
Phenotypic and Molecular Diversity of Wild Populations of Acca sellowiana (Berg.) Burret in the Southern Area of Natural Distribution
 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
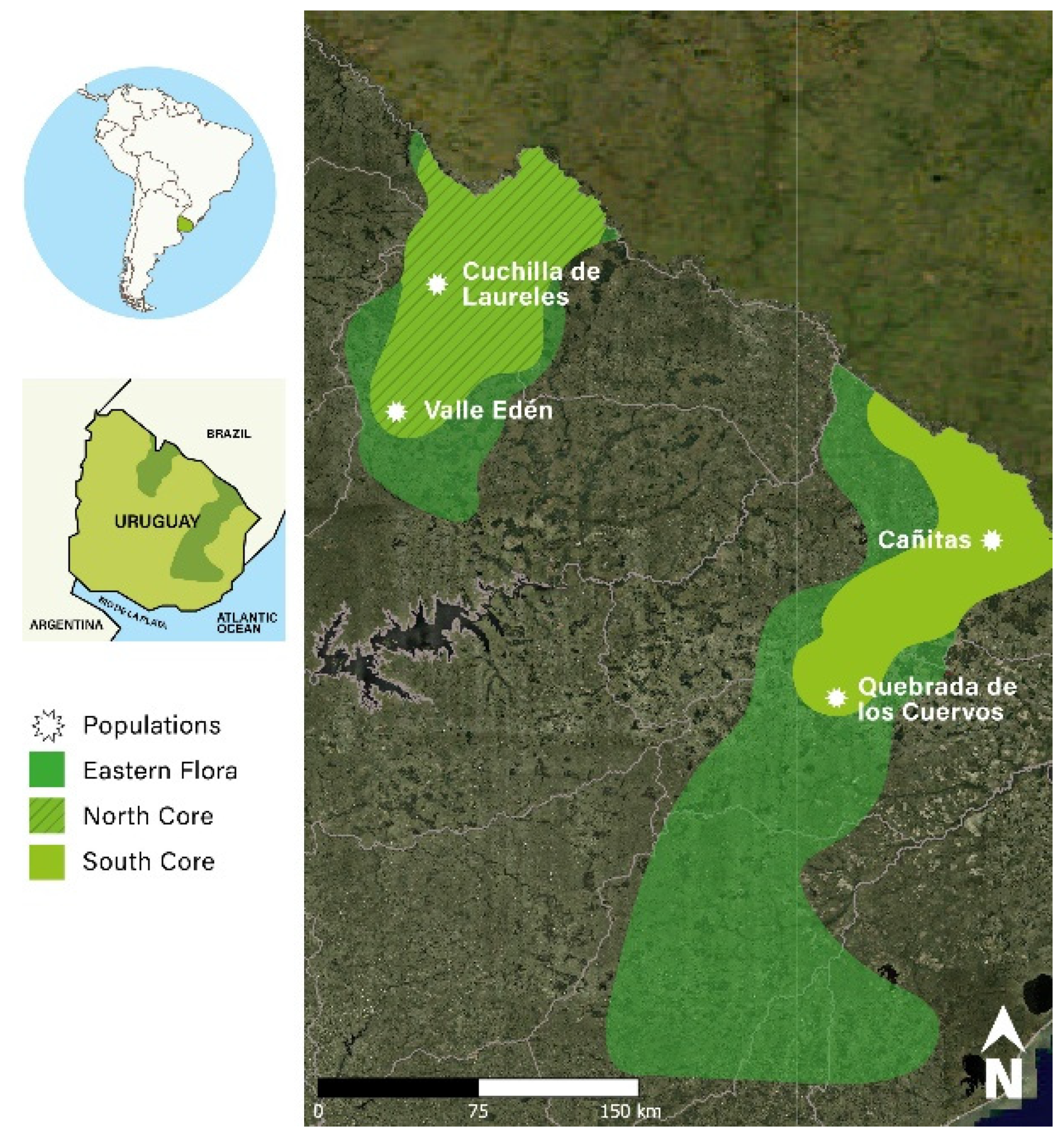
2.1. Plant Material
2.2. Phenotypic Characterization
2.3. Molecular Characterization
2.4. Statistical Analysis of the Phenotypic Data
2.5. Molecular Data Analysis
3. Results
3.1. Characterization of Phenotypic Diversity
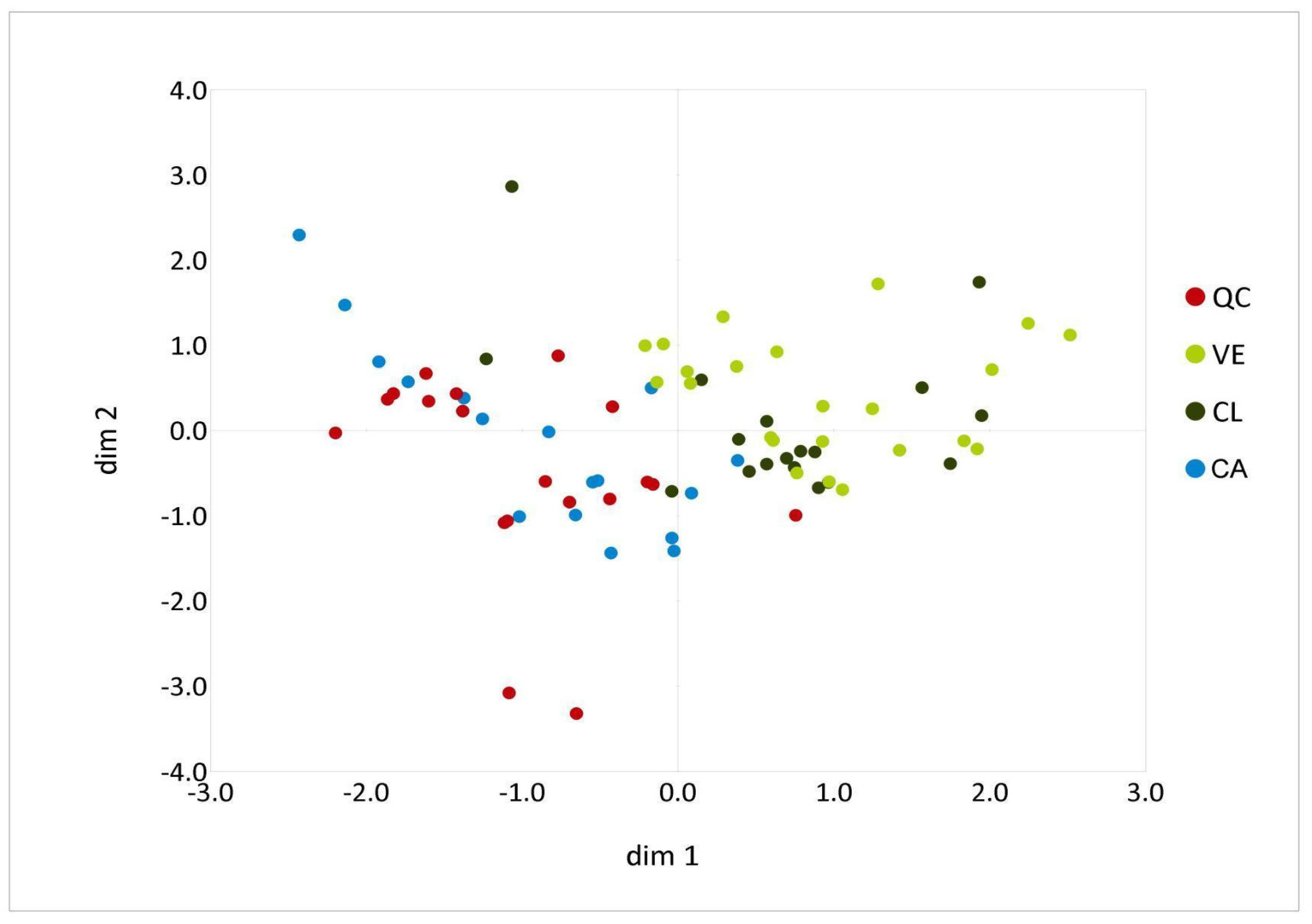
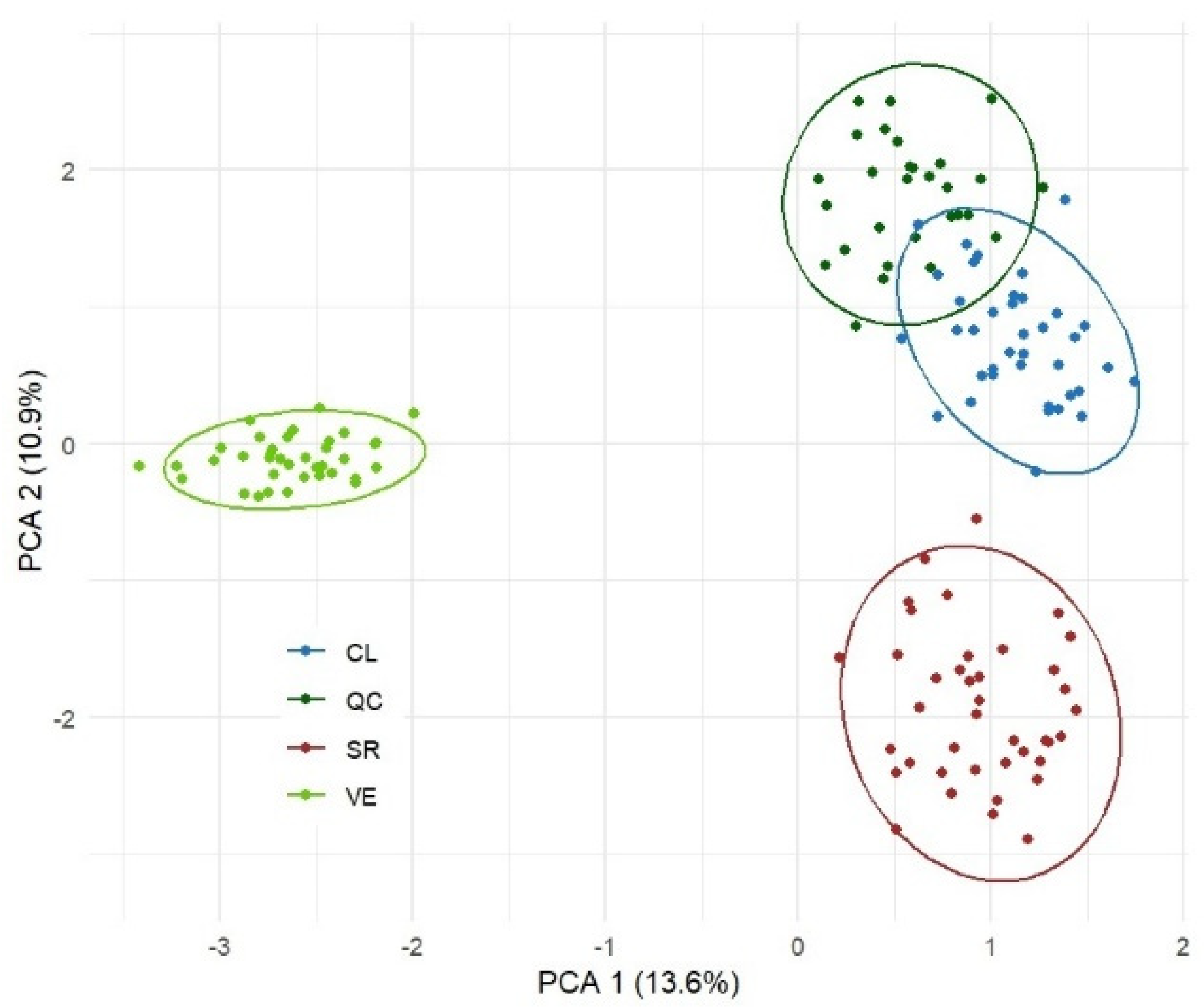
3.2. Multivariate Analysis of Phenotypic Diversity
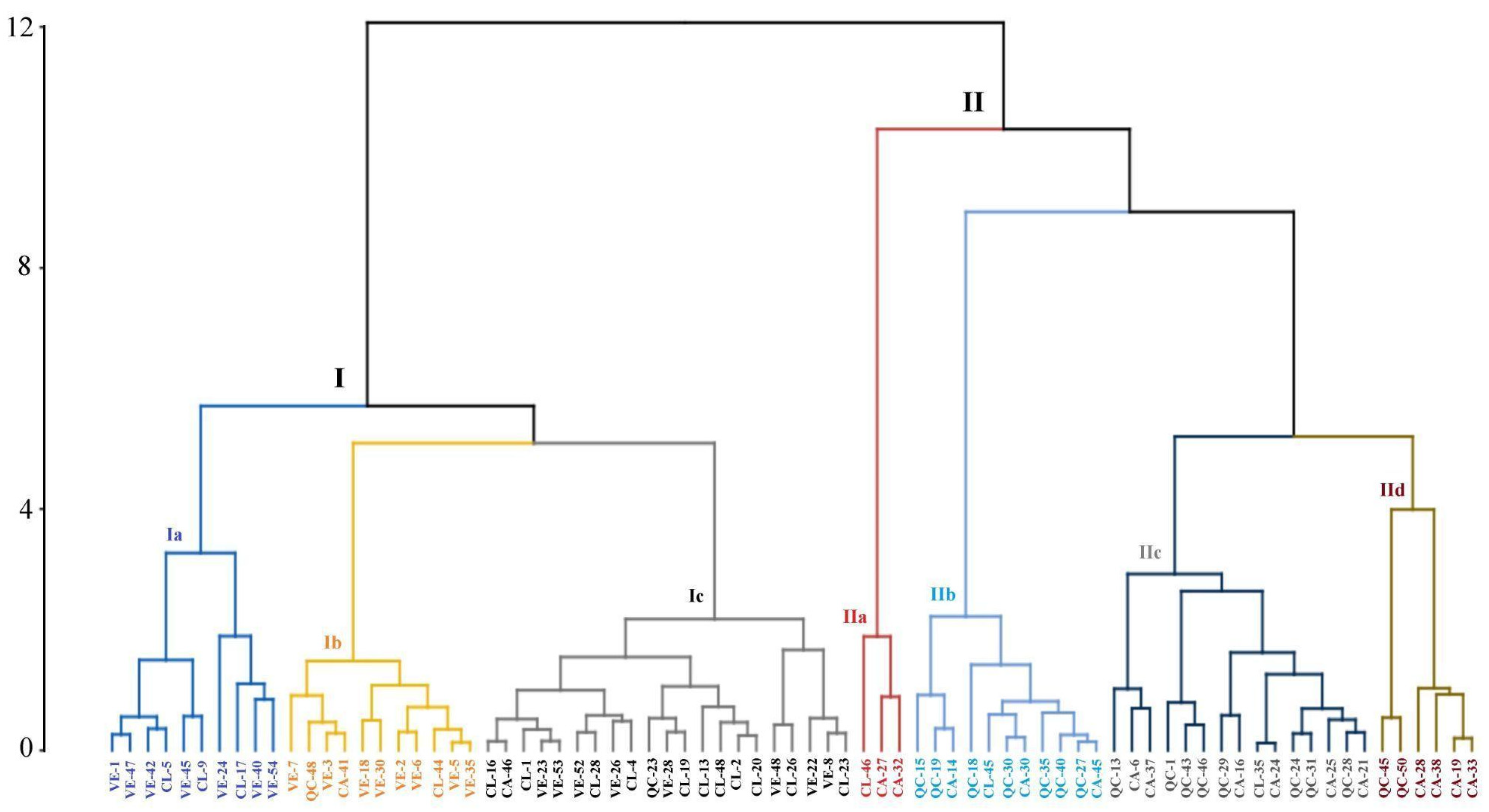
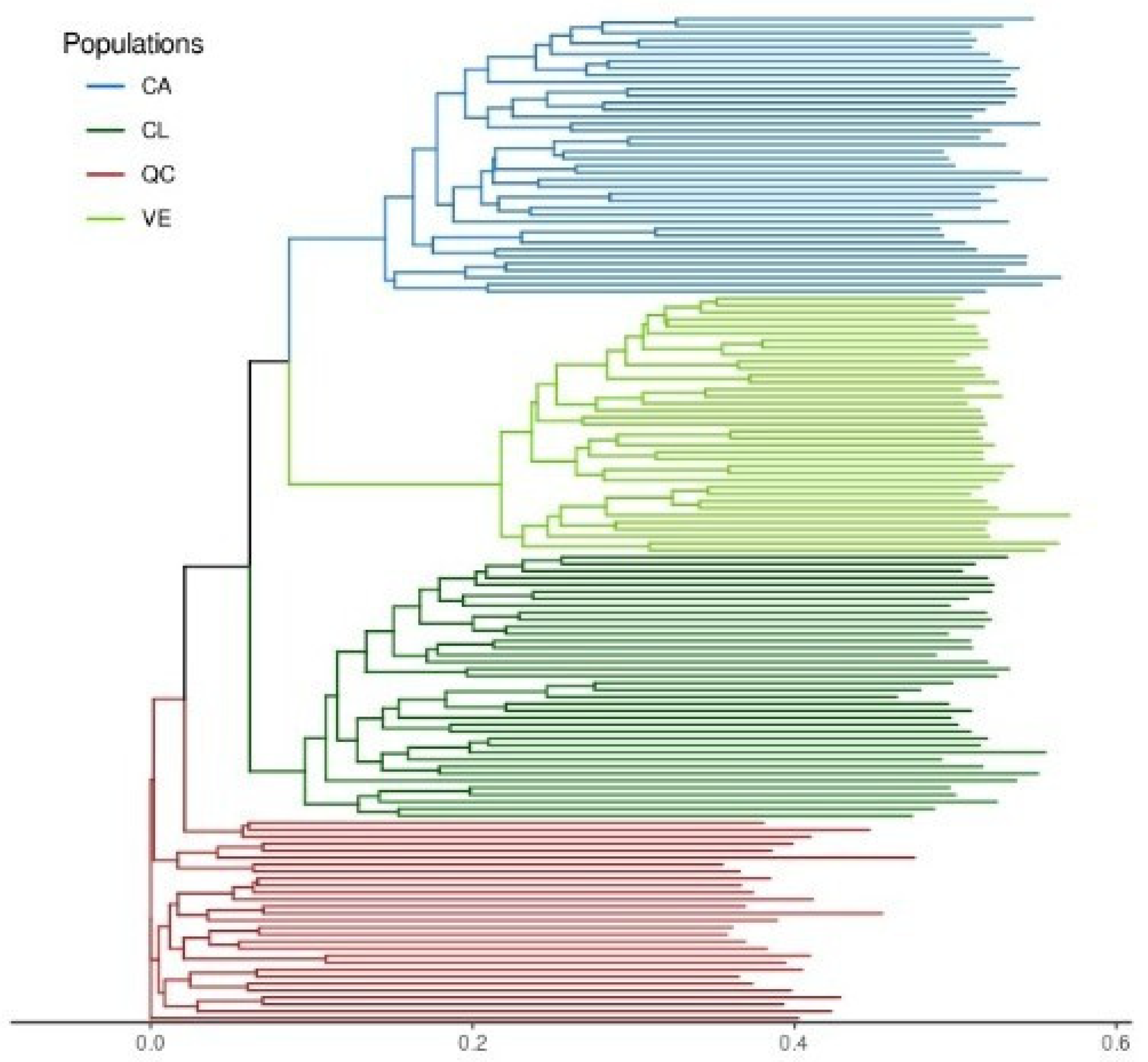
3.3. Characterization of Molecular Diversity
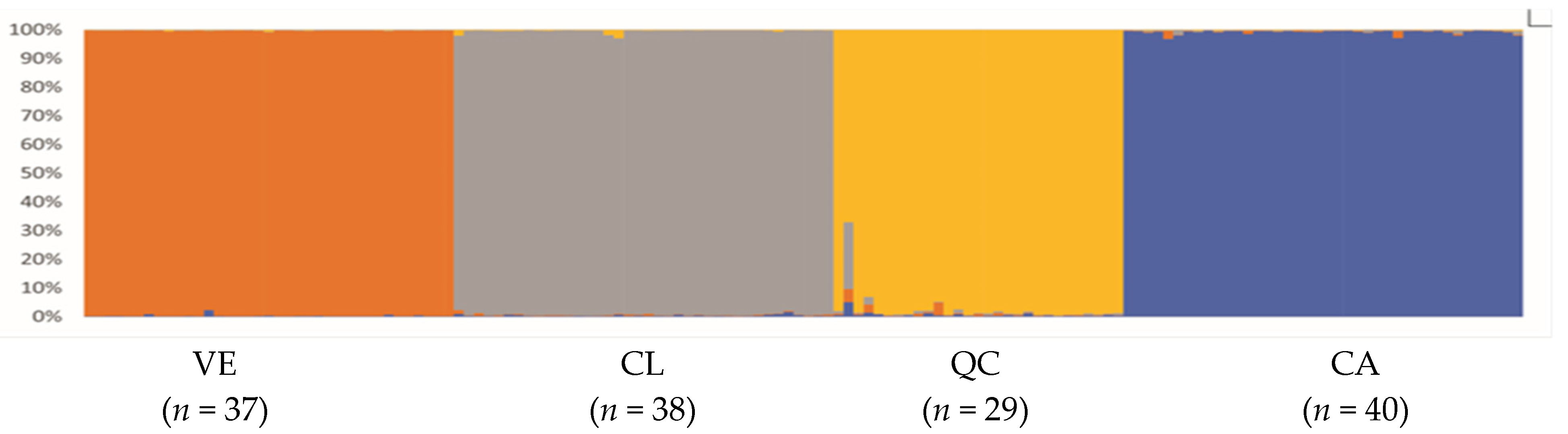
3.4. Estimation of Genetic Diversity per Population
4. Discussion
4.1. Molecular and Phenotypic Diversity
4.2. Population Structure and Geographic Distribution
4.3. Conservation and Utilization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ducroquet, J.P.H.J.; Hickel, E.R.; Nodari, R.O. Goiabeira-Serrana (Feijoa sellowiana); FUNEP: Jaboticabal, Brazil, 2000. [Google Scholar]
- Thorp, G.; Bieleski, R. Feijoas: Origins, Cultivation and Uses; HortResearch: Auckland, New Zealand; David Bateman: Auckland, New Zealand, 2002. [Google Scholar]
- Keller, H.A.; Tressens, S.G. Presencia en Argentina de dos especies de uso múltiple: Acca sellowiana (Myrtaceae) y Casearia lasiophylla (Flacourtiaceae). Darwiniana 2007, 45, 204–212. [Google Scholar]
- Rivas, M.; Ayres, C.; Zoppolo, R.; Cabrera, D.; Dellacassa, E.; Bellenda, B.; García, M.; Silveira, A.C.; Vignale, B.; Záccari, F.; et al. Valorización de los recursos genéticos del guayabo del país. Rev. INIA 2010, 23, 38–41. [Google Scholar]
- Rampoldi, C.; Ruíz, M.F.; Burzaco, P.; Arcia, P.; Sosa, J.; Irisity, M.; Ayres, C.; Rivas, M. Productos de Frutos Nativos: Guayabo del País (Acca sellowiana); ANII: Montevideo, Uruguay, 2013; 24p. [Google Scholar]
- Santos, K.L.; Ciotta, M.N.; Almeida, E.B.C.; Pasa, M.S. Importancia social e económica. In A Cultura de Goiabeira Serrana; Ciotta, M.N., Arioli, C.J., Pinto, F.A.M.F., Santos, K.L., Araujo, L., Pasa, M.S., Eds.; Epagri: Florianópolis, Brazil, 2018; pp. 201–215. 216p. [Google Scholar]
- Ielpo, M.T.; Basile, A.; Miranda, R.; Moscatiello, V.; Nappo, C.; Sorbo, S.; Laghi, E.; Ricciardi, M.M.; Ricciardi, L.; Vuotto, M.L. Inmunopharmacological properties of flavonoids. Fitoterapia 2000, 71, 101–109. [Google Scholar] [CrossRef]
- Lim, T.K. Acca sellowiana . In Edible Medicinal and Non-Medicinal Plants; Fruits; Springer: Dordrecht, The Netherlands, 2012; Volume 3, pp. 601–608. [Google Scholar]
- Silveira, A.C.; Záccari, F.; Oyarzún, D.; Rivas, M. Determinación de algunos atributos de calidad en frutos de guayabo del país [Acca sellowiana (Berg) Burret] en diferentes estados de maduración. Agrociencia 2015, 19, 24–30. [Google Scholar] [CrossRef]
- Pasquariello, M.S.; Mastrobuoni, F.; Di Patre, D.; Zampella, L.; Capuano, R.L.; Scortichini, M.; Petriccione, M. Agronomic, nutraceutical and molecular variability of feijoa (Acca sellowiana (O. Berg) Burret) germplasm. Sci. Hortic. 2015, 191, 1–9. [Google Scholar] [CrossRef]
- Beyhan, O.; Elmastas, M.; Gedikli, F. Total phenolic compounds and antioxidant capacity of leaf, dry fruit and fresh fruit of feijoa (Acca sellowiana, Myrtaceae). J. Med. Plants Res. 2010, 4, 1065–1072. [Google Scholar]
- Weston, R.J. Bioactive products from fruit of the feijoa (Feijoa sellowiana, Myrtaceae): A review. Food Chem. 2010, 121, 923–926. [Google Scholar] [CrossRef]
- Zhu, F. Chemical and biological properties of feijoa (Acca sellowiana). Trends Food Sci. 2018, 81, 121–131. [Google Scholar] [CrossRef]
- Sazima, I.; Sazima, M. Petiscos florais: Pétalas de Acca sellowiana (Myrtaceae) como fonte alimentar para aves em área urbana no Sul do Brasil. Biota Neotrop. 2007, 7, 307–311. [Google Scholar] [CrossRef]
- Mattos, J.R. A Goiabeira Serrana; Instituto de Pesquisas de Recursos Naturais Renováveis: Porto Alegre, Brazil, 1986; 84p. [Google Scholar]
- Nodari, R.O.; Guerra, M.P.; Meler, K.; Ducroquet, J.P. Genetic variability of Feijoa sellowiana germplasm. Acta Hortic. 1997, 452, 41–46. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; Mcmahon, T.A. Updated world map of the Koppen-Geiger climate classification. Hydrol. Earth Syst. Sci. Discuss. Eur. Geosci. Union 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Grela, I. Geografía Florística de las Especies Arbóreas de Uruguay. Propuesta para la Delimitación de Dendrofloras. Master’s Thesis, PEDECIBA, Isidoro de María, Uruguay, 2004; 97p. [Google Scholar]
- Fischer, G.; Miranda, D.; Cayón, G.; Mazorra, M. Cultivo, Poscosecha y Exportación de la Feijoa (Acca sellowiana Berg.); Universidad Nacional de Colombia, Fondo Nacional de Fomento Hortifrutícola, Asociación Hortifrutícola de Colombia: Bogotá, Colombia, 2003. [Google Scholar]
- Moretto, S.P.; Nodari, E.S.; Nodari, R.O. A introdução e os usos da Feijoa ou Goiabeira Serrana (Acca sellowiana): A perspectiva da história ambiental. Front. J. Soc. Technol. Environ. Sci. 2014, 3, 67–69. [Google Scholar] [CrossRef]
- Santos, K.L.; Ciotta, M.N.; Donazzolo, J.; Borsuk, L.J.; Nodari, R.O. Domesticaçao da especie. In A Cultura de Goiabeira Serrana; Ciotta, M.N., Arioli, C.J., Pinto, F.A.M.F., Santos, K.L., Araujo, L., Pasa, M.S., Eds.; Epagri: Florianópolis, Brazil, 2018; 216p. [Google Scholar]
- Sharpe, R.H.; Sherman, W.B.; Miller, E.P. Feijoa history and improvement. Proc. Fla. State Hortic. Soc. 1993, 106, 134–139. [Google Scholar]
- Dettori, M.T.; Palombi, M.A. Identification of Feijoa sellowiana Berg. accessions by RAPD markers. Sci. Hortic. 2000, 86, 279–290. [Google Scholar] [CrossRef]
- Popenoe, F.W. Feijoa sellowiana: Its history, culture and varieties. Pomona Coll. J. Econ. Bot. 1912, II, 217–242. [Google Scholar]
- Miranda, D. Sistemas de propagación y establecimiento del cultivo de feijoa. In Cultivo, Poscosecha y Exportación de la Feijoa (Acca sellowiana Berg.); Fischer, G., Miranda, D., Cayón, G., Mazorra, M., Eds.; Universidad Nacional de Colombia, Fondo Nacional de Fomento Hortofrutícola, Asociación Hortofrutícola de Colombia: Bogotá, Colombia, 2003; pp. 27–48. [Google Scholar]
- Vignale, B.; Bisio, L. Selección de frutales nativos en Uruguay. Agrociencia 2005, 9, 35–39. [Google Scholar]
- Santos, K.L.; Peroni, N.; Guries, R.P.; Nodari, R.O. Traditional knowledge and management of Feijoa (Acca sellowiana) in southern Brazil. Econ. Bot. 2009, 63, 204–214. [Google Scholar] [CrossRef]
- Cabrera, D.; Vignale, B.; Pritsch, C. Acca sellowiana Berg. Burret; PROCISUR/IICA: Montevideo, Uruguay, 2018; Available online: https://www.procisur.org.uy/adjuntos/80ca732ba9b2_Guayabo-PROCISUR.pdf (accessed on 1 March 2024).
- Ducroquet, J.P.H.J.; Santos, K.L.; Andrade, E.R.; Boneti, J.I.S.; Bonin, V.; Nodari, R.O. As Primeiras Cultivares Brasileiras de Goiabeira serrana: SCS 411 Alcantara e SCS 412 Helena. Rev. Agropecu. Catarin. 2007, 20, 77–80. [Google Scholar]
- Ducroquet, J.P.H.J.; Nunes, E.C.; Guerra, M.P.; Nodari, R.O. Novas cultivares brasileiras de goiabeira serrana: SCS 414-Mattos, SCS 415 Nonante. Rev. Agropecu. Catarin. 2008, 21, 79–82. [Google Scholar]
- Cabrera, D.; Vignale, B.; Machado, G.; Rodríguez, P.; Zoppolo, R.; Nebel, J.P. Primeras selecciones registradas de guayabo del país en Uruguay. Rev. INIA 2008, 52, 29–32. [Google Scholar]
- Cunda, N. Caracterización de Plantas de Guayabo del País (Acca sellowiana (Berg.) Burret) Desde un Enfoque Frutícola. Bachelor’s Thesis, Tesis Ing. Agr., Universidad de la República, Montevideo, Uruguay, 2006; 98p. [Google Scholar]
- Puppo, M. Prospección y Caracterización de Poblaciones Silvestres de Acca sellowiana (Berg) Burret. (Guayabo del País). Bachelor’s Thesis, Universidad de la República, Montevideo, Uruguay, 2008; 127p. [Google Scholar]
- Quezada, M. Estudio de la Diversidad Genética de una Colección de Acca sellowiana (Berg.) Burret con alto Potencial Agronómico Mediante el uso de Marcadores Moleculares RAPD. Bachelor’s Thesis, Universidad de la República, Montevideo, Uruguay, 2008. [Google Scholar]
- Santos, K.L.; Ducroquet, J.P.H.J.; Nodari, R.O. Caracterização genética de populações naturais de goiabeira serrana (Acca sellowiana) com marcadores microssatélites heterólogos. Biotemas 2011, 24, 75–83. [Google Scholar] [CrossRef][Green Version]
- Baccino, E. Estructura Genética de Cuatro Poblaciones Silvestres de Acca sellowiana (Berg.) Burret Situadas en el Noreste de Uruguay. Bachelor’s Thesis, Universidad de la República, Montevideo, Uruguay, 2011; 77p. [Google Scholar]
- Donazzolo, J. Conservação Pelo Uso e Domesticação da Feijoa na Serra Gaúcha. Ph.D. Thesis, Tese de Doutorado apresentada ao Programa de Pós-Graduação em Recursos Genéticos Vegetais, Universidade Federal de Santa Catarina, Florianópolis, Brazil, 2012; 312p. [Google Scholar]
- Borzuk, L.J.; Saifert, L.; Otalora, J.M.; Sánchez, F.D.; Nodari, R.O. Phenotypic variability in Feijoa fruits (Acca sellowiana (O. Berg.) Burret) on indigenous lands, quilombolas communities and protected areas in the south of Brazil. Rev. Bras. Frutic. 2017, 39, e-699. [Google Scholar] [CrossRef]
- Donazzolo, J.; Stefenon, V.M.; Guerra, M.P.; Nodari, R.O. On farm management of Acca sellowiana (Myrtaceae) as a strategy for conservation of species genetic diversity. Sci. Hortic. 2020, 259. [Google Scholar] [CrossRef]
- Saifert, L.; Sánchez-Mora, F.D.; Borzuk, L.J.; Donazzolo, J.; Freitas da Costa, N.C.; Nunes Ribeiro, H.; Nodari, R.O. Evaluation of the genetic diversity in the feijoa accessions maintained at Santa Catarina, Brazil. Crop Sci. 2020, 60, 345–356. [Google Scholar] [CrossRef]
- Legrand, D. Las Mirtáceas de Uruguay, III; Boletín Facultad de Agronomía: Montevideo, Uruguay, 1968; 80p. [Google Scholar]
- Evia, G.; Gudynas, E. Ecología del Paisaje del Uruguay; Junta de Andalucía, AECI: Sevilla, Spain, 2000; 173p. [Google Scholar]
- Brazeiro, A. Eco-Regiones de Uruguay: Biodiversidad, Presiones y Conservación; Aportes a la Estrategia Nacional de Biodiversidad, Facultad de Ciencias, CIEDUR, VS-Uruguay, SZU: Montevideo, Uruguay, 2015; 122p. [Google Scholar]
- Brown, A.H.D.; Marshall, D.R. A basica sampling strategy: Theory and practice. In Collecting Plant Genetic Diversity: Technical guidelines; Guarino, L., Ramanatha Rao, V., Reid, R., Eds.; Publisher CAB International: Wallingford, UK, 1995; pp. 75–91. [Google Scholar]
- Puppo, M.; Rivas, M.; Franco, J.; Barbieri, R.L. Propuesta de descriptores para Acca sellowiana (Berg.) Burret. Rev. Bras. De Frutic. 2014, 36, 957–970. [Google Scholar] [CrossRef][Green Version]
- Ducroquet, J.P.H.J.; Ribeiro, P.A. Goiabeira serrana: Velha conocida, nova alternativa. Rev. Agropecu. Catarin. 1991, 4, 27–29. [Google Scholar]
- Kader, A. Feijoa, Recommendations of Maintaining Postharvest Quality; University of California: Davis, CA, USA, 2005; Available online: https://postharvest.ucdavis.edu/ (accessed on 20 December 2023).
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Buntjer, J.B. Cross Checker Fingerprint Analysis Software v2.9; Wageningen University and Research Centre: Wageningen, The Netherlands, 1999. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 7 March 2024).
- LeDien, S.; Pagés, J. Analyse factorielle multiple hiérarchique. Rev. Stat. Appl. 2003, 51, 41–73. [Google Scholar]
- Franco, J.; Crossa, J.; Desphande, S. Hierarchical multiple factor analysis for classifying genotypes and forming core subsets based on phenotypic and genetic data. Crop Sci. 2010, 50, 105–117. [Google Scholar] [CrossRef]
- Ward, J.H., Jr. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Jaccard, P. Contribution au problème de l’immigration post-glaciaire de la flore alpine. Bull. Soc. Vaudoise Sci. Nat. 1900, 37, 547–579. [Google Scholar]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package Version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 October 2023).
- Paradis, E.; Schliep, K. Ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferrednfrom metric distances among DNA haplotypes: Application to human mitochondrialDNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Goudet, J.; Jombart, T. Hierfstat: Estimation and Tests of Hierarchical F-Statistics. R Package Version 0.5-11. 2022. Available online: https://CRAN.R-project.org/package=hierfstat (accessed on 15 October 2023).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A. CLUMPAK: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Res. 2015, 15, 1179–1191. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; Vonholdt, B.M. Structure Harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Slatkin, M. Isolation by distance in equilibrium and non-equilibrium populations. Evolution 1993, 47, 264–279. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Mora, F.D.; Saifert, L.; Borsuk, L.J.; Otalora Villamil, J.M.; Nodari, R.O. Avaliação de frutos de goiabeira-serrana cultivados no meio oeste de Santa Catarina, Brasil. Biotemas 2020, 33, 1–9. [Google Scholar] [CrossRef]
- Thorp, T.G. DSIR’s feijoa breeding programme goes to South America. Orchard. N. Z. 1988, 61, 213–215. [Google Scholar]
- Amarante, C.V.T.; Steffens, C.A.; Ducroquet, J.P.H.J.; Sasso, A. Fruit quality of feijoas in response to storage temperature and treatment with 1-methylcyclopropene. Pesqui. Agropecu. Bras. 2008; 43, 1683–1689. [Google Scholar]
- Degenhardt, J.; Orth, A.I.; Guerra, M.P.; Ducroquet, J.P.; Nodari, R.O. Morfología floral da Goiabeira serrana (Feijoa sellowiana) e suas implicações na polinização. Rev. Bras. Frutic. 2001, 23, 718–721. [Google Scholar] [CrossRef]
- Mara, V. Aportes al Conocimiento de la Biología Floral y Reproductiva y Caracterización de Guayabo del País (Acca sellowiana (Berg) Burret). Bachelor’s Thesis, Universidad de la República, Montevideo, Uruguay, 2012; 64p. [Google Scholar]
- Ducroquet, J.P.H.J.; Hickel, E.R. Birds as pollinators of Feijoa (Acca sellowiana Berg). Acta Hortic. 1997, 452, 37–40. [Google Scholar] [CrossRef]
- Finatto, T.; Santos, K.L.; Steiner, N.; Bizzocchi, L.; Holderbaum, D.; Ducroquet, J.P.H.J.; Guerra, M.; Nodari, R.O. Late-acting self-incompatibility in Acca sellowiana (Myrtaceae). Aust. J. Bot. 2011, 59, 53–60. [Google Scholar] [CrossRef]
- Martínez-Vega, R.R.; Fischer, G.; Herrera, A.; Chaves-Córdoba, B.; Quintero, O.C. Características físico-químicas de frutos de feijoa influenciadas por la posición en el canopi. Rev. Col. Cienc. Hortic. 2008, 2, 21–32. [Google Scholar] [CrossRef]
- Parra-Coronado, A.; Fischer, G.; Chaves-Córdoba, B. Tiempo térmico para estados fenológicos reproductivos de la feijoa (Acca sellowiana (O. Berg) Burret). Acta Biol. Colomb. 2015, 20, 163–173. [Google Scholar] [CrossRef]
- Chebataroff, J. La Vegetación del Uruguay y sus Relaciones Fitogeográficas con la del resto de la América del sur. Rev. Geogr. 1942, 2, 49–90. [Google Scholar]
- Soriano, A. “Río de la Plata grasslands,” in Natural Grasslands; Coupland, R.T., Ed.; Elsevier: Amsterdam, The Netherlands, 1991; pp. 367–407. [Google Scholar]
- Bilenca, D.; Miñarro, F. Identificación de Áreas Valiosas de Pastizal (AVPs) en las Pampas y Campos de Argentina, Uruguay y sur de Brasil; Fundación Vida Silvestre Argentina: Buenos Aires, Argentina, 2004; 323p. [Google Scholar]
- Cabrera-Toledo, D.; Mendoza-Galindo, E.; Larranaga, N.; Herrera-Estrella, A.; Vásquez-Cruz, M.; Hernández-Hernández, T. Genomic and Morphological Differentiation of Spirit Producing Agave angustifolia Traditional Landraces Cultivated in Jalisco, Mexico. Plants 2022, 11, 2274. [Google Scholar] [CrossRef]
- Ruiz-Chután, J.A.; Kalousová, M.; Maňourová, A.; Degu, H.D.; Berdúo-Sandoval, J.E.; Villanueva-González, C.E.; Lojka, B. Core Collection Formation in Guatemalan Wild Avocado Germplasm with Phenotypic and SSR Data. Agronomy 2023, 13, 2385. [Google Scholar] [CrossRef]
- Ministerio de Ambiente. Available online: https://www.gub.uy/ministerio-ambiente/politicas-y-gestion/paisaje-protegido-quebrada-cuervos-sierras-del-yerbal-treinta-tres (accessed on 7 March 2024).
- Ministerio de Ambiente. Available online: https://www.gub.uy/ministerio-ambiente/politicas-y-gestion/paisaje-protegido-paso-centurion-sierra-rios-cerro-largo (accessed on 7 March 2024).
- Unesco. Available online: https://www.unesco.org/en (accessed on 7 March 2024).
- Proyecto REDD+ Uruguay. Informe Población Indígena en Uruguay y su Vínculo con el Bosque; Rodríguez, P., Baccino, E., Carro, I., Justo, C., Martino, D., Miguel, C., y Olivera, J., Eds.; Ministerio de Ganadería, Agricultura y Pesca-Ministerio de Vivienda, Ordenamiento Territorial y Medio Ambiente: Montevideo, Uruguay, 2019. [Google Scholar]
- Puppo, M.; Gianotti, C.; Calvete, A.; Leal, A.; Rivas, M. Landscape, agrobiodiversity, and local knowledge in the protected area “Quebrada de los Cuervos y Sierras del Yerbal,” Uruguay. Front. Sustain. Food Syst. 2023, 7, 1240991. [Google Scholar] [CrossRef]
- Ross, S.; Speroni, G.; Souza-Pérez, M.; Ávila, N.; Pietro, F.; González, A.M.; Speranza, P. Stem-cutting anatomy and biochemical responses associated with competence for adventitious root differentiation in Acca sellowiana (Myrtaceae). Trees 2021, 35, 1221–1232. [Google Scholar] [CrossRef]
- Donazzolo, J.; Sanches, T.O.; Bizzocchi, L.; Vilperte, V.; Nodari, R.O. O Armazenamento refrigerado prolonga a viabilidade de sementes de Goiabeira-serrana. Rev. Bras. Frutic. 2015, 37, 748–754. [Google Scholar] [CrossRef]
- Voss, L.C.; Filippi, M.; Sohne, A.E.; Danner, M.A.; Giesel, A.; Donazzolo, J. Classification and quality of Acca sellowiana (O. Berg) Burret seeds as to tolerance to desiccation. Res. Soc. Dev 2020, 9. [Google Scholar] [CrossRef]
- Gomes, J.P.; Oliveira, L.M.; Saldanha, A.P.; Manfredi, S.; Ferreira, P.I. Secagem e Classificação de Sementes de Acca sellowiana (O. Berg) Burret—Myrtaceae quanto à Tolerância à Dessecação e ao Armazenamento. Floresta Ambiente 2013, 20, 207–215. [Google Scholar] [CrossRef]
- Calvete, A. Contribución al Mejoramiento Genético Participativo de Guayabo del País (Acca sellowiana (Berg. Burret)) en el Paisaje Protegido Quebrada de los Cuervos. Bachelor’s Thesis, Universidad de la República, Montevideo, Uruguay, 2013; 86p. [Google Scholar]
- Phan, A.D.T.; Chaliha, M.; Sultanbawa, Y.; Netzel, M.E. Nutritional Characteristics and Antimicrobial Activity of Australian Grown Feijoa (Acca sellowiana). Foods 2019, 8, 376. [Google Scholar] [CrossRef] [PubMed]
- do Amarante, C.V.T.; de Souza, A.G.; Beninca, T.D.T.; Steffens, C.A. Phenolic content and antioxidant activity of the fruit in Brazilian genotypes of feijoa. Pesqui. Agropecu. Bras. 2017, 52, 1223–1230. [Google Scholar] [CrossRef]
- Silveira, A.C.; Oyarzún, D.; Rivas, M.; Záccari, F. Evaluación de la calidad poscosecha en frutos de guayabo del país [Acca sellowiana (Berg) Burret]. Agrocienc. Urug. 2016, 20, 14–21. [Google Scholar] [CrossRef]
- Silveira, A.C.; Záccari, F. An analysis of the postharvest behavior of four genetic materials of feijoa (Acca sellowiana Berg). Rev. Bras. Frutic. 2023, 45, e-974. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphological Descriptor | Code | Unit/Scale | |
|---|---|---|---|
| Leaf | Leaf shape | FORH | nominal scale (3 states) |
| Apex shape | FORAPH | nominal scale (4 states) | |
| Length | LH | mm | |
| Width | AH | mm | |
| Length/Width ratio | LH/AH | Ratio | |
| Flower | Stamens distribution | DEST | nominal scale (2 states) |
| Distance stigma-stamens | DISTEE | mm | |
| Flower (stamens) opening | AFL | mm | |
| Pistil length | LPIST | mm | |
| Fruit | Fruit shape | FFRUT | nominal scale (4 states) |
| Shell hardness | RCAS | nominal scale (6 states) | |
| Position of sepals | SEP | nominal scale (3 states) | |
| Rugosity | RUG | nominal scale (4 states) | |
| Pulp color | COLP | nominal scale (4 states) | |
| Internal color of shell | COLINT | nominal scale (4 states) | |
| Height | AF | mm | |
| Diameter | DF | mm | |
| Weight | PF | g | |
| Pulp yield | RP | % | |
| Pericarp thickness | ECAS | mm | |
| Total soluble solids | SST | °Brix | |
| 100 seed weight | P100 | g | |
| Number of seeds | NSEM | Number | |
| QC | VE | CL | CA | General * | |
|---|---|---|---|---|---|
| AF (mm) | 33.9 | 22.9 | 27.7 | 24.6 | 27.2 |
| (24.6–52.8) | (16.3–29.8) | (20.3–42) | (18.0–33.6) | (16.3–52.8) | |
| DF (mm) | 29.8 | 18.9 | 23.3 | 21.3 | 23.2 |
| (21.2–41.9) | (12.4–26.3) | (17.4–33.9) | (15.1–29.7) | (12.4–41.9) | |
| PF (g) | 18.1 | 5.2 | 8.8 | 7.2 | 9.8 |
| (7.9–51.8) | (1.9–10.7) | (3.7–25.4) | (2.4–16.7) | (1.9–51.8) | |
| RP (%) | 36.3 | 35.6 | 39.0 | 30.7 | 35.3 |
| (23.6–68.5) | (11.7–68.8) | (30.8–59.7) | (24.1–44.0) | (11.7–68.8) | |
| ECAS (mm) | 3.2 | 2.0 | 2.3 | 2.9 | 3 |
| (1.3–6.7) | (1.1–2.7) | (1.2–4.0) | (1.3–5.1) | (1.1–6.7) | |
| SST (°Brix) | 14.3 | 18.2 | 14.7 | 13.1 | 15.1 |
| (11.8–16.9) | (11.2–23.6) | (11.0–19.0) | (10.6–15.5) | (10.6–23.6) | |
| NSEM (number) | 41.0 | 55.1 | 61.4 | 29.8 | 46.4 |
| (17.0–72.0) | (20.0–98.0) | (39.0–114.0) | (15.0–52.0) | (14.9–113.7) | |
| P100 (g) | 0.22 | 0.14 | 0.15 | 0.19 | 0.18 |
| (0.16–0.33) | (0.08–0.19) | (0.10–0.26) | (0.10–0.31) | (0.08–0.33) | |
| LH (mm) | 42.3 | 50.4 | 49.3 | 39.7 | 45.6 |
| (33.8–56.5) | (40.5–69.5) | (35.5–62.1) | (30.6–47.2) | (30.6–69.5) | |
| AH (mm) | 25.4 | 29.2 | 28.0 | 24.6 | 26.9 |
| (21.2–31.1) | (23.1–39.1) | (21.2–33.7) | (19.5–27.9) | (19.5–39.1) | |
| LH/AH (ratio) | 1.70 | 1.70 | 1.80 | 1.60 | 1.70 |
| (1.47–1.95) | (1.53–2.08) | (1.57–2.02) | (1.45–1.79) | (1.45–2.08) | |
| DISTEE (mm) | 3.4 | 4.4 | 3.8 | 3.2 | 3.7 |
| (0.5–7.3) | (1.3–7.8) | (0.5–7.6) | (−5.2–6.5) | (−5.2–7.8) | |
| AFL (mm) | 3.1 | 2.4 | 2.1 | 1.5 | 2.3 |
| (2.3–3.9) | (1.6–3.7) | (1.6–2.8) | (1.1–1.8) | (1.1–3.9) | |
| LPIST (mm) | 2.6 | 2.3 | 2.3 | 2.1 | 2.3 |
| (2.3–3.5) | (2.2–2.4) | (2.1–2.5) | (1.8–2.3) | (1.8–3.5) |
| Classes/States | Quebrada de los Cuervos (%) | Valle Edén (%) | Cuchilla de Laureles (%) | Cañitas (%) | Total Population (%) | Chi Square | |
|---|---|---|---|---|---|---|---|
| FFRUT | round | 39.1 | 26.9 | 11.1 | 35 | 29 | 0.0192 |
| oval | 56.5 | 46.2 | 72.2 | 65 | 59 | ||
| oblong | 4.4 | 26.9 | 16.7 | 0 | 13 | ||
| SEP | open | 39.1 | 51.9 | 83.3 | 68.4 | 59 | 0.0051 |
| semi-open | 34.8 | 40.7 | 11.1 | 5.3 | 25 | ||
| erect | 26.1 | 7.4 | 5.6 | 26.3 | 16 | ||
| RUG | smooth | 20.8 | 18.5 | 22.2 | 25 | 21 | 0.0795 |
| somewhat rough | 50 | 70.4 | 77.8 | 70 | 66 | ||
| rough | 29.2 | 11.1 | 0 | 5 | 12 | ||
| COLP 1 | greenish white 3 | 0 | 4 | 11.1 | 0 | 3 | <0.0001 |
| cream white 3 | 37.5 | 92 | 55.6 | 80 | 67 | ||
| amber white 2 | 58.3 | 0 | 11.1 | 10 | 21 | ||
| naples yellow 4 | 4.2 | 4 | 22.2 | 10 | 9 | ||
| COLINT 1 | amber white 4 | 58 | 22 | 22 | 21 | 32 | <0.0001 |
| naples yellow 1 | 9 | 0 | 0 | 21 | 7 | ||
| naples yellow 4 | 33 | 56 | 78 | 58 | 55 | ||
| honey yellow 4 | 0 | 22 | 0 | 0 | 7 | ||
| RCAS | soft | 9 | 19 | 11 | 16 | 14 | 0.0491 |
| medium-soft | 9 | 7 | 6 | 0 | 6 | ||
| medium | 34 | 41 | 83 | 42 | 48 | ||
| medium-hard | 13 | 7 | 0 | 11 | 8 | ||
| hard | 26 | 22 | 0 | 31 | 21 | ||
| very hard | 9 | 4 | 0 | 0 | 3 | ||
| FORH | oval | 2 | 0 | 2 | 6.1 | 3 | 0.0264 |
| obovate | 95.9 | 90.4 | 88 | 93.9 | 92 | ||
| elliptic | 2 | 9.6 | 10 | 0 | 11 | ||
| FORAPH | rounded | 46.9 | 73.1 | 68 | 61.2 | 63 | 0.0381 |
| acute | 0 | 0 | 2 | 0 | 1 | ||
| obtuse | 44.9 | 26.9 | 30 | 36.7 | 35 | ||
| emarginated | 8.2 | 0 | 0 | 2 | 3 | ||
| DEST | radial | 7.4 | 86.1 | 88.6 | 47.1 | 65 | <0.0001 |
| random | 92.6 | 13.9 | 11.4 | 52.9 | 35 |
| Source of Variation | df | Sum of Squares | Variance Components | Percentage of Variation |
|---|---|---|---|---|
| Between populations | 3 (P−1) | 808.2538 | 7.143329 | 34.75% |
| Within populations | 140 (n−P) | 1878.1820 | 13.415585 | 65.25% |
| Total | 143 (n−1) | 2686.4358 | 20.558914 |
| VE | CA | CL | QC | |
|---|---|---|---|---|
| VE | 0.664 | 0.763 | 0.760 | |
| CA | 0.430 | 1.335 | 1.016 | |
| CL | 0.396 | 0.272 | 1.583 | |
| QC | 0.397 | 0.330 | 0.240 | |
| Specific Fst | 0.598 | 0.371 | 0.213 | 0.185 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivas, M.; Puppo, M.; Baccino, E.; Quezada, M.; Franco, J.; Pritsch, C. Phenotypic and Molecular Diversity of Wild Populations of Acca sellowiana (Berg.) Burret in the Southern Area of Natural Distribution. Horticulturae 2024, 10, 360. https://doi.org/10.3390/horticulturae10040360
Rivas M, Puppo M, Baccino E, Quezada M, Franco J, Pritsch C. Phenotypic and Molecular Diversity of Wild Populations of Acca sellowiana (Berg.) Burret in the Southern Area of Natural Distribution. Horticulturae. 2024; 10(4):360. https://doi.org/10.3390/horticulturae10040360
Chicago/Turabian StyleRivas, Mercedes, María Puppo, Estela Baccino, Marianella Quezada, Jorge Franco, and Clara Pritsch. 2024. "Phenotypic and Molecular Diversity of Wild Populations of Acca sellowiana (Berg.) Burret in the Southern Area of Natural Distribution" Horticulturae 10, no. 4: 360. https://doi.org/10.3390/horticulturae10040360
APA StyleRivas, M., Puppo, M., Baccino, E., Quezada, M., Franco, J., & Pritsch, C. (2024). Phenotypic and Molecular Diversity of Wild Populations of Acca sellowiana (Berg.) Burret in the Southern Area of Natural Distribution. Horticulturae, 10(4), 360. https://doi.org/10.3390/horticulturae10040360