
Impact of Temperature on Growth, Photosynthetic Efficiency, Yield, and Functional Components of Bud-Leaves and Flowers in Edible Chrysanthemum (Chrysanthemum morifolium Ramat)

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Temperature Treatments and Measurement
2.1.1. Experimental Site and Treatments
2.1.2. Measurement of Parameters
Growth and Development
Yield and Biomass
Photosynthetic Parameters
2.1.3. Preparation of Extraction Solutions
2.2. Analysis of Functional Components
2.2.1. Polyphenols
2.2.2. Free Amino Acids
2.2.3. Flavonoids
2.2.4. Affeoylquinic Acid (CQAs)
2.2.5. Luteolin
2.2.6. Lutein
2.3. Data Analysis
3. Results
3.1. Effects of Temperature on the Growth of Taiwan Hangju No. 1
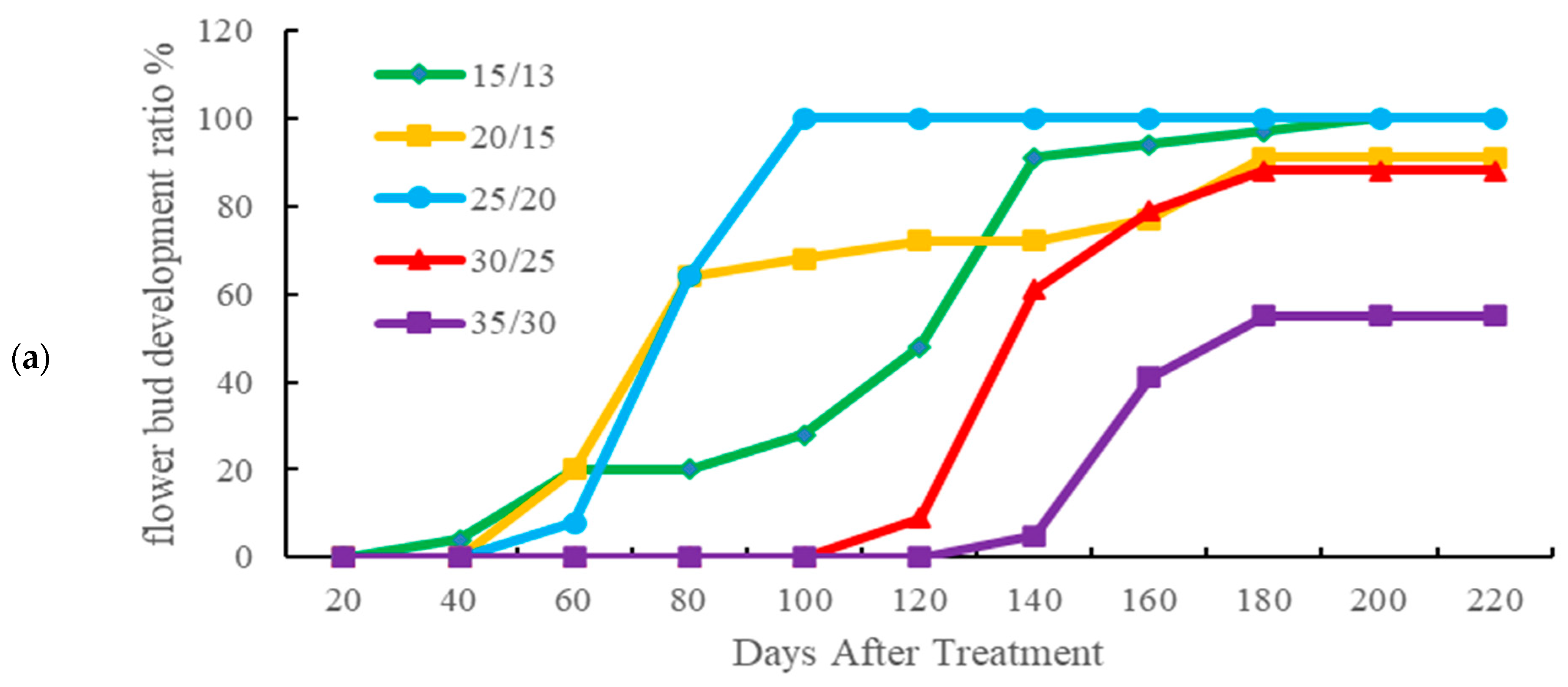
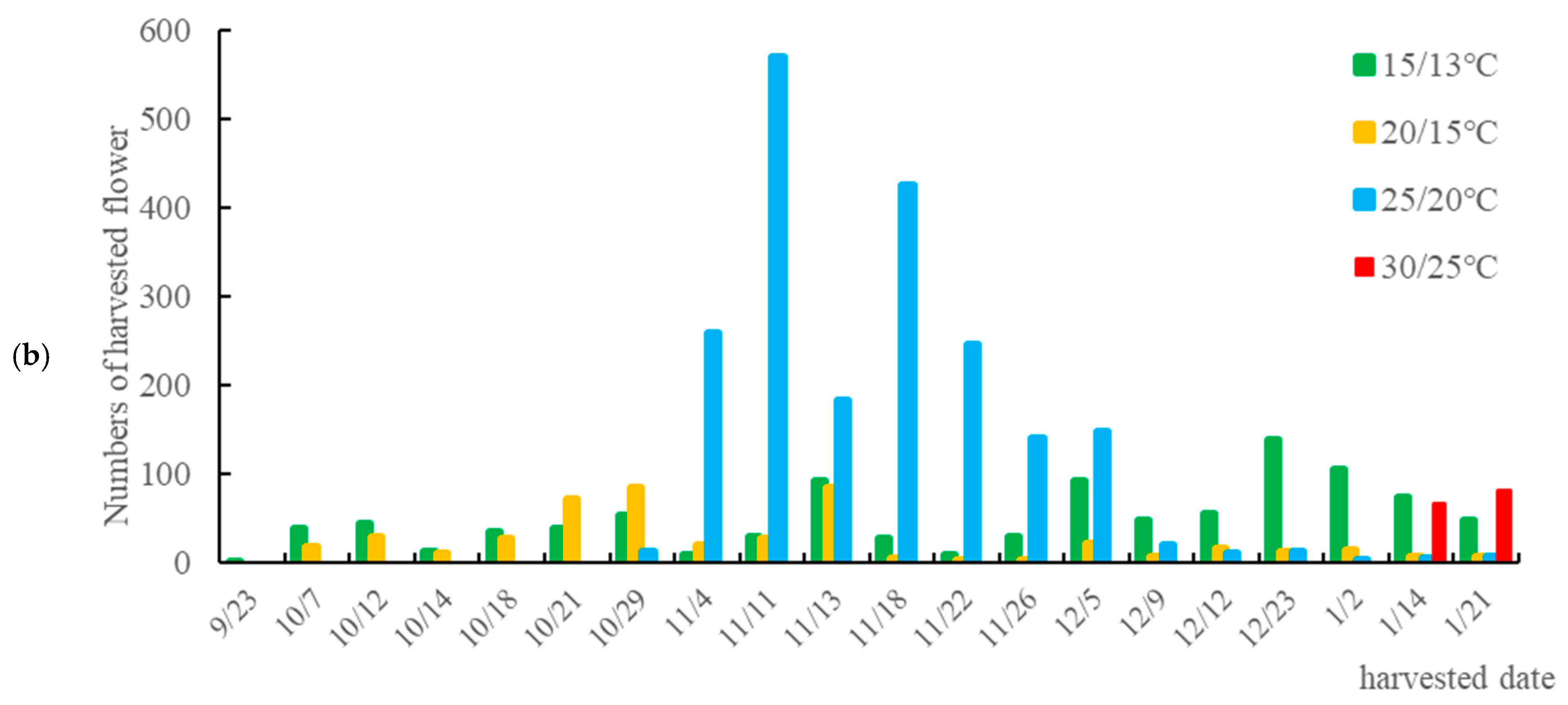
3.2. Effects of Temperature on the Reproductive Growth of Taiwan Hangju No. 1
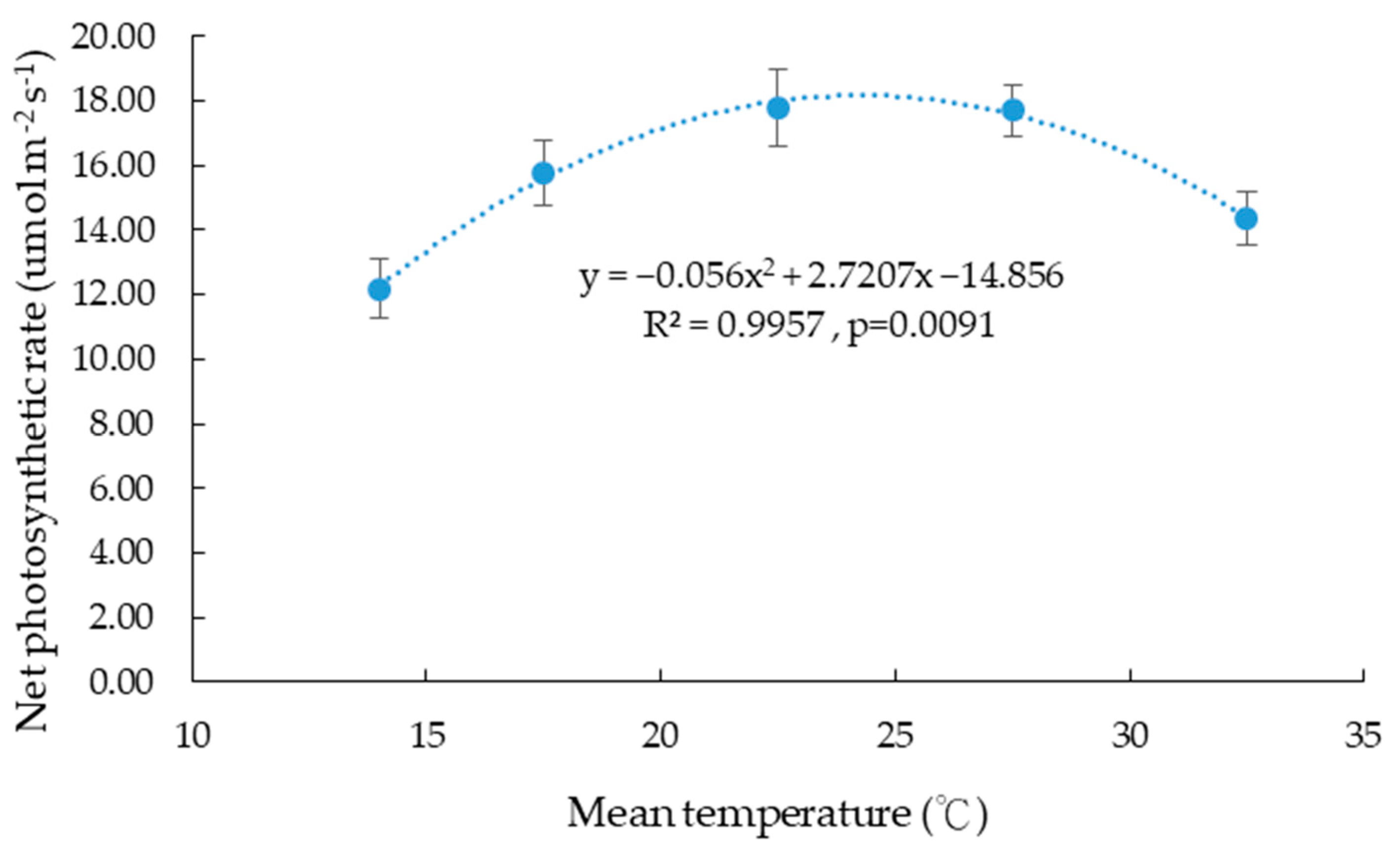
3.3. Effects of Temperature on Photosynthesis
3.4. Concentration and Content of Functional Compounds in Chrysanthemum morifolium Bud-Leaves
3.4.1. The Concentration of Functional Compounds in Bud-Leaves
3.4.2. The Content of Functional Compounds in Bud-Leaves
3.5. Concentration and Content of Functional Compounds in Chrysanthemum morifolium ‘Taiwan Hangju No. 1’ Flowers
3.5.1. The Concentration of Functional Compounds in Flowers
3.5.2. The Content of Functional Compounds in Flowers
4. Discussion
4.1. Effects of Temperature on the Vegetative Growth of Taiwan Hangju No. 1
4.2. Effects of Temperature on the Flowering of Taiwan Hangju No. 1
4.3. Effects of Temperature on Photosynthesis of Taiwan Hangju No. 1
4.4. Effects of Temperature on the Concentration and Content of Functional Components in Chrysanthemum morifolium
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, H.H.; Wu, M.; Wei, W.G.; Ren, W.K.; Zheng, Z.A. Chrysanthemum morifolium Ramat. as a traditional tea material: Unraveling the influence of kill-green process on drying characteristics, phytochemical compounds, and volatile profile. Food Res. Int. 2025, 200, 115478. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.F.; Cheng, R.A.; Li, Y.; Jiang, D.L.; Zhao, H.S.; Wu, X.F.; Shu, Y.C.; Lu, T.L.; Jin, C.S.; Wu, D.L.; et al. Effects of stir-frying on chemical profile, sensory quality and antioxidant activity of Chrysanthemi Flos: A metabolomics and sensory study. Food Res. Int. 2025, 200, 115391. [Google Scholar] [CrossRef] [PubMed]
- Allied Market Research. 2023. Chrysanthemum Tea Market. Available online: https://www.alliedmarketresearch.com/chrysanthemum-tea-market-A110757 (accessed on 7 April 2025).
- KBV Research. 2023. Global Chrysanthemum Tea Market Report. Available online: https://www.kbvresearch.com/chrysanthemum-tea-market/ (accessed on 7 April 2025).
- Li, J.M.; Li, H.Q.; Hu, S.X. Investigation on types of Chrysanthemum morifolium. China Mod. Med. 2016, 6, 93–96. [Google Scholar]
- Zhang, J.; Qian, D.W.; Li, Y.B.; Yin, Z.Q. Chemical Constituents from Chrysanthemum morifolium Ramat. Nat. Prod. Res. Dev. 2006, 1, 71–73. [Google Scholar]
- Chang, W.Y. Studies on Health Components of Chrysanthemum morifolium. Master’s Thesis, Department of Food Science, Tunghai University, Taichung, Taiwan, 2007. [Google Scholar]
- Wang, T.; Shen, X.G.; Guo, Q.S.; Zhou, J.S.; Mao, P.F.; Shen, Z.G. Comparison of major bioactive components from leaves of Chrysanthemum morifolium. China J. Chin. Mater. Medica 2015, 40, 1670–1674. [Google Scholar]
- Wang, C.S. Chemical Composition of Chrysanthemum morifolium Ramat Leaves and Eye-Care Product Development. Master’s Thesis, Graduate Institute of Food Science, Yuanpei University of Medical Technology, Hsinchu, Taiwan, 2018. [Google Scholar]
- Zhu, L.; Guo, J.M.; Yang, N.Y.; Qian, D.W.; Nie, H.; Su, S.L.; Ouyang, Z.; Duan, J.A. Distribution and dynamic changes of chemical constituentes from non-medicial parts of Chrysanthemum morifolium. Chin. Tradit. Herb. Drugs. 2014, 45, 425–431. [Google Scholar]
- Liu, C.F.; Guo, C.C. Taiwan Hang Ju No. 1 (Snow) Flower Tea and Leaf Tea health ingredients. Tea Newsl. 2017, 100, 11–14. [Google Scholar]
- Liu, C.F. Chrysanthemum morifolium Ramat Leaf Tea Processing Technology. Taiwan. Agri. Techno Mart. 2015, 124. Available online: https://tatm.moa.gov.tw/Home/EPaperNewsTempShowHistory.aspx?type=1&typecode=2&etmno=434 (accessed on 24 November 2024).
- Zhang, Q.H.; Zhang, L. Research Advance in Chemical Composition and Pharmacological Action of Chrysanthemum morifolium. Food Drug. 2007, 2, 60–63. [Google Scholar]
- Lo, Y.F.; Chang, D.L.; Chen, L.G.; Wang, C.C. Studies on the Antibiotic Activity and Bio-Activity of Anti-Inflammation of Several Endemic Medicinal Plants. Seed Nurs. 2009, 4, 13–26. [Google Scholar]
- Qu, L.; Wang, T.; Dong, Y.Z.; Zhang, J.H.; Zhang, Y. Research progress on chemical constituents of Chrysanthemum morifoliuin and their pharmacologic activities. Drug Eval. Res. 2015, 1, 98–104. [Google Scholar]
- Zheng, C.; Dong, Q.; Chen, H.; Cong, Q.; Ding, K. Structural characterization of a polysaccharide from Chrysanthemum morifolium flowers and its antioxidant activity. Carbohydr. Polym. 2015, 130, 113–121. [Google Scholar] [CrossRef]
- Chen, D.Y. CMFE (Chrysanthemum morifolium Flower Extract) Prevents Retinal Ganglia Cell and Ischemic Damages in an AOH (Acute Ocular Hypertension) Glaucoma Mouse Model. Master’s Thesis, Department of Mdeical Laboratory and Biotechnology, Chung Shan Medical University, Taichung, Taiwan, 2017. [Google Scholar]
- Wang, D.S.; Huang, Y.M.; Shi, Y. Research Progress on Chemical Constituent and Pharmacological Action of Chrysanthemum. J. Anhui Agric. Sci. 2018, 23, 9–17. [Google Scholar]
- Ai, Y.T. Effect of Chrysanthemum morifolium Ramat. Leaf Extract on Dry Eye. Master’s Thesis, Department of Medical Laboratory and Biotechnology, Chung Shan Medical University, Taichung, Taiwan, 2023. [Google Scholar]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Response to high temperature stress. J. Photochem. Photobiol. B Biol. 2014, 137, 116–126. [Google Scholar] [CrossRef]
- Scafaro, A.P.; Fan, Y.; Posch, B.C.; Garcia, A.; Coast, O.; Atkin, O.K. Responses of leaf respiration to heatwaves. Plant Cell Environ. 2021, 7, 2090–2101. [Google Scholar] [CrossRef]
- Zobayed, S.M.A.; Afreen, F.; Kozai, T. Temperature stress can alter the photosynthetic efficiency and secondary metabolite concentrations in St. John’s Wort. Plant Physiol. Biochem. 2005, 10, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.H.; Jiao, Y.; Zhang, X.K.; Zu, Y.G.; Gao, Y.; Sun, Y.F.; Yang, L.; Zhao, X.J. Effect of high temperature on life cycle forms and physiological metabolisms of Catharanthus roseus. Acta Ecol. Sin. 2006, 11, 3641–3646. [Google Scholar]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 4, 762. [Google Scholar] [CrossRef]
- Sheu, C.S. Chrysanthemum Breeding. In Proceedings of the Symposium on Achievements of Floriculture Research Team of Councial of Agriculture in 2011; Taiwan Agricultural Research Institute, COA: Taichung, Taiwan, 2012; pp. 61–70. [Google Scholar]
- Li, Z.M.; Zhou, H.G.; Chen, Y.; Chen, M.Y.; Yao, Y.T.; Luo, H.H.; Wu, Q.; Wang, F.L.; Zhou, Y.W. Analysis of Transcriptional and Metabolic Differences in the Petal Color Change Response to High-Temperature Stress in Various Chrysanthemum Genotypes. Agronomy 2024, 12, 2863. [Google Scholar] [CrossRef]
- Wu, D.; Wu, Y.X.; Gao, R.Q.; Zhang, Y.H.; Zheng, R.Y.; Fang, M.H.; Li, Y.H.; Zhang, Y.; Guan, L.; Gao, Y.Q. Integrated Metabolomics and Transcriptomics Reveal the Key Role of Flavonoids in the Cold Tolerance of Chrysanthemum. Int. J. Mol. Sci. 2024, 14, 7589. [Google Scholar] [CrossRef]
- Uranishi, R.; Aedla, R.; Alsaadi, D.H.M.; Wang, D.X.; Kusakari, K.; Osaki, H.; Sugimura, K.; Watanabe, T. Evaluation of Environmental Factor Effects on the Polyphenol and Flavonoid Content in the Leaves of Chrysanthemum indicum L. and Its Habitat Suitability Prediction Mapping. Molecules 2024, 5, 927. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, K. Methods of chemical analysis of green tea. Jpn. Agric. R. Q. 1975, 3, 161–164. [Google Scholar]
- Ikegaya, K.; Masuda, M. A New Simple Determination Method of Total Amino Acids in Tea. Natl. R. Ins. Tea 1986, 63, 35–36. [Google Scholar] [CrossRef]
- Li, Y.; Ling, W.Z.; Ye, Y.; Li, M.D.; Lan, F.C.; Xiao, A.L.; Zheng, K.W.; Huang, G.C.; Huang, Y. Study on the Determination of Flavonoids in Eugenia caryophyllata Thunb. by VIS UV Spectrophotometry. Lishizhen Med. Mater. Medica R. 2007, 2, 271–272. [Google Scholar]
- Chinese National Standards-Taiwan (CNS15022 N6384); Method of Test for Catechins Content in Foods. Bureau of Standards, Metrology and Inspection, Ministry of Economic Affairs: Taipei, Taiwan, 2006.
- List of Editorial Members of Taiwan Herbal Pharmacopeia. Chrysanthemi Flos. In Taiwan Herbal Pharmacopeia, 3rd ed.; Ministry of Health and Welfare Taiwan (MOHW): Taipei, Taiwan, 2019; pp. 348–350. [Google Scholar]
- Available online: https://www.fda.gov.tw/upload/133/Content/2012110809292523102.pdf (accessed on 10 September 2019).
- Lo, C.S.; Wang, F.N. Effects of Transplanting Time on the Plant Growth and Nutrient Uptake of Potted Chrysanthemum. Bull. Taoyuan Dist. Agric. R. Ext. Stn. 2000, 41, 27–50. [Google Scholar]
- Sin, Y. Cultivation and Regulation of Flowering in Echinacea purpurea. Master’s Thesis, Department of Horticulture and Landscape Architecture College of Bioresources and Agriculture, National Taiwan University, Taipei, Taiwan, 2017. [Google Scholar]
- Zhang, Q.L.; Li, J.Z.; Wang, Z.M.; Dai, S.L. Research progress on the genetic regulatory mechanism of flowering in Chrysanthemum. Plant Sci. J. 2023, 6, 768–780. [Google Scholar]
- Sun, X.Z.; Zheng, C.S.; Wang, X.F. Effects of high temperature stress on photosynthesis and chlorophyll fluorescence of cut flower chrysanthemum (Dendranthema grandiflora ‘Jinba’). Chin. J. Appl. Ecol. 2008, 10, 2149–2154. [Google Scholar]
- Lyu, J.; Aiwaili, P.; Gu, Z.; Xu, Y.; Zhang, Y.; Wang, Z.; Huang, H.; Zeng, R.; Ma, C.; Gao, J.; et al. Chrysanthemum MAF2 regulates flowering by repressing gibberellin biosynthesis in response to low temperature. Plant J. 2022, 112, 1159–1175. [Google Scholar] [CrossRef]
- Yang, S.B.; Xu, S.S.; Jiang, X.D.; Shi, C.L.; Wang, Y.P.; Shen, S.H. Correcting the Response of Maximum Leaf Photosynthetic Rate to Temperatures in Crop Models. Acta Agron. Sin. 2018, 5, 750–761. [Google Scholar] [CrossRef]
- Kuo, Y.L.; Lin, T.Y.; Yang, Y.Y.; Chen, H.L.; Yang, C.K.; Yu, S.Y. Photosynthetic characteristics and shade tolerance of 440 native woody species in Taiwan. Taiwan. J. Sci. 2021, 3, 189–220. [Google Scholar]
- Woodward, E.I.; Williams, B.G. Climate and Plant Distribution at Global and Local Scales. Vegetatio 1987, 69, 189–197. [Google Scholar] [CrossRef]
- Wang, K.C.; Zhang, Y.; Chen, X.; Guo, Q.H.; Huang, Y. Comparison of photosynthetic characteristics between white and yellow cultivated species in Chrysanthemum morifolium Ramat. J. Nanjing Agric. Univ. 2009, 2, 151–155. [Google Scholar]
- Zhou, J.; Jiang, X.; Agathokleous, E.; Lu, X.; Yang, Z.; Li, R. High temperature inhibits photosynthesis of chrysanthemum (Chrysanthemum morifolium Ramat.) seedlings more than relative humidity. Front. Plant Sci. 2023, 14, 1272013. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.C.; Zhang, H.F. Simultaneous Determination of Two Organic Acids and Two Flavones in Hangzhou White Chrysanthemum by HPLC. China Pharm. 2015, 9, 1578–1580. [Google Scholar]
- Liang, Y.N.; Guo, Q.S.; Zhang, Z.Y.; Wang, T.Y.; Wang, T. Effects of Climate Factors on Quality of Chrysanthemum morifolium Originated from Wenxian county. China J. Chin. Mater. Medica. 2007, 23, 2474–2477. [Google Scholar]
- Cui, M.; Liang, Z.Y.; Liu, Y.X.; Sun, Q.F.; Wu, D.; Luo, L.P.; Hao, Y.B. Flavonoid Profile of Anoectochilus roxburghii (Wall.) Lindl. Under Short-Term Heat Stress. Plant Physiol. Biochem. 2023, 201, 107896. [Google Scholar] [CrossRef]
- Lu, F.; Sun, X.; Dai, X.D.; Zhang, P.Q.; Ma, Y.P.; Xu, Y.F.; Wang, L.; Zhang, J.C. Integrated Multi-Omics Analysis to Investigate the Molecular Mechanisms Underlying the Response of Auricularia heimuer to High-Temperature Stress. J. Fungi 2025, 3, 167. [Google Scholar] [CrossRef]
- Du, X.L.; Huang, J.X.; Yang, Z.J.; Xiong, D.C. Effects of warming on oxidative damage and defense characteristics and their correlation in leaf and fine root of plants: A review. Chin. J. Plant Ecol. 2024, 2, 135–146. [Google Scholar] [CrossRef]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Oravec, M.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; Parella, T.; Peñuelas, J. Warming differentially influences the effects of drought on stoichiometry and metabolomics in shoots and roots. New Phytol. 2015, 3, 591–603. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Survey Date | Days After Treatment | Day/Night Temp. (°C) | Survival Rate (%) | Plant Height (cm) | Plant Width (cm) |
|---|---|---|---|---|---|
| 7/3 | 20 | 15/13 | 100.00 | 15.58 ± 5.11 c | 24.16 ± 3.08 b |
| 20/15 | 100.00 | 18.29 ± 3.16 b | 24.02 ± 2.01 b | ||
| 25/20 | 100.00 | 24.90 ± 3.83 a | 27.32 ± 2.78 a | ||
| 30/25 | 100.00 | 25.10 ± 3.08 a | 26.74 ± 2.53 a | ||
| 35/30 | 100.00 | 18.16 ± 2.68 b | 23.97 ± 2.30 b | ||
| 7/23 | 40 | 15/13 | 100.00 | 17.20 ± 5.40 c | 30.94 ± 3.06 b |
| 20/15 | 96.88 | 19.90 ± 3.40 b | 32.30± 3.22 ab | ||
| 25/20 | 100.00 | 27.90 ± 3.30 a | 33.24 ± 2.96 a | ||
| 30/25 | 100.00 | 27.30 ± 3.30 a | 30.60 ± 5.20 bc | ||
| 35/30 | 87.50 | 19.60 ± 2.30 b | 28.88 ± 2.30 c | ||
| 8/12 | 60 | 15/13 | 100.00 | 19.50 ± 8.00 d | 39.08 ± 4.38 a |
| 20/15 | 96.88 | 28.10 ± 6.50 c | 35.88 ± 4.55 b | ||
| 25/20 | 100.00 | 41.10 ± 6.30 a | 37.66 ± 3.86 ab | ||
| 30/25 | 81.25 | 35.70 ± 4.40 b | 33.52 ± 4.01 c | ||
| 35/30 | 78.13 | 20.10 ± 4.60 d | 27.44 ± 3.31 d | ||
| 9/1 | 80 | 15/13 | 100.00 | 23.30 ± 2.40 d | 37.16 ± 1.07 b |
| 20/15 | 96.88 | 35.40 ± 1.80 c | 36.84 ± 0.80 b | ||
| 25/20 | 100.00 | 47.90 ± 1.30 a | 41.16 ± 1.08 a | ||
| 30/25 | 81.25 | 40.00 ± 0.80 b | 37.20 ± 0.94 b | ||
| 35/30 | 78.13 | 25.60 ± 1.00 d | 29.16 ± 0.52 c | ||
| 9/21 | 100 | 15/13 | 100.00 | 26.30 ± 13.30 c | 39.68 ± 4.18 b |
| 20/15 | 96.88 | 40.40 ± 10.90 b | 37.30 ± 4.83 b | ||
| 25/20 | 96.88 | 61.20 ± 7.50 a | 43.74 ± 6.76 a | ||
| 30/25 | 81.25 | 42.90 ± 5.30 b | 38.02 ± 4.60 b | ||
| 35/30 | 78.13 | 27.00 ± 3.90 c | 33.48 ± 3.26 c | ||
| 10/11 | 120 | 15/13 | 100.00 | 30.50 ± 12.70 c | 37.88 ± 5.55 b |
| 20/15 | 96.88 | 41.20 ± 10.70 b | 33.96 ± 3.79 c | ||
| 25/20 | 96.88 | 66.50 ± 5.40 a | 41.24 ± 5.48 a | ||
| 30/25 | 75.00 | 43.50 ± 6.90 b | 36.13 ± 5.54 bc | ||
| 35/30 | 68.75 | 29.20 ± 3.90 c | 34.10 ± 3.08 c | ||
| 10/31 | 140 | 15/13 | 100.00 | 37.20 ± 12.10 cd | 37.76 ± 4.29 b |
| 20/15 | 96.88 | 41.30 ± 9.60 c | 34.70 ± 3.99 bc | ||
| 25/20 | 93.75 | 67.30 ± 5.90 a | 49.96 ± 6.43 a | ||
| 30/25 | 75.00 | 47.80 ± 8.10 b | 40.83 ± 8.36 b | ||
| 35/30 | 68.75 | 33.00 ± 4.50 d | 39.30 ± 6.33 b | ||
| 11/20 | 160 | 15/13 | 100.00 | 39.23 ± 8.10 c | 39.09 ± 5.90 b |
| 20/15 | 96.88 | 39.44 ± 9.73 c | 35.53 ± 3.92 c | ||
| 25/20 | 93.75 | 66.07 ± 6.04 a | 44.16 ± 6.80 a | ||
| 30/25 | 75.00 | 48.38 ± 8.61 b | 44.79 ± 10.46 a | ||
| 35/30 | 68.75 | 34.25 ± 5.16 d | 37.50 ± 7.01 bc | ||
| 12/10 | 180 | 15/13 | 100.00 | 40.14 ± 8.24 c | 37.16 ± 3.73 b |
| 20/15 | 96.88 | 40.61 ± 9.11 c | 32.81 ± 3.17 c | ||
| 25/20 | 93.75 | 65.50 ± 7.10 a | 38.08 ± 6.01 b | ||
| 30/25 | 75.00 | 50.13 ± 7.17 b | 41.14 ± 6.88 a | ||
| 35/30 | 68.75 | 35.45 ± 5.44 d | 38.16 ± 5.73 ab | ||
| 12/30 | 200 | 15/13 | 100.00 | 42.31 ± 6.51 c | 37.08 ± 8.16 a |
| 20/15 | 96.88 | 40.58 ± 9.68 cd | 31.40 ± 2.80 b | ||
| 25/20 | 93.75 | 65.33 ± 7.05 a | 39.05 ± 5.78 a | ||
| 30/25 | 75.00 | 49.88 ± 12.99 b | 39.90 ± 10.56 a | ||
| 35/30 | 68.75 | 36.84 ± 6.12 d | 39.66 ± 8.82 a | ||
| 1/19 | 220 | 15/13 | 100.00 | 42.00 ± 6.34 c | 36.17 ± 3.50 b |
| 20/15 | 96.88 | 40.37 ± 9.14 c | 30.11 ± 2.06 c | ||
| 25/20 | 93.75 | 68.02 ± 5.82 a | 37.83 ± 5.42 b | ||
| 30/25 | 75.00 | 49.83 ± 11.96 b | 44.75 ± 7.72 a | ||
| 35/30 | 68.75 | 35.40 ± 6.83 d | 37.95 ± 10.26 b |
| Day/Night Temp. (°C) | Total Numbers | Fresh Weight (g) | Dry Weight (g) | Numbers of Bud-Leaves/Pot | One Bud-Leaves Dry Weight (g) |
|---|---|---|---|---|---|
| 15/13 | 74 | 651.74 | 87.04 | 2.47 | 1.18 |
| 20/15 | 175 | 888.81 | 132.67 | 5.83 | 0.76 |
| 25/20 | 385 | 1044.50 | 150.21 | 12.83 | 0.39 |
| 30/25 | 432 | 730.91 | 124.62 | 14.40 | 0.29 |
| 35/30 | 246 | 426.04 | 80.61 | 8.20 | 0.33 |
| Day/Night Temp. (°C) | First Harvest Date | Numbers of Harvest | Total Numbers of Flowers | Total Fresh Weight (g) | Total Dry Weight (g) |
|---|---|---|---|---|---|
| 15/13 | 23 September 2019 | 20 | 988 | 963.25 | 127.18 |
| 20/15 | 7 October 2019 | 19 | 479 | 524.73 | 53.25 |
| 25/20 | 29 October 2019 | 14 | 2049 | 2047.66 | 281.83 |
| 30/25 | 14 January 2020 | 2 | 148 | 81.70 | 14.22 |
| 35/30 | – | – | – | – | – |
| Day/Night Temp. (°C) | A (µmol m−2 s−1) | R (µmol m−2 s−1) | gsw (mol m−2 s−1) | E (mmol m−2 s−1) | A/E |
|---|---|---|---|---|---|
| 15/13 | 12.19 ± 0.91 d | −0.57 ± 0.14 a | 0.34 ± 0.07 b | 1.21 ± 0.32 d | 10.93 ± 3.95 a |
| 20/15 | 15.78 ± 1.02 b | −1.30 ± 0.07 b | 0.54 ± 0.14 b | 2.31 ± 0.24 cd | 7.21 ± 0.97 b |
| 25/20 | 17.79 ± 1.17 a | −1.39 ± 0.27 b | 0.52 ± 0.11 b | 2.45 ± 0.45 bc | 7.41 ± 1.00 b |
| 30/25 | 17.72 ± 0.79 a | −1.50 ± 0.21 b | 0.47 ± 0.18 b | 3.39 ± 1.47 ab | 6.38 ± 2.22 b |
| 35/30 | 14.37 ± 0.80 c | −2.21 ± 0.39 c | 1.43 ± 0.77 a | 4.05 ± 0.91 a | 3.85 ± 0.86 c |
| Day/Night Temp. (°C) | 15/13 | 20/15 | 25/20 | 30/25 | 35/30 |
|---|---|---|---|---|---|
| Polyphenols (mg/g) | 10.98 ± 1.79 c | 12.55 ± 2.26 bc | 15.33 ± 1.98 abc | 16.86 ± 3.53 ab | 19.33 ± 6.64 a |
| Free amino acids (mg/g) | 69.49 ± 6.62 a | 55.99 ± 9.98 b | 41.92 ± 3.21 c | 30.14 ± 3.43 d | 25.21 ± 1.56 e |
| Flavonoids (mg/g) | 12.31 ± 1.41 b | 13.07 ± 1.07 b | 14.34 ± 2.74 ab | 16.34 ± 3.75 ab | 18.34 ± 6.26 a |
| Luteolin (μg/g) | 62.00 ± 0.10 e | 66.50 ± 0.60 d | 70.60 ± 0.30 c | 83.10 ± 2.50 b | 88.00 ± 0.90 a |
| Lutein (μg/g) | 797.60 ± 19.10 c | 880.50 ± 13.80 a | 840.90 ± 15.60 b | 857.90 ± 13.60 ab | 865.50 ± 7.00 ab |
| 3-CQA (mg/g) | 2.12 ± 3.01 c | 7.76 ± 10.42 c | 17.73 ± 1.58 b | 24.23 ± 6.53 ab | 29.04 ± 12.03 a |
| 4-CQA (mg/g) | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.98 ± 0.21 b | 2.19 ± 1.01 a | 2.79 ± 1.52 a |
| 5-CQA (mg/g) | 0.31 ± 0.38 c | 0.37 ± 0.45 c | 0.86 ± 0.19 bc | 1.26 ± 0.49 ab | 1.74 ± 1.05 a |
| 3,4-CQA (mg/g) | 0.33 ± 0.37 d | 0.87 ± 0.24 cd | 1.49 ± 0.62 bc | 2.08 ± 0.55 ab | 2.50 ± 0.78 a |
| 3,5-CQA (mg/g) | 0.00 ± 0.00 b | 2.07 ± 2.77 b | 16.07 ± 7.48 a | 21.33 ± 6.49 a | 22.85 ± 9.43 a |
| 4,5-CQA (mg/g) | 0.10 ± 0.18 b | 0.53 ± 0.7 b | 1.73 ± 0.41 ab | 1.82 ± 1.0 ab | 2.90 ± 1.35 a |
| total CQA (mg/g) | 2.86 ± 3.51 c | 11.60 ± 14.4 c | 38.85 ± 8.99 b | 52.92 ± 15.8 ab | 61.83 ± 25.86 a |
| Day/Night Temp. (°C) | 15/13 | 20/15 | 25/20 | 30/25 | 35/30 | |
|---|---|---|---|---|---|---|
| Polyphenols | (mg) | 519.37 ± 34.91 c | 686.41 ± 165.64 bc | 693.57 ± 54.08 bc | 1481.67 ± 420.80 a | 906.61 ± 76.13 b |
| Free amino acids | 3357.36 ± 639.67 a | 3005.69 ± 348.21 ab | 1900.43 ± 47.60 c | 2631.27 ± 500.97 b | 1371.62 ± 643.19 c | |
| Flavonoids | 584.75 ± 15.89 c | 711.06 ± 100.98 bc | 646.89 ± 91.82 bc | 1436.91 ± 434.98 a | 861.89 ± 83.68 b | |
| Luteolin | 3.13 ± 0.33 b | 3.36 ± 0.21 b | 3.11 ± 0.60 b | 6.39 ± 1.07 a | 5.46 ± 2.41 a | |
| Lutein | 36.77 ± 5.36 b | 46.41 ± 0.82 b | 42.51 ± 5.56 b | 79.30 ± 2.14 a | 41.74 ± 19.25 b | |
| 3-CQA | 93.88 ± 130.80 d | 444.86 ± 597.70 cd | 807.27 ± 92.12 c | 2134.03 ± 714.77 a | 1327.64 ± 150.16 b | |
| 4-CQA | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 44.39 ± 9.36 c | 195.40 ± 101.68 a | 121.61 ± 33.45 b | |
| 5-CQA | 14.97 ± 19.48 c | 21.18 ± 25.54 c | 38.96 ± 7.59 c | 111.53 ± 50.24 a | 73.57 ± 18.18 b | |
| 3,4-CQA | 14.34 ± 16.32 d | 47.64 ± 15.28 cd | 68.80 ± 31.73 c | 182.61 ± 57.51 a | 121.26 ± 36.67 b | |
| 3,5-CQA | 0.00 ± 0.00 c | 117.49 ± 159.78 c | 744.56 ± 375.90 b | 1877.33 ± 669.84 a | 1060.47 ± 304.18 b | |
| 4,5-CQA | 4.49 ± 7.67 d | 30.25 ± 39.32 cd | 78.64 ± 20.15 bc | 162.76 ± 97.42 a | 129.96 ± 15.39 ab | |
| total CQAs | 127.69 ± 151.62 c | 661.41 ± 826.07 c | 1782.62 ± 490.61 b | 4663.66 ± 1670.75 a | 2834.51 ± 526.69 b |
| Day/Night Temp. (°C) | 15/13 | 20/15 | 25/20 | 30/25 |
|---|---|---|---|---|
| Polyphenols (mg/g) | 10.76 ± 0.03 d | 11.86 ± 0.04 c | 16.48 ± 0.01 b | 19.98 ± 0.04 a |
| Free amino acids (mg/g) | 45.59 ± 0.99 b | 64.81 ± 1.68 a | 35.12 ± 0.18 c | 19.33 ± 0.12 d |
| Flavonoids (mg/g) | 14.08 ± 0.10 c | 13.09 ± 0.07 d | 18.13 ± 0.07 a | 16.25 ± 0.08 b |
| Luteolin (μg/g) | 50.59 ± 4.12 d | 61.92 ± 0.83 c | 70.15 ± 0.70 b | 80.18 ± 1.04 a |
| Lutein (μg/g) | 64.83 ± 3.21 b | 77.43 ± 0.84 a | 37.71 ± 0.59 c | 63.96 ± 0.77 b |
| 3-CQA (mg/g) | 1.01 ± 0.23 d | 1.83 ± 0.22 c | 2.79 ± 0.16 b | 4.35 ± 0.61 a |
| 4-CQA (mg/g) | 0.01 ± 0.02 c | 0.02 ± 0.03 c | 0.08 ± 0.03 b | 0.15 ± 0.02 a |
| 5-CQA (mg/g) | 0.10 ± 0.00 d | 0.13 ± 0.01 c | 0.17 ± 0.01 b | 0.20 ± 0.01 a |
| 3,4-CQA (mg/g) | 0.00 ± 0.00 b | 0.13 ± 0.05 b | 0.36 ± 0.08 a | 0.29 ± 0.14 a |
| 3,5-CQA (mg/g) | 0.44 ± 0.06 c | 2.31 ± 0.33 b | 4.67 ± 0.52 a | 3.73 ± 1.12 a |
| 4,5-CQA (mg/g) | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| total CQA (mg/g) | 1.57 ± 0.31 c | 4.40 ± 0.63 b | 8.07 ± 0.79 a | 8.73 ± 1.90 a |
| Day/Night Temp. (°C) | 15/13 | 20/15 | 25/20 | 30/25 | |
|---|---|---|---|---|---|
| Polyphenols | (mg) | 1368.92 ± 3.80 b | 631.58 ± 2.16 c | 4644.39 ± 2.95 a | 284.09 ± 0.62 d |
| Free amino acids | 5797.77 ± 125.30 b | 3450.90 ± 89.28 c | 9896.37 ± 51.66 a | 274.85 ± 1.65 d | |
| Flavonoids | 1790.17 ± 12.99 b | 696.98 ± 3.90 c | 5109.84 ± 18.85 a | 231.04 ± 1.09 d | |
| Luteolin | 6.43 ± 0.52 b | 3.30 ± 0.04 c | 19.77 ± 0.20 a | 1.14 ± 0.01 d | |
| Lutein | 8.24 ± 0.41 b | 4.12 ± 0.04 c | 10.63 ± 0.17 a | 0.91 ± 0.01 d | |
| 3-CQA | 128.65 ± 29.02 b | 97.20 ± 11.83 bc | 785.75 ± 44.73 a | 61.81 ± 8.68 c | |
| 4-CQA | 1.50 ± 2.60 b | 0.89 ± 1.54 b | 21.30 ± 9.74 a | 2.20 ± 0.33 b | |
| 5-CQA | 12.96 ± 0.27 b | 6.66 ± 0.49 c | 48.67 ± 2.73 a | 2.90 ± 0.18 d | |
| 3,4-CQA | 0.00 ± 0.00 b | 6.87 ± 2.57 b | 101.02 ± 22.99 a | 4.15 ± 1.99 b | |
| 3,5-CQA | 56.05 ± 8.26 b | 122.80 ± 17.57 b | 1316.98 ± 145.53 a | 53.10 ± 15.86 b | |
| 4,5-CQA | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | |
| total CQA | 199.16 ± 39.08 c | 234.42 ± 33.49 b | 2273.71 ± 222.83 a | 124.16 ± 26.98 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.-F.; Chen, Y.-J.; Chen, P.-A.; Kuo, C.-C.; Chen, K.-H.; Chen, C.-H.; Su, T.-C.; Chen, I.-Z.; Chang, Y.-S. Impact of Temperature on Growth, Photosynthetic Efficiency, Yield, and Functional Components of Bud-Leaves and Flowers in Edible Chrysanthemum (Chrysanthemum morifolium Ramat). Horticulturae 2025, 11, 448. https://doi.org/10.3390/horticulturae11050448
Liu C-F, Chen Y-J, Chen P-A, Kuo C-C, Chen K-H, Chen C-H, Su T-C, Chen I-Z, Chang Y-S. Impact of Temperature on Growth, Photosynthetic Efficiency, Yield, and Functional Components of Bud-Leaves and Flowers in Edible Chrysanthemum (Chrysanthemum morifolium Ramat). Horticulturae. 2025; 11(5):448. https://doi.org/10.3390/horticulturae11050448
Chicago/Turabian StyleLiu, Chiou-Fang, Yu-Jen Chen, Po-An Chen, Chih-Chun Kuo, Kai-Hsien Chen, Cheng-Hsuan Chen, Tsung-Chen Su, Iou-Zen Chen, and Yu-Sen Chang. 2025. "Impact of Temperature on Growth, Photosynthetic Efficiency, Yield, and Functional Components of Bud-Leaves and Flowers in Edible Chrysanthemum (Chrysanthemum morifolium Ramat)" Horticulturae 11, no. 5: 448. https://doi.org/10.3390/horticulturae11050448
APA StyleLiu, C.-F., Chen, Y.-J., Chen, P.-A., Kuo, C.-C., Chen, K.-H., Chen, C.-H., Su, T.-C., Chen, I.-Z., & Chang, Y.-S. (2025). Impact of Temperature on Growth, Photosynthetic Efficiency, Yield, and Functional Components of Bud-Leaves and Flowers in Edible Chrysanthemum (Chrysanthemum morifolium Ramat). Horticulturae, 11(5), 448. https://doi.org/10.3390/horticulturae11050448