
RrLBD40 Enhances Salt Tolerance in Rosa rugosa via Promoting Root Development


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Subcellular Localization
2.3. Arabidopsis Thaliana Transformation and NaCl Treatment
2.4. R. rugosa Transformation Mediated by Agrobacterium rhizogenes
2.5. Propidium Iodide (PI) Staining
2.6. Measurement of Na+ Flux of Root Tips
2.7. RT-qPCR
2.8. Statistical Analysis
3. Results
3.1. Characterization of RrLBD40
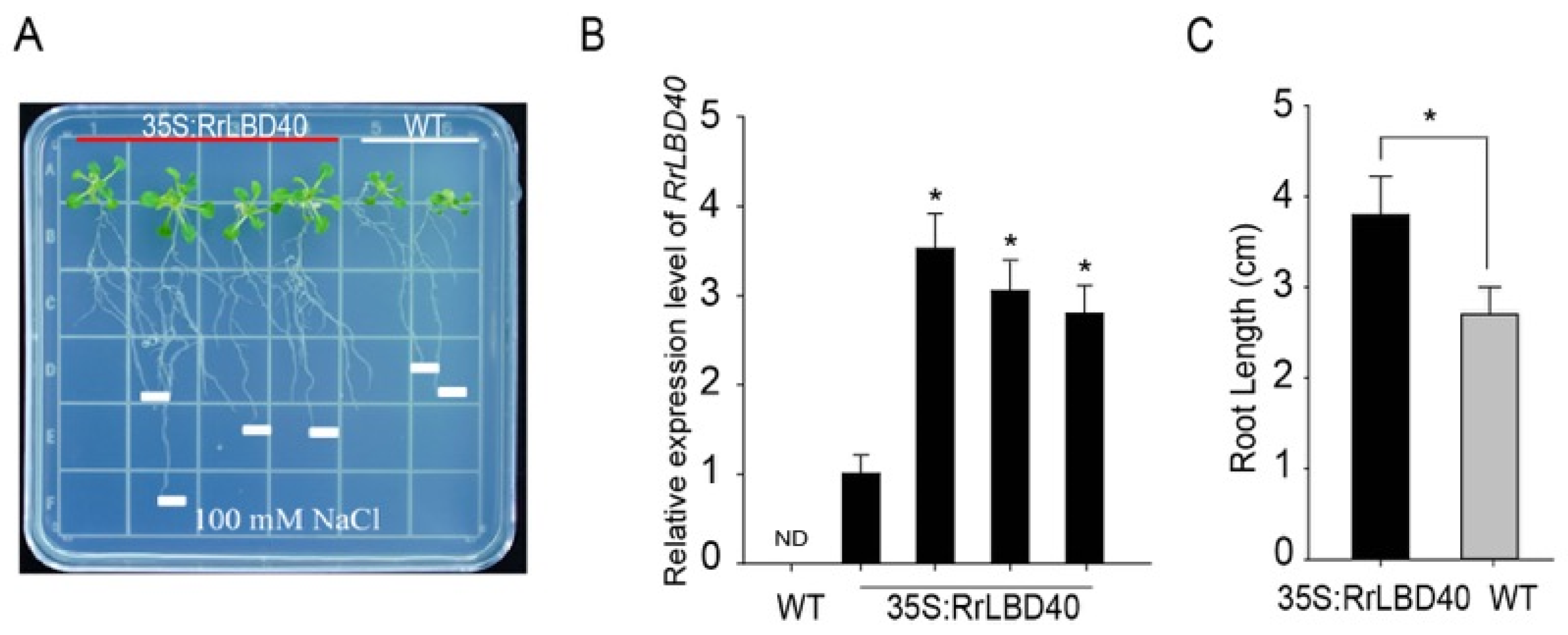
3.2. Ectopic Expression of RrLBD40 Enhances salt Tolerance in Arabidopsis
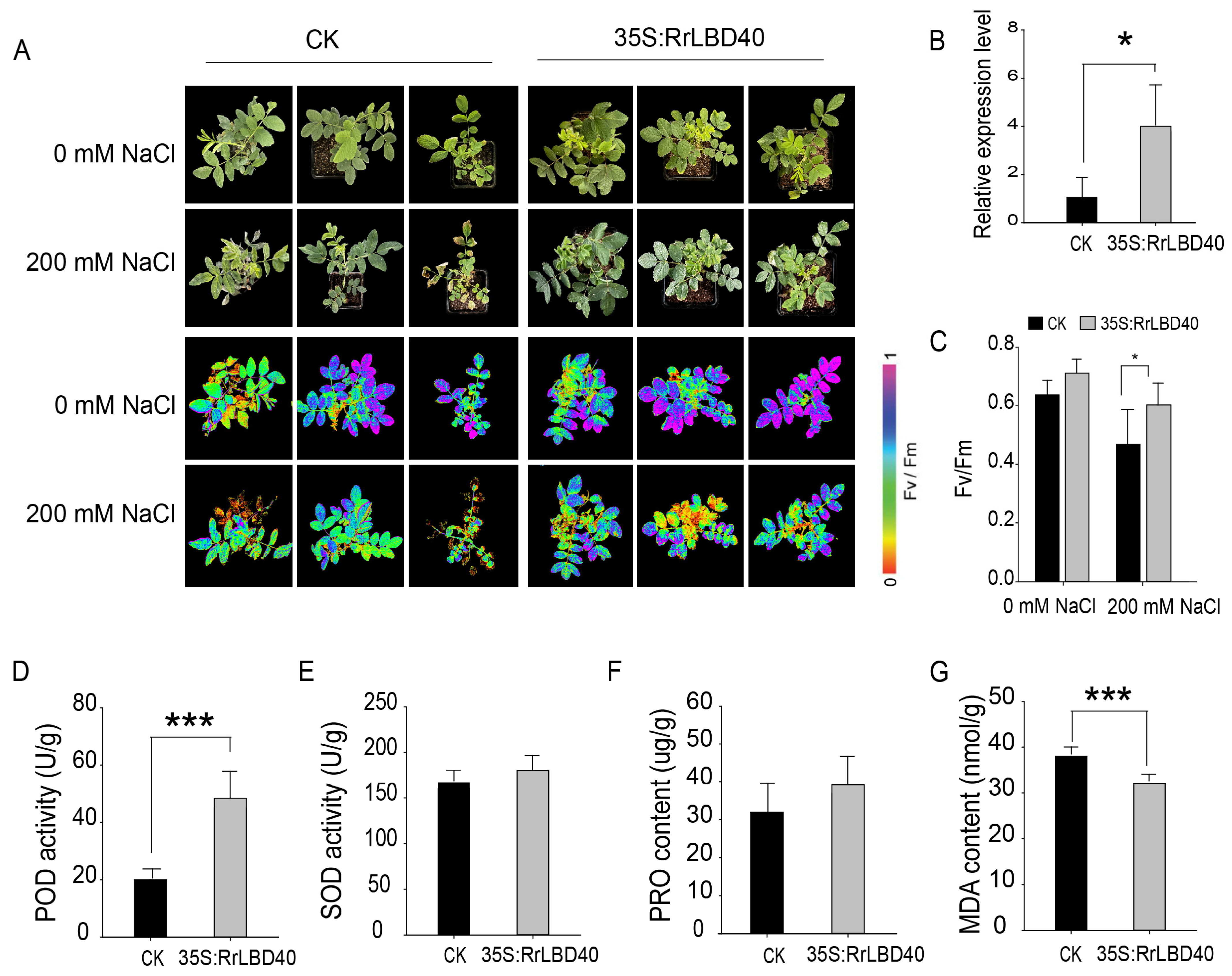
3.3. RrLBD40 Positively Regulates Salt Tolerance in R. rugosa
3.4. Overexpression of RrLBD40 Does Not Affect the Na+ Homeostasis in R. rugosa Roots
3.5. Overexpression of RrLBD40 Promotes Root Development in R. rugosa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Cavusoglu, E.; Sari, U.; Tiryaki, I. Genome-wide identification and expression analysis of Na+/H+ antiporter (NHX) genes in tomato under salt stress. Plant Direct 2023, 7, e543. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. ABA-dependent and ABA-independent signaling in response to osmotic stress in plants. Curr. Opin. Plant Biol. 2014, 21, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Roppolo, D.; De Rybel, B.; Denervaud Tendon, V.; Pfister, A.; Alassimone, J.; Vermeer, J.E.; Yamazaki, M.; Stierhof, Y.D.; Beeckman, T.; Geldner, N. A novel protein family mediates Casparian strip formation in the endodermis. Nature 2011, 473, 380-U564. [Google Scholar] [CrossRef]
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 2007, 19, 118–130. [Google Scholar] [CrossRef]
- Lee, H.W.; Cho, C.; Kim, J. Lateral organ boundaries domain16 and 18 act downstream of the AUXIN1 and LIKE-AUXIN3 auxin influx carriers to control lateral root development in Arabidopsis. Plant Physiol. 2015, 168, 1792-U1177. [Google Scholar] [CrossRef]
- Porco, S.; Larrieu, A.; Du, Y.; Gaudinier, A.; Goh, T.; Swarup, K.; Swarup, R.; Kuempers, B.; Bishopp, A.; Lavenus, J.; et al. Lateral root emergence in Arabidopsis is dependent on transcription factor LBD29 regulation of auxin influx carrier LAX3. Development 2016, 143, 3340–3349. [Google Scholar] [CrossRef]
- Semiarti, E.; Ueno, Y.; Tsukaya, H.; Iwakawa, H.; Machida, C.; Machida, Y. The ASYMMETRIC LEAVES2 gene of Arabidopsis thaliana regulates formation of a symmetric lamina, establishment of venation and repression of meristem-related homeobox genes in leaves. Development 2001, 128, 1771–1783. [Google Scholar] [CrossRef]
- Lin, W.C.; Shuai, B.; Springer, P.S. The Arabidopsis LATERAL ORGAN BOUNDARIES-domain gene ASYMMETRIC LEAVES2 functions in the repression of KNOX gene expression and in adaxial-abaxial patterning. Plant Cell 2003, 15, 2241–2252. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.Z.; Xu, C.Y.; Xu, K.; Hu, Y. LATERAL ORGAN BOUNDARIES DOMAIN transcription factors direct callus formation in Arabidopsis regeneration. Cell Res. 2012, 22, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Cao, H.; Zhang, Q.; Wang, H.; Xin, W.; Xu, E.; Zhang, S.; Yu, R.; Yu, D.; Hu, Y. Control of auxin-induced callus formation by bZIP59-LBD complex in Arabidopsis regeneration. Nat. Plants 2018, 4, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yu, H.; Li, L.G. SUMO modification of LBD30 by SIZ1 regulates secondary cell wall formation in Arabidopsis thaliana. PloS Genet. 2019, 15, e1007928. [Google Scholar] [CrossRef]
- Han, Z.; Yang, T.; Guo, Y.; Cui, W.H.; Yao, L.J.; Li, G.; Wu, A.M.; Li, J.H.; Liu, L.J. The transcription factor PagLBD3 contributes to the regulation of secondary growth in Populus. J. Exp. Bot. 2021, 72, 7092–7106. [Google Scholar] [CrossRef]
- Jiao, P.; Wei, X.; Jiang, Z.; Liu, S.; Guan, S.; Ma, Y. ZmLBD2 a maize (Zea mays L.) lateral organ boundaries domain (LBD) transcription factor enhances drought tolerance in transgenic Arabidopsis thaliana. Front. Plant. Sci. 2022, 13, 1000149. [Google Scholar] [CrossRef]
- Xiong, J.; Mi, X.; Du, L.J.; Wang, X. The LBD Transcription Factor ZmLBD33 Confers Drought Tolerance in Transgenic Arabidopsis. Plants 2025, 14, 1305. [Google Scholar] [CrossRef]
- Li, D.; Chen, X.Z.; Feng, S.Q. The Class II LBD protein MdLBD37 positively regulates the adaptability of apples to drought and salt stress. Biochem. Bioph. Res. Commun. 2025, 754, 151528. [Google Scholar] [CrossRef]
- Liu, Y.; An, X.H.; Liu, H.; Zhang, T.; Li, X.; Liu, R.; Li, C.; Tian, Y.; You, C.; Wang, X.F. Cloning and functional identification of apple LATERAL ORGAN BOUNDARY DOMAIN 3 (LBD3) transcription factor in the regulation of drought and salt stress. Planta 2024, 259, 125. [Google Scholar] [CrossRef]
- Wu, M.; He, W.; Wang, L.; Zhang, X.; Wang, K.; Xiang, Y. PheLBD29, an LBD transcription factor from Moso bamboo, causes leaf curvature and enhances tolerance to drought stress in transgenic Arabidopsis. J. Plant Physiol. 2023, 280, 153865. [Google Scholar] [CrossRef]
- Feng, X.; Xiong, J.; Zhang, W.; Guan, H.; Zheng, D.; Xiong, H.; Jia, L.; Hu, Y.; Zhou, H.; Wen, Y.; et al. ZmLBD5, a class-II LBD gene, negatively regulates drought tolerance by impairing abscisic acid synthesis. Plant J. 2022, 112, 1364–1376. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, J.; Xu, J.; Li, Y.; Guo, L.; Wang, Z.; Zhang, X.; Zhao, B.; Guo, Y.D.; Zhang, N. CRISPR/Cas9 targeted mutagenesis of SlLBD40, a lateral organ boundaries domain transcription factor, enhances drought tolerance in tomato. Plant Sci. 2020, 301, 110683. [Google Scholar] [CrossRef]
- Guan, C.; Wu, B.; Ma, S.; Zhang, J.; Liu, X.; Wang, H.; Zhang, J.; Gao, R.; Jiang, H.; Jia, C. Genome-wide characterization of LBD transcription factors in switchgrass (Panicum virgatum L.) and the involvement of PvLBD12 in salt tolerance. Plant Cell Rep. 2023, 42, 735–748. [Google Scholar] [CrossRef]
- Bai, M.; Liu, J.; Fan, C.; Chen, Y.; Chen, H.; Lu, J.; Sun, J.; Ning, G.; Wang, C. KSN heterozygosity is associated with continuous flowering of Rosa rugosa Purple branch. Hortic. Res. 2021, 8, 26. [Google Scholar] [CrossRef]
- Wei, G.; Chen, Y.; Wang, J.; Feng, L. Molecular cloning and characterization of farnesyl diphosphate synthase from Rosa rugosa Thunb associated with salinity stress. PeerJ 2024, 12, e16929. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cheng, Y.; Shi, X.; Feng, L. GT transcription factors of Rosa rugosa Thunb. involved in salt stress response. Biology 2023, 12, 176. [Google Scholar] [CrossRef] [PubMed]
- Zang, F.; Wu, Q.; Li, Z.; Li, L.; Xie, X.; Tong, B.; Yu, S.; Liang, Z.; Chu, C.; Zang, D.; et al. RrWRKY1, a transcription factor, is involved in the regulation of the salt stress response in Rosa rugosa. Plants 2024, 13, 2973. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shi, Y.; Zhang, W.; Zhu, K.; Feng, L.; Wang, J. C2H2 zinc finger protein family analysis of Rosa rugosa identified a salt-tolerance regulator, RrC2H2-8. Plants 2024, 13, 3580. [Google Scholar] [CrossRef]
- Bao, M.Y.; Xu, Y.; Wei, G.; Bai, M.J.; Wang, J.W.; Feng, L.G. The MYC Gene RrbHLH105 Contributes to Salt Stress-Induced Geraniol in Rose by Regulating Trehalose-6-Phosphate Signalling. Plant Cell Environ. 2025, 48, 1947–1962. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, W.; Cheng, Y.; Feng, L. Genome-Wide Identification of LATERAL ORGAN BOUNDARIES DOMAIN (LBD) Transcription Factors and Screening of Salt Stress Candidates of Rosa rugosa Thunb. Biology 2021, 10, 992. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Wang, Z.; Yamaji, N.; Huang, S.; Zhang, X.; Shi, M.; Fu, S.; Yang, G.; Ma, J.F.; Xia, J. OsCASP1 Is Required for Casparian Strip Formation at Endodermal Cells of Rice Roots for Selective Uptake of Mineral Elements. Plant Cell 2019, 31, 2636–2648. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Nenadić, M.; Vermeer, J.E.M. How to establish a GAPLESS Casparian strip. Nat. Plants 2023, 9, 1585–1586. [Google Scholar] [CrossRef] [PubMed]
- Mahiwal, S.; Andersen, T.G.; Shen, D.F. Establishment and functions of the Casparian strip. Mol. Plant 2025, 18, 1249–1252. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Feng, Z.H.; Tsukagoshi, H.; Yang, M.Y.; Ao, Y.; Fujiwara, T.; Kamiya, T. Early differentiation of Casparian strip mediated by nitric oxide is required for efficient K transport under low K conditions in Arabidopsis. Plant J. 2023, 116, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.G.; Ding, H.; Wang, J.; Wang, M.; Xia, W.; Zang, S.; Sheng, L.X. Molecular cloning and expression analysis of RrNHX1 and RrVHA-c genes related to salt tolerance in wild Rosa rugosa. Saudi J. Biol. Sci. 2015, 22, 417–423. [Google Scholar] [CrossRef]
- Ren, H.R.; Yang, W.J.; Jing, W.K.; Shahid, M.O.; Liu, Y.M.; Qiu, X.H.; Choisy, P.; Xu, T.; Ma, N.; Gao, J.P.; et al. Multi-omics analysis reveals key regulatory defense pathways and genes involved in salt tolerance of rose plants. Hortic. Res. 2024, 11, uhae068. [Google Scholar] [CrossRef]
- Chen, F.; Su, L.Y.; Hu, S.Y.; Xue, J.Y.; Liu, H.; Liu, G.H.; Jiang, Y.F.; Du, J.K.; Qiao, Y.S.; Fan, Y.N.; et al. A chromosome-level genome assembly of rugged rose (Rosa rugosa) provides insights into its evolution, ecology, and floral characteristics. Hortic. Res. 2021, 8, 141. [Google Scholar] [CrossRef]
- Joshi, S.; Hill, K.; Chakrabarti, M.; Perry, S.E. Regulatory mechanisms of the LBD40 transcription factor in Arabidopsis thaliana somatic embryogenesis. Plant Direct 2023, 7, e547. [Google Scholar] [CrossRef]
- Feng, M.Q.; Jiang, N.; Wang, P.B.; Liu, Y.; Xia, Q.M.; Jia, H.H.; Shi, Q.F.; Long, J.M.; Xiao, G.A.; Yin, Z.P.; et al. miR171-targeted SCARECROW-LIKE genes CsSCL2 and CsSCL3 regulate somatic embryogenesis in citrus. Plant Physiol. 2023, 192, 2838–2854. [Google Scholar] [CrossRef] [PubMed]
- Qiao, P.F.; Saleem, N.; Zhao, J.L.; Zhao, C.Z.; Zhang, M. Pan-genome analysis of the LATERAL ORGAN BOUNDARIES domain family in camelina and function investigation of LATERAL ORGAN BOUNDARIES domain 40 in fatty acid synthesis. Int. J. Biol. Macromol. 2025, 330 Pt 3, 148232. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, J.L.; Xu, J.Y.; Li, Y.F.; Lv, H.M.; Wang, F.; Guo, J.X.; Lin, T.; Zhao, B.; Li, X.X.; et al. SlMYC2 promotes SlLBD40-mediated cell expansion in tomato fruit development. Plant J. 2024, 118, 1872–1888. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.L.; Dong, D.H.; Jia, C.Y.; Li, H.X.; Liu, L.; Xu, J.Y.; Cui, H.; Zhang, N.; Guo, Y.D. Fine-tuning of MYC2-mediated Botrytis defense response by the LBD40/42-CRL3BPM4 module in tomato. Plant Cell 2025, 24, koaf258. [Google Scholar] [CrossRef]
- Karahara, I.; Shibaoka, H. The Casparian strip in pea epicotyls: Effects of light on its development. Planta 1994, 192, 269–275. [Google Scholar] [CrossRef]
- Karahara, I.; Ikeda, A.; Kondo, T.; Uetake, Y. Development of the Casparian strip in primary roots of maize under salt stress. Planta 2004, 219, 41–47. [Google Scholar] [CrossRef]
- West, G.; Inze, D.; Beemster, G.T. Cell cycle modulation in the response of the primary root of Arabidopsis to salt stress. Plant Physiol. 2004, 135, 1050–1058. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, J.; Ding, T.; Lin, X.; Hu, H.; Ding, Z.; Tian, H. MYB2 and MYB108 regulate lateral root development by interacting with LBD29 in Arabidopsis thaliana. J. Integr. Plant Biol. 2024, 66, 1675–1687. [Google Scholar] [CrossRef]
- Geng, L.P.; Tan, M.F.; Deng, Q.Y.; Wang, Y.J.; Zhang, T.; Hu, X.S.; Ye, M.M.; Lian, X.M.; Zhou, D.X.; Zhao, Y. Transcription factors WOX11 and LBD16 function with histone demethylase JMJ706 to control crown root development in rice. Plant Cell 2024, 36, 1777–1790. [Google Scholar] [CrossRef]
- Zhang, F.; Tao, W.; Sun, R.; Wang, J.; Li, C.; Kong, X.; Tian, H.; Ding, Z. Correction: PRH1 mediates ARF7-LBD dependent auxin signaling to regulate lateral root development in Arabidopsis thaliana. PLoS Genet. 2022, 18, e1010125. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, M.; Wang, Y.; Shi, Y.; Wang, J.; Feng, L. RrLBD40 Enhances Salt Tolerance in Rosa rugosa via Promoting Root Development. Horticulturae 2025, 11, 1379. https://doi.org/10.3390/horticulturae11111379
Bai M, Wang Y, Shi Y, Wang J, Feng L. RrLBD40 Enhances Salt Tolerance in Rosa rugosa via Promoting Root Development. Horticulturae. 2025; 11(11):1379. https://doi.org/10.3390/horticulturae11111379
Chicago/Turabian StyleBai, Mengjuan, Yue Wang, Yuqing Shi, Jianwen Wang, and Liguo Feng. 2025. "RrLBD40 Enhances Salt Tolerance in Rosa rugosa via Promoting Root Development" Horticulturae 11, no. 11: 1379. https://doi.org/10.3390/horticulturae11111379
APA StyleBai, M., Wang, Y., Shi, Y., Wang, J., & Feng, L. (2025). RrLBD40 Enhances Salt Tolerance in Rosa rugosa via Promoting Root Development. Horticulturae, 11(11), 1379. https://doi.org/10.3390/horticulturae11111379