

Genotyping of Croatian Olive Germplasm with Consensus SSR Markers


Abstract
1. Introduction
- To genotype a large and representative number of olive samples using SSR markers, including olive traditional varieties and unnamed genotypes of economic importance;
- To identify synonyms and homonyms of Croatian varieties;
- To create a database of olive SSR profiles comparable to publicly available olive SSR databases and to detect genotypes which are unique only to the Croatian part of the Mediterranean.
2. Materials and Methods
2.1. Plant Material and DNA Isolation
2.2. Molecular Analysis
2.3. Allele Lenght Standardization
2.4. Data Analysis
3. Results
3.1. Synonyms and Homonyms
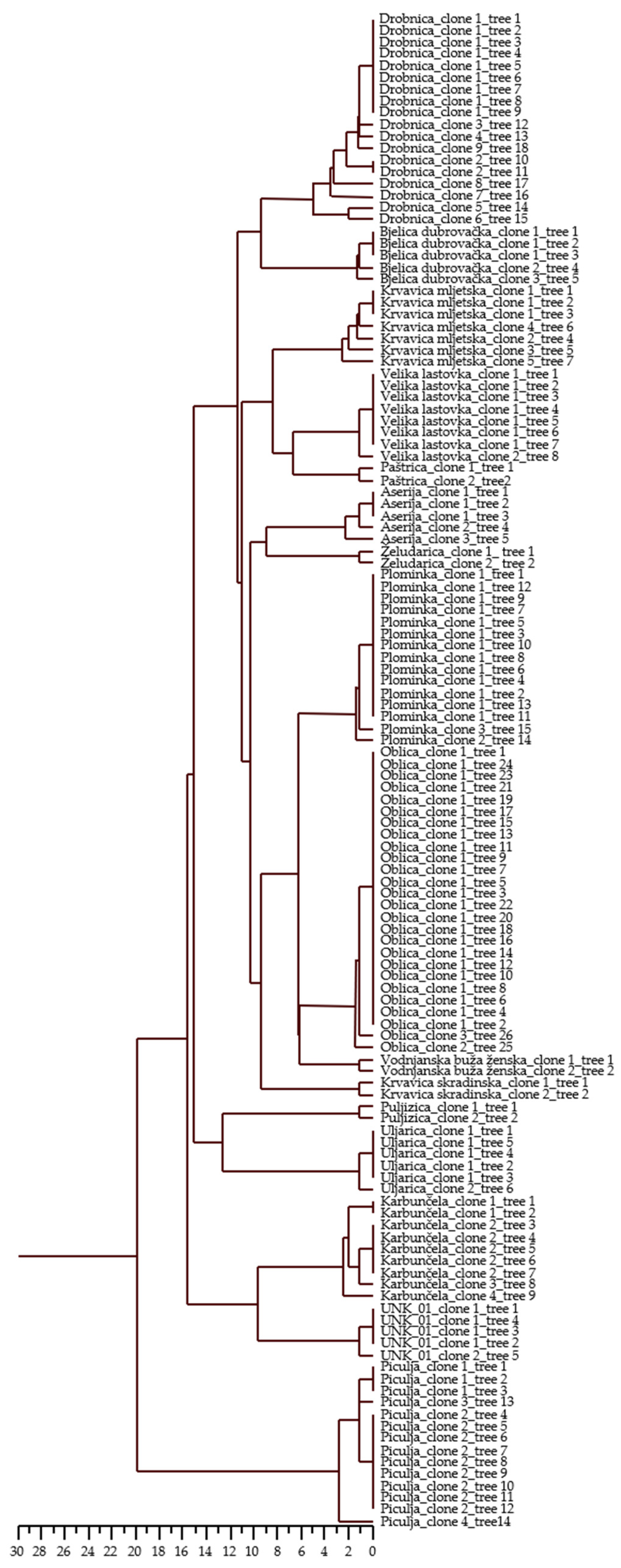
3.2. Genetic Distances and Relationships of Croatian Olive Germplasm
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cipriani, G.; Marrazo, M.T.; Marconi, R.; Cimato, A.; Testolin, R. Microsatellite markers isolated in olive (Olea europaea L.) are suitable for individual fingerprinting and reveal polymorphism within ancient cultivars. Theor. Appl. Genet. 2002, 104, 223–228. [Google Scholar] [CrossRef]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World-The Origin and Spread of Domesticated Plants in South-West Asia, Europe, and the Mediterranean Basin, 4th ed.; Oxford University Press: Oxford, UK, 2012; p. 5. [Google Scholar] [CrossRef]
- Kaniewski, D.; Van Campo, E.; Boiy, T.; Terral, J.F.; Khadari, B.; Besnard, G. Primary domestication and early uses of the emblematic olive tree: Palaeobotanical, historical and molecular evidence from the Middle East. Biol. Rev. 2012, 87, 885–899. [Google Scholar] [CrossRef]
- Besnard, G.; Khadari, B.; Navascués, M.; Fernández-Mazuecos, M.; El Bakkali, A.; Arrigo, N.; Baali-Cherif, D.; Brunini-Bronzini de Caraffa, V.; Santoni, S.; Vargas, P.; et al. The complex history of the olive tree: From Late Quaternary diversification of Mediterranean lineages to primary domestication in the northern Levant. Proc. R. Soc. B. 2013, 280, 20122833. [Google Scholar] [CrossRef]
- Angiolillo, A.; Mencuccini, M.; Baldoni, L. Olive genetic diversity assessed using amplified fragment length polymorphisms. Theor. Appl. Genet. 1999, 98, 411–421. [Google Scholar] [CrossRef]
- Bartolini, G. Olea Databases. 2007. Available online: www.oleadb.it (accessed on 13 July 2023).
- IOC International Olive Council. Available online: https://www.internationaloliveoil.org/the-world-of-olive-oil/ (accessed on 13 July 2023).
- Miljković, I. Suvremeno Voćarstvo; Nakladni zavod Znanje: Zagreb, Hrvatska, 1991; pp. 491–518. [Google Scholar]
- Zec, J. Sortiment maslina u Dalmaciji. Biljn. Proizv. 1951, 1, 3–20. [Google Scholar]
- Croatian Bureau of Statistics. Available online: https://web.dzs.hr/ (accessed on 13 July 2023).
- Šatović, Z.; Liber, Z.; Belaj, A.; Radosavljević, I.; Šindrak, Z.; Benčić, Đ. Genetic diversity of Croatian olive cultivars. In Proceedings of the 46th Croatian and 6th International Symposium on Agriculture, Opatija, Croatia, 14–18 February 2011; Pospišil, M., Ed.; University of Zagreb, Faculty of Agriculture: Zagreb, Croatia, 2011; pp. 98–99. [Google Scholar]
- Ozimec, R.; Karoglan Kontić, J.; Maletić, E.; Matotan, Z.; Strikić, F. Tradicijske Sorte i Pasmine Dalmacije; UNDP Croatia: Zagreb, Croatia, 2015; pp. 90–131. [Google Scholar]
- Bulić, S. Građa za Dalmatinsku Elajografiju; Poljoprivredno odjeljenje Pokrajinske uprave za Dalmaciju: Šibenik, Kingdom of Serbs Croats and Slovenes, 1921; pp. 2–32. [Google Scholar]
- Marčić, M. Uzgoj Maslina na Istočnim Obalama Jadrana; Zadružni savez. Leonova Tiskara: Split, Kingdom of Serbs Croats and Slovenes, 1923; pp. 5–158. [Google Scholar]
- Bakarić, P. Olive cultivars in the Dubrovnik littoral. Pomol. Croat. 1995, 8, 11–29. [Google Scholar]
- Bakarić, P. Sorte Maslina Dubrovačkog Primorja; Alfa 2: Dubrovnik, Croatia, 2002; pp. 7–145. [Google Scholar]
- Bakarić, P. Stare Konavoske Sorte Maslina; Samizdat: Dubrovnik, Croatia, 2005; pp. 5–47. [Google Scholar]
- Bakarić, P. Main olive varieties in the area of Dubrovnik-Neretva County with special reference to autochtonous varieties of the Pelješac peninsula. Pomol. Croat. 2005, 11, 15–21. [Google Scholar]
- Bakarić, P. Autohtone Sorte Maslina Elafita; Samizdat: Dubrovnik, Croatia, 2007; pp. 7–85. [Google Scholar]
- Hugues, C. Elaiografia Istriana; Ceres: Zagreb, Hrvatska, 1999; pp. 7–159. [Google Scholar]
- Belaj, A.; Satovic, Z.; Cipriani, G.; Baldoni, L.; Testolin, R.; Rallo, L.; Trujillo, I. Comparative study of the discriminating capacity of RAPD, AFLP and SSR markers and of their effectiveness in establishing genetic relationships in olive. Theor. Appl. Genet. 2003, 107, 736–744. [Google Scholar] [CrossRef]
- Muzzalupo, I.; Perri, E. Genetic characterizationof olive germplasm by molecular markers. Eur. J. Plant Sci. Biotechnol. 2008, 2, 60–68. [Google Scholar]
- Rafalski, J.A.; Vogel, J.M.; Morgante, M.; Powell, W.; Andre, C.; Tingey, S.V. Generating and using DNA markers in plants. In Nonmammalian Genomic Analysis; Birren, B., Lai, E., Eds.; Academic Press: Boca Raton, FL, USA, 1996; pp. 75–134. [Google Scholar] [CrossRef]
- Kumar, P.; Gupta, V.K.; Misra, A.K.; Modi, D.R.; Pandey, B.K. Potential of molecular markers in plant biotechnology. Plant Omics 2009, 2, 141–162. [Google Scholar]
- Štambuk, S.; Sutlović, D.; Bakarić, P.; Petričević, S.; Anđelinović, Š. Forensic botany: Potential usefulness of microsatellite-based genotyping of Croatian olive (Olea europaea L.) in forensic casework. Croat Med. J. 2007, 48, 556–562. [Google Scholar]
- Poljuha, D.; Sladonja, B.; Bubola, K.B.; Radulović, M.; Brščić, K.; Šetić, E.; Krapac, M.; Milotić, A. A Multidisciplinary Approach to the Characterisation of Autochthonous Istrian Olive (Olea europaea L.) Varieties. Food Technol. Biotechnol. 2008, 46, 347–354. [Google Scholar]
- Poljuha, D.; Sladonja, B.; Šetić, E.; Milotić, A.; Bandelj, D.; Jakše, J.; Javornik, B. DNA fingerprinting of olive varieties in Istria (Croatia) by microsatellite markers. Sci. Hortic. 2008, 115, 223–230. [Google Scholar] [CrossRef]
- Bolarić, S.; Kauf, Z.; Kožarić Silov, G.; Šindrak, Z.; Kozumplik, V.; Benčić, Đ.; Vokurka, A. Morphological and genetic characterization of olive variety of hinterland of Skradin. In Proceedings of the 46th Croatian and 6th International Symposium on Agriculture, Opatija, Croatia, 14–18 February 2011; Pospišil, M., Ed.; University of Zagreb, Faculty of Agriculture: Zagreb, Croatia, 2011; pp. 100–101. [Google Scholar]
- Miljković, I.; Žužić, I.; Pucci, C.; Baldoni, L.; Mariotti, M.; Cultrera, N.G.M. Molecular characterization of an ancient Olea europaea tree located on the Brijuni islands of (Croatia) by SSR markers analysis. Pomol. Croat. 2010, 16, 3–11. [Google Scholar] [CrossRef]
- Ercisli, S.; Bencic, D.; Ipek, A.; Barut, E.; Liber, Z. Genetic relationships among olive (Olea europaea L.) cultivars native to Croatia and Turkey. J. Appl. Bot. Food Qual. 2012, 85, 144–149. [Google Scholar]
- Vokurka, A.; Batelja Lodeta, K.; Bolarić, S.; Hropić, L.; Majić, M.; Benčić, Đ. Determination of genetic variability within the olive variety ‘Piculja’ by molecular and morphological markers. In Proceedings of the 46th Croatian and 16th International Symposium on Agriculture, Vodice, Croatia, 5–11 September 2021; Rozman, V., Antunović, Z., Eds.; University Josip Juraj Strossmayer, Faculty of Agrobiotechnical Sciences: Osijek, Croatia, 2021; pp. 373–377. [Google Scholar]
- Contento, A.; Ceccarelli, M.; Gelati, M.T.; Maggini, F.; Baldoni, L.; Cionini, P.G. Diversity of Olea genotypes and the origin of cultivated olives. Theor. Appl. Genet. 2002, 104, 1229–1238. [Google Scholar] [CrossRef]
- Sari, V.; Baldoni, L.; Porceddu, A.; Cultrera, N.G.M.; Contento, A.; Frediani, M.; Belaj, A.; Trujillo, I.; Cionini, P.G. Microsatellite markers are powerful tools for discriminating among olive cultivars and assigning them to geographically defined populations. Genome 2006, 49, 1606–1615. [Google Scholar] [CrossRef]
- Muzzalupo, I.; Perri, E. Genetic diversity in olive tree cultivars from Italy and other countries of the Mediterranean basin as revealed by RAPD and SSR molecular marker. Adv. Hortic. Sci. 2009, 23, 263–275. [Google Scholar]
- Lazović, B.; Adakalić, M.; Pucci, C.; Perović, T.; Bandelj, D.; Belaj, A.; Mariotti, R.; Baldoni, L. Characterizing ancient and local olive germplasm from Montenegro. Sci. Hortic. 2016, 209, 117–123. [Google Scholar] [CrossRef]
- Haouane, H.; El Bakkali, A.; Moukhli, A.; Tollon, C.; Santoni, S.; Oukabli, A.; Al Modafar, C.; Khadari, B. Genetic structure and core collection of the World Olive Germplasm Bank of Marrakech, towards the optimised management and use of Mediterranean olive genetic resources. Genetica 2011, 139, 1083–1094. [Google Scholar] [CrossRef]
- Trujillo, I.; Ojeda, M.A.; Urdiroz, N.M.; Potter, D.; Barranco, D.; Rallo, L.; Díez, C.M. Identification of the Worldwide Olive Germplasm Bank of Córdoba (Spain) using SSR and morphological markers. Tree Genet. Genomes 2013, 10, 141–155. [Google Scholar] [CrossRef]
- El Bakkali, A.; Essalouh, L.; Tollon, C.; Rivallan, R.; Mournet, P.; Moukhli, A.; Zaher, H.; Mekkaoui, A.; Hadidou, A.; Sikaoui, L.; et al. Characterization of Worldwide Olive Germplasm Banks of Marrakech (Morocco) and Córdoba (Spain): Towards management and use of olive germplasm in breeding programs. PLoS ONE 2019, 14, e0223716. [Google Scholar] [CrossRef] [PubMed]
- Muzzalupo, I.; Muto, A.; Badolati, G.; Veizi, A.; Chiappetta, A. Genotyping of Albania olive (Olea europaea) germplasm by SSR molecular marker. Emir. J. Food Agric. 2018, 30, 573–580. [Google Scholar] [CrossRef]
- Dervishi, A.; Jakše, J.; Ismaili, H.; Javornik, B.; Štajner, N. Genetic structure and core collection of olive germplasm from Albania revealed by microsatellite markers. Genes 2021, 12, 256. [Google Scholar] [CrossRef] [PubMed]
- Carriero, E.; Fontanazza, G.; Cellini, F.; Giorio, G. Identification of simple sequence repeats (SSRs) in olive (Olea europaea L.). Theor. Appl. Genet. 2002, 104, 301–307. [Google Scholar] [CrossRef]
- Muzzalupo, I.; Stefanizzi, F.; Salimonti, A.; Falabella, R.; Perri, E. Microsatellite markers for identification of a group of italian olive accessions. Sci. Agric. 2009, 66, 685–690. [Google Scholar] [CrossRef][Green Version]
- Muzzalupo, I.; Vendramin, G.G.; Chiapetta, A. Genetic biodiversity of Italian olives (Olea europaea) germplasm analyzed by SSR markers. Sci. World J. 2014, 2014, 296590. [Google Scholar] [CrossRef] [PubMed]
- Ercisli, S.; Ipek, A.; Barut, E. SSR Marker-Based DNA Fingerprinting and Cultivar Identification of Olives (Olea europaea). Biochem. Genet. 2011, 49, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Işık, N.; Doğanlar, S.; Frary, A. Genetic Diversity of Turkish Olive Varieties Assessed by Simple Sequence Repeat and Sequence-Related Amplified Polymorphism Markers. Crop Sci. 2011, 51, 1646–1654. [Google Scholar] [CrossRef]
- Ipek, A.; Barut, E.; Gulen, H.; Ipek, M. Assessment of inter- and intra-cultivar variations in olive using SSR markers. Sci. Agric. 2012, 69, 327–335. [Google Scholar] [CrossRef]
- Unver, H.; Sakar, E.; Ulas, M.; Ercisli, S.; Bekir Erol, A.K. Molecular characterization of indigenous olive genotypes based on SSR analysis. Genetika 2016, 48, 1017–1025. [Google Scholar] [CrossRef]
- Roubos, K.; Moustakas, M.; Aravanopoulos, F.A. Molecular identification of Greek olive (Olea europaea) cultivars based on microsatellite loci. Genet. Mol. Res. 2010, 9, 1865–1876. [Google Scholar] [CrossRef] [PubMed]
- Noormohammadi, Z.; Hosseini-Mazinani, M.; Trujillo, I.; Rallo, L.; Belaj, A.; Sadeghizadeh, M. Identification and classification of main Iranian olive cultivars using microsatellite markers. HortScience 2007, 42, 1545–1550. [Google Scholar] [CrossRef]
- Rony, C.; Baalbaki, R.; Kalaitzis, P.; Talhouk, S.N. Molecular characterization of Lebanese olive germplasm. Tree Genet. Genomes 2009, 5, 109–115. [Google Scholar] [CrossRef]
- Rekik, I.; Salimonti, A.; Kamoun, N.G.; Muzzalupo, I.; Lepais, O.; Gerber, S.; Perri, E.; Rebai, A. Characterization and Identification of Tunisian Olive Tree Varieties by Microsatellite Markers. HortScience 2008, 43, 1371–1376. [Google Scholar] [CrossRef]
- Fendri, M.; Trujillo, I.; Trigui, A.; Rodrıguez-Garcıa, M.I.; Ramırez, J.D.A. Simple Sequence Repeat Identification and Endocarp haracterization of Olive Tree Accessions in a Tunisian Germplasm Collection. HortScience 2010, 45, 1429–1436. [Google Scholar] [CrossRef]
- Abdelhamid, S.; Omri, A.; Grati-Kamoun, N.; Paolo Marra, F.; Caruso, T. Molecular characterization and genetic relationships of cultivated Tunisian olive varieties (Olea europaea L.) using SSR markers. J. New Sci. Agric. Biotechnol. 2017, 40, 2175–2185. [Google Scholar]
- Abdessemed, S.; Muzzalupo, I.; Benbouza, H. Assessment genetic diversity among Algerian olive (Olea europaea L) cultivars using SSR marker. Sci. Hortic. 2015, 192, 10–20. [Google Scholar] [CrossRef]
- Obaid, R.; Abu-Qaoud, H.; Arafeh, R. Molecular characterization of three common olive (Olea europaea L.) cultivars in Palestine, using simple sequence repeat (SSR) markers. Biotechnol. Biotechnol. Equip. 2014, 28, 813–817. [Google Scholar] [CrossRef]
- Sakar, E.; Unver, H.; Ercisli, S. Genetic Diversity Among Historical Olive (Olea europaea L.) Genotypes from Southern Anatolia Based on SSR Markers. Biochem Genet. 2016, 54, 842–853. [Google Scholar] [CrossRef]
- La Mantia, M.; Lain, O.; Caruso, T.; Testolin, R. SSR-based DNA fingerprints reveal the genetic diversity of Sicilian olive (Olea europaea L.) germplasm. J. Hortic. Sci. Biotechnol. 2005, 80, 628–632. [Google Scholar] [CrossRef]
- Las Casas, G.; Scollo, F.; Distefano, G.; Continella, A.; Gentile, A.; La Malfa, S. Molecular characterization of olive (Olea europaea L.) Sicilian cultivars using SSR markers. Biochem. Syst. Ecol. 2014, 57, 15–19. [Google Scholar] [CrossRef]
- Alba, V.; Montemurro, C.; Sabetta, W.; Pasqualone, A.; Blanco, A. SSR-based identification key of cultivars of Olea europaea L. diffused in Southern-Italy. Sci. Hortic. 2009, 123, 11–16. [Google Scholar] [CrossRef]
- Marra, F.P.; Caruso, T.; Costa, F.; Di Vaio, C.; Mafrica, R.; Marchese, A. Genetic relationships, structure and parentage simulation among the olive tree (Olea europaea L. subsp. europaea) cultivated in Southern Italy revealed by SSR markers. Tree Genet. Genomes. 2013, 9, 961–973. [Google Scholar] [CrossRef]
- Díez, C.M.; Trujillo, I.; Barrio, E.; Belaj, A.; Barranco, D.; Rallo, L. Centennial olive trees as a reservoir of genetic diversity. Ann. Bot. 2011, 108, 797–807. [Google Scholar] [CrossRef]
- Brito, G.; Loureiro, J.; Lopes, T.; Rodriguez, E.; Santos, C. Genetic characterisation of olive trees from Madeira Archipelago using flow cytometry and microsatellite markers. Genet. Resour. Crop Evol. 2008, 55, 657–664. [Google Scholar] [CrossRef]
- Bandelj, D.; Jakše, J.; Javornik, B. DNA fingerprinting of olive varieties by microsatellite markers. Food Technol. Biotechnol. 2002, 40, 185–190. [Google Scholar]
- Bandelj, D.; Jakše, J.; Javornik, B. Assessment of genetic variability of olive varieties by microsatellite and AFLP markers. Euphytica 2004, 136, 93–102. [Google Scholar] [CrossRef]
- Mousavi, S.; Mariotti, R.; Regni, L.; Nasini, L.; Bufacchi, M.; Pandolfi, S.; Baldoni, L.; Proietti, P. The first molecular identification of an olive collection applying standard simple sequence repeats and novel expressed sequence Tag markers. Front. Plant Sci. 2017, 8, 1283. [Google Scholar] [CrossRef]
- di Rienzo, V.; Sion, S.; Taranto, F.; D’Agostino, N.; Montemurro, C.; Fanelli, V.; Sabetta, W.; Boucheffa, S.; Tamendjari, A.; Pasqualone, A.; et al. Genetic flow among olive populations within the Mediterranean basin. PeerJ 2018, 6, e5260. [Google Scholar] [CrossRef]
- Gomes, S.; Martins-Lopes, P.; Lopes, J.; Guedes-Pinto, H. Assessing Genetic Diversity in Olea europaea L. Using ISSR and SSR Markers. Plant. Mol. Biol. Rep. 2009, 27, 365–373. [Google Scholar] [CrossRef]
- Belaj, A.; Cipriani, G.; Testolin, R.; Rallo, L.; Trujillo, I. Characterization and identification of the main Spanish and Italian olive cultivars by simple-sequence repeat markers. HortScience 2004, 39, 1557–1561. [Google Scholar] [CrossRef]
- Doveri, S.; Gil, F.S.; Dıaz, A.; Reale, S.; Busconi, M.; Machado, A.D.; Martın, A.; Fogher, C.; Donini, P.; Lee, D. Standardization of a set of microsatellite markers for use in cultivar identification studies in olive (Olea europaea L.). Sci. Hortic. 2008, 116, 367–373. [Google Scholar] [CrossRef]
- Baldoni, L.; Cultrera, N.G.; Mariotti, R.; Riccioloni, C.; Arcioni, S.; Vendramin, G.G.; Buonamici, A.; Porceddu, A.; Sarri, V.; Ojeda, M.A.; et al. A consensus list of microsatellites markers for olive genotyping. Mol. Breed. 2009, 24, 213–231. [Google Scholar] [CrossRef]
- Bracci, T.; Busconi, M.; Fogher, C.; Sebastiani, L. Molecular studies in olive (Olea europaea L.): Overview on DNA markers applications and recent advances in genome analysis. Plant Cell Rep. 2011, 30, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Veloso, M.M.; Simões-Costa, M.C.; Carneiro, L.C.; Guimarães, J.B.; Mateus, C.; Fevereiro, P.; Pinto-Ricardo, C. Olive tree (Olea europaea L.) diversity in traditional small farms of Ficalho, Portugal. Diversity 2018, 10, 5. [Google Scholar] [CrossRef]
- Hosseini-Mazinani, M.; Mariotti, R.; Torkzaban, B.; Sheikh-Hassani, M.; Ataei, S.; Cultrera, N.G.; Pandolfi, S.; Baldoni, L. High genetic diversity detected in olives beyond the boundaries of the Mediterranean Sea. PLoS ONE 2014, 9, e93146. [Google Scholar] [CrossRef]
- Zaher, H.; Boulouha, B.; Baaziz, M.; Sikaoui, L.; Gaboun, F.; Udupa, S.M. Morphological and genetic diversity in olive (Olea europaea subsp. europaea L.) clones and varieties. Plant Omics 2011, 4, 370–376. [Google Scholar]
- Muzzalupo, I.; Russo, A.; Chuappetta, A.A.; Benincasa, C.; Perri, E. Evaluation of genetic diversity in Italian olives (Olea europaea L.) cultivars by SSR markers. J. Biotechnol. 2010, 150, 472–480. [Google Scholar] [CrossRef]
- Noormohammadi, Z.; Hosseini-Mazinani, M.; Trujillo, I.; Belaj, A. Study of intracultivar variation among main Iranian olive cultivars using SSR markers. Acta. Biol. Szeged 2009, 53, 27–32. [Google Scholar]
- Lazović, B.; Klepo, T.; Adakalić, M.; Šatović, Z.; Baruca Arbeiter, A.; Hladnik, M.; Strikić, F.; Liber, Z.; Bandelj, D. Intra-varietal variability and genetic relationships among the homonymic East Adriatic olive (Olea europaea L.) varieties. Sci. Hortic. 2018, 236, 175–185. [Google Scholar] [CrossRef]
- Sefc, K.M.; Lopes, M.S.; Mendonça, D.; Rodrigues Dos Santos, M.; Laimer Da Câmara Machado, M.; Da Câmara Machado, A. Identification of microsatellite loci in olive (Olea europaea) and their characterization in Italian and Iberian olive trees. Mol. Ecol. 2000, 9, 1171–1193. [Google Scholar] [CrossRef]
- De La Rosa, R.; James, C.; Tobutt, K.R. Isolation and characterization of polymorphic microsatellite in olive Olea europaea L. and their transferability to other genera in the Oleaceae. Mol. Ecol. Notes 2002, 2, 265–267. [Google Scholar] [CrossRef]
- Arnaud-Haond, S.; Belkhir, K. Genclone: A computer program to analyse genotypic data, test for clonality and describe spatial clonal organization. Mol. Ecol. Notes 2007, 7, 7–11. [Google Scholar] [CrossRef]
- Nei, M. Analysis of Gene Diversity in Subdivided Populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed]
- Botstein, D.; White, R.L.; Sholnick, M.; David, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Paetkau, D.; Calvert, W.; Stirling, I.; Strobeck, C. Microsatellite analysis of population structure in Canadian polar bears. Mol. Ecol. 1995, 4, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.W.; Sefc, K.M. Identity 1.0; Centre for Applied Genetics, University of Agricultural Sciences: Vienna, Austria, 1999. [Google Scholar]
- Kloosterman, A.D.; Budowe, B.; Riley, E.L. Population data of the HLA DQα locus in Dutch caucasians. Int. J. Leg. Med. 1993, 105, 233–238. [Google Scholar] [CrossRef]
- Bowcock, A.M.; Ruiz-Linarez, A.; Tomfhorde, J.; Minch, E.; Kidd, J.R.; Cavalli-Sforza, L.L. High resolution human evolutionary trees with polamorphic microsatellites. Nature 1994, 368, 455–457. [Google Scholar] [CrossRef]
- Minch, E.; Ruiz-Linares, A.; Goldstein, D.; Feldman, M.; Cavalli-Sforza, L.L. MICROSAT: A Computer Program for Calculating Various Statistics on Microsatellite Allele Data, Ver.1.5; Stanford University Medical Center: Stanford, CA, USA, 1996; Available online: https://hpgl.stanford.edu/projects/microsat/microsat.html (accessed on 13 December 2023).
- Fitch, W.M.; Margoliash, E. Construction of phylogenetic trees: A method based on mutation distances as estimated from cytochrome C sequences is of general applicability. Science 1967, 155, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. PHYLIP-Phylogeny Inference Package (Version 3.2). Cladistics 1989, 5, 164–166. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, F.J. Exeter Software NTSYS-Pc: Numerical Taxonomy and Multivariate Analysis System; Applied Biostatistics, Inc. Exeter Software: Setauket, NY, USA, 2009. [Google Scholar]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Dent, E.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y.; Vigouroux, Y.; Goodman, M.M.; Sanchez, J.G.; Buckler, E.; Doebley, J. A single domestication for maize shown by multilocus microsatellite genotyping. Proc. Natl. Acad. Sci. USA 2002, 99, 6080–6084. [Google Scholar] [CrossRef] [PubMed]
- Angiolillo, A.; Reale, S.; Pilla, F.; Baldoni, L. Molecular analysis of olive cultivars in the Molise region of Italy. Genet. Resour. Crop Evol. 2006, 53, 289–295. [Google Scholar] [CrossRef]
- Corrado, G.; La Mura, M.; Ambrosino, O.; Pugliano, G.; Varricchio, P.; Rao, R. Relationships of Campanian olive cultivars: Comparative analysis of molecular and phenotypic data. Genome 2009, 52, 692–700. [Google Scholar] [CrossRef]
- Halkett, F.; Simon, J.C.; Balloux, F. Tackling the population genetics of clonal and partially clonal organisms. Trends Ecol. Evol. 2005, 20, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Dervishi, A.; Jakše, J.; Ismaili, H.; Javornik, B.; Štajner, N. Comparative assessment of genetic diversity in Albanian olive (Olea europaea L) using SSRs from anonymous and transcribed genomic regions. Tree Genet. Genomes 2018, 14, 53. [Google Scholar] [CrossRef]
- Strikić, F.; Gugić, J.; Klepo, T. Stanje hrvatskog maslinarstva. Glas. Zaštite Bilja 2012, 12, 271–276. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Reference | Abbreviated Primer Name | Dye |
|---|---|---|---|
| ssrOeUA-DCA3 | [78] | DCA3 | 6FAM |
| ssrOeUA-DCA5 | [78] | DCA5 | NED |
| ssrOeUA-DCA9 | [78] | DCA9 | VIC |
| ssrOeUA-DCA14 | [78] | DCA14 | NED |
| ssrOeUA-DCA16 | [78] | DCA16 | 6FAM |
| ssrOeUA-DCA18 | [78] | DCA18 | NED |
| UDO099-043 | [1] | UDO43 | VIC |
| GAPU-71B | [41] | GAPU71B | 6FAM |
| GAPU-101 | [41] | GAPU101 | 6FAM |
| GAPU-103A | [41] | GAPU103A | VIC |
| EMO90 | [79] | EMO90 | VIC |
| SSR Loci | Range of Allele Size | Na | Ne | Nu | NDG | Ho | He | PIC | F | PI |
|---|---|---|---|---|---|---|---|---|---|---|
| DCA3 | 232–257 | 12 | 5.10 | 3 | 21 | 0.911 | 0.804 | 0.78 | −0.133 | 0.065 |
| DCA5 | 194–214 | 10 | 2.57 | 2 | 19 | 0.681 | 0.611 | 0.56 | −0.115 | 0.210 |
| DCA9 | 162–208 | 16 | 7.20 | 3 | 42 | 0.793 | 0.861 | 0.86 | 0.080 | 0.034 |
| DCA14 | 171–193 | 9 | 2.62 | 1 | 18 | 0.622 | 0.619 | 0.58 | −0.006 | 0.019 |
| DCA16 | 124–182 | 18 | 5.19 | 7 | 34 | 0.756 | 0.807 | 0.78 | 0.064 | 0.059 |
| DCA18 | 159–199 | 14 | 4.52 | 2 | 33 | 0.822 | 0.779 | 0.75 | −0.056 | 0.078 |
| UDO43 | 168–216 | 19 | 5.98 | 6 | 38 | 0.985 | 0.833 | 0.82 | −0.183 | 0.044 |
| GAPU71B | 119–148 | 9 | 4.13 | 3 | 19 | 0.837 | 0.758 | 0.73 | −0.104 | 0.091 |
| GAPU101 | 182–219 | 9 | 5.85 | 1 | 28 | 0.919 | 0.829 | 0.81 | −0.108 | 0.051 |
| GAPU103A | 136–190 | 17 | 5.75 | 6 | 35 | 0.881 | 0.826 | 0.81 | −0.067 | 0.053 |
| EMO90 | 186–198 | 5 | 1.36 | - | 11 | 0.844 | 0.701 | 0.65 | −0.205 | 0.140 |
| Mean | 12.5 | 4.75 | 3 | 27 | 0.823 | 0.766 | 0.74 | −0.076 | ||
| Total | 138 | 34 | ||||||||
| Combined | 5.8 × 10−13 |
| Genotype | SSR Loci | Rare Alleles (bp) |
|---|---|---|
| Drobnica_clone 5 | GAPU71B | 134 * |
| Drobnica_clone 8 | UDO43 | 202 |
| Drobnica_clone 9 | GAPU103A | 166 * |
| Dužica | DCA16 | 180 |
| GAPU71B | 119 * | |
| GAPU103A | 181 | |
| Karbona | DCA16 | 182 |
| Krvavica mljetska_clone 5 | UDO43 | 206 |
| Oblica_clone 2 | DCA16 | 178 |
| Plominka_clone 2 | GAPU101 | 209 |
| Puljizica_clone 1 | DCA14 | 193 |
| UNK_01_clone 1 | DCA9 | 198 |
| UNK_23 | GAPU71B | 148 |
| UNK_26 | UDU43 | 200 |
| UNK_28 | DCA3 | 257 |
| UNK_32 | DCA9 | 178 * |
| GAPU103A | 161 * | |
| UNK_36 | DCA16 | 168 * |
| UNK_39 | DCA9 | 180 |
| UNK_40 | GAPU103A | 170 |
| UNK_41 | DCA5 | 200 |
| DCA16 | 172 | |
| UNK_47 | DCA16 | 162 * |
| UNK_48 | DCA3 | 247 |
| DCA5 | 204 | |
| DCA16 | 152 * | |
| DCA18 | 191 * | |
| UDO43 | 186 | |
| UNK_49 | UDO43 | 190 * |
| UDO43 | 192 * | |
| UNK_56 | DCA3 | 241 |
| Žabarka | DCA18 | 197 * |
| Želudarica_clone 2 | GAPU103A | 188 |
| Uljarica_clone 2 | GAPU103A | 164 |
| No. | The Common Name of the Variety | Site | Number of Trees | Synonyms for the Location Where Samples Were Taken 1 |
|---|---|---|---|---|
| 1 | ‘Aserija’ | Zadar | 4 | Galka |
| Ugljan | 1 | Tvrđaka | ||
| 2 | ‘Bjelica dubrovačka’ | Dubrovnik | 1 | Žutica |
| Mljet | 2 | Bjelica | ||
| Dubrovnik | 1 | - | ||
| 3 | ‘Buhavica’ | Lastovo | 1 | - |
| 4 | ‘Buža puntoža’ | Southern Istria | 2 | - |
| 5 | ‘Drobnica’ | Pašman | 1 | Krstunjača |
| Dubrovnik | 2 | Sitnica | ||
| Dugi Otok | 4 | - | ||
| Cres | 1 | Rošulja | ||
| Korčula | 1 | - | ||
| Krk | 1 | Naška | ||
| Ugljan | 2 | - | ||
| Northern Istria | 1 | Starinska bilica, Brambolina | ||
| Dubrovnik | 1 | Česvinka | ||
| 6 | ‘Duška’ | Murter | 3 | - |
| 7 | ‘Dužica’ | Dugi Otok | 1 | Lavdara |
| Mljet | 1 | - | ||
| Dubrovnik | 1 | - | ||
| 8 | ‘Grozdenjača’ | Mljet | 1 | - |
| 9 | ‘Istarska bjelica’ | Istria | 1 | - |
| 10 | ‘Jeruzalemka’ | Dubrovnik | 2 | - |
| 11 | ‘Kamasa’ | Koločep | 1 | - |
| 12 | ‘Karbona’ | Istria | 1 | Buva, Črna, Karbonaca |
| 13 | ‘Karbunčela’ | Dugi Otok | 6 | Grambučela, Krombunčela |
| 14 | ‘Kosmača’ | Dubrovnik | 1 | - |
| 15 | ‘Krvavica mljetska’ | Mljet | 7 | Puntoguza |
| 16 | ‘Krvavica skradinska’ | Šibenik (the area of Skradin) | 1 | - |
| 17 | ‘Lastovka’ | Lastovo | 1 | - |
| 18 | ‘Levantinka’ | Lastovo | 1 | - |
| 19 | ‘Mezanica’ | Dubrovnik | 1 | - |
| 20 | ‘Mrčakinja’ | Dubrovnik | 1 | - |
| 21 | ‘Murgulja’ | Dubrovnik | 1 | Krvavica |
| Dubrovnik | 1 | - | ||
| Mljet | 2 | Šorgula | ||
| 22 | ‘Oblica’ | Zadar | 2 | - |
| Dubrovnik | 1 | Lumbardeška | ||
| Dugi Otok | 3 | Orkula | ||
| Cres | 4 | Slatka, Mastrinka, Istrijanka | ||
| Krk | 1 | - | ||
| Mljet | 2 | Balunjača | ||
| Murter | 1 | - | ||
| Ugljan | 3 | Orkula, Orkula kaljska | ||
| Kornati | 3 | - | ||
| Zadar | 2 | Orkula | ||
| 23 | ‘Oštrica’ | Ugljan | 1 | - |
| 24 | ‘Pačica’ | Koločep | 1 | Kalamata |
| Mljet | 2 | - | ||
| Šipan | 2 | Šljivača, Lituša | ||
| 25 | ‘Paštrica’ | Dubrovnik | 2 | - |
| 26 | ‘Piculja’ | Lastovo | 7 | - |
| Mljet | 4 | - | ||
| Šipan | 3 | - | ||
| 27 | ‘Plominka’ | Dubrovnik | 2 | Crna dubrovačka, Crnica |
| Cres | 2 | Žižolača | ||
| Krk | 2 | - | ||
| Northern Istria | 3 | Buža puntoža, Črnica, Marana | ||
| Southern Istria | 1 | Buža muška | ||
| 28 | ‘Puljizica’ | Šipan | 2 | Krivuja |
| 29 | ‘Puljka’ | Dugi Otok | 2 | - |
| 30 | ‘Resulja’ | Mljet | 1 | - |
| 31 | ‘Rosulja’ | Krk | 1 | - |
| 32 | ‘Slivnjača’ | Cres | 3 | Simjača |
| 33 | ‘Šljivovica’ | Šibenik (the region of Skradin) | 1 | Šljivarica |
| 34 | ‘Uljarica’ | Dubrovnik | 3 | - |
| Dubrovnik | 1 | Zuzorka | ||
| Šipan | 1 | Mezanica | ||
| Dubrovnik | 1 | Zuzorka | ||
| 35 | ‘Velika lastovka’ | Kornati | 3 | Duška |
| Dubrovnik | 1 | - | ||
| 36 | ‘Vodnjanska buža ženska’ | Southern Istria | 1 | - |
| 37 | ‘Vrhuljača’ | Dugi Otok | 1 | Puljižica |
| Murter | 2 | - | ||
| Kornati | 1 | - | ||
| 38 | ‘Žabarka’ | Dubrovnik | 1 | - |
| 39 | ‘Želudarica’ | Dubrovnik | 2 | Dubravka |
| 40 | ‘Žižolera’ | Southern Istria | 1 | - |
| Genotype | No. of Trees (No. of SSR Profiles) | SSR Loci | Allele Combinations (bp); (in Brackets: The Numbers of Trees with the Same Allele Combination) |
|---|---|---|---|
| Drobnica | 18 (9) | DCA9 | 176/204 (1), 184/204 (1), 204/204 (14), 206/206 (2) |
| DCA14 | 179/191 (1), 187/191 (2), 191/191 (15) | ||
| UDO43 | 172/112 (1), 176/212 (15), 176/214 (1), 202/210 (1) | ||
| Gapu71B | 124/130 (17), 124/134 (1) | ||
| Gapu103A | 159/166 (1), 159/174 (13), 159/177 (2), 159/179 (2) | ||
| Krvavica mljetska | 7 (5) | DCA9 | 162/162 (1), 162/206 (6) |
| DCA16 | 126/154 (6), 126/156 (1) | ||
| DCA18 | 173/181 (2), 173/185 (5) | ||
| UDO43 | 174/212 (5), 174/214 (1), 176/206 (1) | ||
| Piculja | 14 (4) | DCA9 | 172/172 (1), 172/196 (4), 172/204 (9) |
| DCA18 | 171/175 (1), 171/177 (13) | ||
| UDO43 | 178/180 (1), 180/182 (13) | ||
| Karbunčela | 9 (4) | DCA9 | 182/184 (1), 184/202 (2), 184/206 (6) |
| DCA18 | 179/185 (8), 179/187 (1) | ||
| UDO43 | 176/210 (2), 176/212 (1), 176/216 (6) | ||
| Aserija | 5 (3) | DCA9 | 162/194 (1), 162/204 (3), 162/206 (1) |
| UDO43 | 176/208 (1), 176/210 (4) | ||
| Plominka | 15 (3) | DCA3 | 239/249 (14), 239/251 (1) |
| GAPU101 | 191/209 (1), 191/219 (14) | ||
| Oblica | 26 (3) | DCA9 | 162/192 (1), 162/204 (25) |
| DCA16 | 150/174 (25), 150/178 (1) | ||
| Bjelica dubrovačka | 5 (3) | DCA9 | 172/204 (1), 204/204 (4) |
| UDO43 | 174/176 (1), 176/178 (4) | ||
| Velika lastovka | 8 (2) | DCA9 | 162/204 (7), 162/208 (1) |
| Uljarica | 6 (2) | Gapu103A | 157/164 (1), 157/174 (5) |
| UNK_01 | 5 (2) | DCA9 | 198/206 (1), 206/206 (4) |
| Paštrica | 2 (2) | DCA9 | 162/196 (1), 162/204 (1) |
| Krvavica skradinska | 2 (2) | DCA9 | 194/204 (1), 194/206 (1) |
| Vodnjanska buža ženska | 2 (2) | UDO43 | 174/210 (1), 174/212 (1) |
| Puljizica | 2 (2) | DCA14 | 173/191 (1), 173/193 (1) |
| Želudarica | 2 (2) | GAPU103A | 150/186 (1), 150/188 (1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolarić, S.; Vokurka, A.; Batelja Lodeta, K.; Benčić, Đ. Genotyping of Croatian Olive Germplasm with Consensus SSR Markers. Horticulturae 2024, 10, 417. https://doi.org/10.3390/horticulturae10040417
Bolarić S, Vokurka A, Batelja Lodeta K, Benčić Đ. Genotyping of Croatian Olive Germplasm with Consensus SSR Markers. Horticulturae. 2024; 10(4):417. https://doi.org/10.3390/horticulturae10040417
Chicago/Turabian StyleBolarić, Snježana, Aleš Vokurka, Kristina Batelja Lodeta, and Đani Benčić. 2024. "Genotyping of Croatian Olive Germplasm with Consensus SSR Markers" Horticulturae 10, no. 4: 417. https://doi.org/10.3390/horticulturae10040417
APA StyleBolarić, S., Vokurka, A., Batelja Lodeta, K., & Benčić, Đ. (2024). Genotyping of Croatian Olive Germplasm with Consensus SSR Markers. Horticulturae, 10(4), 417. https://doi.org/10.3390/horticulturae10040417