
- Alatar, A.A.; Ahmad, N.; Javed, S.B.; Abdel-Salam, E.M.; Basahi, R.; Faisal, M. Two-Way Germination System of Encapsulated Clonal Propagules of Vitex trifolia L.: An Important Medicinal Plant. J. Hortic. Sci. Biotechnol. 2017, 92, 175–182. [Google Scholar] [CrossRef]
- Hatzilazarou, S.; Kostas, S.; Nendou, T.; Economou, A. Conservation, Regeneration, and Genetic Stability of Regenerants from Alginate-Encapsulated Shoot Explants of Gardenia jasminoides Ellis. Polymers 2021, 13, 1666. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Tripathi, M.K.; Tiwari, S.; Tripathi, N.; Tejovathi, G.; Ahuja, A.; Tiwari, S. Production of Synthetic Seed from Encapsulated Nodal Segments of Giloe (Tinospora cordifolia Willd.). In Research Developments in Science and Technology; BP International: Hooghly, West Bengal, India, 2022; Volume 2, pp. 156–170. [Google Scholar]
- Kumar, G.K.; Thomas, T.D. High-Frequency Somatic Embryogenesis and Synthetic Seed Production in Clitoria ternatea Linn. Plant Cell Tissue Organ Cult. 2012, 110, 141–151. [Google Scholar] [CrossRef]
- Mahdavi, Z.; Daylami, S.D.; Fadavi, A.; Vahdati, K. Artificial Seed Production of Phalaenopsis Orchid: Effect of Encapsulation Materials, Temperature, Light Spectra, and Storage Period. Plant Cell Tissue Organ Cult. 2023, 155, 797–808. [Google Scholar] [CrossRef]
- Qahtan, A.A.; Abdel-Salam, E.M.; Alatar, A.A.; Wang, Q.C.; Faisal, M. An Introduction to Synthetic Seeds: Production, Techniques, and Applications. In Synthetic Seeds: Germplasm Regeneration, Preservation and Prospects; Springer: Cham, Switzerland, 2019; pp. 1–20. [Google Scholar]
- Gantait, S.; Mitra, M. Applications of Synthetic Seed Technology for Propagation, Storage, and Conservation of Orchid Germplasms. In Synthetic Seeds: Germplasm Regeneration, Preservation and Prospects; Springer: Cham, Switzerland, 2019; pp. 301–321. [Google Scholar]
- Ekinci, H.; Çiftçi, Y.Ö.; Nadarajan, J. Medium- and Long-Term Conservation of Ornamental Plants Using Synthetic Seed Technology. In Synthetic Seeds: Germplasm Regeneration, Preservation and Prospects; Springer: Cham, Switzerland, 2019; pp. 259–281. [Google Scholar]
- Ghanbarali, S.; Abdollahi, M.R.; Zolnorian, H.; Moosavi, S.S.; Seguí-Simarro, J.M. Optimization of the Conditions for Production of Synthetic Seeds by Encapsulation of Axillary Buds Derived from Minituber Sprouts in Potato (Solanum tuberosum). Plant Cell Tiss. Organ Cult. 2016, 126, 449–458. [Google Scholar] [CrossRef]
- de Andrade, S.A.L.; de Oliveira, V.H.; Mazzafera, P. Metabolomics of Nutrient-Deprived Forest Trees. In Monitoring Forest Damage with Metabolomics Methods; Springer: Cham, Switzerland, 2024; pp. 235–265. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Kareem, F.; El-Mahrouk, M.E.; Fuller, M.P. Artificial Seeds: Principle, Aspects, and Applications. Agronomy 2017, 7, 71. [Google Scholar] [CrossRef]
- Raspor, M.; Motyka, V.; Kaleri, A.R.; Ninković, S.; Tubić, L.; Cingel, A.; Ćosić, T. Integrating the Roles for Cytokinin and Auxin in de Novo Shoot Organogenesis: From Hormone Uptake to Signaling Outputs. Int. J. Mol. Sci. 2021, 22, 8554. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, E.; Kinoshita, I.; Ishii, K.; Shigenaga, H.; Ohra, K.; Saito, A. Alginate-Encapsulation Technology for the Propagation of the Tropical Forest Trees: Cedrela odorata L., Guazuma crinita Mart., and Jacaranda mimosaefolia D. Don. Silvae Genet. 1997, 46, 17–23. [Google Scholar]
- Jayaprakash, C.M.; Divakaran, M.; Madhusoodanan, P.V. Role of Activated Charcoal in the Micropropagation and Conservation of Celastrus paniculatus Willd. Med. Plants-Int. J. Phytomedicines Relat. Ind. 2022, 14, 448–455. [Google Scholar] [CrossRef]
- Balaji, R.; Sriram, S.; Vigneshwaran, K.; Bharathi, A. Micropropagation of Acmella calva (DC.) RK Jansen (Asteraceae) via Encapsulation of In Vitro Nodal Explants. Plant Cell Tissue Organ Cult. 2016, 127, 1–9. [Google Scholar] [CrossRef]
- Largia, M.J.V.; Pandian, S.K.; Ramesh, M. Genetic Fidelity Assessment of Encapsulated In Vitro Tissues of Bacopa monnieri after Six Months of Storage Using ISSR and RAPD Markers. Turk. J. Bot. 2013, 37, 1008–1017. [Google Scholar] [CrossRef]
- Fatima, N.; Ahmad, N.; Anis, M.; Ahmad, I. An Improved In Vitro Encapsulation Protocol, Biochemical Analysis, and Genetic Integrity Using DNA-Based Molecular Markers in Regenerated Plants of Withania somnifera L. Ind. Crops Prod. 2013, 50, 468–477. [Google Scholar] [CrossRef]
- Naz, R.; Anis, M.; Alatar, A.A.; Ahmad, A.; Naaz, A. Nutrient Alginate Encapsulation of Nodal Segments of Althaea officinalis L., for Short-Term Conservation and Germplasm Exchange. Plant Biosyst. 2018, 152, 1256–1262. [Google Scholar] [CrossRef]
- Benson, E.E.; Harding, K.; Ryan, M.; Petrenko, A.; Petrenko, Y.; Fuller, A. Alginate Encapsulation to Enhance Biopreservation Scope and Success: A Multidisciplinary Review of Current Ideas and Applications in Cryopreservation and Non-Freezing Storage. CryoLetters 2018, 39, 14–38. [Google Scholar]
- Kocak, M.; Sevindik, B.; Izgu, T.; Tutuncu, M.; Mendi, Y.Y. Synthetic Seed Production of Flower Bulbs. In Synthetic Seeds: Germplasm Regeneration, Preservation and Prospects; Springer: Cham, Switzerland, 2019; pp. 283–299. [Google Scholar]
- Javed, S.B.; Anis, M. Cobalt-Induced Augmentation of In Vitro Morphogenic Potential in Erythrina variegata L.: A Multipurpose Tree Legume. Plant Cell Tissue Organ Cult. 2015, 120, 463–474. [Google Scholar] [CrossRef]
- Tian, S.; Qin, G.; Li, B. Reactive Oxygen Species Involved in Regulating Fruit Senescence and Fungal Pathogenicity. Plant Mol. Biol. 2013, 82, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.R.; Anis, M. Changes in Activity of Antioxidant Enzymes and Photosynthetic Machinery during Acclimatization of Micropropagated Cassia alata L. Plantlets. In Vitro Cell. Dev. Biol. Plant 2014, 50, 601–609. [Google Scholar] [CrossRef]
- Khan, T.A.; Fariduddin, Q.; Yusuf, M. Lycopersicon esculentum under Low-Temperature Stress: An Approach toward Enhanced Antioxidants and Yield. Environ. Sci. Pollut. Res. 2015, 22, 14178–14188. [Google Scholar] [CrossRef] [PubMed]
- Faisal, M.; Alatar, A.A. Synthetic Seeds; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Akhtar, G.; Jaskani, M.J.; Sajjad, Y.; Akram, A. Effect of Antioxidants, Amino Acids, and Plant Growth Regulators on In Vitro Propagation of Rosa centifolia. Iran. J. Biotechnol. 2016, 14, 51. [Google Scholar] [CrossRef]
- Javed, S.B.; Alatar, A.A.; Anis, M.; El-Sheikh, M.A. In Vitro Regeneration of Coral Tree from Three Different Explants Using Thidiazuron. HortTechnology 2019, 29, 946–951. [Google Scholar] [CrossRef]
- Kader, A.; Sinha, S.N.; Ghosh, P. A Strategy for Development of Genetically Stable Synthetic Seeds of Azadirachta indica A. Juss. (Neem) Suitable for In Vitro Storage. Plant Cell Tissue Organ Cult. 2022, 151, 47–58. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sodium Alginate (%w/v) | Conversion Response (%) into Synseeds | Remark |
|---|---|---|
| 0 | NA | No bead formation |
| 1 | 16.35 ± 0.31 e | Fragile |
| 2 | 33.84 ± 0.64 c | Soft with loose texture |
| 3 | 51.6 ± 0.98 a | Soft with good texture |
| 4 | 37.23 ± 0.70 b | Hard |
| 5 | 26.41 ± 0.50 d | Hard |
| CaCl2 (mM) | Percentage Response | Remark |
|---|---|---|
| 0 | NA | No bead formation |
| 50 | 17.52 ± 0.33 e | Fragile and difficult to handle |
| 75 | 34.25 ± 0.65 c | Soft with loose texture |
| 100 | 51.6 ± 0.98 a | Soft with good texture |
| 125 | 42.7 ± 0.81 b | Hard |
| 150 | 19.36 ± 0.37 d | Hard |
| Time (min) | Response into Synseed | Remark |
|---|---|---|
| 10 | 19.32 ± 0.37 d | Fragile beads difficult to handle |
| 20 | 24.51 ± 0.47 c | Soft with loose texture |
| 30 | 51.6 ± 0.98 a | Soft with good texture |
| 40 | 31.3 ± 0.60 b | Hard |
| 50 | 12.8 ± 0.24 e | Hard |
| Media Used in Germination | Single Layered | Double Layered | ||
|---|---|---|---|---|
| Concentration (µM) | Conversion Response into Plantlets | Mean Number of Shoots Per Bead | Conversion Response into Plantlets | Mean Number of Shoots Per Bead |
| ½ MS | 38.2 ± 0.73 e | 1.9 ± 0.03 e | 46.3 ± 0.88 f | 2.1 ± 0.04 e |
| MS | 46.9 ± 0.89 d | 2.5 ± 0.05 d | 52.6 ± 1.01 d | 2.8 ± 0.05 d |
| MS + BA (5.0) + NAA (0.5) | 69.8 ± 1.33 b | 3.5 ± 0.07 b | 75.4 ± 1.44 b | 4.6 ± 0.09 b |
| MS + mT (5.0) + NAA (0.5) | 87.3 ± 1.66 a | 6.4 ± 0.12 a | 93.6 ± 1.78 a | 7.2 ± 0.13 a |
| MS + Kn (5.0) + NAA (0.5) | 51.5 ± 0.98 c | 2.8 ± 0.05 c | 58.5 ± 1.11 c | 3.5 ± 0.06 c |
| Storage Time (Weeks) | Conversion Response of Single-Layered Encapsulated Nodal Segment | Conversion Response of Double-Layered Encapsulated Nodal Segment | Conversion Response of Non-Encapsulated Nodal Segment |
|---|---|---|---|
| 0 | 87.3 ± 1.66 a | 93.6 ± 1.78 a | 96.0 ± 1.98 a |
| 1 | 76.5 ± 1.46 b | 84.5 ± 1.61 b | 67.3 ± 1.28 b |
| 2 | 64.7 ± 1.23 c | 76.2 ± 1.45 c | 53.6 ± 1.02 c |
| 4 | 58.6 ± 1.11 d | 66.7 ± 1.27 d | 33.7 ± 0.64 d |
| 6 | 38.8 ± 0.73 e | 44.3 ± 0.84 e | 19.4 ± 0.36 e |
| 8 | 18.2 ± 0.35 f | 24.4 ± 0.46 f | 9.2 ± 0.17 f |
| S. No. | Primer | Sequences (5′-3′) | Total Bands Screened |
|---|---|---|---|
| 1 | SCoT5 | CAACAATGGCTACCACGA | 54 |
| 2 | SCoT8 | CAACAATGGCTACCACGT | 81 |
| 3 | SCoT10 | CAACAATGGCTACCAGCC | 63 |
| 4 | SCoT13 | ACGACATGGCGACCATCG | 63 |
| 5 | SCoT18 | ACCATGGCTACCACCGCC | 72 |
| 6 | SCoT20 | ACCATGGCTACCACCGCG | 63 |
| 7 | SCoT29 | CCATGGCTACCACCGGCC | 63 |
| 8 | SCoT30 | CCATGGCTACCACCGGCG | 63 |
| 9 | SCoT33 | CCATGGCTACCACCGCAG | 72 |
| 10 | SCoT36 | GCAACAATGGCTACCACC | 72 |
| Total | 666 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reshi, Z.A.; Ahmad, W.; Husain, F.M.; Khanam, M.N.; Javed, S.B. Optimizing Storage and Regeneration of Clonal Propagules of Salix tetrasperma Through Double-Layered Encapsulation. Horticulturae 2025, 11, 486. https://doi.org/10.3390/horticulturae11050486
Reshi ZA, Ahmad W, Husain FM, Khanam MN, Javed SB. Optimizing Storage and Regeneration of Clonal Propagules of Salix tetrasperma Through Double-Layered Encapsulation. Horticulturae. 2025; 11(5):486. https://doi.org/10.3390/horticulturae11050486
Chicago/Turabian StyleReshi, Zubair Altaf, Waquar Ahmad, Fohad Mabood Husain, Mehrun Nisha Khanam, and Saad Bin Javed. 2025. "Optimizing Storage and Regeneration of Clonal Propagules of Salix tetrasperma Through Double-Layered Encapsulation" Horticulturae 11, no. 5: 486. https://doi.org/10.3390/horticulturae11050486
APA StyleReshi, Z. A., Ahmad, W., Husain, F. M., Khanam, M. N., & Javed, S. B. (2025). Optimizing Storage and Regeneration of Clonal Propagules of Salix tetrasperma Through Double-Layered Encapsulation. Horticulturae, 11(5), 486. https://doi.org/10.3390/horticulturae11050486