
Mandarin Variety Significantly Affects the Metabolites Present in the Leaves

 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Metabolomics Analysis of Leaves by 1H NMR
2.3. Statistical Analysis
3. Results and Discussion
3.1. Analysis of Variance by ANOVA
3.1.1. Amino Acids
3.1.2. Organic Acids
3.1.3. Sugars
3.1.4. Other Metabolites
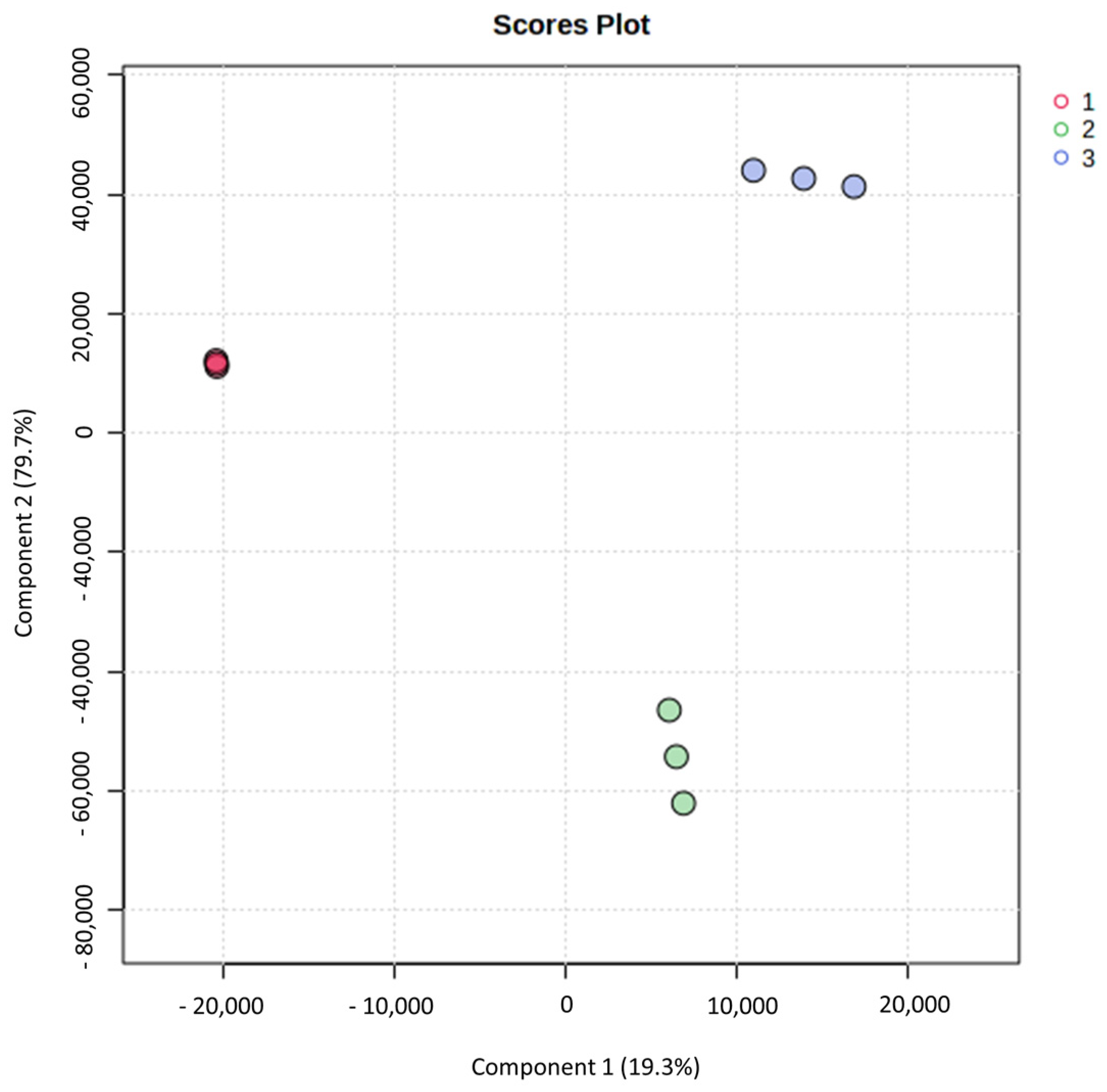
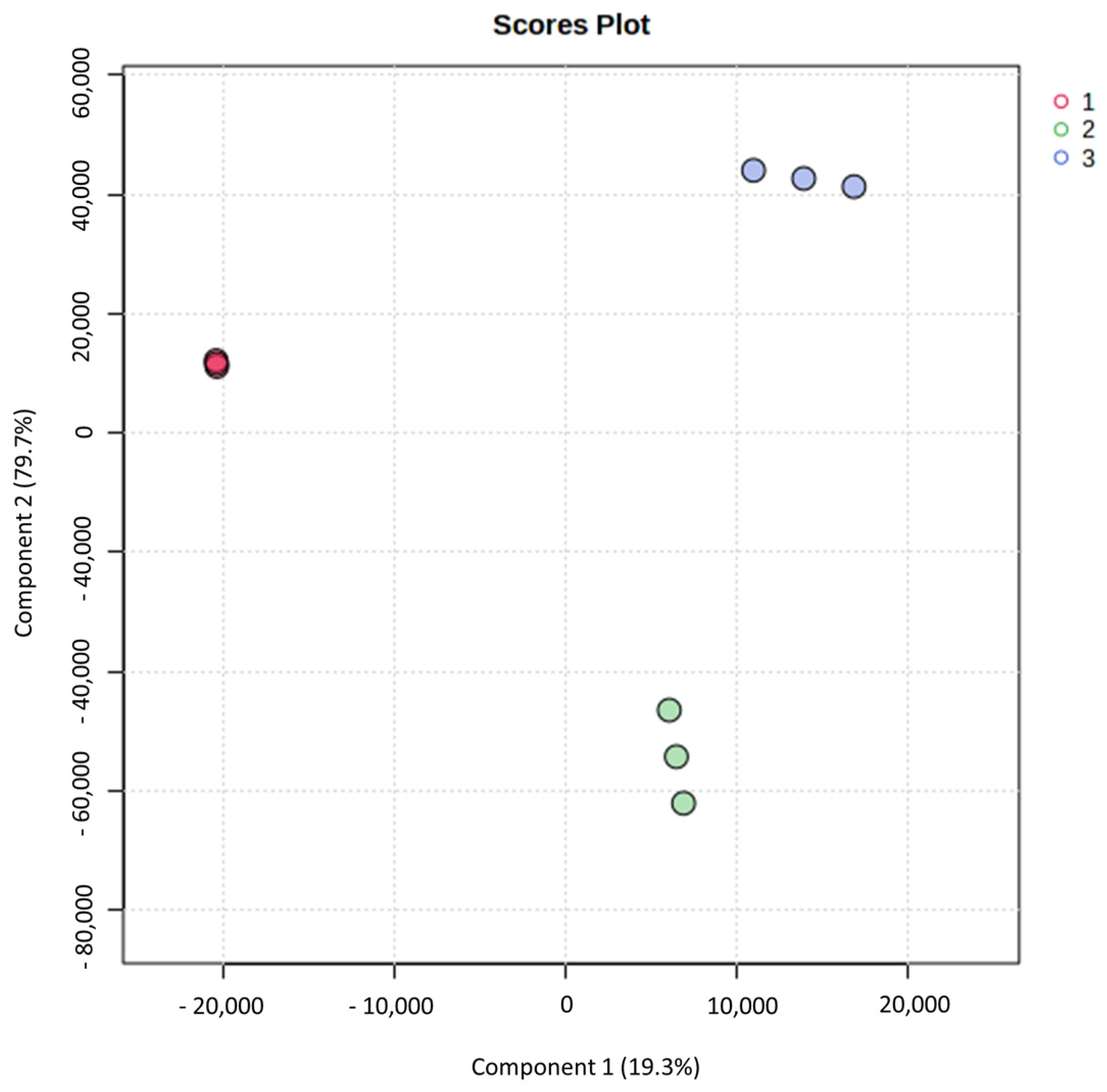
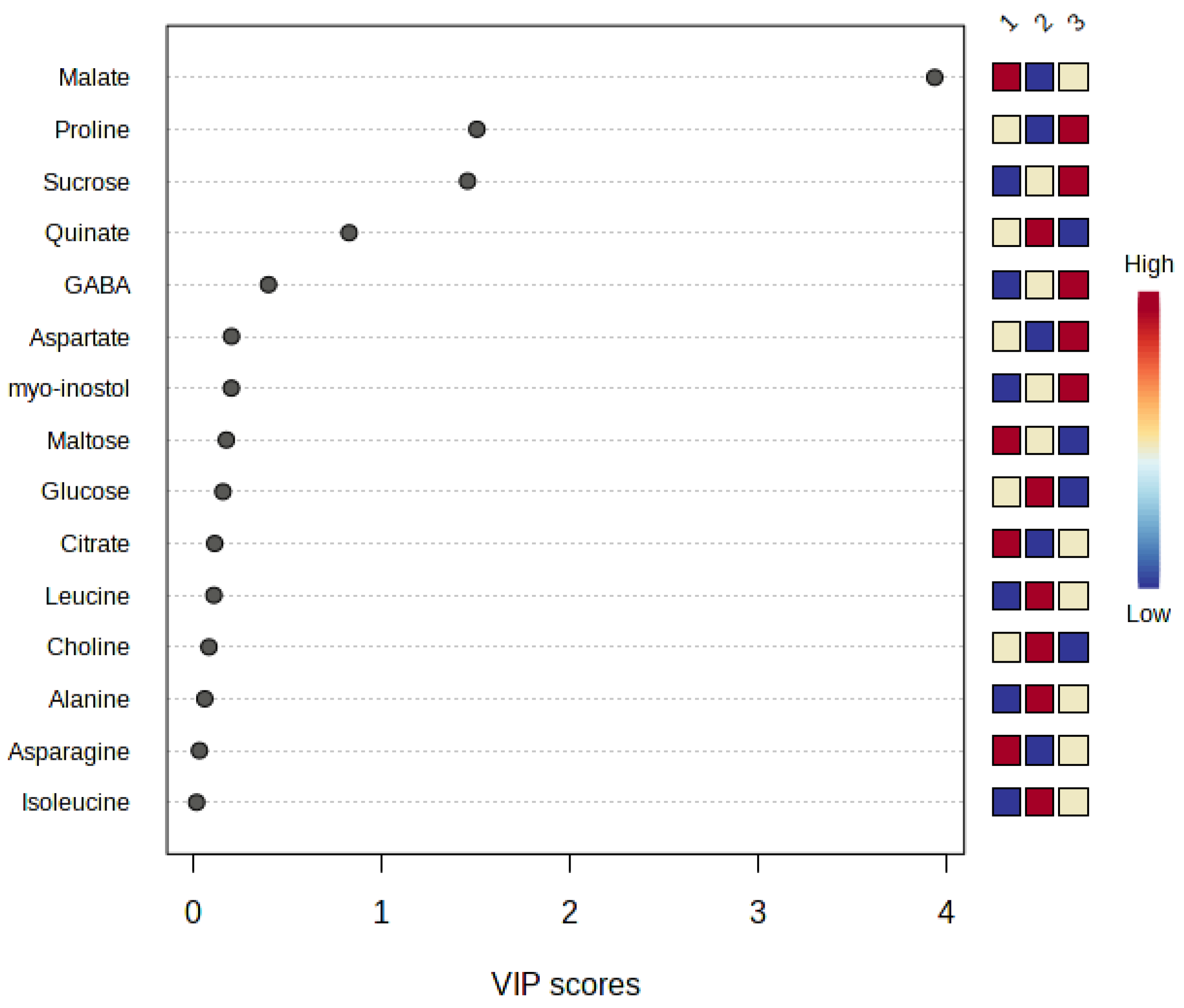
3.2. Principal Component Analysis and Partial Least Squares-Discriminant Analysis
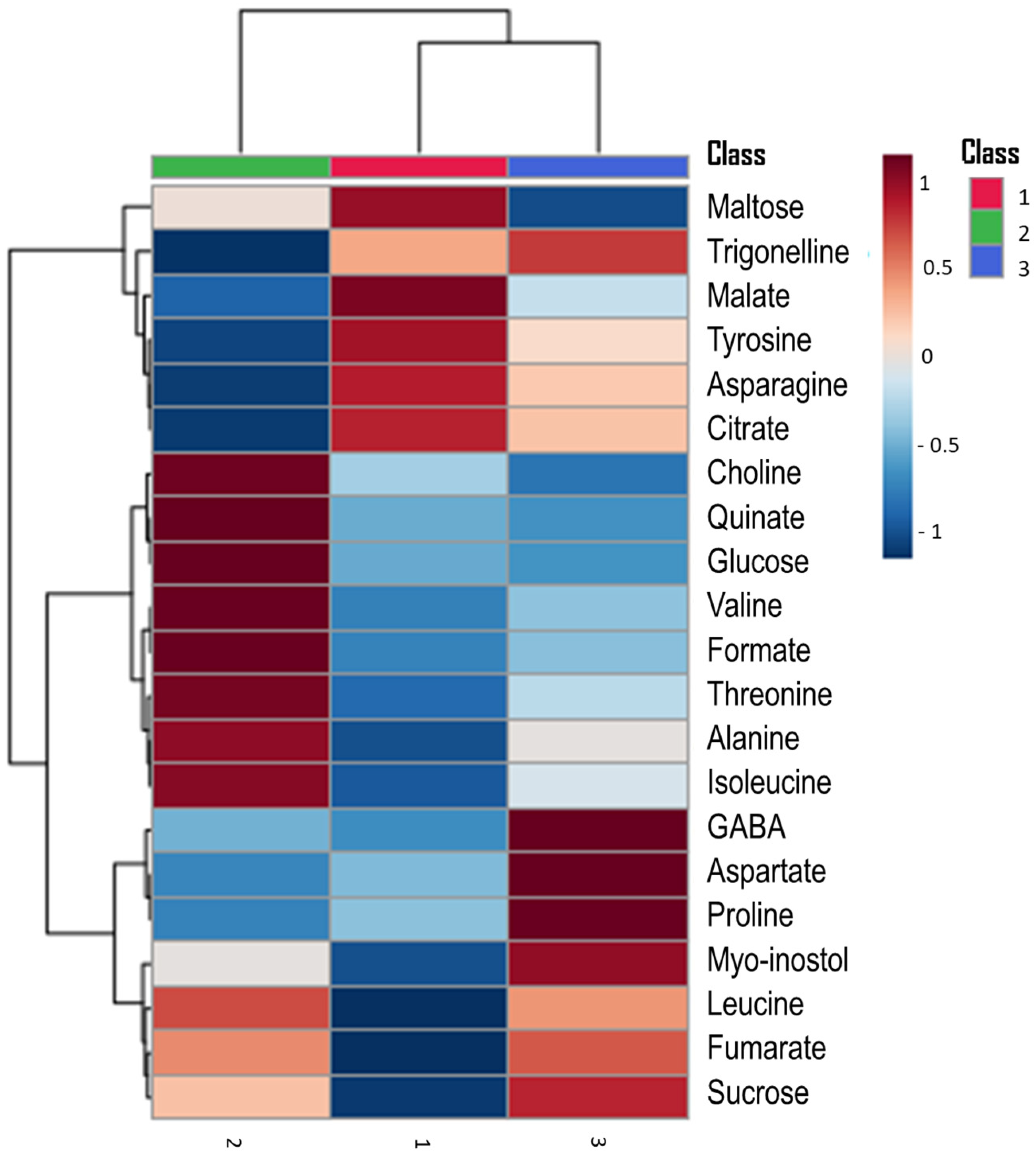
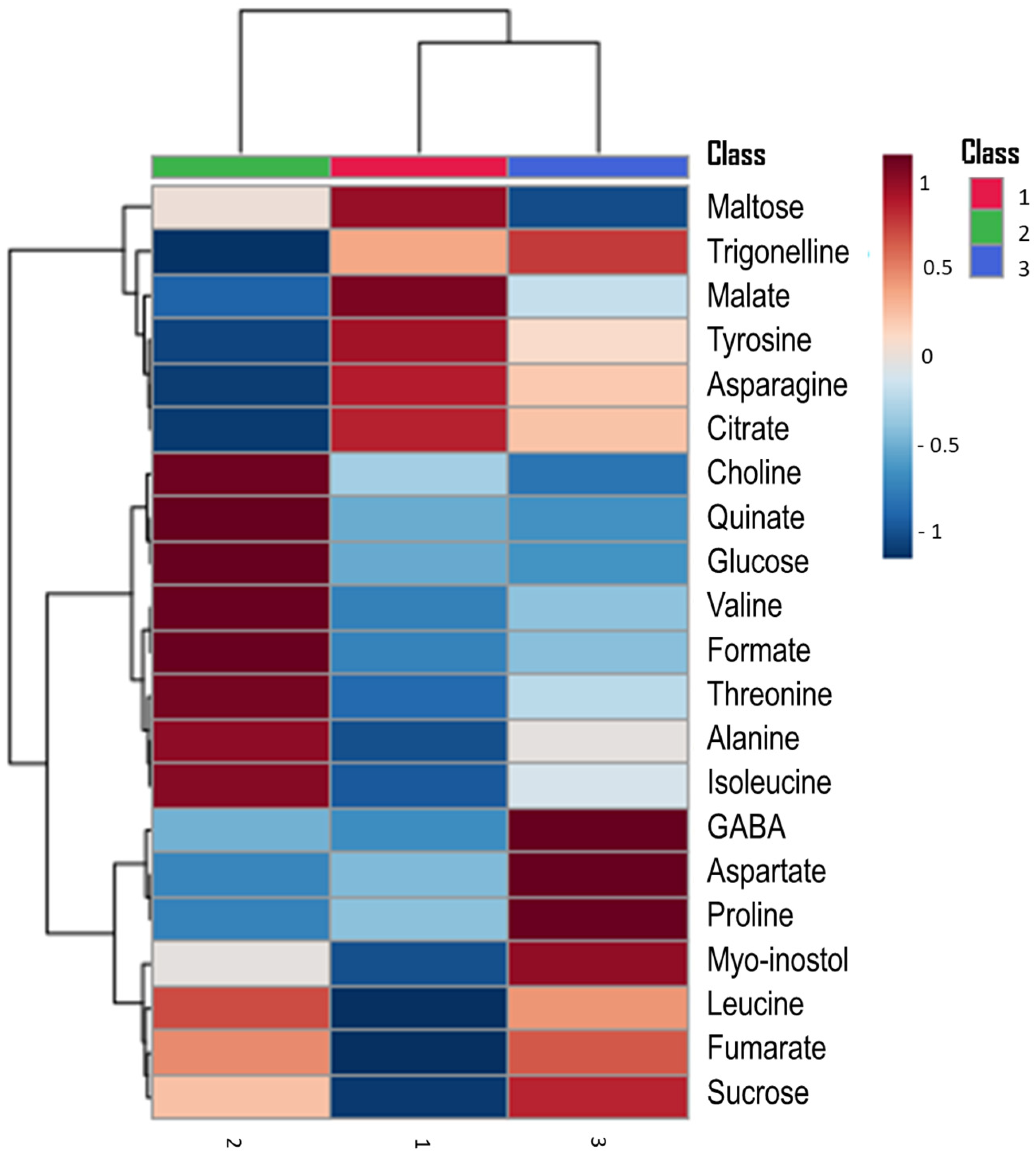
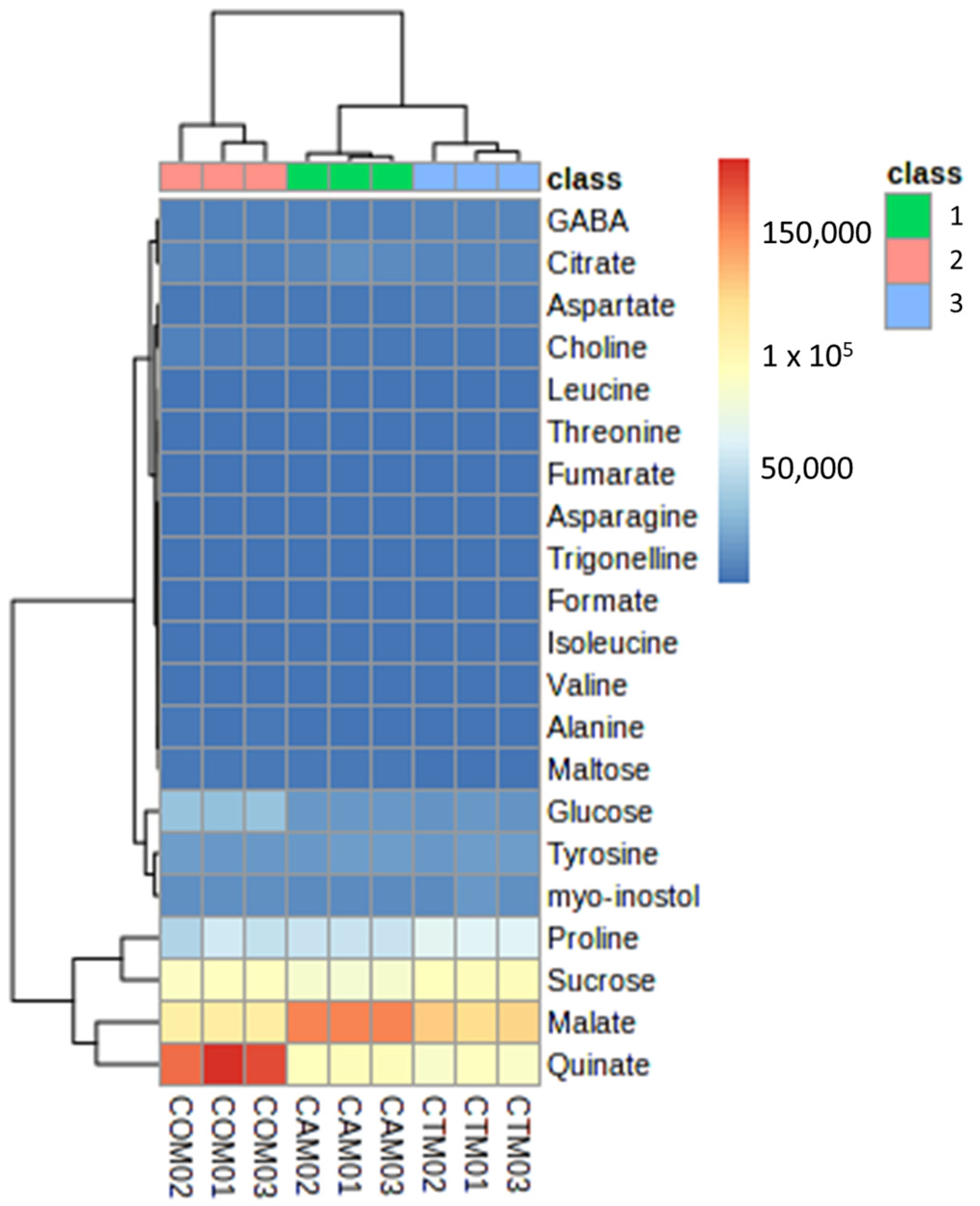
3.3. Hierarchical Clustering Heatmap
3.4. Significance Analysis of Microarray
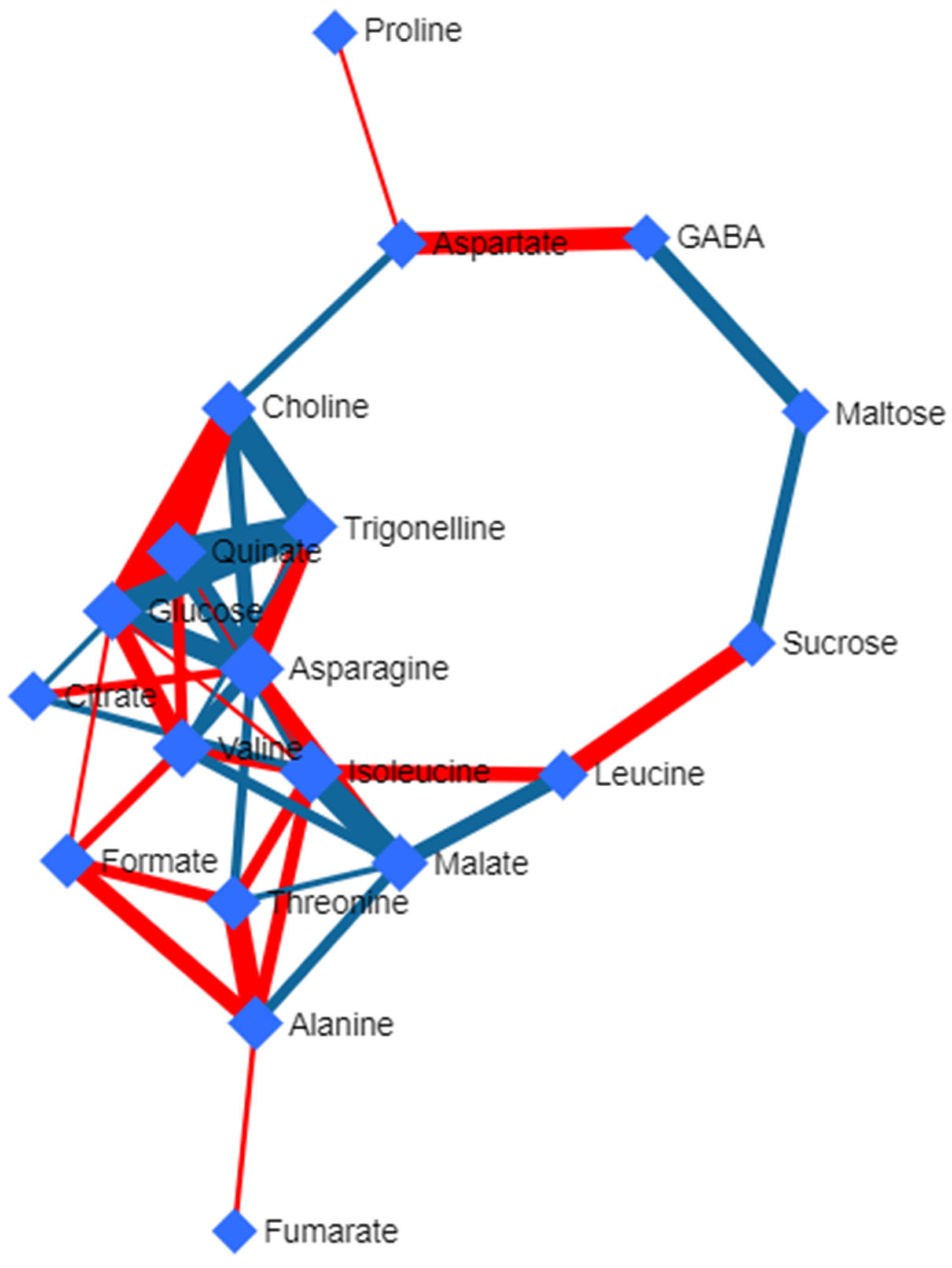
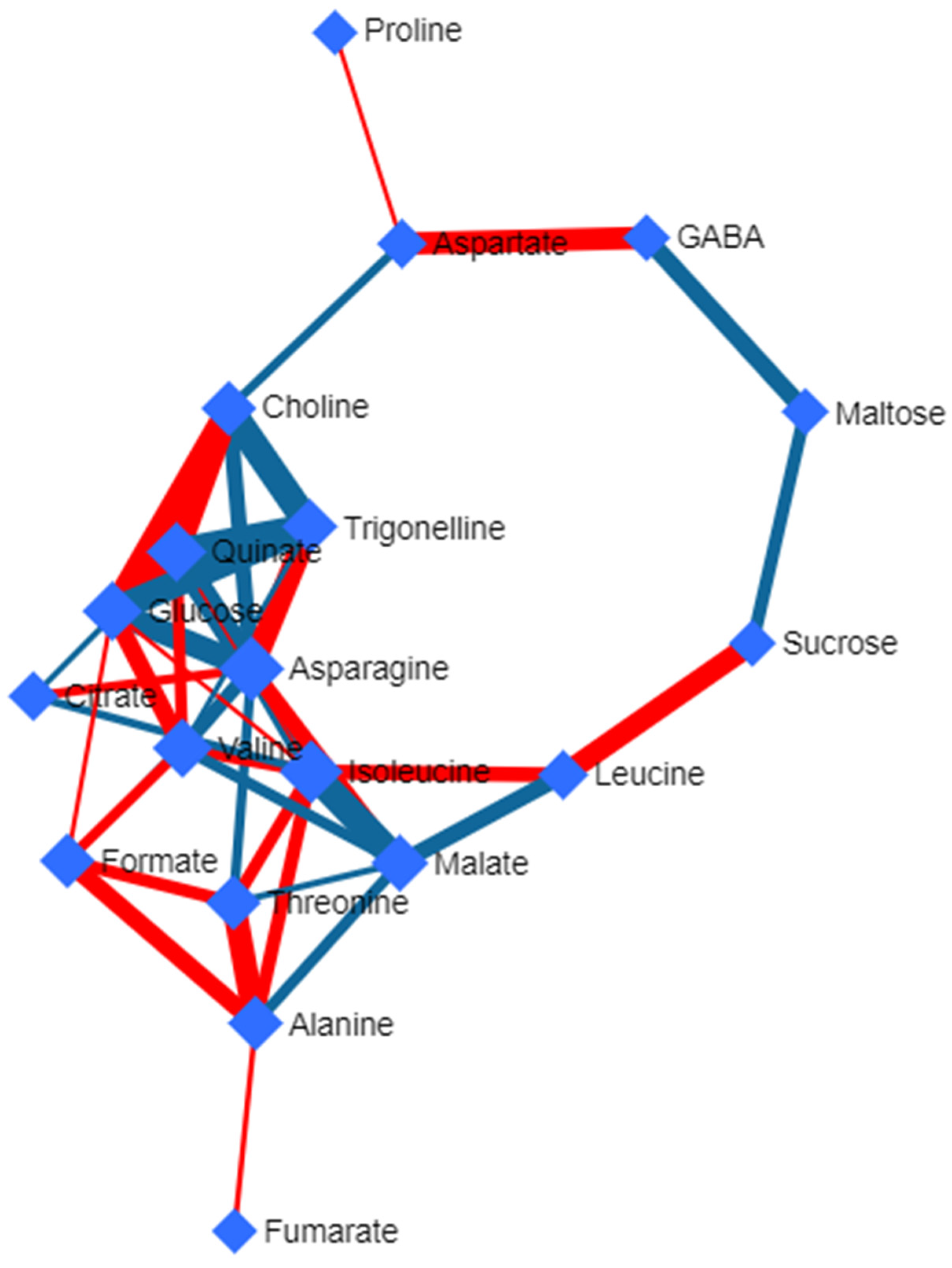
3.5. Deviated Scattered Partial Correlation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO Food and Agriculture Organization: Citrus. Available online: https://www.fao.org/economic/est/est-commodities/citrus-fruit/en/ (accessed on 27 January 2022).
- Shorbagi, M.; Fayek, N.M.; Shao, P.; Farag, M.A. Citrus reticulata Blanco (the Common Mandarin) Fruit: An Updated Review of Its Bioactive, Extraction Types, Food Quality, Therapeutic Merits, and Bio-Waste Valorization Practices to Maximize Its Economic Value. Food Biosci. 2022, 47, 101699. [Google Scholar] [CrossRef]
- Maciá-Vázquez, A.A.; Núñez-Gómez, D.; Martínez-Nicolás, J.J.; Legua, P.; Melgarejo, P. Morphological and Biochemical Characterization of Late-Season Varieties of Mandarin Growing in Spain under Homogeneous Growing Conditions. Agronomy 2023, 13, 1825. [Google Scholar] [CrossRef]
- Nadori, E.B. Mandarin Tangerine Called Nadorcott. US 08/787,028, 7 July 1998. [Google Scholar]
- Vardi, A.; Spiegel-roy, P.; Frydman-shani, A.; Elchanati, A.; Neumann, H. Citrus Tree Named ‘Orri’. US 09/716,482, 4 March 2003. [Google Scholar]
- Roose, M.L.; Williams, T.E. Mandarin Variety Named “Tango”. US 11/220,875, 8 March 2007. [Google Scholar]
- Tarancón, P.; Giménez-Sanchis, A.; Aleza, P.; Besada, C. Selection of New Late-Season Mandarin Cultivars Based on Sensory Changes and Consumer Acceptance after Fruit Cold Storage. Agronomy 2021, 11, 116. [Google Scholar] [CrossRef]
- Simón-Grao, S.; Gimeno, V.; Simón, I.; Lidón, V.; Nieves, M.; Balal, R.M.; Carbonell-Barrachina, A.A.; Manera, F.J.; Hernández, F.; García-Sánchez, F. Fruit Quality Characterization of Eleven Commercial Mandarin Cultivars in Spain. Sci. Hortic. 2014, 165, 274–280. [Google Scholar] [CrossRef]
- Maciá-Vázquez, A.A.; Martínez-Nicolás, J.J.; Núñez-Gómez, D.; Melgarejo, P.; Legua, P. Influence of Rootstock on Yield, Morphological, Biochemical and Sensory Characteristics of “Afourer” Variety Mandarins. Sci. Hortic. 2024, 325, 112644. [Google Scholar] [CrossRef]
- Asai, T.; Matsukawa, T.; Kajiyama, S. Metabolomic Analysis of Primary Metabolites in Citrus Leaf during Defense Responses. J. Biosci. Bioeng. 2017, 123, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, T.M. Synthesis versus Degradation: Directions of Amino Acid Metabolism during Arabidopsis Abiotic Stress Response. Plant Mol. Biol. 2018, 98, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Aharoni, A.; Brandizzi, F. High-Resolution Measurements in Plant Biology. Plant J. 2012, 70, 1–4. [Google Scholar] [CrossRef]
- Rolin, D.; Deborde, C.; Maucourt, M.; Cabasson, C.; Fauvelle, F.; Jacob, D.; Canlet, C.; Moing, A. High-Resolution 1H-NMR Spectroscopy and Beyond to Explore Plant Metabolome. Adv. Bot. Res. 2013, 67, 1–66. [Google Scholar]
- Okada, T.; Mochamad Afendi, F.; Altaf-Ul-Amin, M.; Takahashi, H.; Nakamura, K.; Kanaya, S. Metabolomics of Medicinal Plants: The Importance of Multivariate Analysis of Analytical Chemistry Data. Curr. Comput. Aided-Drug Des. 2010, 6, 179–196. [Google Scholar] [CrossRef]
- Kumar, D. Nuclear Magnetic Resonance (NMR) Spectroscopy For Metabolic Profiling of Medicinal Plants and Their Products. Crit. Rev. Anal. Chem. 2016, 46, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Gogna, N.; Hamid, N.; Dorai, K. Metabolomic Profiling of the Phytomedicinal Constituents of Carica papaya L. Leaves and Seeds by 1H NMR Spectroscopy and Multivariate Statistical Analysis. J. Pharm. Biomed. Anal. 2015, 115, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Melgarejo, P.; Núñez-Gómez, D.; Martínez-Nicolás, J.J.; Hernández, F.; Martínez-Font, R.; Lidón, V.; García-Sánchez, F.; Legua, P. Metabolomic Profile of Citrus Limon Leaves (‘Verna’ Variety) by 1H-NMR and Multivariate Analysis Technique. Agronomy 2022, 12, 1060. [Google Scholar] [CrossRef]
- Sousa, A.R.d.O.; Silva, E.M.d.A.; Filho, M.A.C.; Costa, M.G.C.; Filho, W.d.S.S.; Micheli, F.; Maserti, B.; Gesteira, A.d.S. Metabolic Responses to Drought Stress and Rehydration in Leaves and Roots of Three Citrus Scion/Rootstock Combinations. Sci. Hortic. 2022, 292, 110490. [Google Scholar] [CrossRef]
- Tristán, A.I.; Abreu, A.C.; Aguilera-Sáez, L.M.; Peña, A.; Conesa-Bueno, A.; Fernández, I. Evaluation of ORAC, IR and NMR Metabolomics for Predicting Ripening Stage and Variety in Melon (Cucumis melo L.). Food Chem. 2022, 372, 131263. [Google Scholar] [CrossRef] [PubMed]
- Castejón-Porcel, G.; Espín-Sánchez, D.; Ruiz-Álvarez, V.; García-Marín, R.; Moreno-Muñoz, D. Runoff Water as A Resource in the Campo de Cartagena (Region of Murcia): Current Possibilities for Use and Benefits. Water 2018, 10, 456. [Google Scholar] [CrossRef]
- SIAM Informe Agrometeorológico—TORRE PACHECO Station. Available online: http://siam.imida.es/apex/f?p=101:46:3131252762679599 (accessed on 29 March 2024).
- van der Sar, S.; Kim, H.K.; Meissner, A.; Verpoorte, R.; Choi, Y.H. Nuclear Magnetic Resonance Spectroscopy for Plant Metabolite Profiling. In The Handbook of Plant Metabolomics; Wiley: Hoboken, NJ, USA, 2013; pp. 57–76. [Google Scholar]
- Melgarejo, P.; Núñez-Gómez, D.; Hernández, F.; Martínez-Font, R.; Lidón Noguera, V.; Martínez-Nicolás, J.J.; Legua, P. Lemon Peel and Juice: Metabolomic Differentiation. Horticulturae 2023, 9, 510. [Google Scholar] [CrossRef]
- Huang, S.; Ying Lim, S.; Lau, H.; Ni, W.; Fong Yau Li, S. Effect of Glycinebetaine on Metabolite Profiles of Cold-Stored Strawberry Revealed by 1H NMR-Based Metabolomics. Food Chem. 2022, 393, 133452. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.P.; Rai, N.; Farag, M.A.; Maurya, S.; Yerasu, S.R.; Bisen, M.S.; Prabha, R.; Shukla, R.; Behera, T.K. Metabolic Diversity, Biosynthetic Pathways, and Metabolite Biomarkers Analysed via Untargeted Metabolomics and the Antioxidant Potential Reveal for High Temperature Tolerance in Tomato Hybrid. Plant Stress 2024, 11, 100420. [Google Scholar] [CrossRef]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Cao, X.; Khan, M.A.R. Proline, a Multifaceted Signalling Molecule in Plant Responses to Abiotic Stress: Understanding the Physiological Mechanisms. Plant Biol. 2022, 24, 227–239. [Google Scholar] [CrossRef]
- Yamada, N.; Promden, W.; Yamane, K.; Tamagake, H.; Hibino, T.; Tanaka, Y.; Takabe, T. Preferential Accumulation of Betaine Uncoupled to Choline Monooxygenase in Young Leaves of Sugar Beet—Importance of Long-Distance Translocation of Betaine under Normal and Salt-Stressed Conditions. J. Plant Physiol. 2009, 166, 2058–2070. [Google Scholar] [CrossRef]
- Cho, Y.; Lightfoot, D.A.; Wood, A.J. Trigonelline Concentrations in Salt Stressed Leaves of Cultivated Glycine Max. Phytochemistry 1999, 52, 1235–1238. [Google Scholar] [CrossRef]
- Cheng, P.; Yue, Q.; Zhang, Y.; Zhao, S.; Khan, A.; Yang, X.; He, J.; Wang, S.; Shen, W.; Qian, Q.; et al. Application of γ-Aminobutyric Acid (GABA) Improves Fruit Quality and Rootstock Drought Tolerance in Apple. J. Plant Physiol. 2023, 280, 153890. [Google Scholar] [CrossRef] [PubMed]
- Pandita, D. Jasmonates: Key Players in Plant Stress Tolerance. In Emerging Plant Growth Regulators in Agriculture; Elsevier: Amsterdam, The Netherlands, 2022; pp. 165–192. [Google Scholar]
- Surekha Devi, V.; Sharma, H.C.; Arjuna Rao, P. Influence of Oxalic and Malic Acids in Chickpea Leaf Exudates on the Biological Activity of CryIAc towards Helicoverpa armigera. J. Insect Physiol. 2013, 59, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Gaufichon, L.; Reisdorf-Cren, M.; Rothstein, S.J.; Chardon, F.; Suzuki, A. Biological Functions of Asparagine Synthetase in Plants. Plant Sci. 2010, 179, 141–153. [Google Scholar] [CrossRef]
- Lourkisti, R.; Antoine, S.; Pailly, O.; Luro, F.; Gibon, Y.; Oustric, J.; Santini, J.; Berti, L. GABA Shunt Pathway Is Stimulated in Response to Early Defoliation-Induced Carbohydrate Limitation in Mandarin Fruits. Heliyon 2023, 9, e15573. [Google Scholar] [CrossRef]
- Katz, E.; Fon, M.; Lee, Y.J.; Phinney, B.S.; Sadka, A.; Blumwald, E. The Citrus Fruit Proteome: Insights into Citrus Fruit Metabolism. Planta 2007, 226, 989–1005. [Google Scholar] [CrossRef]
- Xiong, H.; Luo, Y.; Rennenberg, H.; Wang, J.; Hu, B.; Zhao, H.; Tang, X.; Zhang, Y.; Shi, X. Enhancing Proline Turnover Is the Key Physiological Response of Mature Citrus Leaves to Fruiting. Sci. Hortic. 2023, 315, 111979. [Google Scholar] [CrossRef]
- Zhang, S.; Hu, Y.; Wang, Y.; Li, M.; Han, D.; Gao, S.; Hu, Y.; Guo, J.; Zhang, T.; Shi, L. Integrated Transcriptomics and Metabolomics Reveals That Regulating Biosynthesis and Metabolism of HCN and GABA Plays a Key Role in Drought Resistance of Wild Soybean. Env. Exp. Bot. 2023, 215, 105505. [Google Scholar] [CrossRef]
- Potel, F.; Valadier, M.; Ferrario-Méry, S.; Grandjean, O.; Morin, H.; Gaufichon, L.; Boutet-Mercey, S.; Lothier, J.; Rothstein, S.J.; Hirose, N.; et al. Assimilation of Excess Ammonium into Amino Acids and Nitrogen Translocation in Arabidopsis thaliana—Roles of Glutamate Synthases and Carbamoylphosphate Synthetase in Leaves. FEBS J. 2009, 276, 4061–4076. [Google Scholar] [CrossRef]
- Antunes, F.; Aguilar, M.; Pineda, M.; Sodek, L. Nitrogen Stress and the Expression of Asparagine Synthetase in Roots and Nodules of Soybean (Glycine max). Physiol. Plant 2008, 133, 736–743. [Google Scholar] [CrossRef]
- Galili, G. The Aspartate-Family Pathway of Plants. Plant Signal Behav. 2011, 6, 192–195. [Google Scholar] [CrossRef]
- Fait, A.; Angelovici, R.; Less, H.; Ohad, I.; Urbanczyk-Wochniak, E.; Fernie, A.R.; Galili, G. Arabidopsis Seed Development and Germination Is Associated with Temporally Distinct Metabolic Switches. Plant Physiol. 2006, 142, 839–854. [Google Scholar] [CrossRef]
- Schneidereit, J.; Häusler, R.E.; Fiene, G.; Kaiser, W.M.; Weber, A.P.M. Antisense Repression Reveals a Crucial Role of the Plastidic 2-oxoglutarate/Malate Translocator DiT1 at the Interface between Carbon and Nitrogen Metabolism. Plant J. 2006, 45, 206–224. [Google Scholar] [CrossRef]
- Lindén, P.; Keech, O.; Stenlund, H.; Gardeström, P.; Moritz, T. Reduced Mitochondrial Malate Dehydrogenase Activity Has a Strong Effect on Photorespiratory Metabolism as Revealed by 13C Labelling. J. Exp. Bot. 2016, 67, 3123–3135. [Google Scholar] [CrossRef]
- Dao, O.; Kuhnert, F.; Weber, A.P.M.; Peltier, G.; Li-Beisson, Y. Physiological Functions of Malate Shuttles in Plants and Algae. Trends Plant Sci. 2022, 27, 488–501. [Google Scholar] [CrossRef]
- Narayanamma, V.L.; Sharma, H.C.; Gowda, C.L.L.; Sriramulu, M. Incorporation of Lyophilized Leaves and Pods into Artificial Diets to Assess the Antibiosis Component of Resistance to Pod Borer Helicoverpa armigera (Lepidoptera: Noctuidae) in Chickpea. Int. J. Trop. Insect Sci. 2007, 27, 191. [Google Scholar] [CrossRef]
- Yoshida, M.; Cowgill, S.E.; Wightman, J.A. Roles of Oxalic and Malic Acids in Chickpea Trichome Exudate in Host-Plant Resistance to Helicoverpa armigera. J. Chem. Ecol. 1997, 23, 1195–1210. [Google Scholar] [CrossRef]
- Carrington, Y.; Guo, J.; Le, C.H.; Fillo, A.; Kwon, J.; Tran, L.T.; Ehlting, J. Evolution of a Secondary Metabolic Pathway from Primary Metabolism: Shikimate and Quinate Biosynthesis in Plants. Plant J. 2018, 95, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Leiss, K.A.; Maltese, F.; Choi, Y.H.; Verpoorte, R.; Klinkhamer, P.G.L. Identification of Chlorogenic Acid as a Resistance Factor for Thrips in Chrysanthemum. Plant Physiol. 2009, 150, 1567–1575. [Google Scholar] [CrossRef]
- Rocha-Santos, T.; Duarte, A.C. Introduction to the Analysis of Bioactive Compounds in Marine Samples. In Comprehensive Analytical Chemistry; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1–13. [Google Scholar]
- Chen, X.; Hao, R.; Chen, W.; Jia, H.; Qin, S.; Wang, Q.; Zhang, D.; Han, Z.; Li, Y. Effect of Choline Amino Acid Ionic Liquids on Maize Seed Germination and Endogenous Plant Hormone Levels. RSC Adv. 2024, 14, 382–389. [Google Scholar] [CrossRef]
- Kelebek, H.; Selli, S. Determination of Volatile, Phenolic, Organic Acid and Sugar Components in a Turkish Cv. Dortyol (Citrus sinensis L. Osbeck) Orange Juice. J. Sci. Food Agric. 2011, 91, 1855–1862. [Google Scholar] [CrossRef] [PubMed]
- Katz, E.; Boo, K.H.; Kim, H.Y.; Eigenheer, R.A.; Phinney, B.S.; Shulaev, V.; Negre-Zakharov, F.; Sadka, A.; Blumwald, E. Label-Free Shotgun Proteomics and Metabolite Analysis Reveal a Significant Metabolic Shift during Citrus Fruit Development. J. Exp. Bot. 2011, 62, 5367–5384. [Google Scholar] [CrossRef] [PubMed]
- Mohamadi, N.; Sharififar, F.; Pournamdari, M.; Ansari, M. A Review on Biosynthesis, Analytical Techniques, and Pharmacological Activities of Trigonelline as a Plant Alkaloid. J. Diet. Suppl. 2018, 15, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Nuccio, M.L.; McNeil, S.D.; Ziemak, M.J.; Hanson, A.D.; Jain, R.K.; Selvaraj, G. Choline Import into Chloroplasts Limits Glycine Betaine Synthesis in Tobacco: Analysis of Plants Engineered with a Chloroplastic or a Cytosolic Pathway. Metab. Eng. 2000, 2, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Syvertsen, J.P.; Garcia-Sanchez, F. Multiple Abiotic Stresses Occurring with Salinity Stress in Citrus. Environ. Exp. Bot. 2014, 103, 128–137. [Google Scholar] [CrossRef]
- Rodríguez-Gamir, J.; Primo-Millo, E.; Forner, J.B.; Forner-Giner, M.A. Citrus Rootstock Responses to Water Stress. Sci. Hortic. 2010, 126, 95–102. [Google Scholar] [CrossRef]
- Pérez-Pérez, J.G.; Syvertsen, J.P.; Botía, P.; García-Sánchez, F. Leaf Water Relations and Net Gas Exchange Responses of Salinized Carrizo Citrange Seedlings during Drought Stress and Recovery. Ann. Bot. 2007, 100, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Jolliffe, I.T.; Cadima, J. Principal Component Analysis: A Review and Recent Developments. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374, 20150202. [Google Scholar] [CrossRef]
- Janková, J.; van de Geer, S. Confidence Intervals for High-Dimensional Inverse Covariance Estimation. Electron. J. Stat. 2015, 9, 1205–1229. [Google Scholar] [CrossRef]
- Lee, L.C.; Liong, C.-Y.; Jemain, A.A. Partial Least Squares-Discriminant Analysis (PLS-DA) for Classification of High-Dimensional (HD) Data: A Review of Contemporary Practice Strategies and Knowledge Gaps. Analyst 2018, 143, 3526–3539. [Google Scholar] [CrossRef]
- Wilkinson, L.; Friendly, M. The History of the Cluster Heat Map. Am. Stat. 2009, 63, 179–184. [Google Scholar] [CrossRef]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance Analysis of Microarrays Applied to the Ionizing Radiation Response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef]
- Basu, S.; Duren, W.; Evans, C.R.; Burant, C.F.; Michailidis, G.; Karnovsky, A. Sparse Network Modeling and Metscape-Based Visualization Methods for the Analysis of Large-Scale Metabolomics Data. Bioinformatics 2017, 33, 1545–1553. [Google Scholar] [CrossRef] [PubMed]
- Ireland, R.J.; Lea, P.J. The Enzymes of Glutamine, Glutamate, Asparagine and Aspartate Metabolism; Dekker, M. Inc.: New York, NY, USA; Basel, Switzerland; Hong Kong, China, 1999. [Google Scholar]
- Gorissen, S.H.M.; Phillips, S.M. Branched-Chain Amino Acids (Leucine, Isoleucine, and Valine) and Skeletal Muscle. In Nutrition and Skeletal Muscle; Elsevier: Amsterdam, The Netherlands, 2019; pp. 283–298. [Google Scholar]
- Kumari, A. Citric Acid Cycle. In Sweet Biochemistry; Elsevier: Amsterdam, The Netherlands, 2023; pp. 9–15. [Google Scholar]
- Duggleby, R.G.; McCourt, J.A.; Guddat, L.W. Structure and Mechanism of Inhibition of Plant Acetohydroxyacid Synthase. Plant Physiol. Biochem. 2008, 46, 309–324. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types of Compounds | Compounds | Chemical Shift (ppm) 1 |
|---|---|---|
| Amino acids | GABA | 2.3 (t) |
| Alanine | 1.46 (d) | |
| Asparagine | 2.94 (dd) | |
| Aspartate | 2.81 (dd) | |
| Isoleucine | 0.98 (d) | |
| Leucine | 0.95 (t) | |
| Proline | 2.0 (m) | |
| Threonine | 1.3 (d) | |
| Tyrosine | 6.9 (d) | |
| Valine | 1.0 (d) | |
| Organic acids | Citrate | 2.69 (d) |
| Formate | 8.43 (s) | |
| Fumarate | 6.53 (s) | |
| Malate | 2.7 (d) | |
| Quinate | 1.7 (dd) | |
| Sugars | Glucose | 5.2 (d) |
| Maltose | 5.44 (d) | |
| myo-inostol | 3.3 (t) | |
| Sucrose | 5.4 (d) | |
| Other metabolites | Choline | 3.2 (s) |
| Trigonelline | 9.1 (s) |
| Types of Compounds | Compounds | ‘Afourer’ | ‘Orri’ | ‘Tango’ |
|---|---|---|---|---|
| Amino acids | GABA | 0.58 ± 0.01 a | 0.62 ± 0.03 a | 0.89 ± 0.03 b |
| Alanine | 0.11 ± 0.01 a | 0.21 ± 0.06 a | 0.16 ± 0.01 a | |
| Asparagine | 0.09 ± 0 b | ND | 0.07 ± 0 a | |
| Aspartate | 0.3 ± 0.02 a | 0.28 ± 0.02 a | 0.46 ± 0 b | |
| Isoleucine | 0.03 ± 0 a | 0.06 ± 0 b | 0.04 ± 0 a | |
| Leucine | 0.03 ± 0.01 a | 0.12 ± 0.04 a | 0.11 ± 0 a | |
| Proline | 5.26 ± 0.05 a | 5.01 ± 0.65 a | 6.39 ± 0.05 a | |
| Threonine | 0.06 ± 0.01 a | 0.09 ± 0.02 a | 0.07 ± 0.01 a | |
| Tyrosine | 1.67 ± 0.04 a | 1.65 ± 0.01 a | 1.66 ± 0.06 a | |
| Valine | 0.04 ± 0 a | 0.08 ± 0.01 a | 0.04 ± 0.02 a | |
| Organic acids | Citrate | 1 ± 0.15 a | 0.72 ± 0.09 a | 0.91 ± 0.02 a |
| Formate | 0.02 ± 0 a | 0.03 ± 0.01 a | 0.02 ± 0 a | |
| Fumarate | 0.08 ± 0.01 a | 0.09 ± 0.02 a | 0.09 ± 0.01 a | |
| Malate | 15.46 ± 0.05 c | 10.8 ± 0.04 a | 12.51 ± 0.38 b | |
| Quinate | 9.34 ± 0.05 a | 17.28 ± 1.01 b | 8.72 ± 0.29 a | |
| Sugars | Glucose | 1.57 ± 0.02 a | 3.33 ± 0.16 b | 1.45 ± 0.15 a |
| Maltose | 0.29 ± 0.06 a | 0.23 ± 0 a | 0.16 ± 0.02 a | |
| myo-inostol | 1.09 ± 0.01 a | 1.17 ± 0.03 a | 1.25 ± 0.24 a | |
| Sucrose | 8.26 ± 0.13 a | 9.02 ± 0.11 b | 9.35 ± 0.12 b | |
| Other metabolites | Choline | 0.37 ± 0 b | 0.56 ± 0.01 c | 0.31 ± 0.02 a |
| Trigonelline | 0.07 ± 0 b | 0.04 ± 0 a | 0.08 ± 0 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maciá-Vázquez, A.A.; Núñez-Gómez, D.; Martínez-Nicolás, J.J.; Legua, P.; Melgarejo, P. Mandarin Variety Significantly Affects the Metabolites Present in the Leaves. Horticulturae 2024, 10, 359. https://doi.org/10.3390/horticulturae10040359
Maciá-Vázquez AA, Núñez-Gómez D, Martínez-Nicolás JJ, Legua P, Melgarejo P. Mandarin Variety Significantly Affects the Metabolites Present in the Leaves. Horticulturae. 2024; 10(4):359. https://doi.org/10.3390/horticulturae10040359
Chicago/Turabian StyleMaciá-Vázquez, Alejandro Andy, Dámaris Núñez-Gómez, Juan José Martínez-Nicolás, Pilar Legua, and Pablo Melgarejo. 2024. "Mandarin Variety Significantly Affects the Metabolites Present in the Leaves" Horticulturae 10, no. 4: 359. https://doi.org/10.3390/horticulturae10040359
APA StyleMaciá-Vázquez, A. A., Núñez-Gómez, D., Martínez-Nicolás, J. J., Legua, P., & Melgarejo, P. (2024). Mandarin Variety Significantly Affects the Metabolites Present in the Leaves. Horticulturae, 10(4), 359. https://doi.org/10.3390/horticulturae10040359