
Seed Morphological Analysis in Species of Vitis and Relatives

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Seeds of Vitis Species and Relatives
2.2. Photography
2.3. General Morphological Descriptors
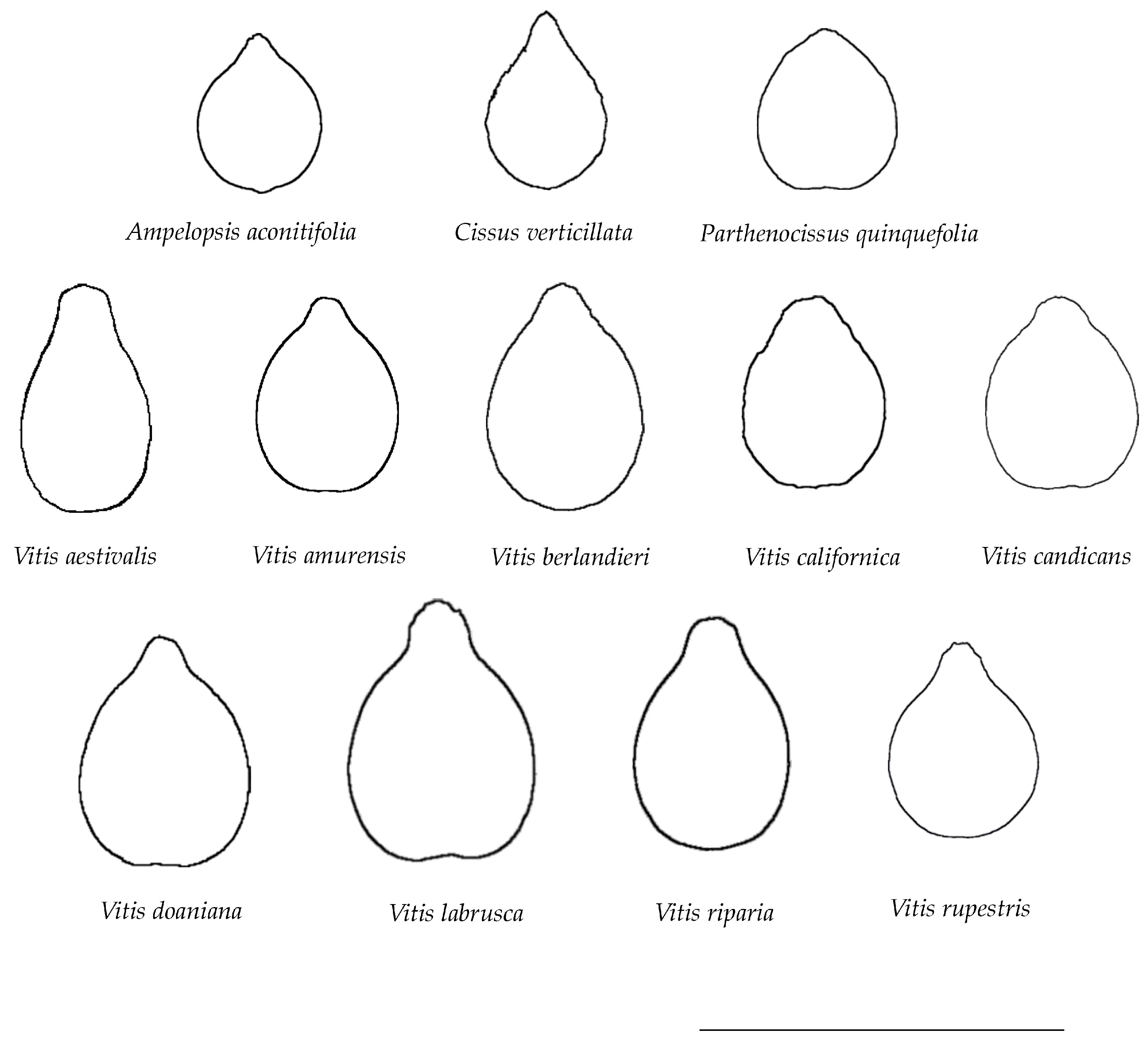
2.4. Average Silhouettes for Each Species
2.5. Quantification of Seed Shape by the J Index

2.6. Symmetry Analysis
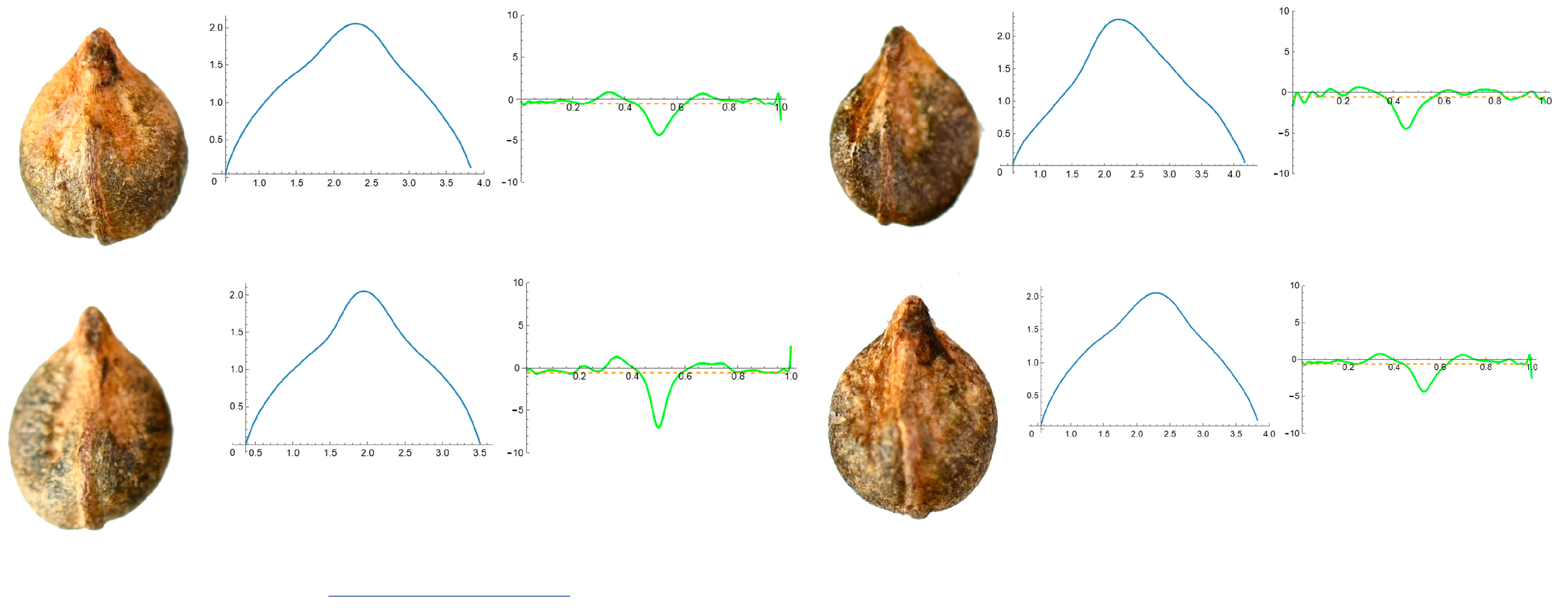
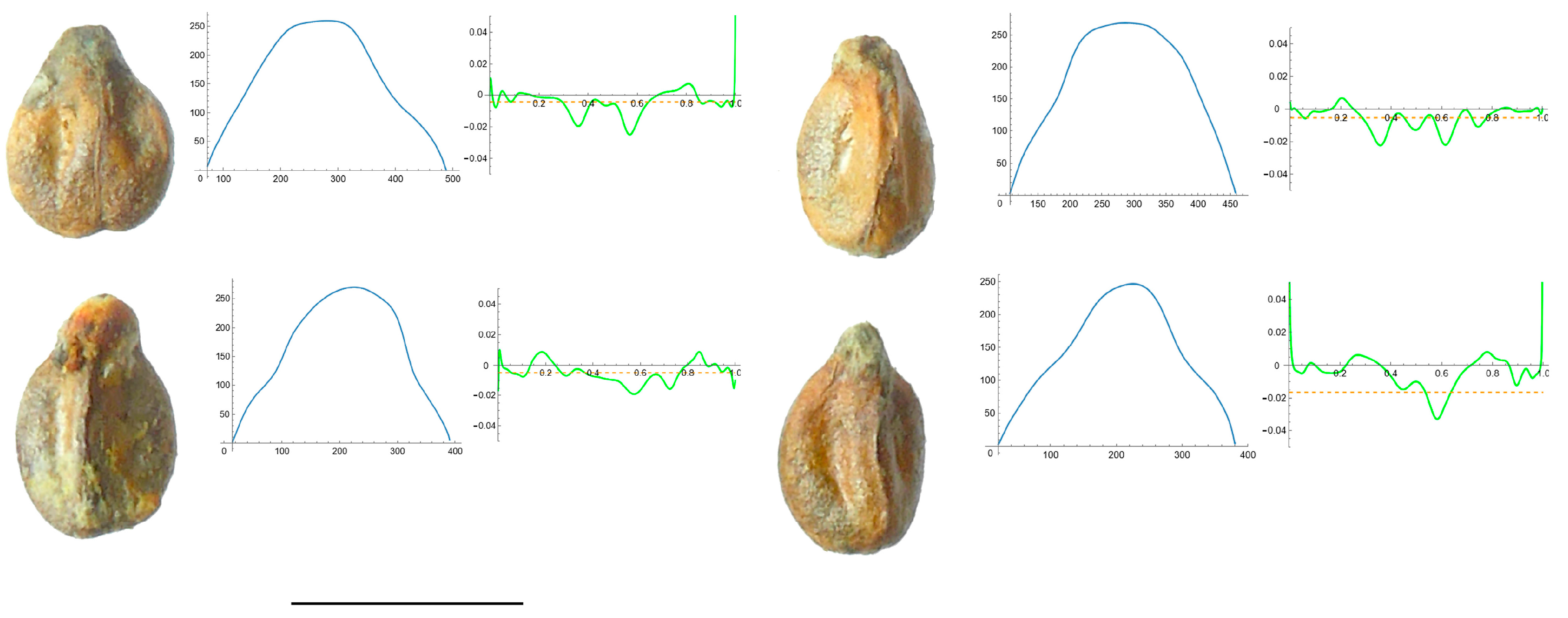
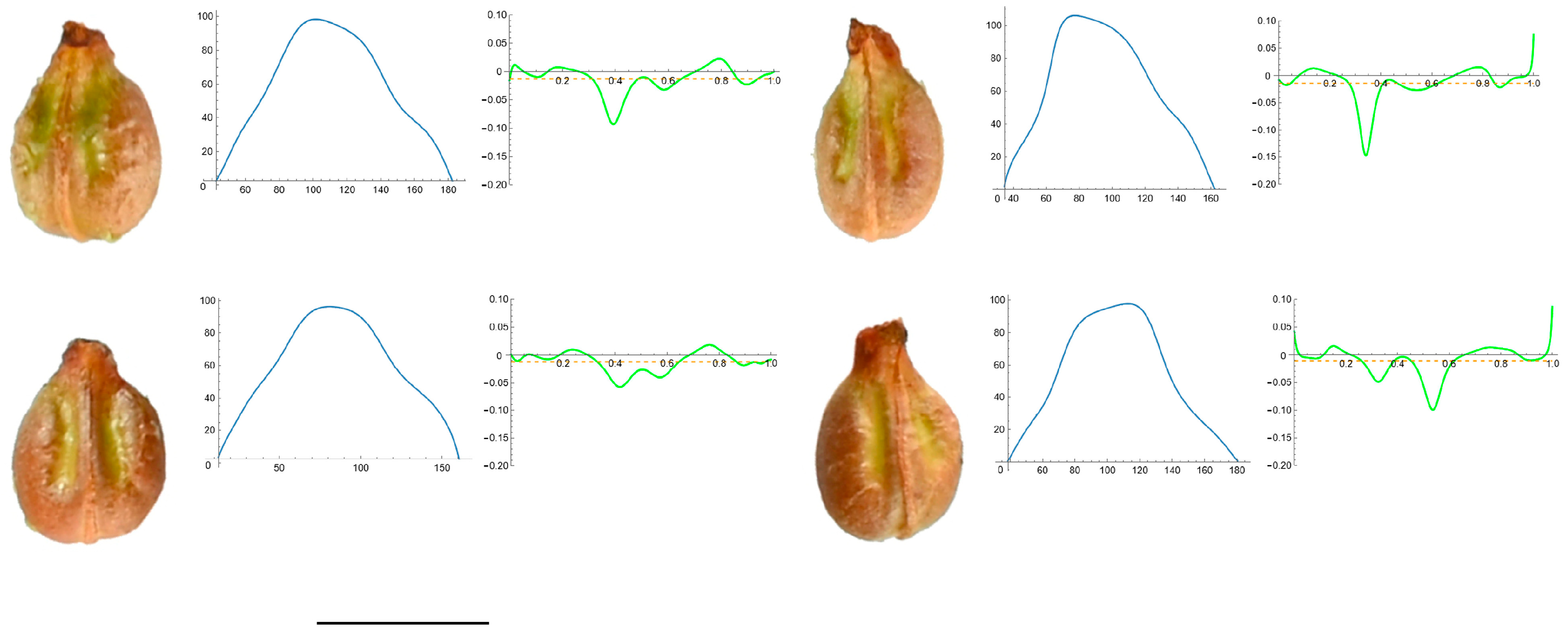
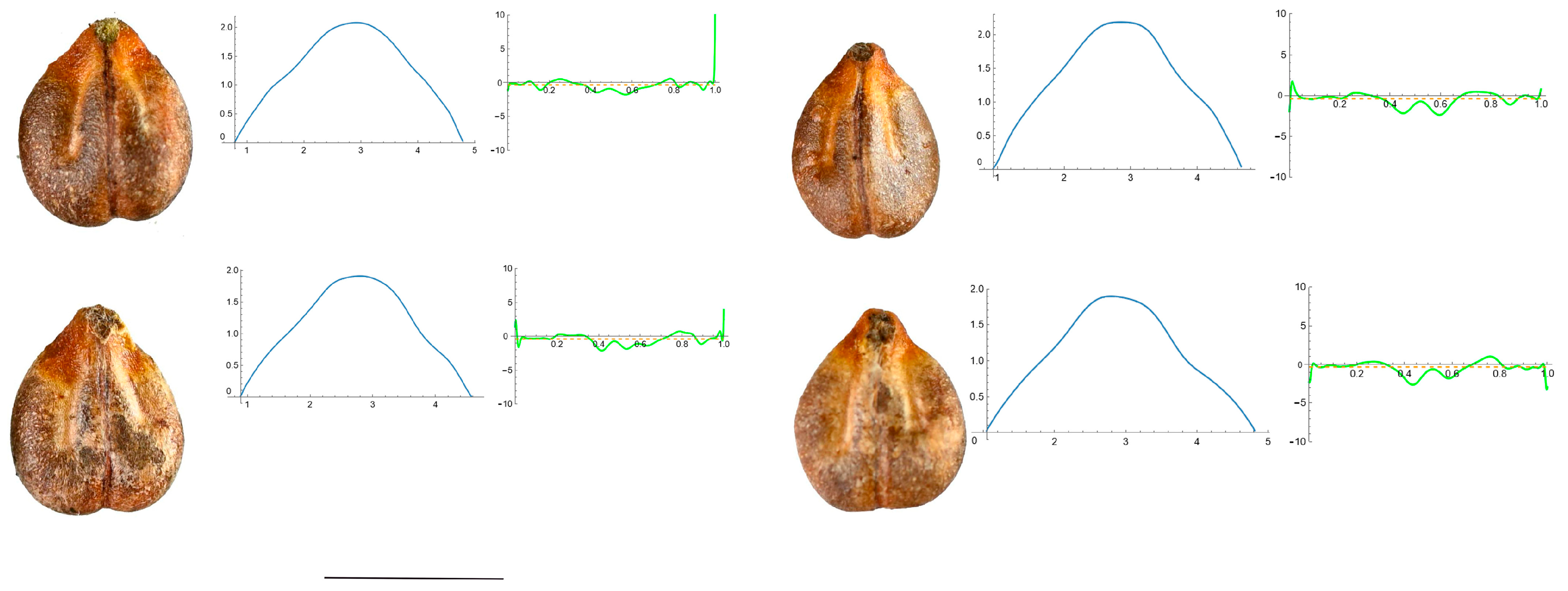
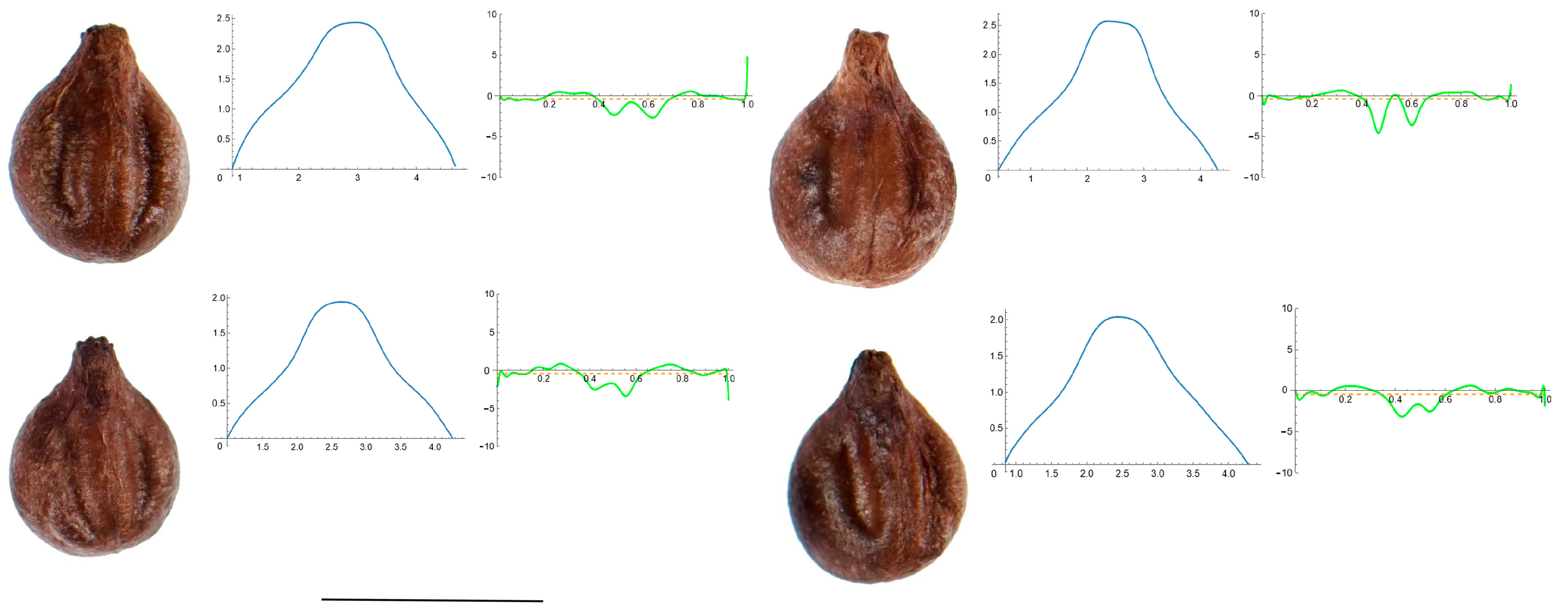
2.7. Curvature Analysis
2.8. Statistical Analysis
3. Results
3.1. General Morphological Measurements
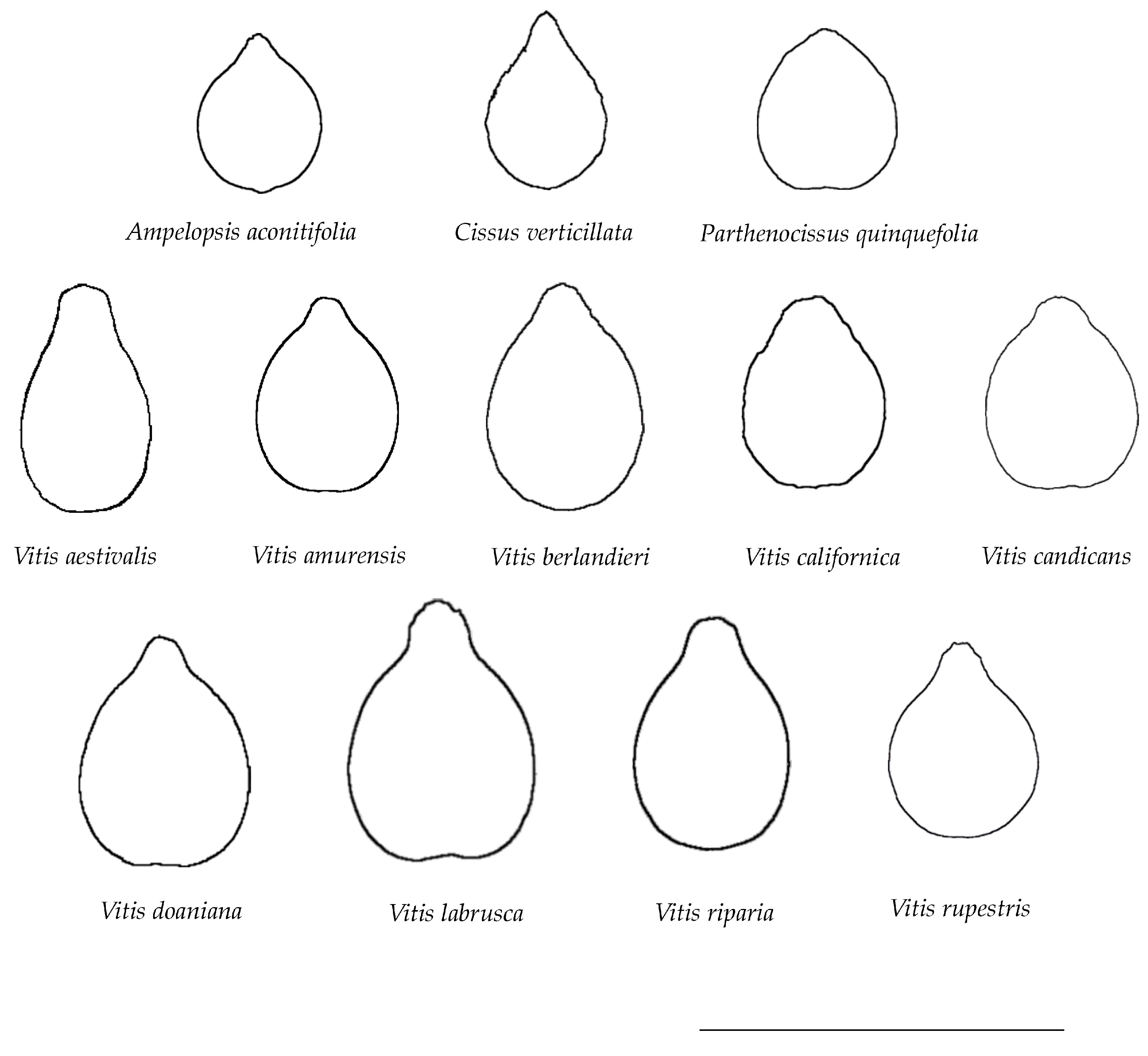
3.2. Average Silhouettes of Representative Species
3.3. Comparison of Seed Shape between Species by the J Index
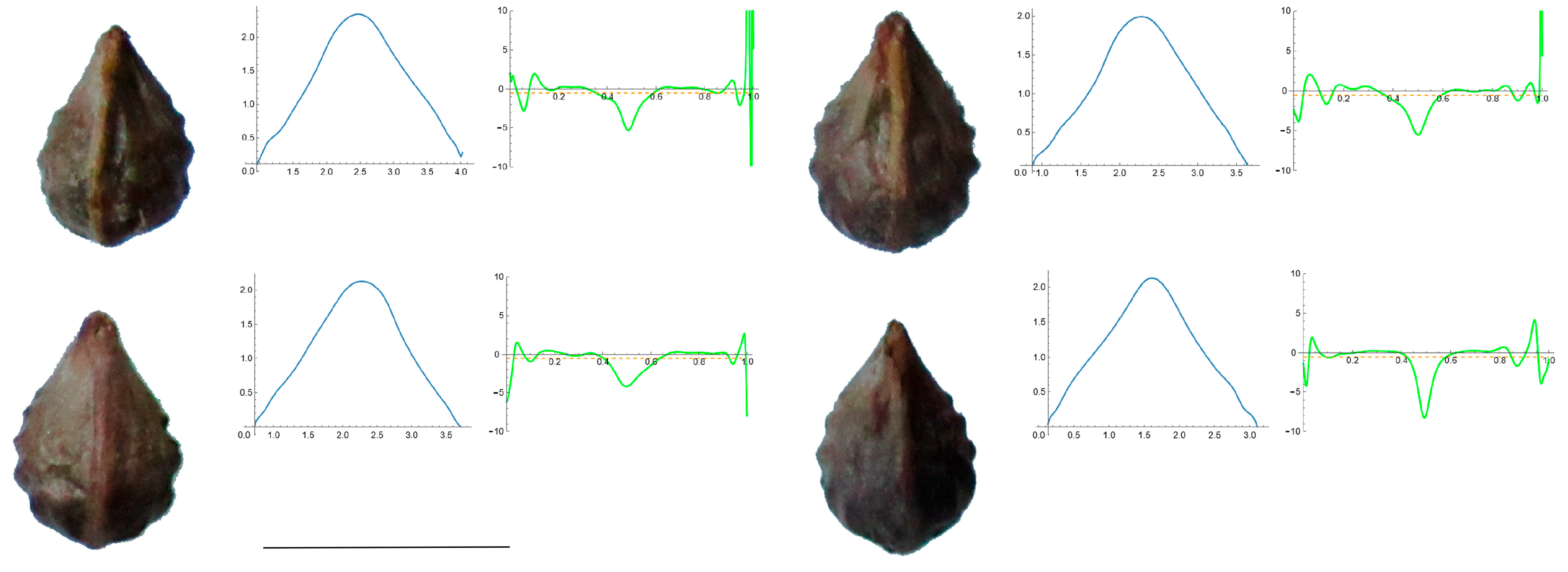
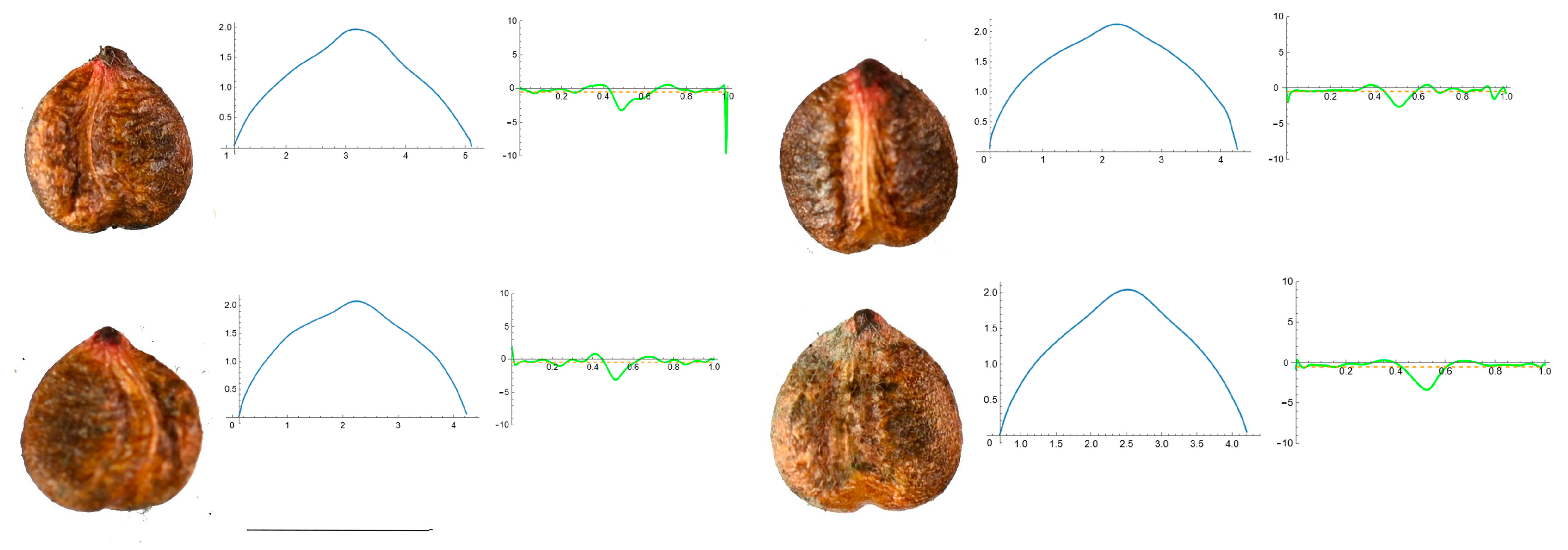
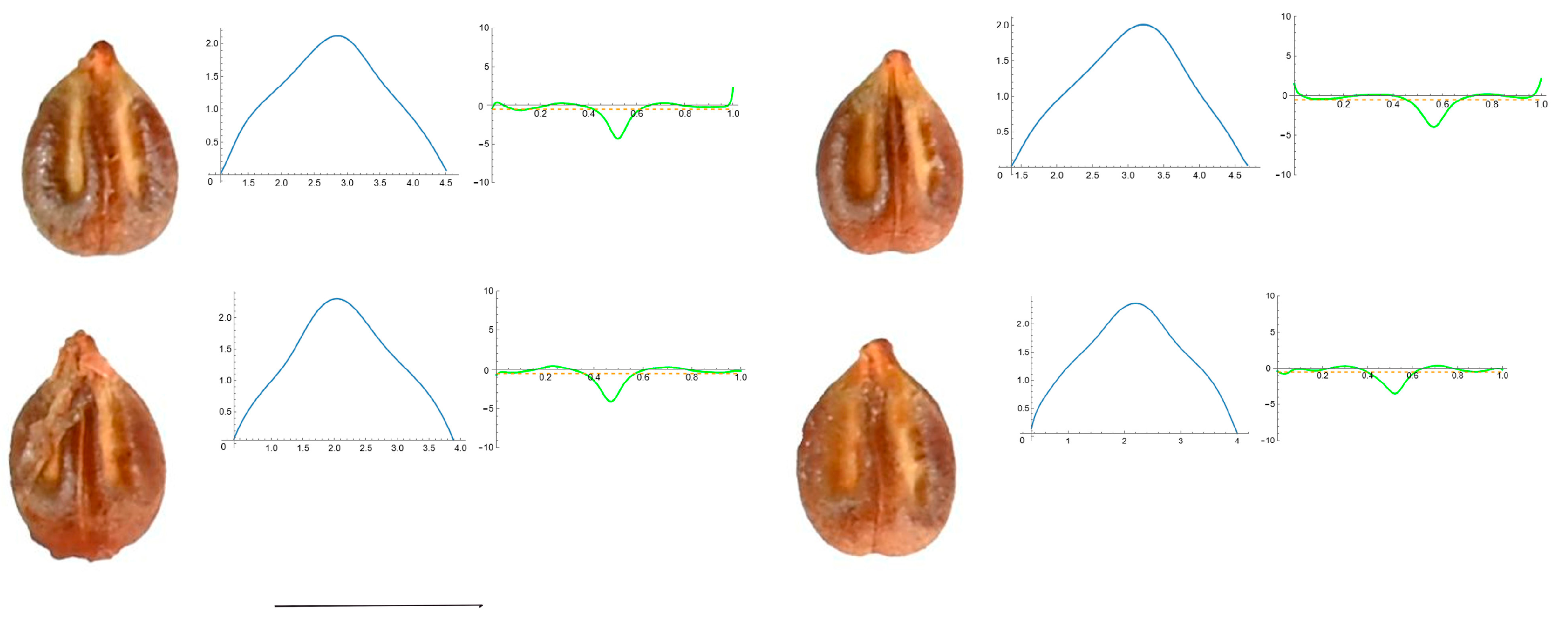
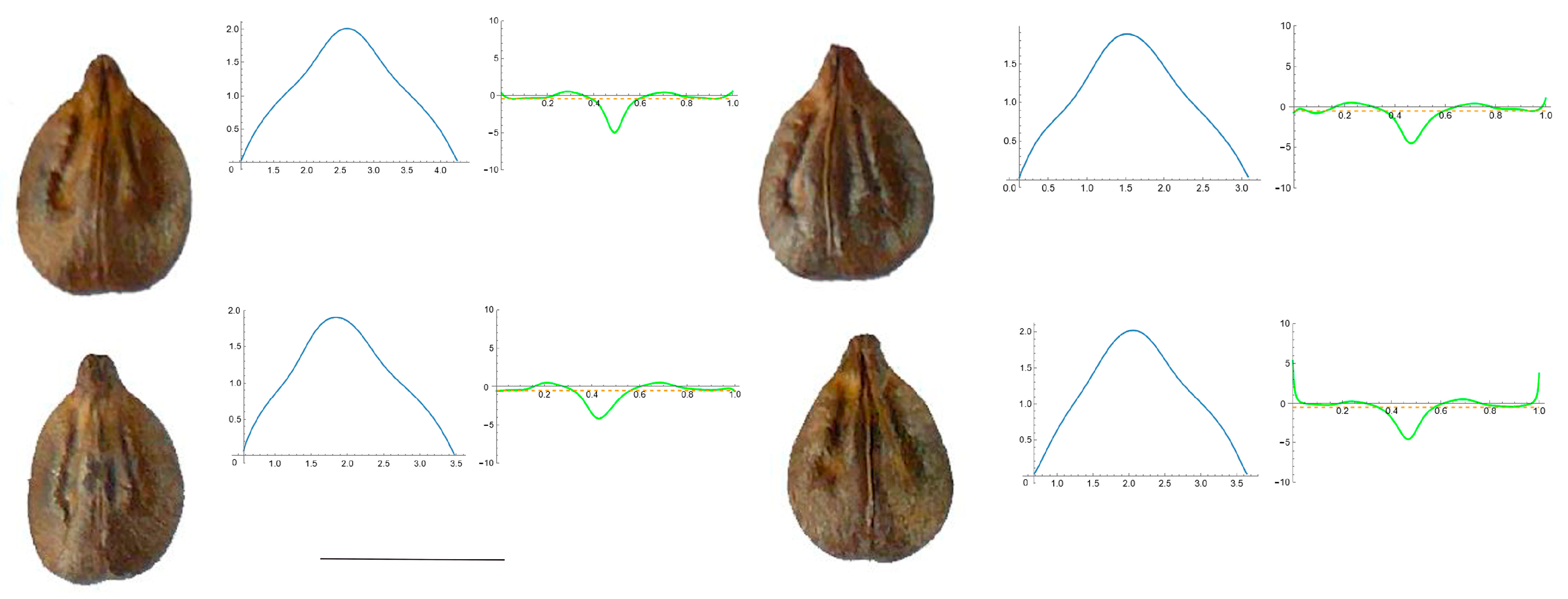
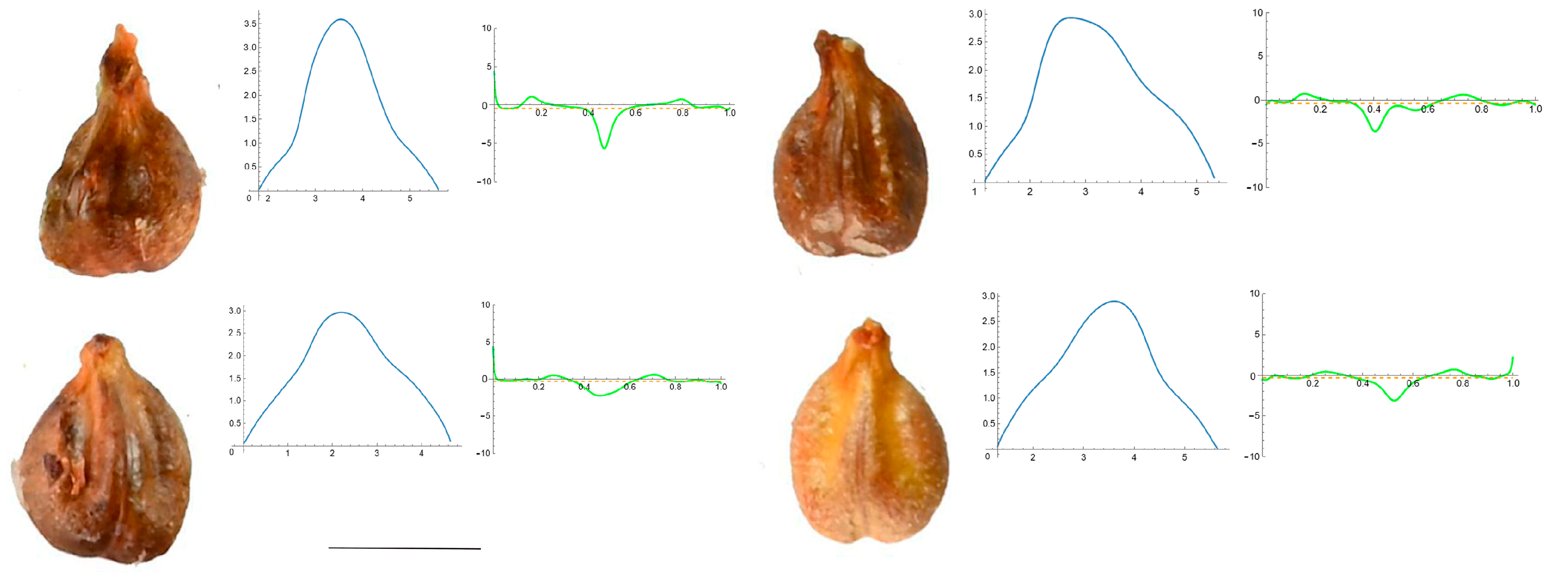
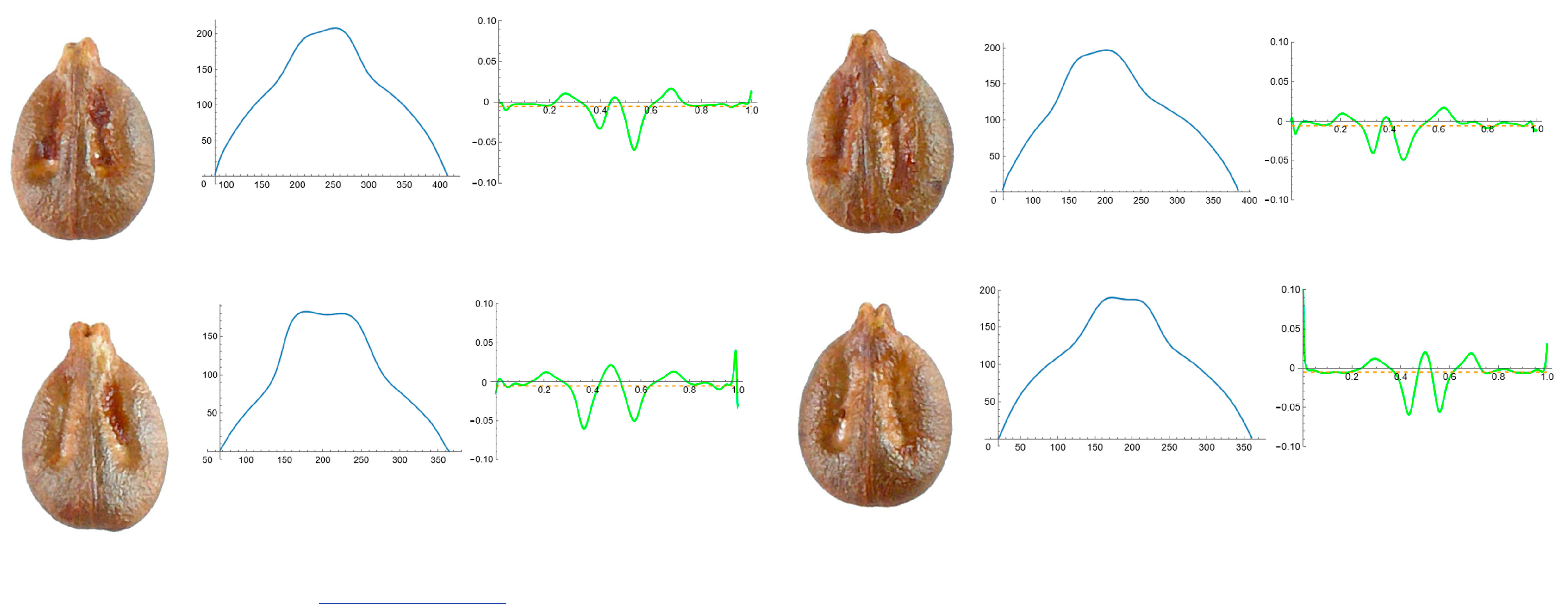
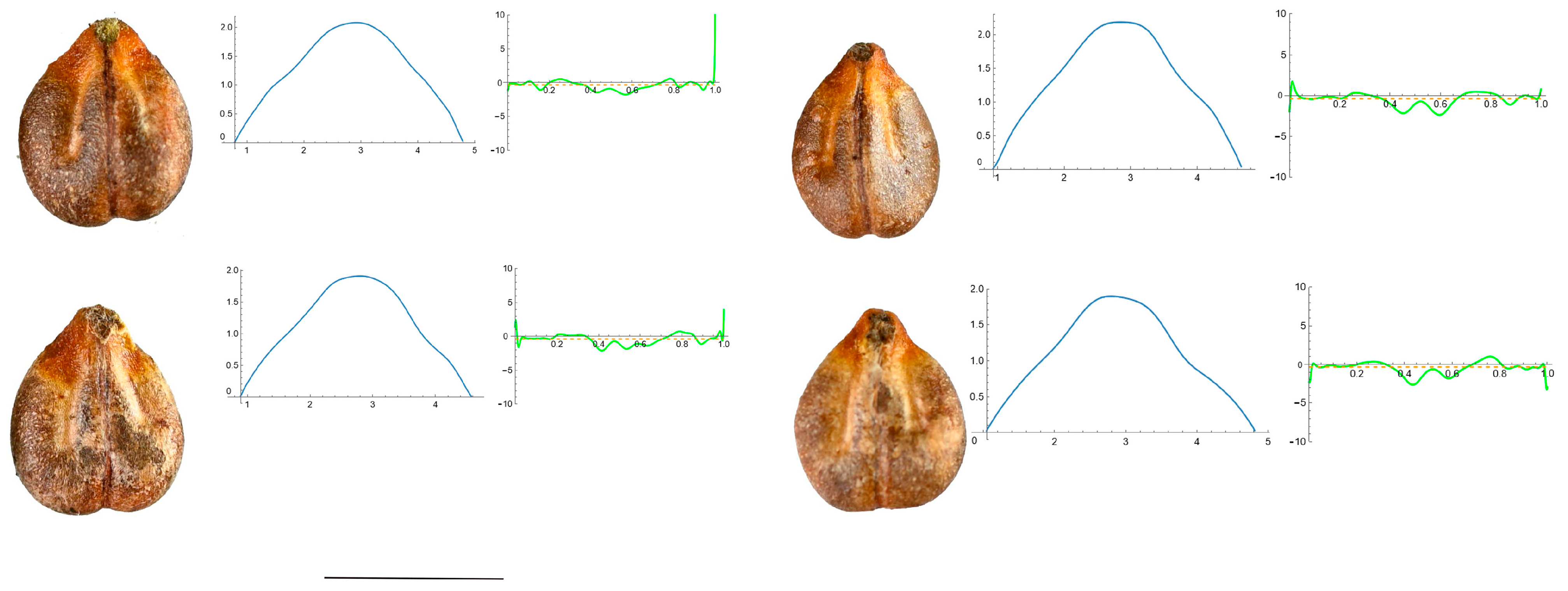
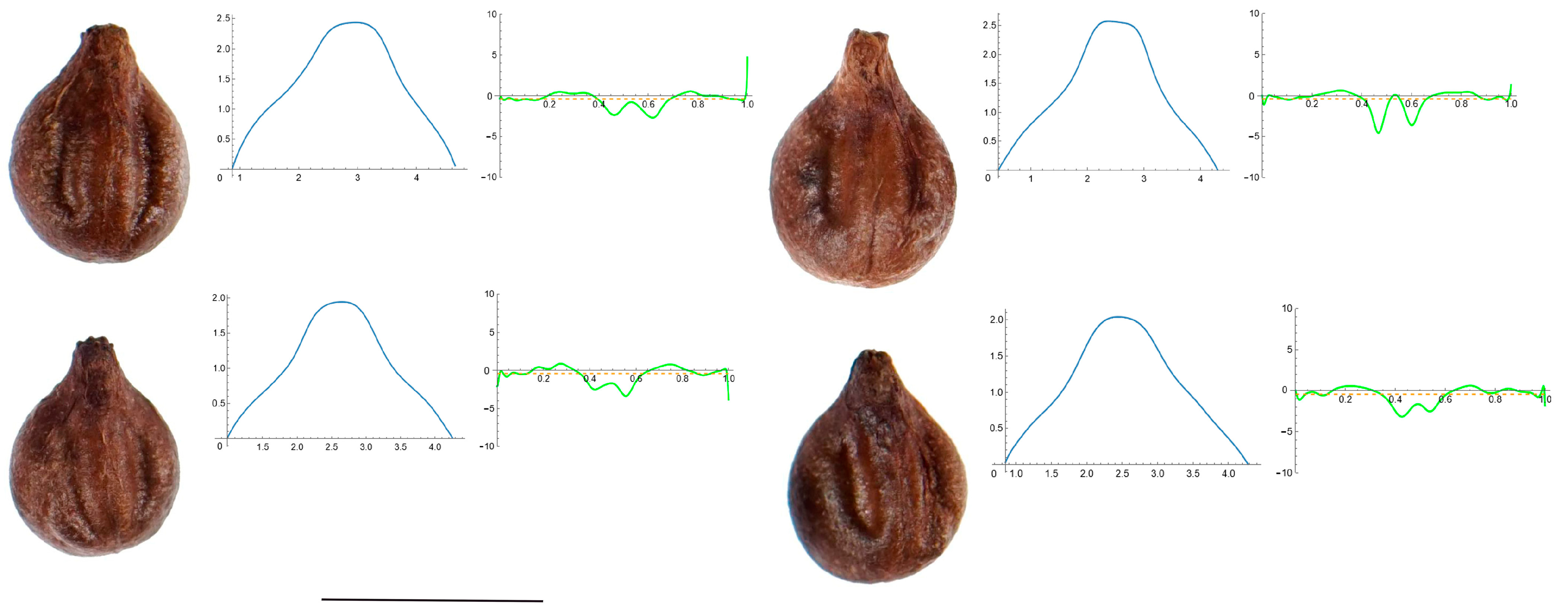
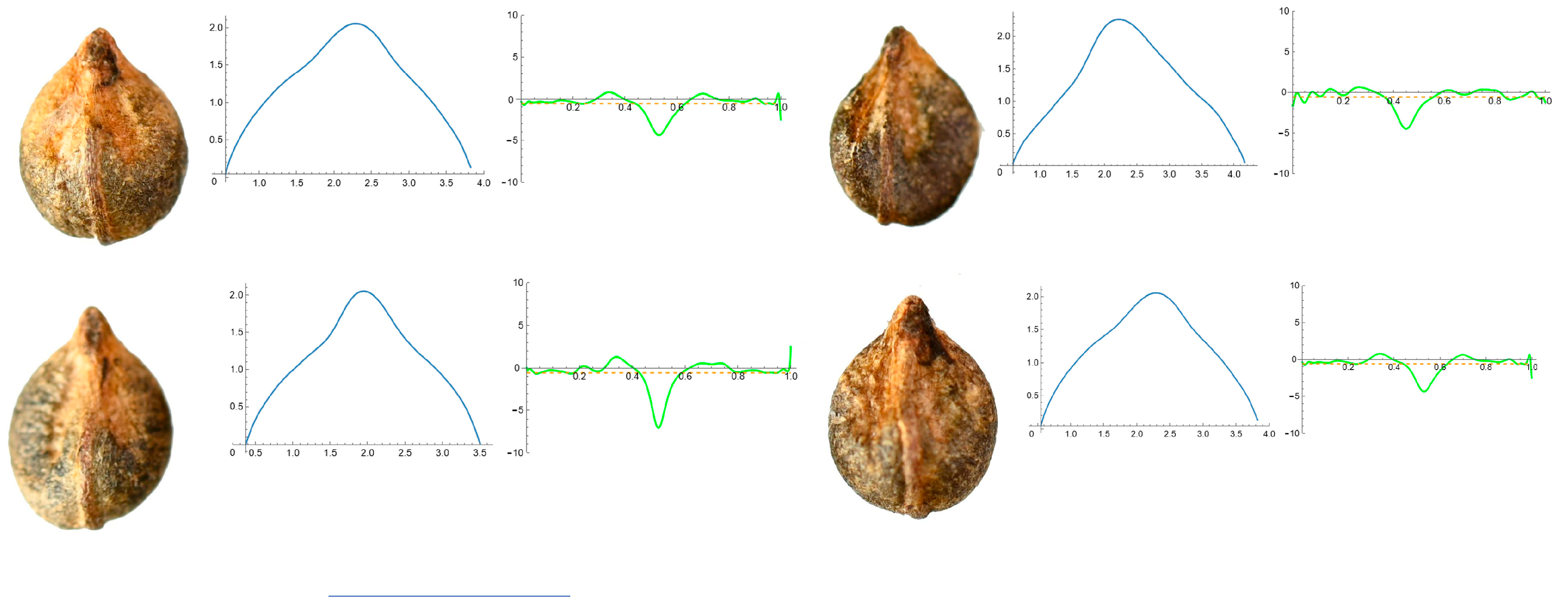
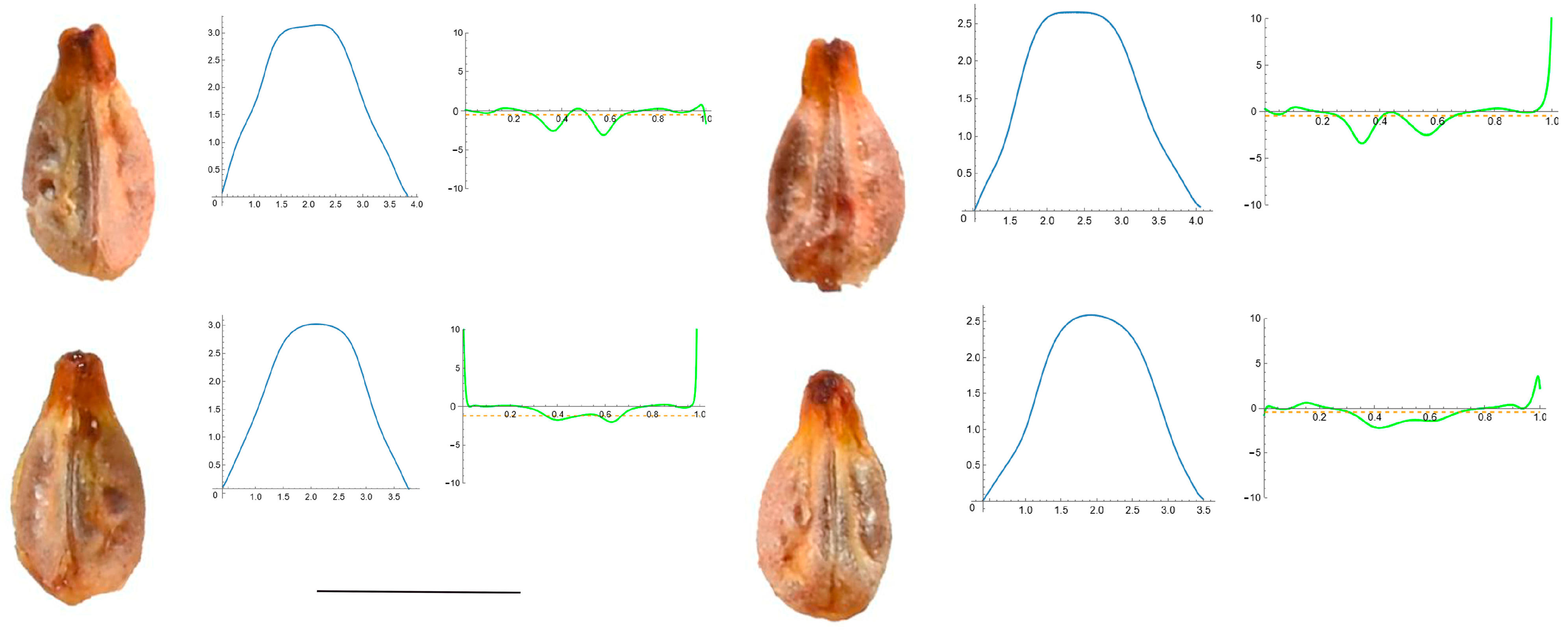
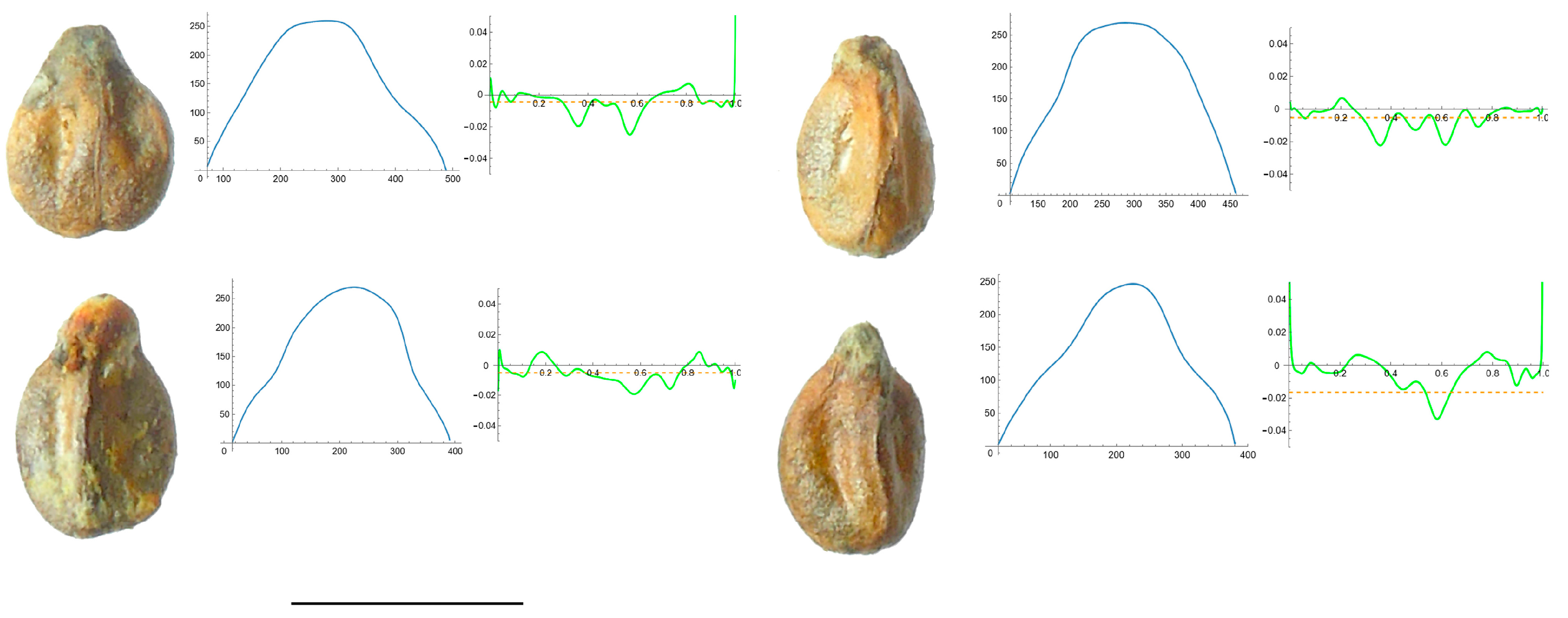
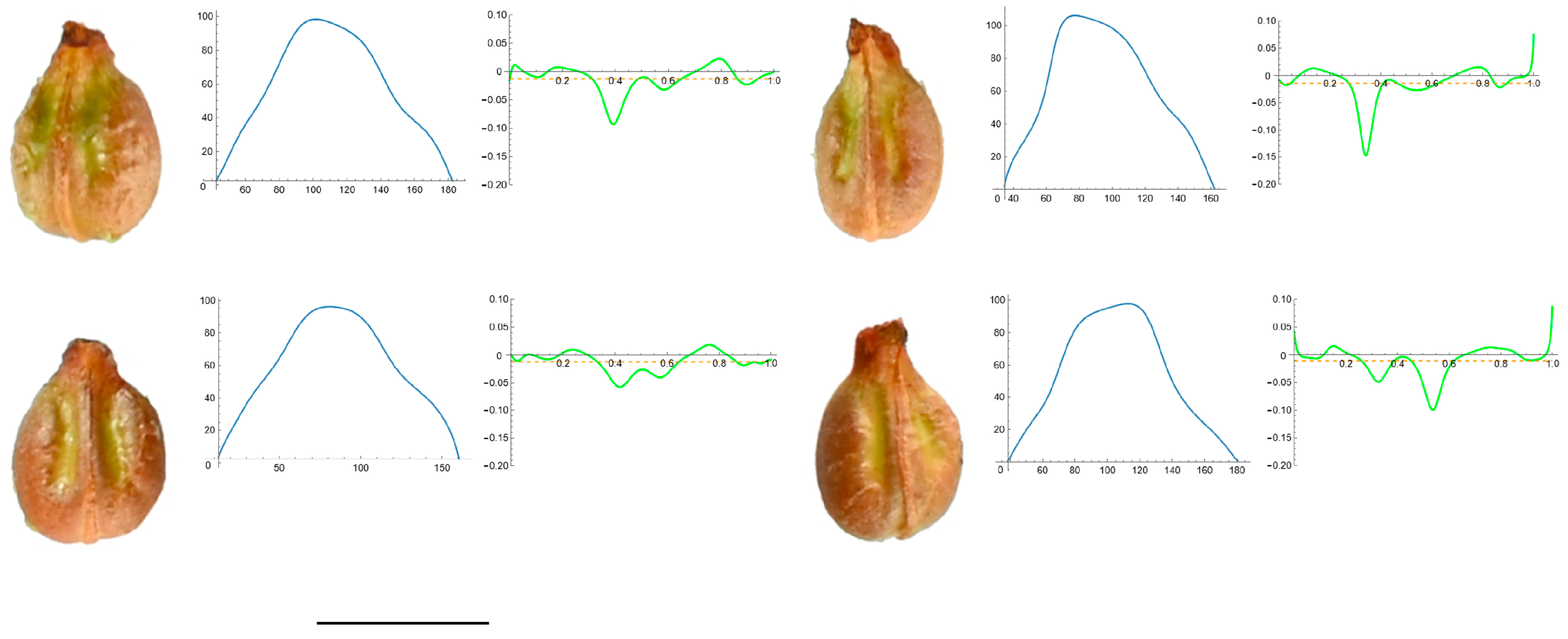
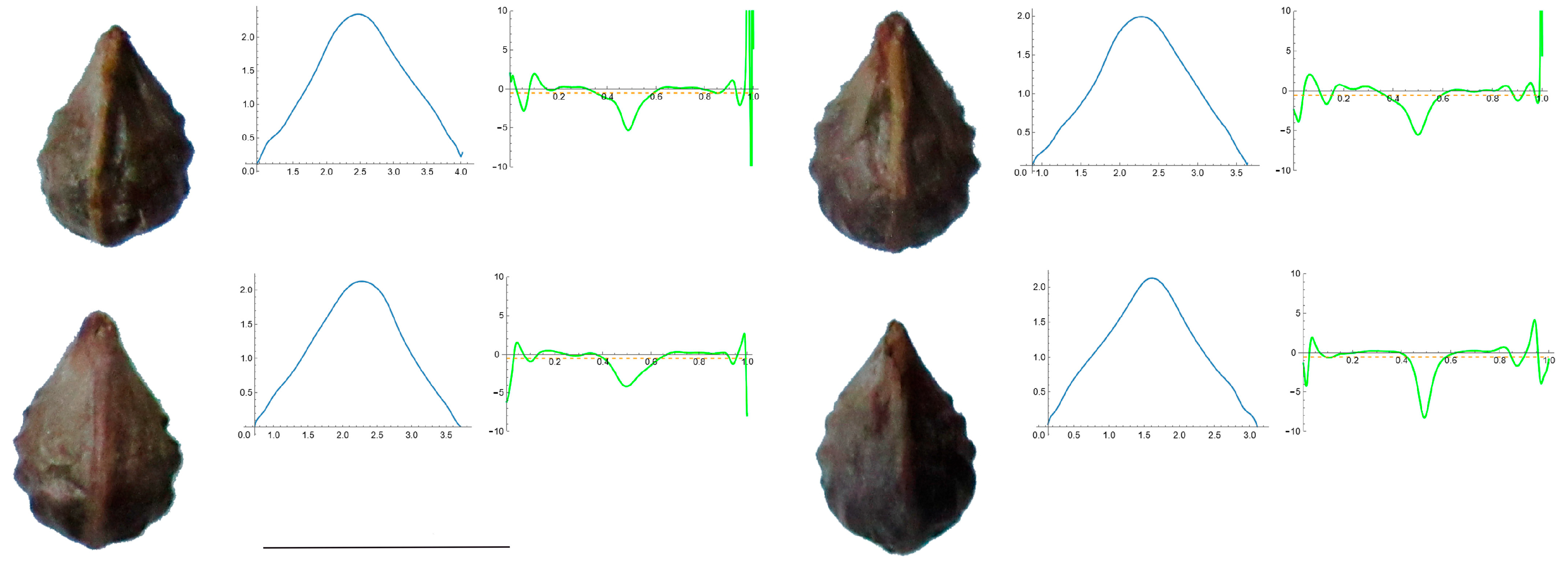
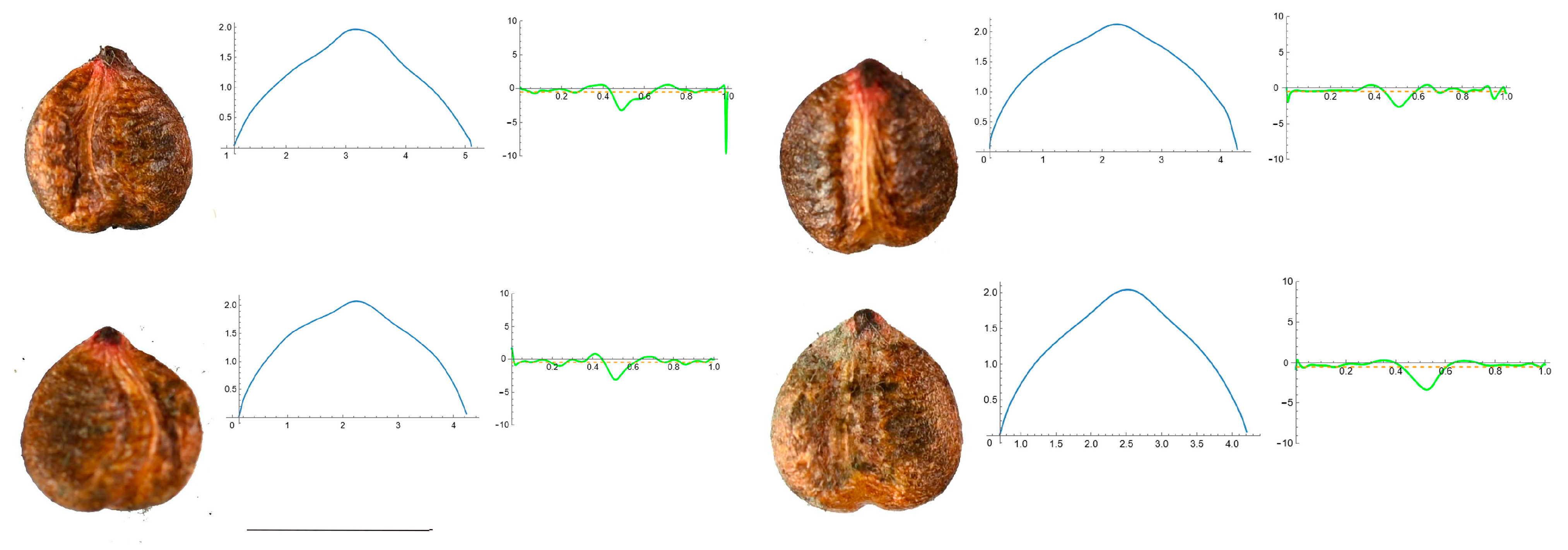
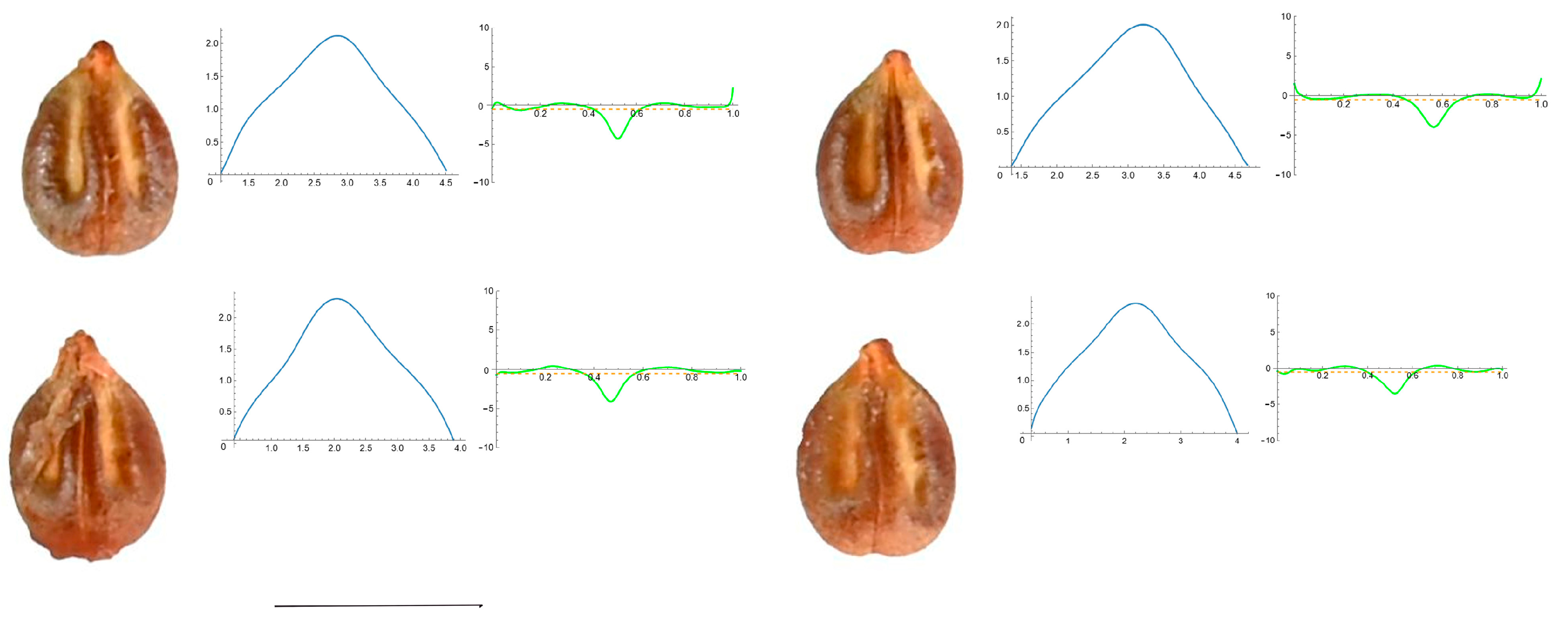
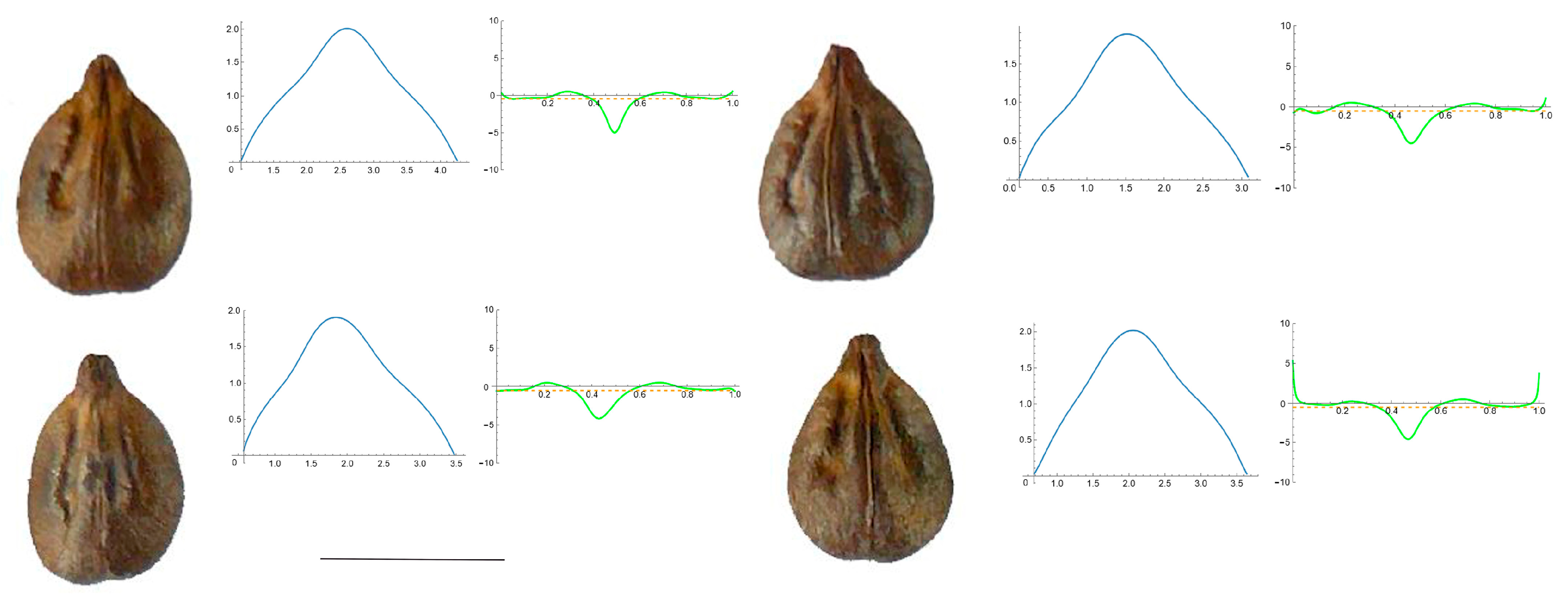
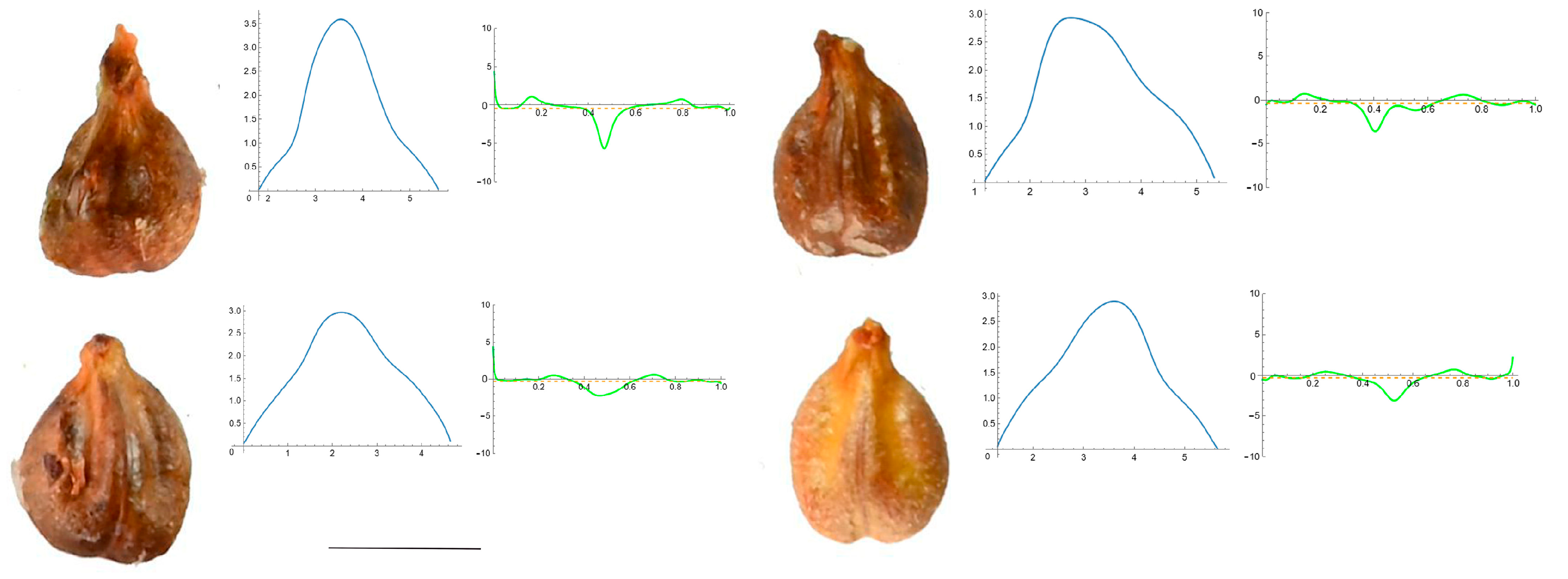
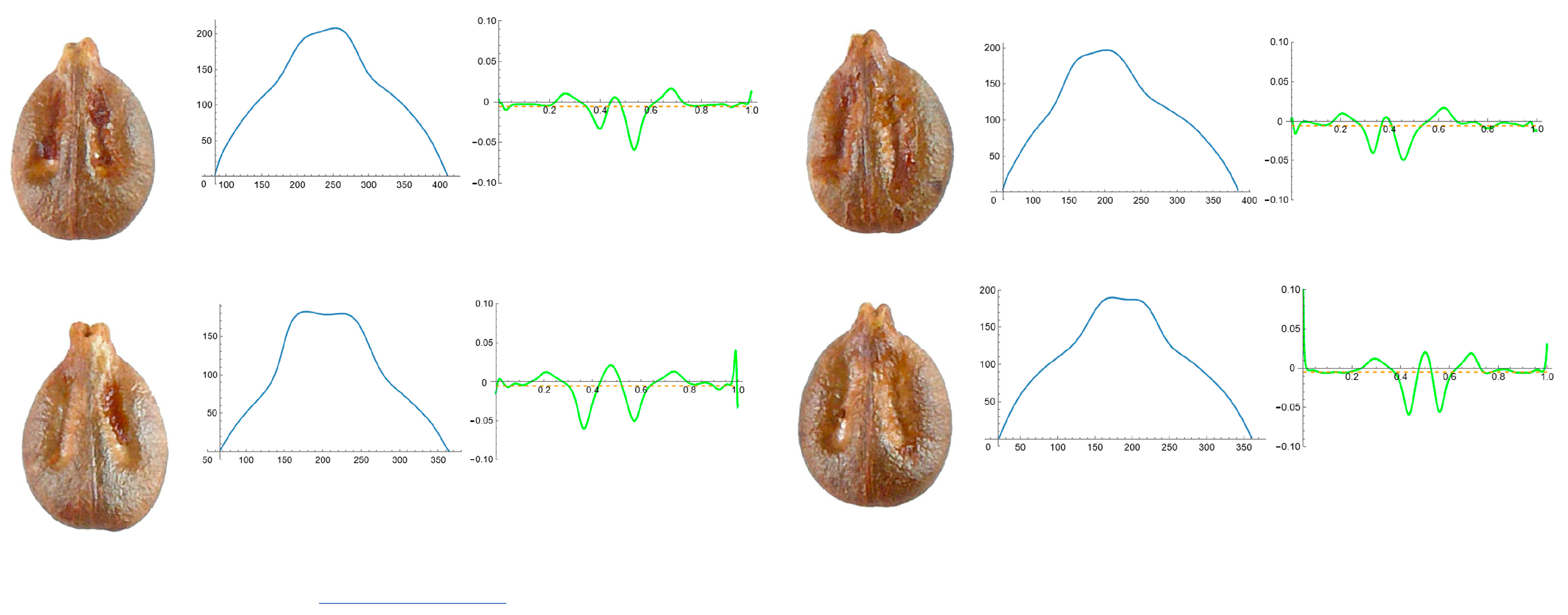
3.4. Curvature Analysis
3.4.1. Curvature Measurements
3.4.2. Seed Morphological Analysis
Section 1: Genera and Species with One Point of Maximum Curvature at Their Seed Apex (Acute)
Section 2: Species with Two Points of Maximum Curvature at Their Apex (Plane)
Section 3: Species of an Intermediate or Mixed Type at Their Apex (Acute and Plane)
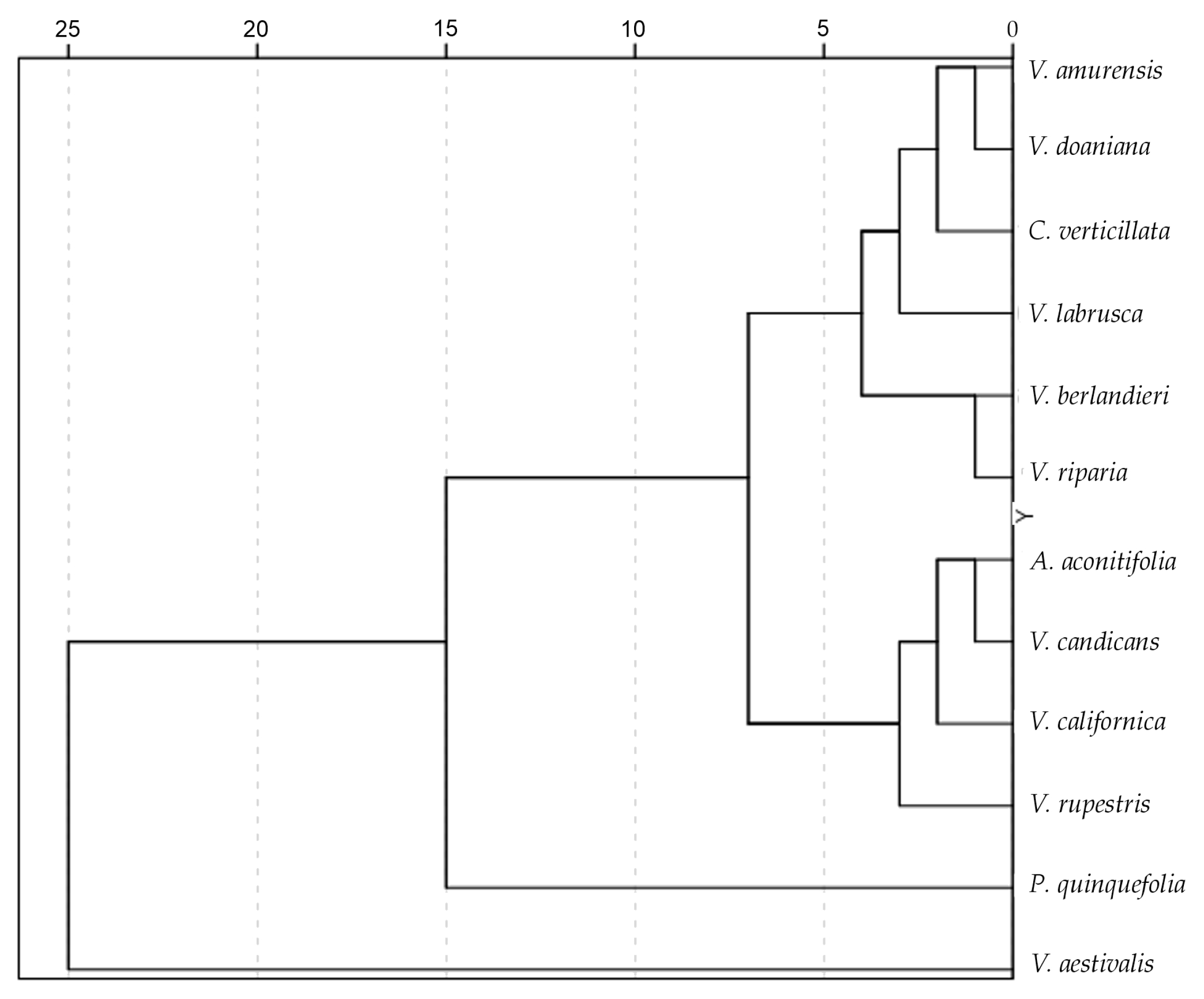
3.5. Symmetry Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
References
- FAO. Food and Agriculture Organization of the United Nations. Available online: http://faostat.fao.org (accessed on 18 February 2024).
- OIV. Office International de la Vigne et du Vin. Available online: http://www.oiv.int/ (accessed on 18 February 2024).
- Vivier, M.A.; Pretorius, I.S. Genetically tailored grapevines for the wine industry. Trends Biotechnol. 2002, 20, 472–478. [Google Scholar] [CrossRef]
- Wen, J. Vitaceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 9, pp. 467–479. [Google Scholar]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet. 2023. Available online: http://www.plantsoftheworldonline.org/ (accessed on 18 February 2024).
- Arrigo, N.; Arnold, C. Naturalised Vitis Rootstocks in Europe and consequences to native wild grapevine. PLoS ONE 2007, 2, e521. [Google Scholar] [CrossRef]
- Pipia, I.; Gogniashvili, M.; Tabidze, V.; Beridze, T.; Gamkrelidze, M.; Gotsiridze, V.; Melyan, G.; Musayev, M.; Salimov, V.; Beck, J.; et al. Plastid DNA sequence diversity in wild grapevine samples (Vitis vinifera subsp. sylvestris) from the Caucasus region. Vitis 2012, 51, 119–124. [Google Scholar]
- Laguna Lumbreras, E. Sobre las formas naturalizadas de Vitis L. (Vitaceae) en la Comunidad Valenciana, I. Especies. Flora Montiberica 2003, 23, 46–82. [Google Scholar]
- Jacquat, C.H.; Martinoli, D. Vitis vinifera L.: Wild or cultivated? Study of the grape pips found at Petra, Jordan; 150 B.C.-A.D. 40. Veget. Hist. Archaeobot. 1999, 8, 25–30. [Google Scholar] [CrossRef]
- Gitea, M.A.; Bungau, S.G.; Gitea, D.; Pasca, B.M.; Purza, A.L.; Radu, A. Evaluation of the phytochemistry-therapeutic activity relationship for grape seeds Oil. Life 2023, 13, 178. [Google Scholar] [CrossRef]
- Barbagallo, M.G.; Patti, D.; Pisciotta, A. Phenotypic traits of berries and seeds of Sicilian grape cultivars (Vitis vinifera L.). Sci. Hortic. 2020, 261, 109006. [Google Scholar] [CrossRef]
- Sharafan, M.; Malinowska, M.A.; Kubicz, M.; Kubica, P.; Gémin, M.; Abdallah, C.; Ferrier, M.; Hano, C.; Giglioli-Guivarc’h, N.; Sikora, E.; et al. Shoot Cultures of Vitis vinifera (Vine Grape) Different cultivars as a promising innovative cosmetic raw material—Phytochemical profiling, antioxidant potential, and whitening activity. Molecules 2023, 28, 6868. [Google Scholar] [CrossRef]
- Rivera, D.; Miralles, B.; Obón, C.; Carreño, E.; Palazón, J.A. Multivariate analysis of Vitis subgenus Vitis seed morphology. Vitis 2007, 46, 158–167. [Google Scholar]
- Planchon, J.E. Ampelideae, Monographie des Ampélidées Vraies. In Monographiae Phanerogamarum; De Candolle, A.P., Ed.; Treuttel et Würtz: Paris, France, 1887; Volume 5, pp. 305–368. [Google Scholar]
- Viala, P.; Péchoutre, P. Morphologie externe de la graine In Ampélographie; Viala, P., Vermorel, V., Eds.; Masson et Cie.: Paris, France, 1910; pp. 156–166. [Google Scholar]
- Ocete, R.; Cantos, M.; López, M.A.; Gallardo, A.; Pérez, M.A.; Troncoso, A.; Lara, M.; Failla, O.; Ferragut, F.J.; Liñán, J. Caracterización y Conservación del Recurso Fitogenético vid Silvestre en Andalucía; Consejería de Medio Ambiente, Junta de Andalucía: Seville, Spain, 2007. [Google Scholar]
- Hajnalová, M.; Látková, M.; Kajanová, M.; Eliáš jun, P.; Ďurišová, Ľ. Wild or cultivated? A study of Vitis sylvestris in natura in Slovakia and implications for archaeology and archaeobotany (morphometric approach). Veg. Hist. Archaeobotany 2023, 32, 321–337. [Google Scholar] [CrossRef]
- Orrú, M.; Grillo, O.; Lovicu, G.; Venora, G.; Bacchetta, G. Morphological characterisation of Vitis vinifera L. seeds by image analysis and comparison with archaeological remains. Veget. Hist. Archaeobot. 2012, 22, 231–242. [Google Scholar] [CrossRef]
- Orrú, M.; Grillo, O.; Venora, G.; Bacchetta, G. Seed morpho-colorimetric analysis by computer vision: A helpful tool to identify grapevine (Vitis vinifera L.) cultivars. Grape Wine Res. 2015, 21, 508–519. [Google Scholar] [CrossRef]
- Sonka, M.; Hlavac, V.; Boyle, R. Image Processing Analysis and Machine Vision, 3rd ed.; Thomson Learning: Toronto, ON, Canada, 2008; 829p. [Google Scholar]
- Ucchesu, M.; Orrú, M.; Grillo, O.; Venora, G.; Usai, A.; Serreli, P.F.; Bacchetta, G. Earliest evidence of a primitive cultivar of Vitis vinifera L. during the Bronze Age in Sardinia (Italy). Veget. Hist. Archaeobot. 2015, 24, 587–600. [Google Scholar] [CrossRef]
- Ucchesu, M.; Orrú, M.; Grillo, O.; Venora, G.; Paglietti, G.; Ardu, A.; Bacchetta, G. Predictive Method for Correct Identification of Archaeological Charred Grape Seeds: Support for Advances in Knowledge of Grape Domestication Process. PLoS ONE 2016, 11, e0149814. [Google Scholar] [CrossRef]
- Milanesi, C.; Costantini, L.; Firmati, M.; Antonucci, F.; Faleri, C.; Buracchi, A.; Cresti, M. Geometric morphometry and archaeobotany: Characterisation of grape seeds (Vitis vinifera L.) by analysis of form. Open Access Libr. J. 2014, 1, e634. [Google Scholar] [CrossRef]
- Martín-Gómez, J.J.; Gutiérrez del Pozo, D.; Ucchesu, M.; Bacchetta, G.; Cabello Sáenz de Santamaría, F.; Tocino, Á.; Cervantes, E. Seed morphology in the Vitaceae based on geometric models. Agronomy 2020, 10, 739. [Google Scholar] [CrossRef]
- Cervantes, E.; Martín-Gómez, J.J.; Espinosa-Roldán, F.E.; Muñoz-Organero, G.; Tocino, Á.; Cabello-Sáenz de Santamaría, F. Seed morphology in key Spanish grapevine cultivars. Agronomy 2021, 11, 734. [Google Scholar] [CrossRef]
- Martín-Gómez, J.J.; Gutiérrez del Pozo, D.; Rodríguez-Lorenzo, J.L.; Tocino, Á.; Cervantes, E. Geometric analysis of seed shape diversity in the Cucurbitaceae. Seeds 2023, 3, 40–55. [Google Scholar] [CrossRef]
- Cervantes, E.; Tocino, A. Geometric analysis of Arabidopsis root apex reveals a new aspect of the ethylene signal transduction pathway in development. J. Plant Physiol. 2005, 162, 1038–1045. [Google Scholar] [CrossRef]
- Noriega, A.; Tocino, A.; Cervantes, E. Hydrogen peroxide treatment results in reduced curvature values in the Arabidopsis root apex. J. Plant Physiol. 2009, 166, 554–558. [Google Scholar] [CrossRef]
- Martín-Gómez; Rewicz; Goriewa-Duba; Wiwart; Tocino; Cervantes Morphological Description and classification of wheat kernels based on geometric models. Agronomy 2019, 9, 399. [CrossRef]
- Cervantes, E.; Martín-Gómez, J.J.; Espinosa-Roldán, F.E.; Muñoz-Organero, G.; Tocino, Á.; Cabello Sáenz de Santamaría, F. Seed apex curvature in key Spanish grapevine cultivars. Vitic. Data J. 2021, 3, e66478. [Google Scholar] [CrossRef]
- Wen, J.; Lu, L.-M.; Nie, Z.-L.; Liu, X.-Q.; Zhang, N.; Ickert-Bond, S.; Gerrath, J.; Manchester, S.R.; Boggan, J.; Chen, Z.-D. A new phylogenetic tribal classification of the grape family (Vitaceae). J. Syst. Evol. 2018, 56, 262–272. [Google Scholar] [CrossRef]
- Ferreira, T.; Rasband, W. Imagej User Guide-Ij1.46r. 2012; 186 p. Available online: http://imagej.nih.gov/ij/docs/guide (accessed on 19 May 2020).
- Cox, E.P. A method of assigning numerical and percentage values to the degree of roundness of sand grains. J. Paleontol. 1927, 1, 179–183. [Google Scholar]
- Riley, N.A. Projection sphericity. J. Sediment. Pet. 1941, 11, 94–97. [Google Scholar]
- Schwartz, H. Two-dimensional feature-shape indexes. Mikroskopie 1980, 37, 64–67. [Google Scholar]
- Sokal, R.R.; Braumann, C.A. Significance tests for coefficients of variation and variability profiles. Syst. Zool. 1980, 29, 50. [Google Scholar] [CrossRef]
- Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Tocino, Á.; Janoušek, B.; Juan, A.; Cervantes, E. The outline of seed silhouettes: A morphological approach to Silene (Caryophyllaceae). Plants 2022, 11, 3383. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, E.; Martín-Gómez, J.J.; Gutiérrez del Pozo, D.; Tocino, Á. Seed Geometry in the Vitaceae. Plants 2021, 10, 1695. [Google Scholar] [CrossRef]
- Burnham, R.J. Climbers. Censusing Lianas in Mesic Biomes of Eastern Regions. Vitis aestivalis Michx. Available online: https://climbers.lsa.umich.edu/vitis-aestivalis/ (accessed on 18 February 2024).
- Ardenghi, N.M.G.; Galasso, G.; Banfi, E.; Cauzzi, P. Vitis × novae-angliae (Vitaceae): Systematics, distribution and history of an “illegal” alien grape in Europe. Willdenowia 2015, 45, 197–207. [Google Scholar] [CrossRef]
- Wan, Y.; Schwaninger, H.R.; Baldo, A.M.; Labate, J.A.; Zhong, G.Y.; Simon, C.J. A phylogenetic analysis of the grape genus (Vitis L.) reveals broad reticulation and concurrent diversification during neogene and quaternary climate change. BMC Evol. Biol. 2013, 13, 141. [Google Scholar] [CrossRef] [PubMed]
- da Costa, A.F.; Teodoro, P.E.; Bhering, L.L.; Tardin, F.D.; Daher, R.F.; Campos, W.F.; Viana, A.P.; Pereira, M.G. Molecular analysis of genetic diversity among vine accessions using DNA markers. Genet. Mol. Res. 2017, 16, gmr16029586. [Google Scholar] [CrossRef]
- Péros, J.P.; Launay, A.; Peyrière, A.; Berger, G.; Roux, C.; Lacombe, T.; Boursiquot, J.M. Species relationships within the genus Vitis based on molecular and morphological data. PLoS ONE 2023, 18, e0283324. [Google Scholar] [CrossRef]
- Kerekes, A.; Tóth-Lencsés, K.A.; Kiss, E.; Szőke, A. Phylogeny of Vitis species based on a VvMybA1 marker analysis. Acta Hortic. 2019, 1248, 135–140. [Google Scholar] [CrossRef]
- Hall, B.K. Homology and homoplasy. In Handbook of the Philosophy of Science, Philosophy of Biology; Matthen, M., Stephens, C., Eds.; Elsevier: Amstercam, The Netherlands, 2007; pp. 429–453. [Google Scholar]
- Olmo, H. The origin and domestication of the Vinifera grape. In The Origins and Ancient History of Wine; McGovern, P.E., Fleming, S.J., Katz, S.H., Eds.; Gordon and Breach: Amsterdam, The Netherlands, 1995; pp. 31–43. [Google Scholar]
- Cunha, J.; Baleiras-Couto, M.; Cunha, J.P.; Banza, J.; Soveral, A.; Carneiro, L.C.; Eiras-Dias, J.E. Characterization of Portuguese populations of Vitis vinifera L. ssp. sylvestris (Gmelin) Hegi. Genet. Resour. Crop. Evol. 2007, 54, 981–988. [Google Scholar] [CrossRef]
- Arroyo-García, R.; Lefort, F.; De Andrés, M.; Ibáñez, J.; Borrego, J.; Cabello, F.; Martínez-Zapater, J.M. Haplotypic polymorphisms for chloroplast microsatellites analysis in Vitis. Genome 2002, 45, 1142–1149. [Google Scholar] [CrossRef] [PubMed]
- Webb, D.A. Vitis. In Flora Europaea 2; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.M., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 246. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Tribe |
|---|---|
| Ampelopsis aconitifolia Bunge | Ampelopsideae |
| Cissus verticillata (L.) Nicolson and C.E. Jarvis | Cisseae |
| Parthenocissus quinquefolia (L.) Planch. | Parthenocisseae |
| Vitis aestivalis Michx. | Viteae |
| Vitis amurensis Rupr. | Viteae |
| Vitis berlandieri Planch. | Viteae |
| Vitis californica Parry | Viteae |
| Vitis candicans Engelm. ex A.Gray | Viteae |
| Vitis x doaniana Munson ex Viala | Viteae |
| Vitis labrusca L. | Viteae |
| Vitis riparia A. Gray | Viteae |
| Vitis rupestris Scheele | Viteae |
| Species | A | P | L | W | C | AR | R | S |
|---|---|---|---|---|---|---|---|---|
| Ampelopsis aconitifolia Bunge | 9.71 a (6.4) | 12.37 a (4.4) | 3.90 a (4.1) | 3.17 a (3.7) | 0.80 efg (4.0) | 1.23 b (4.5) | 0.81 e (4.5) | 0.975 e (0.8) |
| Cissus verticillata (L.) Nicolson and C.E. Jarvis | 11.65 b (5.5) | 14.44 b (3.6) | 4.55 b (3.1) | 3.26 b (3.1) | 0.70 a (5.5) | 1.40 d (2.8) | 0.72 c (2.7) | 0.957 b (0.6) |
| Parthenocisus quinquefolia (L.) Planch. | 14.84 d (6.3) | 15.24 c (3.2) | 4.64 b (4.9) | 4.07 f (5.0) | 0.80 fg (2.8) | 1.14 a (7.6) | 0.88 f (7.6) | 0.982 f (0.4) |
| Vitis aestivalis Michx. | 17.92 f (8.5) | 17.87 f (4.3) | 6.21 gh (6.0) | 3.68 c (6.4) | 0.70 a (4.1) | 1.69 f (9.4) | 0.60 a (9.8) | 0.961 bc (1.62) |
| Vitis amurensis Rupr. | 13.95 c (4.90) | 14.73 b (3.0) | 4.85 c (3.3) | 3.66 c (3.4) | 0.81 g (2.6) | 1.32 c (4.4) | 0.76 d (4.4) | 0.973 de (0.42) |
| Vitis berlandieri Planch. | 17.40 ef (5.2) | 15.95 e (3.0) | 5.70 e (4.2) | 3.89 de (4.7) | 0.76 c (3.0) | 1.47 e (7.2) | 0.68 b (7.2) | 0.971 d (0.5) |
| Vitis californica Parry | 14.67 cd (10.0) | 15.19 c (5.5) | 4.99 c (7.7) | 3.75 cd (7.7) | 0.80 defg (3.7) | 1.34 c (12.1) | 0.76 de (11.7) | 0.981 f (0.5) |
| Vitis candicans Engelm. ex A.Gray | 16.85 e (7.9) | 16.62 de (4.4) | 5.26 d (4.2) | 4.08 ef (7.1) | 0.77 cd (7.1) | 1.30 bc (8.9) | 0.78 def (8.7) | 0.981 f (0.46) |
| Vitis doaniana Munson ex Viala | 22.21 h (5.3) | 18.46 g (2.5) | 5.96 f (3.4) | 4.54 g (4.3) | 0.78 cdef (2.7) | 1.32 c (5.8) | 0.76 d (5.6) | 0.975 e (0.4) |
| Vitis labrusca L. | 23.81 i (6.6) | 20.25 h (5.1) | 6.39 h (5.0) | 4.75 h (3.9) | 0.73 b (5.9) | 1.35 c (5.9) | 0.75 d (5.9) | 0.949 a (1.4) |
| Vitis riparia A. Gray | 19.35 g (7.4) | 18.09 f (3.8) | 6.07 fg (4.5) | 4.06 f (5.0) | 0.74 b (3.4) | 1.50 e (6.1) | 0.67 b (6.1) | 0.963 c (0.8) |
| Vitis rupestris Scheele | 15.62 d (13.8) | 15.90 cd (7.7) | 5.05 c (9.0) | 3.93 def (6.0) | 0.77 cde (3.8) | 1.29 bc (6.4) | 0.78 de (6.2) | 0.965 c (0.9) |
| Species | Model | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MAac | MCve | MPqu | MVae | MVam | MVbe | MVca | MVcan | MVdo | MVla | MVri | MVru | |
| A. aconitifolia | 90.0 f (1.6) | 83.9 abc (3.2) | 88.7 e (1.9) | 73.2 b (4.3) | 88.5 cdef (2.8) | 86.5 bcd (2.7) | 87.3 cde (1.5) | 90.9 e (1.7) | 89.1 cd (2.2) | 84.6 bc (3.3) | 81.7 b (3.8) | 90.4 e (1.7) |
| C. verticillata | 86.1 cd (1.8) | 89.3 g (0.9) | 82.1 c (2.0) | 80.1 d (2.0) | 89.3 ef (1.3) | 89.8 fg (1.2) | 87.9 de (0.9) | 89.1 c (1.1) | 89.6 d (1.2) | 88.6 f (1.0) | 87.6 f (1.0) | 90.0 e (1.6) |
| P. quinquefolia | 87.5 de (2.4) | 80.5 a (5.6) | 90.2 f (1.9) | 66.6 a (5.2) | 84.2 ab (5.5) | 79.2 a (6.3) | 83.6 b (4.7) | 88.6 cde (4.6) | 85.7 b (4.9) | 81.6 a (4.4) | 77.7 a (5.6) | 85.2 b (3.9) |
| V. aestivalis | 76.9 a (6.2) | 83.5 abc (3.8) | 71.7 a (6.7) | 87.5 f (4.3) | 81.8 a (5.6) | 84.0 b (4.8) | 80.6 a (5.3) | 79.9 a (6.0) | 81.6 a (5.3) | 83.1 ab (4.7) | 84.9 cde (3.8) | 81.0 a (5.2) |
| V. amurensis | 87.8 e (2.1) | 87.1 f (1.2) | 85.4 d (2.8) | 77.1 c (2.9) | 91.1 g (1.1) | 89.3 efg (2.1) | 88.3 e (1.0) | 91.2 e (1.7) | 91.2 e (1.0) | 87.9 ef (1.8) | 86.1 de (1.8) | 90.3 e (1.1) |
| V. berlandieri | 84.8 c (3.8) | 86.3 def (1.6) | 80.4 bc (5.0) | 81.9 d (4.4) | 89.0 ef (3.1) | 89.9 g (2.5) | 86.9 cde (3.0) | 87.9 bc (3.7) | 88.4 cd (3.6) | 86.3 cde (2.6) | 87.0 ef (1.9) | 86.9 bcd (3.6) |
| V. californica | 86.4 cde (4.0) | 82.6 ab (4.0) | 85.6 de (6.5) | 75.6 bc (8.3) | 86.7 bc (2.5) | 85.7 bcd (4.1) | 86.5 bcde (3.3) | 88.8 cde (3.9) | 86.9 b (2.4) | 84.4 b (2.7) | 81.8 bc (6.1) | 86.4 bc (2.9) |
| V. candicans | 87.9 e (2.3) | 84.4 bcd (2.6) | 87.0 de (4.1) | 75.6 bc (5.2) | 89.0 def (2.6) | 86.1 bcde (4.6) | 87.4 de (2.0) | 91.4 e (2.0) | 89.0 cd (2.0) | 85.6 bcd (2.7) | 83.2 bcd (4.1) | 88.8 d (1.7) |
| V. doaniana | 87.9 e (2.2) | 86.4 ef (2.7) | 85.3 d (3.6) | 77.2 c (4.3) | 90.4 fg (1.8) | 87.8 cdef (2.9) | 87.7 de (1.7) | 90.9 e (1.8) | 90.7 e (2.0) | 87.5 def (2.3) | 84.8 cd (3.0) | 90.0 e (2.0) |
| V. labrusca | 85.0 c (3.2) | 85.4 cde (2.2) | 81.9 c (4.4) | 77.9 c (3.1) | 87.4 cde (3.3) | 85.3 bc (3.1) | 85.3 bc (2.9) | 88.9 cd (2.6) | 88.7 cd (2.6) | 88.6 f (2.1) | 83.6 bc (2.7) | 87.9 cd (3.1) |
| V. riparia | 82.5 b (3.2) | 87.3 f (1.6) | 78.0 b (3.5) | 84.5 e (3.4) | 87.5 cd (2.7) | 89.3 fg (1.8) | 86.2 bcd (2.0) | 86.4 b (3.0) | 87.3 bc (2.5) | 86.9 de (2.2) | 88.1 g (1.6) | 85.9 b (2.9) |
| V. rupestris | 88.3 e (2.4) | 86.8 ef (2.6) | 87.3 de (2.0) | 77.2 c (3.7) | 89.3 ef (1.3) | 88.3 def (2.4) | 87.7 de (1.4) | 90.8 de (1.3) | 90.3 de (1.1) | 87.8 ef (2.2) | 85.0 cd (3.3) | 90.9 e (1.4) |
| Species | N | Max C. | Mean C. | Ratio |
|---|---|---|---|---|
| Ampelopsis aconitifolia Bunge | 20 | 5.07 f (22.87) | 0.59 g (4.58) | 8.60 def (25.05) |
| Cissus verticillata Nicolson and C.E. Jarvis | 20 | 5.03 f (26.65) | 0.65 g (30.73) | 10.0 ef (25.26) |
| Parthenocisus quinquefolia (L.) Planch. | 21 | 3.08 d (15.87) | 0.47 de (14.66) | 6.56 c (12.73) |
| Vitis aestivalis Michx. | 18 | 2.53 b (17.92) | 0.64 f (57.89) | 4.98 b (57.81) |
| Vitis amurensis Rupr. | 20 | 4.15 e (14.31) | 0.52 ef (5.38) | 7.94 de (12.91) |
| Vitis berlandieri Planch. | 10 | 4.79 f (8.72) | 0.48 d (3.25) | 10.08 f (10.63) |
| Vitis californica Parry | 20 | 2.16 a (66.69) | 0.50 def (17.75) | 4.57 a (91.66) |
| Vitis candicans Engelm. ex A.Gray | 20 | 2.54 bc (19.13) | 0.37 ab (11.03) | 6.91 c (19.31) |
| Vitis doaniana Munson ex Viala | 19 | 4.13 e (14.43) | 0.48 d (11.00) | 8.74 ef (17.32) |
| Vitis labrusca L. | 19 | 3.39 d (41.78) | 0.35 a (15.70) | 9.91 ef (45.46) |
| Vitis riparia A. Gray | 22 | 3.12 cd (38.34) | 0.44 c (12.85) | 7.03 cd (34.27) |
| Vitis rupestris Scheele | 22 | 3.30 d (21.17) | 0.40 b (15.15) | 8.39 def (22.97) |
| Species | Symmetry |
|---|---|
| Ampelopsis aconitifolia Bunge | 92.28 cde (2.1) |
| Cissus verticillata (L.) Nicolson and C.E. Jarvis | 91.23 bcd (1.6) |
| Parthenocissus quinquefolia (L.) Planch. | 92.40 e (1.8) |
| Vitis aestivalis Michx. | 90.85 b (1.6) |
| Vitis amurensis Rupr. | 91.25 bc (0.8) |
| Vitis berlandieri Planch. | 91.70 bcde (1.7) |
| Vitis californica Parry | 92.13 bcde (1.9) |
| Vitis candicans Engelm. ex A.Gray | 91.33 bcd (1.3) |
| Vitis doaniana Munson ex Viala | 90.99 bcde (3.0) |
| Vitis labrusca L. | 88.57 a (2.6) |
| Vitis riparia A. Gray | 91.53 bcde (1.6) |
| Vitis rupestris Scheele | 92.51 de (1.4) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Gutiérrez del Pozo, D.; Cabello Sáez de Santamaría, F.; Muñoz-Organero, G.; Tocino, Á.; Cervantes, E. Seed Morphological Analysis in Species of Vitis and Relatives. Horticulturae 2024, 10, 285. https://doi.org/10.3390/horticulturae10030285
Martín-Gómez JJ, Rodríguez-Lorenzo JL, Gutiérrez del Pozo D, Cabello Sáez de Santamaría F, Muñoz-Organero G, Tocino Á, Cervantes E. Seed Morphological Analysis in Species of Vitis and Relatives. Horticulturae. 2024; 10(3):285. https://doi.org/10.3390/horticulturae10030285
Chicago/Turabian StyleMartín-Gómez, José Javier, José Luis Rodríguez-Lorenzo, Diego Gutiérrez del Pozo, Félix Cabello Sáez de Santamaría, Gregorio Muñoz-Organero, Ángel Tocino, and Emilio Cervantes. 2024. "Seed Morphological Analysis in Species of Vitis and Relatives" Horticulturae 10, no. 3: 285. https://doi.org/10.3390/horticulturae10030285
APA StyleMartín-Gómez, J. J., Rodríguez-Lorenzo, J. L., Gutiérrez del Pozo, D., Cabello Sáez de Santamaría, F., Muñoz-Organero, G., Tocino, Á., & Cervantes, E. (2024). Seed Morphological Analysis in Species of Vitis and Relatives. Horticulturae, 10(3), 285. https://doi.org/10.3390/horticulturae10030285