

Epidemiology and Management of Bean Common Mosaic Virus (BCMV) in Traditional Phaseolus vulgaris L. Landraces within Protected Geographical Indications




Abstract
1. Introduction
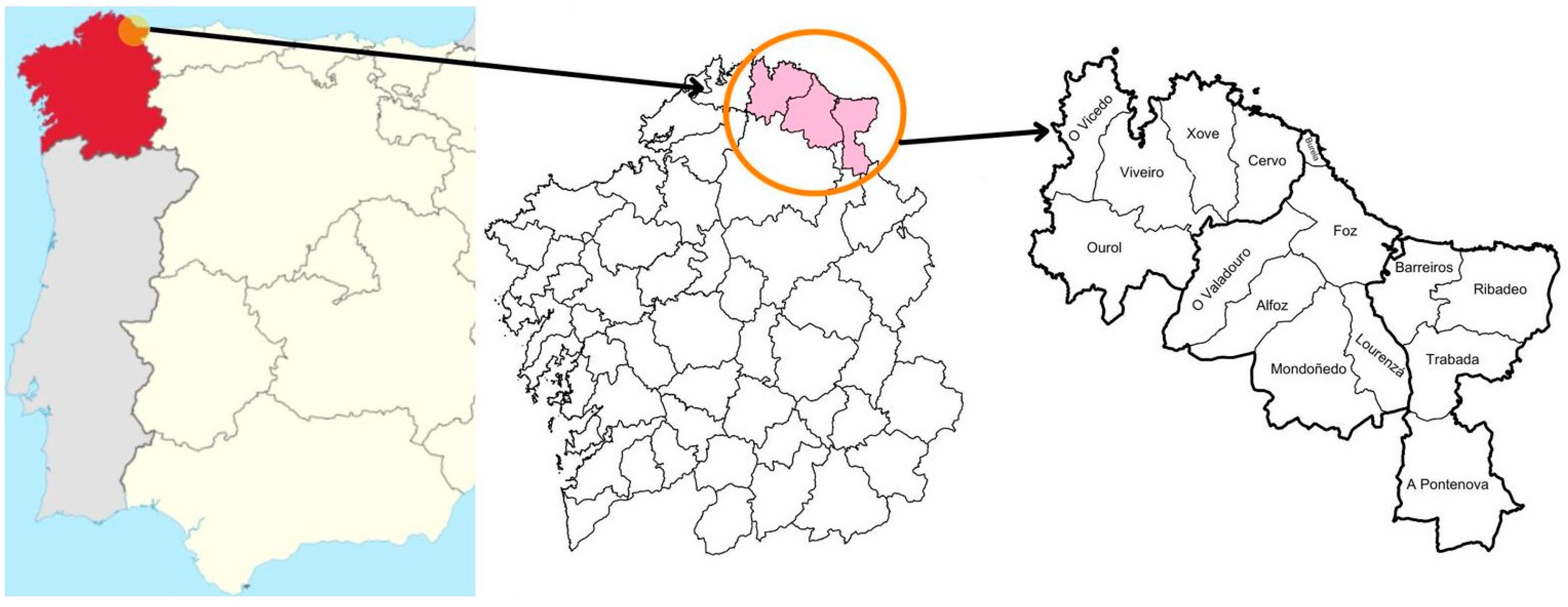
2. Materials and Methods
3. Results
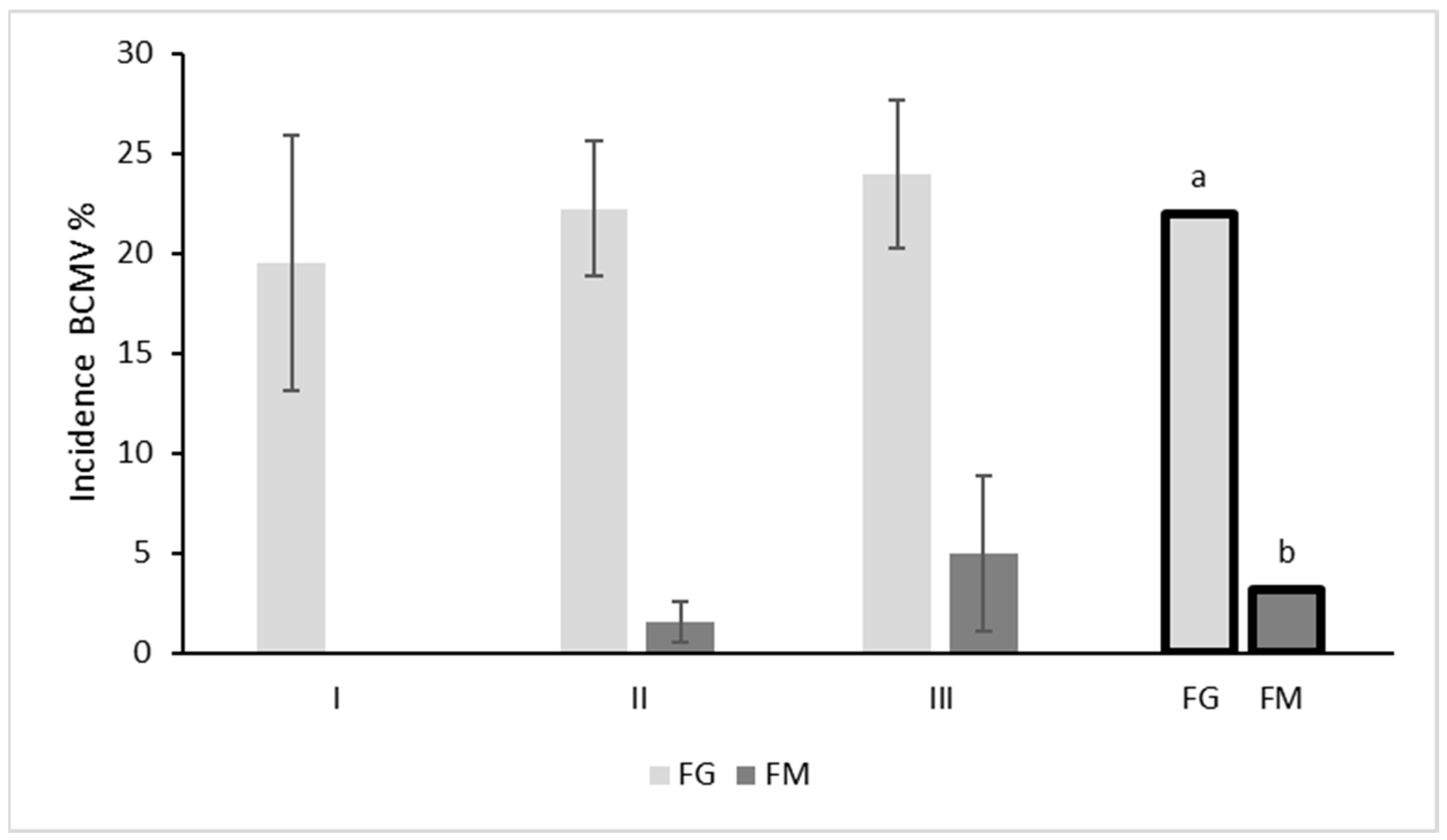
3.1. BCMV and Bacteria in Seed Lots
3.2. Transmission of BCMV to Seeds
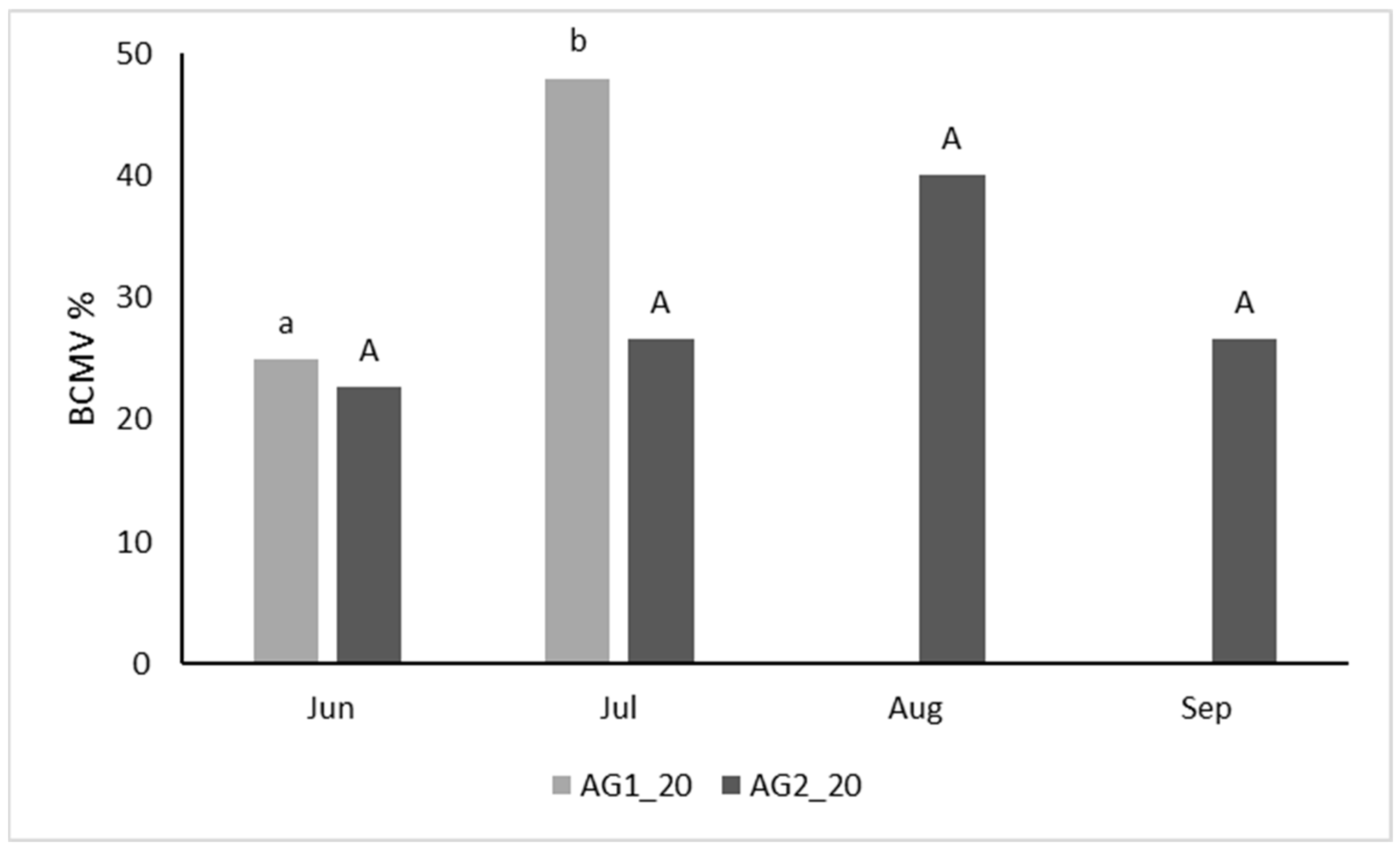
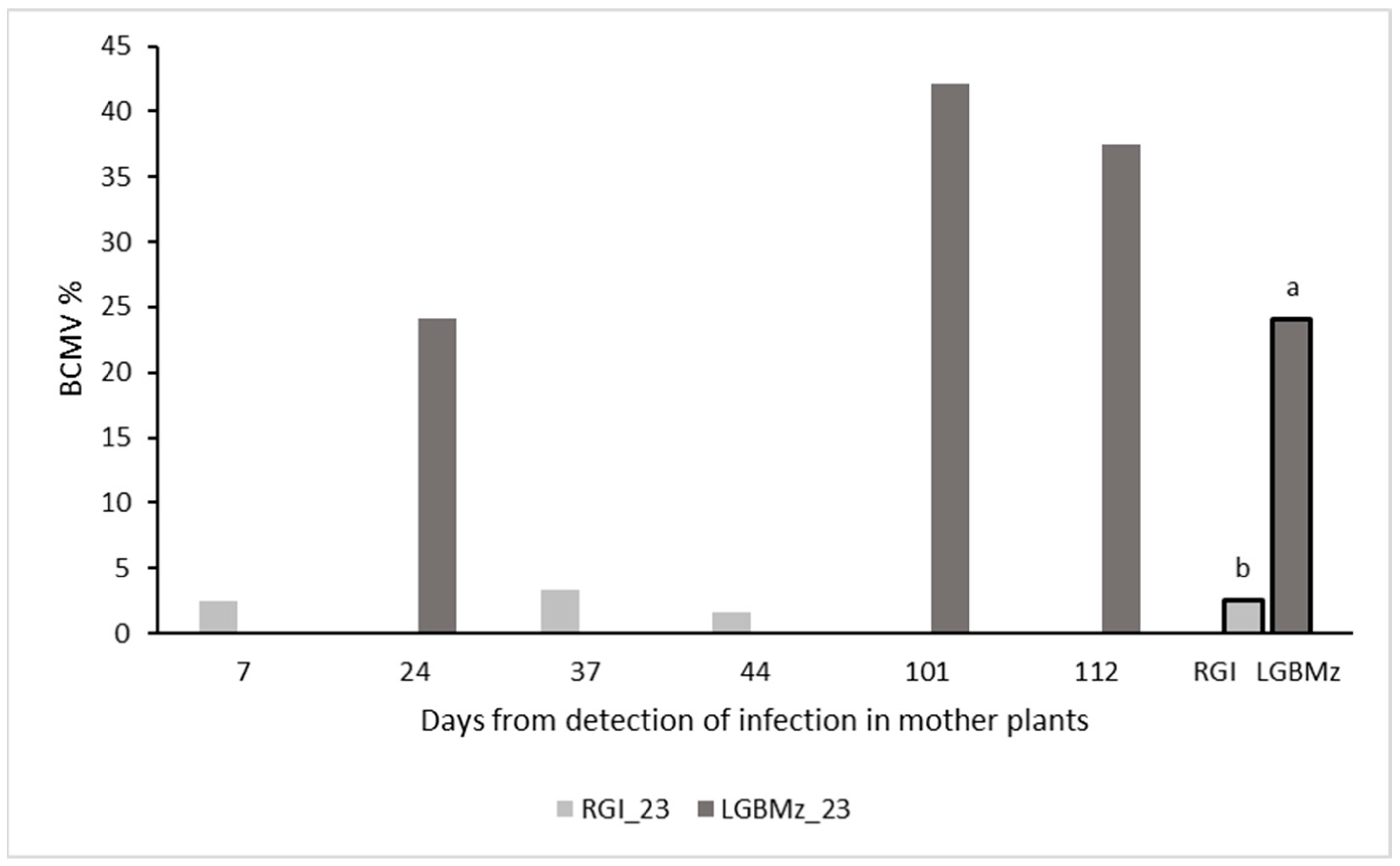
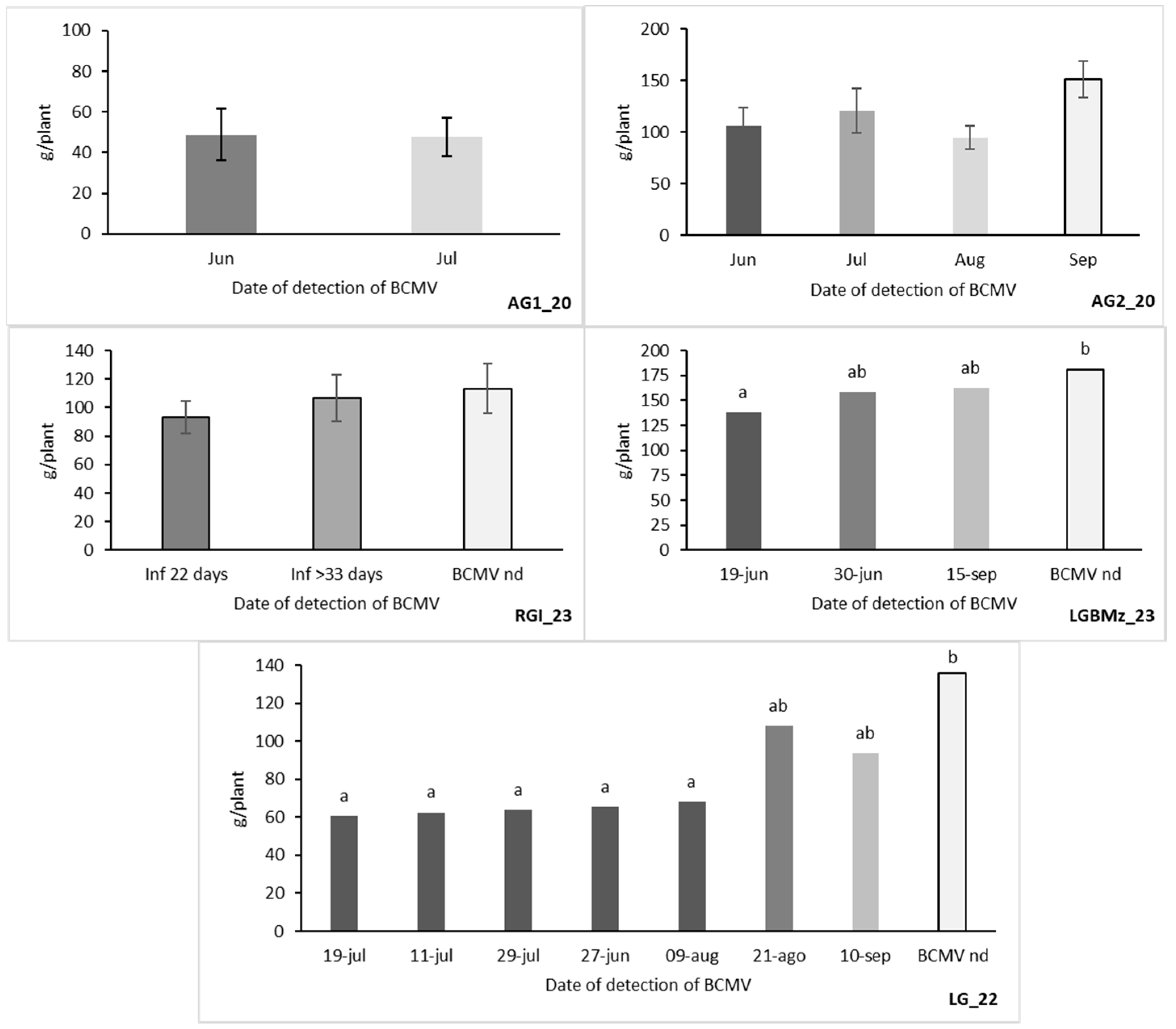
3.3. Harvest
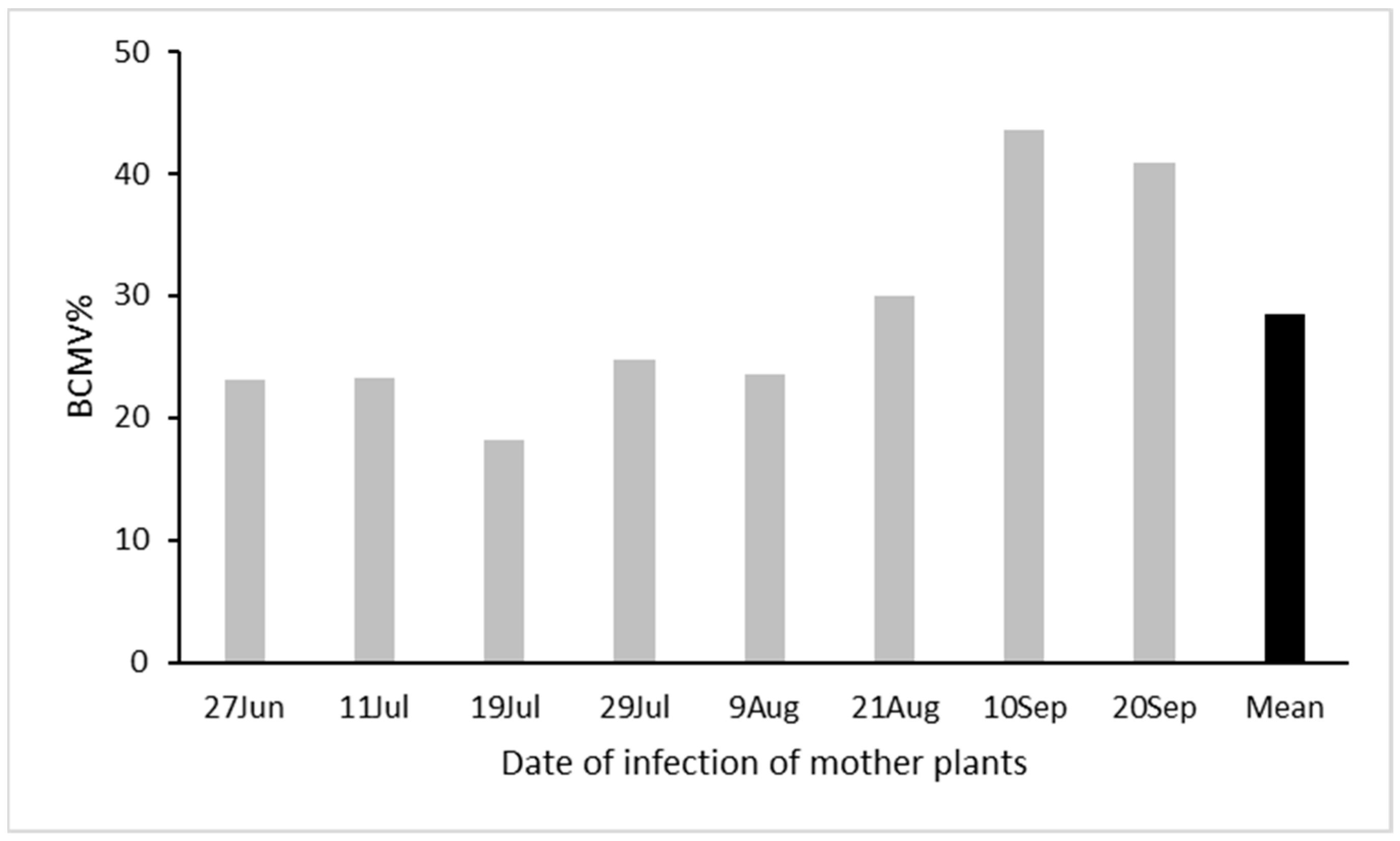
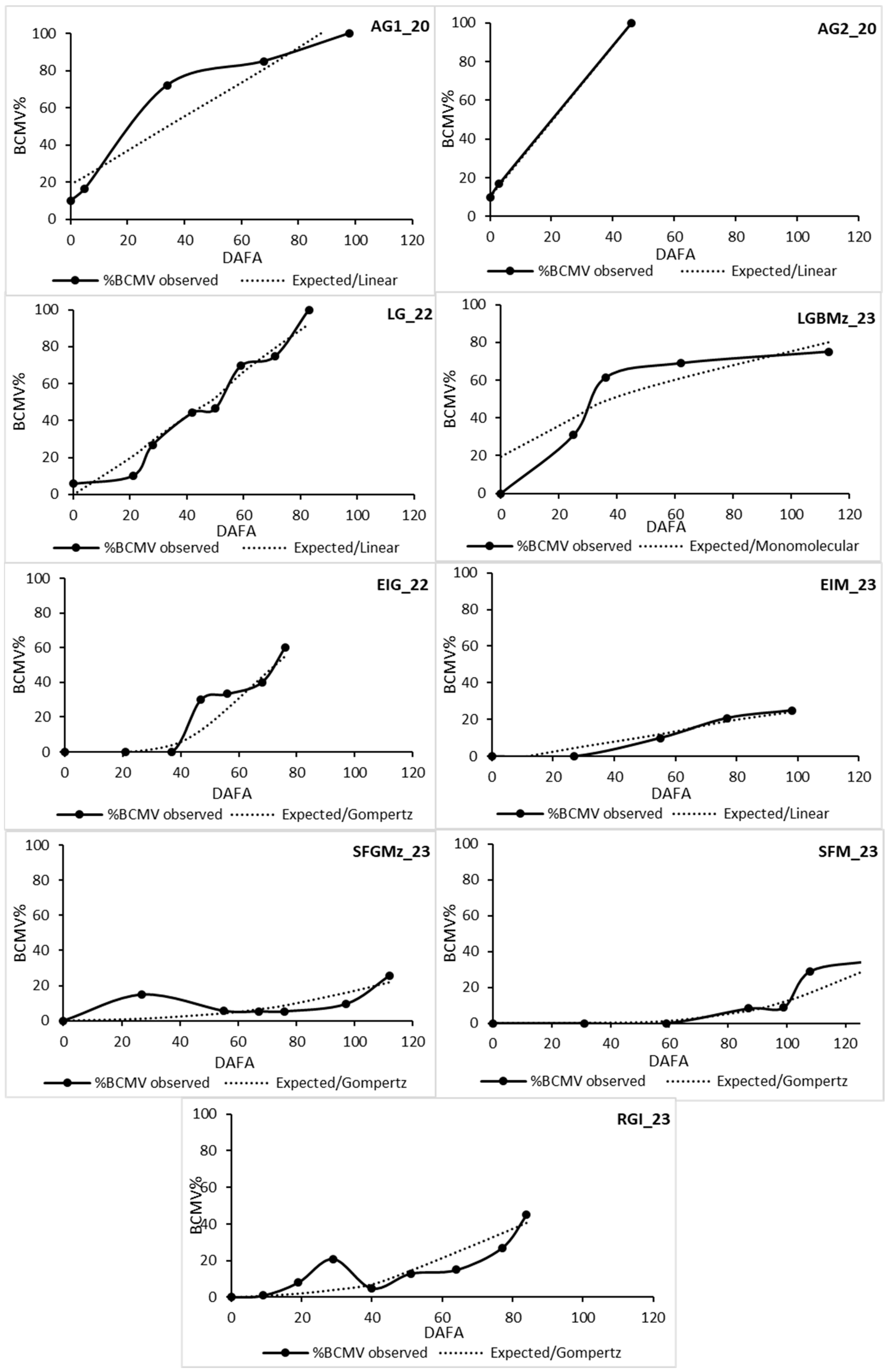
3.4. Epidemiology and Control Strategies
4. Discussion
5. Conclusions
- (a)
- locate plots for seed production far from the growing area where a combination of techniques for preventing virus transmission will be applied by trained growers using an IPM approach,
- (b)
- test representative samples of the seeds produced in those plots and use exclusively lots with the lowest possible virus level, and
- (c)
- train growers to pay special attention to soil and plant management from the beginning of crop planting to delay virus transmission.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santalla, M.; Rodiño, A.P.; De Ron, A.M. Allozyme evidence supporting southwestern Europe as a secondary center of genetic diversity for common bean. Theor. Appl. Genet. 2002, 104, 934–944. [Google Scholar] [CrossRef] [PubMed]
- De Ron, A.M.; Rodiño, A.P.; Santalla, M.; Martínez-Sierra, V. Denominación de Origen Protegida Faba de Lourenzá. In Nuevos Retos y Oportunidades de las Leguminosas en el Sector Agroalimentario Español; De los Mozos, M., Giménez, M.J., Rodríguez Conde, M.F., Sánchez, R., Eds.; Consejería de Agricultura de Castilla—La Mancha: Toledo, Spain, 2006; pp. 387–392. [Google Scholar]
- Boletín Oficial del estado-A-2018-6283. Orden APM/477/2018, de 26 de abril, por la que se dispone la inclusión de diversas variedades de distintas especies en el Registro de Variedades Comerciales: 20170135 faba galaica: 49361-49364. Available online: https://www.boe.es/diario_boe/txt.php?id=BOE-A-2018-6283 (accessed on 31 May 2024).
- Ministerio de Ciencia e Innovación/Consejo Superior de Investigaciones Científicas. Ficha “Faba do Marisco”. Available online: https://www.mbg.csic.es/wp-content/uploads/2021/03/Ficha-Faba-do-Marisco.pdf (accessed on 31 May 2024).
- Pagán, I. Transmission through seeds: The unknown life of plant viruses. PLoS Pathog. 2022, 18, e1010707. [Google Scholar] [CrossRef] [PubMed]
- Dell Olmo, E.; Tiberini, A.; Sigillo, L. Leguminous seedborne pathogens: Seed health and sustainable crop management. Plants 2023, 19, 2040. [Google Scholar] [CrossRef] [PubMed]
- Saiz, M.; de Blas, C.; Carazo, G.; Fresno, J.; Romero, J.; Castro, S. Incidence and characterization of bean common mosaic virus isolates in Spanish bean fields. Plant Dis. 1993, 79, 79–81. [Google Scholar] [CrossRef]
- Melgarejo, P.; García-Jiménez, J.; Jordá, M.C.; López, M.M.; Andrés, M.F.; Duran-Vila, N. Patógenos de plantas descritos en España; Ministerio de Medio Ambiente y Medio Rural y Marino: Madrid, Spain, 2010; p. 49. [Google Scholar]
- Romero, J. Mosaicos de la judía. In Enfermedades de las Plantas Causadas por Virus y Viroides; Ayllón, M.A., Cambra, M., Llave, C., Moriones, E., Eds.; Bubok Publishing S.L. y Sociedad Española de Fitopatología: Madrid, Spain, 2016; pp. 455–466. [Google Scholar]
- Walkey, D.G.A. Ecology and Epidemiology of Plant Viruses 2nd ed. In Applied Plant Virology; Springer: Dordrecht, The Netherlands, 2012; Chapter 8; pp. 210–225. [Google Scholar]
- Morales, F.J.; Castaño, M. Seed transmission characteristics of selected bean common mosaic virus strains in differential bean cultivars. Plant Dis. 1987, 71, 51–53. [Google Scholar] [CrossRef]
- Shukla, D.D.; Ward, C.W.; Brunt, A.A. The Potyviridae; CAB International: Wallingford, UK, 1994; 516p. [Google Scholar]
- Aishwarya, P.; Rangaswamy, K.T.; Basavaraju, S.; Achari, R.; Prameela, H.A. Evaluation of the seed-borne nature of bean common mosaic virus (BCMV) in cowpea. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 239–245. [Google Scholar] [CrossRef]
- Tang, M.; Feng, X. Bean Common Mosaic Disease: Etiology, Resistance Resource, and Future Prospects. Agronomy 2023, 13, 58. [Google Scholar] [CrossRef]
- Gonzalez-Cruces, A.; Arista-Carmona, E.; Díaz-Arias, K.V.; Ramírez-Razo, K.; Hernández-Livera, A.; Acevedo-Sánchez, G.; Mendoza-Ramos, C.; Mora-Aguilera, G. Epidemiology of bean common mosaic virus and Alternaria alternata in 12 Phaseolus vulgaris genotypes. Mex. J. Phytopathol. 2022, 40, 188–220. [Google Scholar] [CrossRef]
- Galvez, E. Aphid-transmitted viruses. In Bean Production Problems: Disease, Insect, Soil and Climatic Constraints of Phaseolus vulgaris; Schwartz, H.F., Gálvez, E., Guillermo, E., Eds.; Centro Internacional de Agricultura Tropical (CIAT): Cali, Colombia, 1980; pp. 211–238. [Google Scholar]
- Meziadi, C.; Blanchet, S.; Geffroy, V.; Pflieger, S. Genetic resistance against viruses in Phaseolus vulgaris L.: State of the art and future prospects. Plant Sci. 2017, 265, 39–50. [Google Scholar] [CrossRef]
- Mangeni, B.H.; Were, K.; Ndong’a, M.; Mukoye, B. Incidence and severity of bean common mosaic disease and resistance of popular bean cultivars to the disease in western Kenya. J. Phytopathol. 2020, 168, 501–515. [Google Scholar] [CrossRef]
- Worrall, E.A.; Wamonje, F.O.; Mukeshimana, G.; Harvey, J.J.W.; Carr, J.P.; Mitter, N. Bean common mosaic virus and bean common mosaic necrosis virus: Relationships, biology, and prospects for control. Adv. Virus Res. 2015, 93, 1–46. [Google Scholar]
- Sastry, K.S. Seed-Borne Plant Virus Diseases; Springer: New Delhi, India, 2013; 353p. [Google Scholar]
- Hampton, R.O. The nature of bean yield reduction by bean yellow and bean common mosaic virus. Phytopathology 1975, 65, 1342–1346. [Google Scholar] [CrossRef]
- Morales, F.J. Common Bean. In Virus and Virus-like Diseases of Major Crops in Developing Countries; Loebenstein, G., Thottappilly, G., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 425–445. [Google Scholar]
- Cifuentes, G.; Castro, S.; Romero, J. Los Potyvirus de judía transmitidos por semilla: Incidencia de la enfermedad y su efecto en los rendimientos. In Proceedings of the II Seminario de la Judía de la Península Ibérica, Villaviciosa, Spain, 5–7 September 2000. [Google Scholar]
- Çelik, A.; Emiralioğlu, O.; Yeken, M.Z.; Çiftçi, V.; Özer, G.; Kim, Y.; Baloch, F.S.; Chung, Y.S. A novel study on bean common mosaic virus accumulation shows disease resistance at the initial stage of infection in Phaseolus vulgaris. Front. Genet. 2023, 14, 1136794. [Google Scholar] [CrossRef] [PubMed]
- Walkley, A.D.G.; Dance, C.M. The effect of oil sprays on aphid transmission of Turnip Mosaic, Beet Yellows, Bean Common and Bean Yellow Mosaic Viruses. Plant Dis. 1979, 63, 877–881. [Google Scholar]
- Dupuis, B.; Cadby, J.; Goy, G.; Tallant, M.; Derron, J.; Schwaerzel, R.; Steinger, T. Control of potato virus Y (PVY) in seed potatoes by oil spraying, straw mulching and intercropping. Plant Pathol. 2017, 66, 960–969. [Google Scholar] [CrossRef]
- Dupuis, B.; Schwaerzel, R.; Goy, G.; Tallant, M.; Derron, J. Stepwise development of an efficient method to control Potato virus Y spread in seed potato fields. In Proceedings of the European Association for Potato Research, Virology Section, Hamar, Norway, 4–9 July 2010; Spetz, C., Blystad, D.R., Eds.; Bioforsk Fokus: Hamar, Norway, 2010; Volume 5, p. 22. [Google Scholar]
- Hooks, C.R.R.; Fereres, A. Protecting crops from non-persistently aphid-transmitted viruses: A review on the use of barrier plants as a management tool. Virus Res. 2006, 120, 1–16. [Google Scholar] [CrossRef]
- Martin-Lopez, B.; Varela, I.; Marnotes, S.; Cabaleiro, C. Use of oils combined with low doses of insecticide for the control of Myzus persicae and PVY epidemics. Pest. Manag. Sci. 2006, 62, 372–378. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Gautam, S.; Rasmussen, D.A.; Srinivasan, R. Aphid transmission of Potyvirus: The largest plant-Infecting RNA virus genus. Viruses 2020, 12, 773. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aziz, M.H. Three modern serological methods to detect plant viruses. J. Plant Sci. Phytopathol. 2019, 3, 101–106. [Google Scholar] [CrossRef]
- Cabaleiro, C.; Marnotes, S.; Couceiro, C.; Martínez, M.A.; Lastra, B.; Martín, B.; García, L.; Alvarez, S. Uso de cultivos borde o asociados para el control de virus no persistentes en patata (PVY) y alubias (BCMV). In Proceedings of the XIV Congreso de la SEF, Lugo, Spain, 15–19 September 2008. [Google Scholar]
- Legorburu, F.J.; Ruiz de Galarreta, J.I.; Abad, J.M.; Pérez de San Román, C. Bean Common (BCMV) and Bean common mosaic necrosis (BCMNV) potyvirus in relation to bean landraces in the Basque Country. Investig. Agr. Prod. Prot. Veg. 1998, 13, 153–158. [Google Scholar]
- Lin, N.S.; Hsu, Y.H.; Hsu, H.T. Immunological detection of plant viruses and a mycoplasmalike organism by direct tissue blotting on nitrocellulose membranes. Phytopathol. 1990, 80, 824–828. [Google Scholar] [CrossRef]
- Cambra, M.; Gorris, M.T.; Roman, M.P.; Terrada, E.; Garnsey, S.M.; Camarasa, E.; Olmos, A.; Colomer, M. Routine detection of Citrus tristeza virus by direct Immunoprinting-ELISA method using specific monoclonal and recombinant antibodies. In Proceedings of the 14th Conference IOCV, Campinas, Brazil, 13–18 September 1998; da Graça, J.V., Lee, R.F., Yokomi, R.K., Eds.; International Organization of Citrus Virologists c/o Department of Plant Pathology: Riverside, CA, USA, 2000. [Google Scholar]
- Madden, L.V.; Hughes, G.; van den Bosch, F. The Study of Plant Disease Epidemics; American Phytopathological Society: St. Paul, MN, USA, 2007; 421p. [Google Scholar]
- Nutter, F.W., Jr.; Eggenberger, S.K.; Littlejohn, K.J. Visualizing, describing, and modeling Disease Progress Curves using EPIMODEL. In Exercises in Plant Disease Epidemiology, 2nd ed.; Stevenson, K.L., Jeger, M.J., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2015; pp. 21–30. [Google Scholar]
- Mwaipopo, B.; Nchimbi-Msolla, S.; Njau, P.; Tairo, F.; William, M.; Binagwa, P.; Kweka, E.; Kilango, M.; Mbanzibwa, D. Viruses infecting common bean (Phaseolus vulgaris L.) in Tanzania: A review on molecular characterization, detection and disease management options. Afr. J. Agric. Res. 2017, 12, 1486–1500. [Google Scholar]
- Makkouk, K.M.; Kumari, S.G. Detection of ten viruses by the tissue-blot immunoassay (TBIA). Arab. J. Plant Protection 1996, 14, 3–9. [Google Scholar]
- European Plant Protection Organization. ELISA tests for plant pathogenic bacteria. Bull. OEPP/EPPO 2010, 40, 369–372. [Google Scholar] [CrossRef]
- Simmons, H.; Munkvold, G. Seed Transmission in the Potyviridae. In Global Perspectives on the Health of Seeds and Plant Propagation Material; Gullino, M.L., Munkvold, G., Eds.; Plant Pathology in the 21st Century 6; Springer Science+Business Media: Dordrecht, The Netherlands, 2014; pp. 3–15. [Google Scholar]
- Campelo, M.P.; Reinoso, B.; González, A.J. Incidencia y transmisión de Potyvirus en semillas de judía de la I.G.P. “Alubia de la Bañeza-León”. In Proceedings of the XIII Congreso Nacional de la Sociedad Española de Fitopatología, Murcia, Spain, 18–22 September 2006. [Google Scholar]
- González, A.J. Virus fitopatógenos transmisibles por semilla en judía tipo “granja asturiana”. Bol. San. Veg. Plagas 2004, 30, 595–603. [Google Scholar]
- Carazo, G.; Romero, J. Malas hierbas reservorios de los Potyvirus que infectan judías. In Proceedings of the IV Seminario de Judías de la Península Ibérica, Valladolid, Spain, 2–4 September 2008. [Google Scholar]
- Odedara, O.O.; Kumar, P.L. Incidence and diversity of viruses in cowpeas and weeds in the unmanaged farming systems of savanna zones in Nigeria. Arch. Phytopathol. Plant Prot. 2017, 50, 1–12. [Google Scholar] [CrossRef]
- Boiteau, G.; Singh, M.; Lavoie, J. Crop border and mineral oil sprays used in combination as physical control methods of the aphid-transmitted potato virus Y in potato. Pest. Manag. Sci. 2009, 65, 255–259. [Google Scholar] [CrossRef]
- Lacomme, C.; Pickup, J.; Fox, A.; Glais, L.; Dupuis, B.; Steinger, T.; Rolot, J.L.; Valkonen, J.P.T.; Kruger, K.; Nie, X.; et al. Transmission and Epidemiology of Potato virus Y. In Potato virus Y: Biodiversity, Pathogenicity, Epidemiology and Management; Lacomme, C., Glais, L., Bellstedt, D., Dupuis, B., Karasev, A., Jacquot, E., Eds.; Springer: Cham, Germany, 2017; pp. 141–176. [Google Scholar]
- Santalla, M.; Rodiño, A.P.; Casquero, P.; de Ron, A.M. Interactions of bush bean intercropped with field and sweet maize. Eur. J. Agron. 2001, 15, 185–196. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Group | Year | #Lots | Identification 1 |
|---|---|---|---|---|
| CSIC | I | 2014 | 1 | C_G (1) |
| 2017 | 1 | C_G (2) | ||
| 2018 | 1 | C_G (3) | ||
| 2019 | 1 | C_G (4) | ||
| 2021 | 1 | C_G (5) | ||
| TERRAS DA MARIÑA S.COOP.GALEGA (LOURENZÁ) | II | 2018 | 2 | CTM_G (1–2) |
| 2019 | 6 | CTM_G (3–8) | ||
| 2021 | 3 | CTM_M (1–2.7) | ||
| 2022 | 4 | CTM_M (3–6) | ||
| 2022 | 19 | CTM_G (9–27) | ||
| ALFOZ & LOURENZÁ (out of cooperative) | III | 2019 | 3 | AL_G (1–3) |
| 2019 | 3 | L_G (1–3) | ||
| 2019 | 1 | LuC_G (1) | ||
| 2020 | 1 | L_G (4) | ||
| 2021 | 1 | L_G (5) | ||
| 2021 | 1 | L_G (6) | ||
| 2021 | 1 | LuC_G (2) | ||
| <2021 | 2 | LuC_M (1–2) | ||
| 2022 | 5 | LuC_M (3–7) | ||
| Total # lots | 57 |
| Plot | Year | cv | Location | Study | |||
|---|---|---|---|---|---|---|---|
| DPC 1 | Harvest | TTS 2 | One Test 3 | ||||
| AG1_20 | 2020 | FG | Alfoz | + | + | + | |
| AG2_20 | 2020 | FG | Alfoz | + | + | + | |
| LG_20 | 2020 | FG | Lourenzá | - | - | - | + |
| LG_21 | 2021 | FG | Lourenzá | - | - | + | + |
| LG_22 | 2022 | FG | Lourenzá | + | + | + | |
| LGBMz_23 | 2023 | FG | Lourenzá | + | + | + | |
| MGI_22 | 2022 | FG | Mondoñedo | - | - | - | |
| MGBMz_22 | 2022 | FG | Mondoñedo | - | - | - | |
| RGI_23 | 2023 | FG | Ribadeo | + | + | + | |
| EIG_22 | 2022 | FG | Lugo | + | - | - | |
| EIM_23 | 2023 | FM | Lugo | + | - | - | |
| SFM_23 | 2023 | FM | Lugo | + | - | - | |
| SFGMz_23 | 2023 | FG | Lugo | + | - | - | |
| LGMz1_23 | 2023 | FG | Lourenzá | - | - | - | + |
| LGMz2_23 | 2023 | FG | Lourenzá | - | - | - | + |
| LM_23 | 2023 | FM | Lourenzá | - | - | - | + |
| Plot | % BCMV Seedlings | Sampling | Frequency | Samples/Date |
|---|---|---|---|---|
| AG1_20 | 10% | Same plants 1 | monthly | 107 |
| AG2_20 | 10% | Same plants 1 | monthly | 104 |
| LG_20 | nt 2 (>20%) | Random | at harvest | >500 |
| LG_21 | nt (>20%) | All plants in 2 lines | at harvest | 200 |
| LG_22 | nt | Same plants 1 | 10 days | 200 (300) |
| LGBMz_23 | 0 | Random & same | monthly | 335 |
| MGI_22 | 0 | All plants | 10 days | 360 |
| MGBMz_22 | 0 | All plants | 10 days | 450 |
| RGI_23 | 0 | All plants | 10 days | 220 |
| EIG_22 | 0 | All plants | 15 days | 50 |
| EIM_23 | 0 | All plants | 15–30 days | 50 |
| SFM_23 | 0 | All plants | 15–30 days | 50 |
| SFGMz_23 | 0 | All plants | 15–30 days | 50 |
| LGMz1_23 | nt | Random | At harvest | 75 |
| LGMz2_23 | nt | Random | At harvest | 75 |
| LM_23 | nt | Random | At harvest | 75 |
| Reactive | Company | Procedure | Dilution | Buffers |
|---|---|---|---|---|
| BCMV + AP | Loewe® Biochemical GmbH, Sauerlach, Germany | Direct | 1:800 | BCB |
| BCMV − Ab | Indirect | 1:200 | Carbonate buffer | |
| Anty-rabbit + AP | Bio-Rad Laboratories, S.A., Madrid, Spain | Indirect | 1:3000 | BCB |
| Ps·ph + AP | Agdia Inc. Elkhart, IN, USA | Direct | 1:500 | BCB |
| Xa·ph + AP | Direct | 1:100 | BCB | |
| Ps·ph − Ab | Indirect | 1:100 | Carbonate buffer | |
| Xa·ph − Ab | Indirect | 1:100 | Carbonate buffer |
| Plot | LA 1 | Model | R2 | Slope | SEEy | % BCMV Expected | % BCMV Observed |
|---|---|---|---|---|---|---|---|
| AG1_20 | bh | Linear | 0.89 | 0.009 | 0.153 | >100 | 100 |
| AG2_20 | bh | Linear | 0.99 | 0.02 | 0.007 | 100 | 100 |
| LG_20 | bh | - | 97 | ||||
| LG_21 | bh | - | 100 | ||||
| LG_22 | bh | Linear | 0.95 | 0.017 | 0.077 | 92 | 98 |
| LGBMz_23 | h | Monomolecular | 0.82 | 0.012 | 0.28 | 80 | 75 |
| MGI_22 | 35 d 2 | - | - | 40 | |||
| MGBMz_22 | 56 d 2 | - | - | 75 | |||
| RGI_23 | h | Gompertz | 0.74 | 0.02 | 0.750 | 40.5 | 45 |
| EIG_22 | h | Gompertz | 0.80 | 0.04 | 0.62 | 55 | 60 |
| EIM_23 | h | Linear | 0.92 | 0.003 | 0.037 | 24 | 25 |
| SFGMz_23 | bh | Gompertz | 0.55 | 0.01 | 0.49 | 22 | 25.7 |
| SFM_23 | h | Gompertz | 0.84 | 0.019 | 0.418 | 31 | 34.8 |
| LGMz1_23 | bh | - | 52.6 | ||||
| LGMz2_23 | bh | - | 59.7 | ||||
| LM_23 | h | - | 45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Expósito-Goás, S.; Pinacho-Lieti, L.G.; Lago-Pena, F.; Cabaleiro, C. Epidemiology and Management of Bean Common Mosaic Virus (BCMV) in Traditional Phaseolus vulgaris L. Landraces within Protected Geographical Indications. Horticulturae 2024, 10, 699. https://doi.org/10.3390/horticulturae10070699
Expósito-Goás S, Pinacho-Lieti LG, Lago-Pena F, Cabaleiro C. Epidemiology and Management of Bean Common Mosaic Virus (BCMV) in Traditional Phaseolus vulgaris L. Landraces within Protected Geographical Indications. Horticulturae. 2024; 10(7):699. https://doi.org/10.3390/horticulturae10070699
Chicago/Turabian StyleExpósito-Goás, Sonia, Lautaro Gabriel Pinacho-Lieti, Fernando Lago-Pena, and Cristina Cabaleiro. 2024. "Epidemiology and Management of Bean Common Mosaic Virus (BCMV) in Traditional Phaseolus vulgaris L. Landraces within Protected Geographical Indications" Horticulturae 10, no. 7: 699. https://doi.org/10.3390/horticulturae10070699
APA StyleExpósito-Goás, S., Pinacho-Lieti, L. G., Lago-Pena, F., & Cabaleiro, C. (2024). Epidemiology and Management of Bean Common Mosaic Virus (BCMV) in Traditional Phaseolus vulgaris L. Landraces within Protected Geographical Indications. Horticulturae, 10(7), 699. https://doi.org/10.3390/horticulturae10070699