
Exogenous Application of Methyl Jasmonate Enhanced the Cold Tolerance of Jasminum sambac through Changes in Metabolites and Antioxidants


Abstract
1. Introduction
2. Materials and Methods
2.1. Overview of Plant Materials and Experimental Conditions
2.2. Experimental Design and Sample Collection
2.3. Measurement of Physiological Indices
2.4. Data Collection and Analysis
3. Results
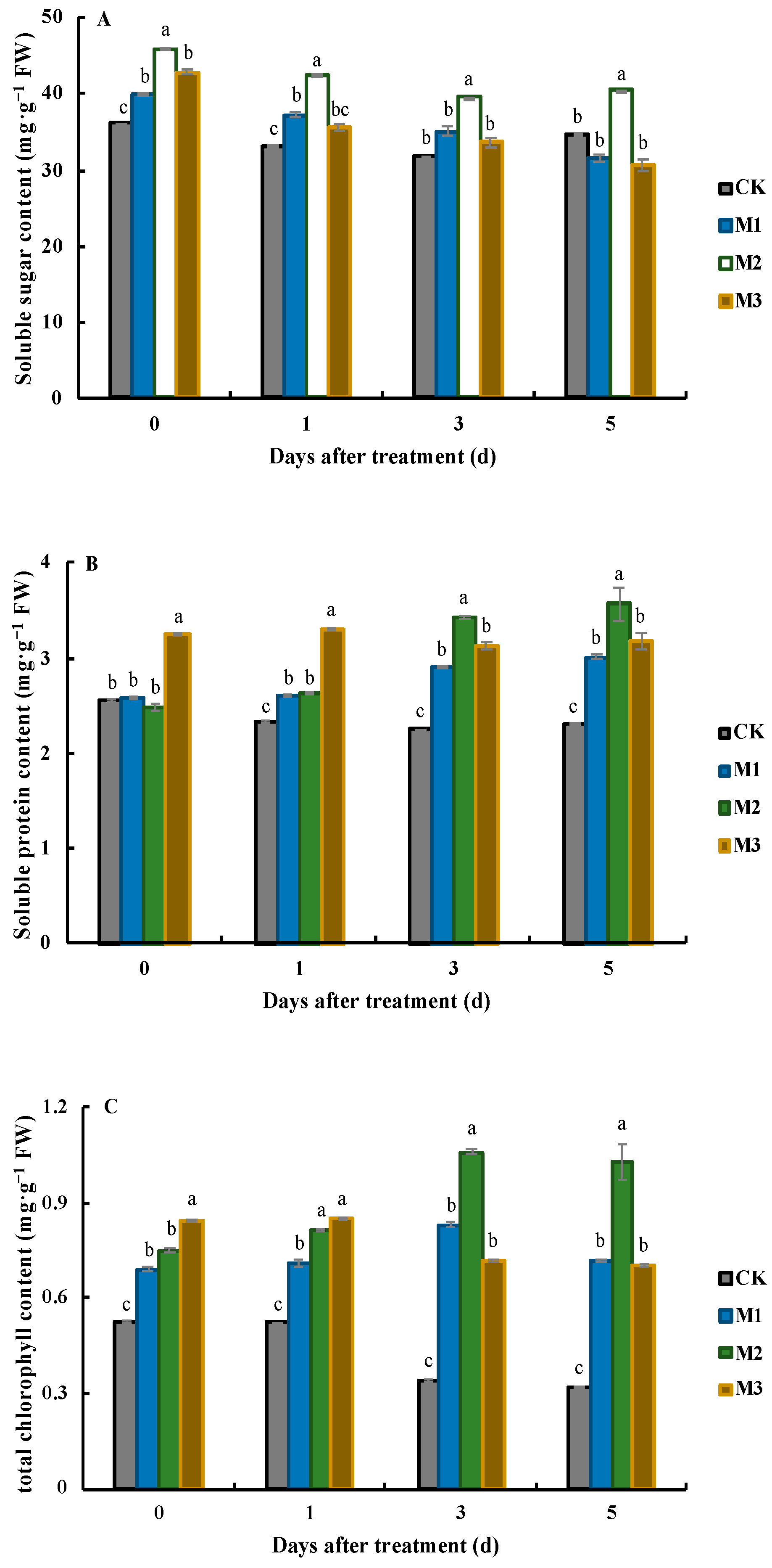
3.1. Effects of MeJA on Nutrient and Chlorophyll Contents in J. sambac Leaves
3.2. Effects of MeJA on MDA and H2O2 Contents in J. sambac Leaves
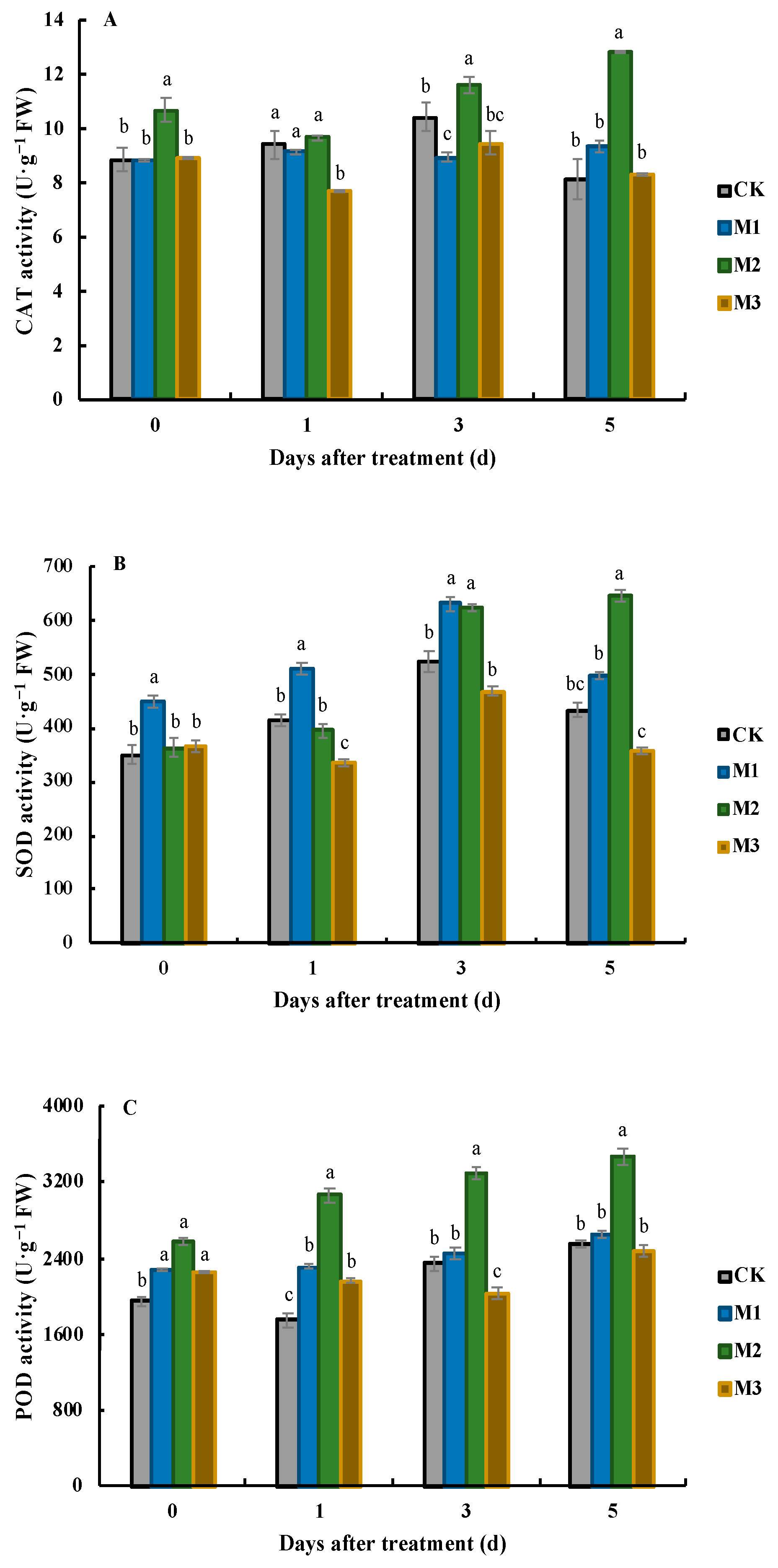
3.3. Effects of MeJA on CAT, SOD, and POD Activities in J. sambac Leaves
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, Y.; Liu, Z.Y.; Lyu, M.L.; Yuan, Y.; Wu, B.H. Characterization of JsWOX1 and JsWOX4 during callus and root induction in the shrub species Jasminum sambac. Plants 2019, 8, 79. [Google Scholar] [CrossRef] [PubMed]
- Supanivatin, P.; Siriwattanayotin, S.; Thipayarat, A.; Ekkaphan, P.; Wongwiwat, J. Effect of overfilled solvent and storage time of subcritical extraction of Jasminum sambac on yield, antioxidant activity, antimicrobial activity and tentative volatile compounds. Plants 2023, 12, 585. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.T.; Zhang, H.L.; Wan, C.; He, X.; Huang, J.F.; Lyu, M.L.; Yuan, Y.; Wu, B.H. Characterization of two BAHD acetyltransferases highly expressed in the flowers of Jasminum sambac (L.) Aiton. Plants 2021, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Al-Snafi, A.E. Pharmacological and therapeutic effects of Jasminum sambac—A review. Indo Am. J. Pharm. Sci. 2018, 5, 1766–1778. [Google Scholar]
- Wei, A.; Shibamoto, T. Antioxidant activities and volatile constituents of various essential oils. J. Agric. Food Chem. 2007, 55, 1737–1742. [Google Scholar] [CrossRef]
- Zhang, H.L.; Wang, W.; Huang, J.F.; Wang, Y.T.; Hu, L.; Yuan, Y.; Lyu, M.L.; Wu, B.H. Role of gibberellin and its three GID1 receptors in Jasminum sambac stem elongation and flowering. Planta 2022, 255, 17. [Google Scholar] [CrossRef]
- Dhanasekaran, D. Influence of growth regulating chemicals on growth and flowering in Jasmine (Jasminum sambac Ait.). J. Hortic. Sci. 2018, 13, 221–226. [Google Scholar] [CrossRef]
- Deng, Y.M.; Jia, X.P.; Liang, L.J.; Gu, C.S.; Sun, X.B. Morphological anatomy, sporogenesis and gametogenesis in flowering process of jasmine (Jasminum sambac Aiton). Sci. Hortic. 2016, 198, 257–266. [Google Scholar] [CrossRef]
- Bera, P.; Mukherjee, C.; Mitra, A. Enzymatic production and emission of floral scent volatiles in Jasminum sambac. Plant Sci. 2016, 256, 25. [Google Scholar] [CrossRef]
- Deng, Y.M.; Qi, X.Y. Research progress on effects of external factors on growth of jasmine. Jiangsu Agric. Sci. 2019, 47, 62–65. (In Chinese) [Google Scholar]
- Niu, S.H. Discussion on the cultivation and management technology of Jasminum sambac. Hortic. Seed 2023, 43, 37–41. (In Chinese) [Google Scholar]
- Cai, H.; He, M.Y.; Ma, K.; Huang, Y.G.; Wang, Y. Salicylic acid alleviates cold-induced photosynthesis inhibition and oxidative stress in Jasminum sambac. Turk. J. Biol. 2015, 39, 241–247. [Google Scholar] [CrossRef]
- Aslam, M.; Fakher, B.; Ashraf, M.A.; Cheng, Y.; Wang, B.R.; Qin, Y. Plant low-temperature stress: Signaling and response. Agronomy 2022, 12, 702. [Google Scholar] [CrossRef]
- Najeeb, S.; Mahender, A.; Anandan, A.; Hussain, W.; Li, Z.; Ali, J. Genetics and breeding of low-temperature stress tolerance in rice. In Rice Improvement: Physiological, Molecular Breeding and Genetic Perspectives; Ali, J., Wani, S.H., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 221–280. [Google Scholar]
- Bao, G.Z.; Tang, W.Y.; An, Q.R.; Liu, Y.X.; Tian, J.Q.; Zhao, N.; Zhu, S.N. Physiological effects of the combined stresses of freezing-thawing, acid precipitation and deicing salt on alfalfa seedlings. BMC Plant Biol. 2020, 20, 204. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.K.; Agarwal, P.; Reddy, M.K.; Sopory, S.K. Role of DREB transcription factors in abiotic and biotic stress tolerance in plants. Plant Cell Rep. 2006, 25, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Zhang, Q.Q.; Ou, L.N.; Ji, D.Z.; Liu, T.; Lan, R.M.; Li, X.Y.; Jin, L.H. Response to the cold stress signaling of the tea plant (Camellia sinensis) elicited by chitosan oligosaccharide. Agronomy 2020, 10, 915. [Google Scholar] [CrossRef]
- Huan, C.; Yang, X.H.; Wang, L.F.; Kebbeh, M.; Wang, Y.X.; Dai, B.G.; Shen, S.L.; Zheng, X.L.; Zhou, H.J. Methyl jasmonate treatment regulates α-linolenic acid metabolism and jasmonate acid signaling pathway to improve chilling tolerance in both stony hard and melting flesh peaches. Postharvest Biol. Technol. 2022, 190, 111960. [Google Scholar] [CrossRef]
- Gul, N.; Masoodi, K.Z.; Ramazan, S.; Mir, J.I.; Aslam, S. Study on the impact of exogenously applied methyl jasmonate concentrations on Solanum lycopersicum under low temperature stress. BMC Plant Biol. 2023, 23, 437. [Google Scholar] [CrossRef]
- Wang, X.Q. Principles and Techniques of Plant Physiological and Biochemical Tests; Higher Education Press: Beijing, China, 2006. (In Chinese) [Google Scholar]
- Ranieri, A.; Castagna, A.; Baldan, B.; Soldatini, G.F. Iron deficiency differently affects peroxidase isoforms in sunflower. J. Exp. Bot. 2001, 52, 25–35. [Google Scholar] [CrossRef]
- Jiang, M.Y.; Zhang, J.H. Water stress -induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulated the activities of antioxidant enzymes in the maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Ni, M.; Yu, F.Y. A study on petal morphological and physiological characteristics of Styrax japonicus during the flowering period. Agronomy 2021, 11, 1498. [Google Scholar] [CrossRef]
- Jeon, J.; Kim, J. Cold stress signaling networks in Arabidopsis. J. Plant Biol. 2013, 56, 69–76. [Google Scholar] [CrossRef]
- Liu, L.L.; Song, H.; Shi, K.J.; Liu, B.; Zhang, Y.; Tang, L.; Cao, W.X.; Zhu, Y. Response of wheat grain quality to low temperature during jointing and booting stages-on the importance of considering canopy temperature. Agric. For. Meteorol. 2019, 278, 107658. [Google Scholar] [CrossRef]
- Zhang, W.J.; Zhao, Y.; Li, L.Y.; Xu, X.; Yang, L.; Luo, Z.; Wang, B.B.; Ma, S.Y.; Fan, Y.H.; Huang, Z.L. The effects of short-term exposure to low temperatures during the booting stage on starch synthesis and yields in wheat grain. Front. Plant Sci. 2021, 12, 684784. [Google Scholar] [CrossRef]
- Han, X.; Li, Y.H.; Yao, M.H.; Yao, F.; Wang, Z.L.; Wang, H.; Li, H. Transcriptomics reveals the effect of short-term freezing on the signal transduction and metabolism of grapevine. Int. J. Mol. Sci. 2023, 24, 3884. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.M.; Li, C.C.; Shao, Q.S.; Ye, X.Q.; She, J.M. Differential response of double petal and multi petal jasmine to shading: I. Photosynthetic characteristics and chloroplast ultrastructure. Plant Physiol. Bioch. 2012, 55, 93–102. [Google Scholar] [CrossRef]
- Wang, P.J.; Fang, J.P.; Lin, H.Z.; Yang, W.W.; Yu, J.X.; Hong, Y.P.; Jiang, M.; Gu, M.; Chen, Q.; Zheng, Y.; et al. Genomes of single- and double-petal jasmines (Jasminum sambac) provide insights into their divergence time and structural variations. Plant Biotechnol. J. 2022, 20, 1232–1234. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Xu, H.F.; Liu, W.J.; Wang, N.; Qu, C.Z.; Jiang, S.H.; Fang, H.C.; Zhang, Z.Y.; Chen, X.S. Methyl jasmonate enhances apple’ cold tolerance through the JAZ-MYC2 pathway. Plant Cell 2019, 136, 75–84. [Google Scholar] [CrossRef]
- Saadati, S.; Baninasab, B.; Mobli, M.; Gholami, M. Enhancement of freezing tolerance of olive leaves by foliar application of methyl jasmonate and 24-epibrassinolide through changes in some metabolites and antioxidant activity. Sci. Hortic. 2021, 284, 110127. [Google Scholar] [CrossRef]
- Serna-Escolano, V.; Martínez-Romero, D.; Giménez, M.J.; Serrano, M.; García-Martínez, S.; Valero, D.; Valverde, J.M.; Zapata, P.J. Enhancing antioxidant systems by preharvest treatments with methyl jasmonate and salicylic acid leads to maintain lemon quality during cold storage. Food Chem. 2021, 338, 128044. [Google Scholar] [CrossRef]
- Chen, C.; Lai, J.Y.; Chen, H.; Yu, F.Y. Zinc oxide nanoparticles enhanced growth of tea trees via modulating antioxidant activity and secondary metabolites. Horticulturae 2023, 9, 631. [Google Scholar] [CrossRef]
- Wang, K.T.; Lei, C.Y.; Tan, M.L.; Wang, J.S.; Li, C.H.; Zou, Y.Y. Increased soluble sugar accumulation in postharvest peaches in response to different defense priming elicitors. Hortic. Environ. Biotechnol. 2023, 64, 115–131. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Ren, Y.; Yang, D.Y.; Liu, H.D.; Zhang, Y.S.; Wang, X.J.; Bai, F.J.; Cheng, S.R. Foliar methyl jasmonate (MeJA) application increased 2-acetyl-1-Pyrroline (2-AP) content and modulated antioxidant attributes and yield formation in fragrant rice. J. Plant Physiol. 2023, 282, 153946. [Google Scholar] [CrossRef] [PubMed]
- George, I.S.; Pascovici, D.; Mirzaei, M.; Haynes, P.A. Quantitative proteomic analysis of cabernet sauvignon grape cells exposed to thermal stresses reveals alterations in sugar and phenylpropanoid metabolism. Proteomics 2015, 15, 3048–3060. [Google Scholar] [CrossRef]
- Zhu, A.D.; Li, W.Y.; Ye, J.L.; Sun, X.H.; Ding, Y.D.; Cheng, Y.J.; Deng, X.X. Microarray expression profiling of postharvest ponkan mandarin (Citrus reticulata) fruit under cold storage reveals regulatory gene candidates and implications on soluble sugars metabolism. J. Int. Plant Biol. 2011, 53, 358–374. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.L.; Yang, W.B.; Dong, Z.G.; Tang, L.X.; Liu, Y.Y.; Xie, S.Y.; Yang, Y.D. Integrated transcriptomic and metabolomics analyses reveal molecular responses to cold stress in coconut (Cocos nucifera L.) seedlings. Int. J. Mol. Sci. 2023, 24, 14563. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.W.; Wang, R.J.; Gong, J.J.; Zhu, T.Q.; Long, S.; Guo, H.; Liu, T.Y.; Yang, P.Z.; Xu, Y.F. Combined cold and drought stress-induced response of photosynthesis and osmotic adjustment in Elymus nutans Griseb. Agronomy 2023, 13, 2368. [Google Scholar] [CrossRef]
- Zhao, Y.Q.; Han, Q.H.; Ding, C.B.; Huang, Y.; Liao, J.Q.; Chen, T.; Feng, S.L.; Zhou, L.J.; Zhang, Z.W.; Chen, Y.E.; et al. Effect of low temperature on chlorophyll biosynthesis and chloroplast biogenesis of rice seedlings during greening. Int. J. Mol. Sci. 2020, 21, 1390. [Google Scholar] [CrossRef]
- Fan, L.L.; Wang, Q.; Lv, J.W.; Gao, L.P.; Zuo, J.H.; Shi, J.Y. Amelioration of postharvest chilling injury in cowpea (Vigna sinensis) by methyl jasmonate (MeJA) treatments. Sci. Hortic. 2016, 203, 95–101. [Google Scholar] [CrossRef]
- Shahzad, R.; Waqas, M.; Khan, A.L.; Hamayun, M.; Kang, S.M.; Lee, I.J. Foliar application of methyl jasmonate induced physio-hormonal changes in Pisum sativum under diverse temperature regimes. Plant Physiol. Biochem. 2015, 96, 406–416. [Google Scholar] [CrossRef]
- Elatafi, E.; Elshahat, A.; Yu, X.; Li, S.N.; Lu, S.W.; Dong, T.Y.; Fang, J.G. Effects of different storage temperatures and methyl jasmonate on grape quality and antioxidant activity. Horticulturae 2023, 9, 1282. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.M.; Al-Kahtani, M.A.; El-Kersh, M.A.; Al-Omair, M.A. Free radical-scavenging, anti-inflammatory/anti-fibrotic and hepatoprotective actions of taurine and silymarin against CCl4 induced rat liver damage. PLoS ONE 2015, 10, e0144509. [Google Scholar] [CrossRef] [PubMed]
- Meloni, D.A.; Oliva, M.A.; Martinez, C.A.; Cambraia, J. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot. 2003, 49, 69–76. [Google Scholar] [CrossRef]
- Repkina, N.; Ignatenko, A.; Holoptseva, E.; Miszalskl, Z.; Kaszycki, P.; Talanova, V. Exogenous methyl jasmonate improves cold tolerance with parallel induction of two cold-regulated (COR) genes expression in Triticum aestivum L. Plants 2021, 10, 1421. [Google Scholar] [CrossRef]
- Nyanasaigran, L.; Ramasamy, S.; Gautam, A.; Guleria, P.; Kumar, V.; Yaacob, J.S. Methyl jasmonate elicitation improves the growth performance and biosynthesis of antioxidant metabolites in Portulaca oleracea through ROS modulation. Ind. Crop. Prod. 2024, 216, 118709. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Day | MeJA | Sampling | Temperature | Sample Label |
|---|---|---|---|---|
| 1 | • | Normal | ||
| 2 | • | |||
| 3 | • | • | 0 d | |
| 4 | • | Low | 1 d | |
| 5 | ||||
| 6 | • | 3 d | ||
| 7 | ||||
| 8 | • | 5 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Chen, H.; Yang, K. Exogenous Application of Methyl Jasmonate Enhanced the Cold Tolerance of Jasminum sambac through Changes in Metabolites and Antioxidants. Horticulturae 2024, 10, 688. https://doi.org/10.3390/horticulturae10070688
Chen C, Chen H, Yang K. Exogenous Application of Methyl Jasmonate Enhanced the Cold Tolerance of Jasminum sambac through Changes in Metabolites and Antioxidants. Horticulturae. 2024; 10(7):688. https://doi.org/10.3390/horticulturae10070688
Chicago/Turabian StyleChen, Chen, Hong Chen, and Kaibo Yang. 2024. "Exogenous Application of Methyl Jasmonate Enhanced the Cold Tolerance of Jasminum sambac through Changes in Metabolites and Antioxidants" Horticulturae 10, no. 7: 688. https://doi.org/10.3390/horticulturae10070688
APA StyleChen, C., Chen, H., & Yang, K. (2024). Exogenous Application of Methyl Jasmonate Enhanced the Cold Tolerance of Jasminum sambac through Changes in Metabolites and Antioxidants. Horticulturae, 10(7), 688. https://doi.org/10.3390/horticulturae10070688