
Dynamics of Sugars, Organic Acids, Hormones, and Antioxidants in Grape Varieties ‘Italia’ and ‘Bronx Seedless’ during Berry Development and Ripening


 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Identification of Sugar in Grape Varieties with HPLC
2.3. Identification of Organic Acids in Grape Varieties with HPLC
2.4. Identification of Antioxidants in Grape Varieties with HPLC
2.5. Identification of Hormones in Grape Varieties with HPLC
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- OIV. OIV Statistical Report on World Vitiviniculture. 2019. Available online: http://www.oiv.int/en/oiv-life/oiv-2019-report-on-the-world-vitivinicultural-situation (accessed on 10 October 2023).
- Liu, H.F.; Wu, B.H.; Fan, P.G.; Li, S.H.; Li, L.S. Sugar and acid concentrations in 98 grape cultivars analyzed by principal component analysis. J. Sci. Food Agric. 2006, 86, 1526–1536. [Google Scholar] [CrossRef]
- Julius, B.T.; Leach, K.A.; Tran, T.M.; Mertz, R.A.; Braun, D.M. Sugar transporters in plants: New insights and discoveries. Plant Cell Physiol. 2017, 58, 1442–1460. [Google Scholar] [CrossRef]
- Lalonde, S.; Tegeder, M.; Frommer, W.B.; Patrick, J.W.; Throne-Holst, M. Phloem loading and unloading of sugars and amino acids. Plant Cell Environ. 2003, 26, 37–56. [Google Scholar] [CrossRef]
- Kambiranda, D.; Vasanthaiah, H.; Basha, S.M. Relationship between acid invertase activity and sugar content in grape species. J. Food Biochem. 2011, 35, 1646–1652. [Google Scholar] [CrossRef]
- Chenu, A.; Scholes, G.D. Coherence in energy transfer and photosynthesis. Annu. Rev. Phys. Chem. 2015, 66, 69–96. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Cao, H.L.; Wang, L.; Zhou, Y.H.; Huang, Y.T.; Hao, X.Y.; Wang, Y.C.; Wang, B.; Yang, Y.J.; Wang, X.C. Effects of cold acclimation on sugar metabolism and sugar-related gene expression in tea plant during the winter season. Plant Mol. Biol. 2015, 88, 591–608. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L.; et al. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef] [PubMed]
- Davies, C.; Boss, P.K.; Geros, H.; Lecourieux, F.; Delrot, S. Source/sink relationships and molecular biology of sugar accumulation in grape berries. In The Biochemistry of the Grape Berry; Bentham Science Publishers: Sharjah, United Arab Emirates, 2012; pp. 44–66. [Google Scholar]
- Zhang, S.; Wang, H.; Wang, T.; Zhang, J.; Liu, W.; Fang, H.; Zhang, Z.; Peng, F.; Chen, X.; Wang, N. Abscisic acid and regulation of the sugar transporter gene MdSWEET9b promote apple sugar accumulation. Plant Physiol. 2023, 192, 2081–2101. [Google Scholar] [CrossRef]
- Lamikanra, O.; Inyang, I.D.; Leong, S. Distribution and effect of grape maturity on organic acid content of red muscadine grapes. J. Agric. Food Chem. 1995, 43, 3026–3028. [Google Scholar] [CrossRef]
- Kaya, O. Effect of manual leaf removal and its timing on yield, the presence of lateral shoots and cluster characteristics with the grape variety ‘Karaerik’. Mitt Klosterneubg Rebe Wein Obstbau Früchteverwertung 2019, 69, 83–92. [Google Scholar]
- Kaya, O.; Ates, F.; Kara, Z.; Turan, M.; Gutiérrez-Gamboa, G. Study of Primary and Secondary Metabolites of Stenospermocarpic, Parthenocarpic and Seeded Raisin Varieties. Horticulturae 2022, 8, 1030. [Google Scholar] [CrossRef]
- Coombe, B.G. Research on development and ripening of the grape berry. Am. J. Enol. Vitic. 1992, 43, 101–110. [Google Scholar] [CrossRef]
- Kliewer, W.M. Sugars and organic acids of Vitis vinifera. Plant Physiol. 1966, 41, 923–931. [Google Scholar] [CrossRef]
- Ali, K.; Maltese, F.; Fortes, A.M.; Pais, M.S.; Choi, Y.H.; Verpoorte, R. Monitoring biochemical changes during grape berry development in Portuguese cultivars by NMR spectroscopy. Food Chem. 2011, 124, 1760–1769. [Google Scholar] [CrossRef]
- Gutiérrez-Escobar, R.; Aliaño-González, M.J.; Cantos-Villar, E. Wine polyphenol content and its influence on wine quality and properties: A Review. Molecules 2021, 26, 718. [Google Scholar] [CrossRef]
- Ali, K.; Maltese, F.; Choi, Y.H.; Verpoorte, R. Metabolic constituents of grapevine and grape-derived products. Phytochem. Rev. 2010, 9, 357–378. [Google Scholar] [CrossRef] [PubMed]
- CooMbe, B.G.; McCarthy, M.G. Dynamics of grape berry growth and physiology of ripening. Aust. J. Grape Wine Res. 2000, 6, 131–135. [Google Scholar] [CrossRef]
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.C.P.; Tavares, R.M.; Sousa, M.J.; Agasse, A.; Delrot, S.; Geros, H. Biochemical changes throughout grape berry development and fruit and wine quality. Food 2007, 1, 1–22. [Google Scholar]
- Lund, C.M.; Thompson, M.K.; Benkwitz, F.; Wohler, M.W.; Triggs, C.M.; Gardner, R.; Heymann, H.; Nicolau, L. New Zealand sauvignon blanc distinct flavor characteristics: Sensory, chemical, and consumer aspects. Am. J. Enol. Vitic. 2009, 60, 1–12. [Google Scholar] [CrossRef]
- McAtee, P.; Karim, S.; Schaffer, R.; David, K. A dynamic interplay between phytohormones is required for fruit development, maturation, and ripening. Front. Plant Sci. 2013, 4, 79. [Google Scholar] [CrossRef]
- Symons, G.M.; Davies, C.; Shavrukov, Y.; Dry, I.B.; Reid, J.B.; Thomas, M.R. Grapes on steroids. Brassinosteroids are involved in grape berry ripening. Plant Physiol. 2006, 140, 150–158. [Google Scholar] [CrossRef]
- Fortes, A.M.; Teixeira, R.T.; Agudelo-Romero, P. Complex interplay of hormonal signals during grape berry ripening. Molecules 2015, 20, 9326–9343. [Google Scholar] [CrossRef]
- Lorenz, D.H.; Eichhorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Growth Stages of the Grapevine: Phenological Growth Stages of the Grapevine (Vitis vinifera L. ssp. Vinifera)—Codes and Descriptions According to the Extended BBCH Scale. Aust. J. Grape Wine Res. 1995, 1, 100–103. [Google Scholar] [CrossRef]
- Ma, C.M.; Sun, Z.; Chen, C.B.; Zhang, L.L.; Zhu, S.H. Simultaneous separation and determination of fructose, sorbitol, glucose and sucrose in fruits by HPLC–ELSD. Food Chem. 2014, 145, 784–788. [Google Scholar] [CrossRef]
- Keskin, N.; Kunter, B.; Çelik, H.; Kaya, O.; Keskin, S. ANOM approach for the statistical evaluation of organic acid contents of clones of the grape variety ‘Kalecik Karası’. Mitteilungen Klosterneubg. 2021, 71, 126–138. [Google Scholar]
- Modesti, M.; Petriccione, M.; Forniti, R.; Zampella, L.; Scortichini, M.; Mencarelli, F. Methyl jasmonate and ozone affect the antioxidant system and the quality of wine grape during postharvest partial dehydration. Food Res. Int. 2018, 112, 369–377. [Google Scholar] [CrossRef]
- Keskin, N.; Kaya, O.; Ates, F.; Turan, M.; Gutiérrez-Gamboa, G. Drying grapes after the application of different dipping solutions: Effects on hormones, minerals, vitamins, and antioxidant enzymes in Gök Üzüm (Vitis vinifera L.) raisins. Plants 2022, 11, 529. [Google Scholar] [CrossRef]
- Minucci, A.; Giardina, B.; Zuppi, C.; Capoluongo, E. Glucose-6-Phosphate Dehydrogenase Laboratory Assay: How, When, and Why? IUBMB Life 2009, 61, 27–34. [Google Scholar] [CrossRef]
- Angelini, R.; Manes, F.; Federico, R. Spatial and Functional Correlation between Diamine-Oxidase and Peroxidase Activities and Their Dependence upon de-Etiolation and Wounding in Chick-Pea Stems. Planta 1990, 182, 89–96. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 8 August 2023).
- Wickham, H. Data Analysis; Springer International Publishing: Cham, Switzerland, 2016; pp. 189–201. [Google Scholar]
- Durán-Soria, S.; Pott, D.M.; Osorio, S.; Vallarino, J.G. Sugar signaling during fruit ripening. Front. Plant Sci. 2020, 11, 564917. [Google Scholar] [CrossRef]
- Swanson, C.A.; El-Shishiny, E.D.H.; Candela, M.I.; Fisher, E.G.; Hewitt, E.J. Translocation of Sugars in the Concord Grape. Plant Physiol. 1958, 33, 33–37. [Google Scholar] [CrossRef]
- Guo, Y.S.; Niu, Z.Z.; Su, K. Composition and content analysis of sugars and organic acids for 45 grape cultivars from the northeast region of China. Pak. J. Bot. 2017, 49, 155–160. [Google Scholar]
- Zhang, X.; Liu, Q.; Niu, S.; Liu, C.; Fan, X.; Zhang, Y.; Sun, L.; Jiang, J. Varietal differences among the fruit quality characteristic of 15 spine grapes (Vitis davidii Foëx). HortScience 2022, 57, 1282–1288. [Google Scholar] [CrossRef]
- Basson, C.E.; Groenewald, J.H.; Kossmann, J.; Cronje, C.; Bauer, R. Sugar and acid-related quality attributes and enzyme activities in strawberry fruits: Invertase is the main sucrose hydrolyzing enzyme. Sci. Hortic. 2010, 121, 1156–1162. [Google Scholar] [CrossRef]
- Li, M.; Feng, F.; Cheng, L. Expression patterns of genes involved in sugar metabolism and accumulation during apple fruit development. PLoS ONE 2012, 7, e33055. [Google Scholar] [CrossRef]
- Wen, Y.-Q.; Jing, C.; Zhang, Y.; Duan, C.-Q.; Pan, Q.-H. Comparison of organic acid levels and L-IdnDH expression in Chinese-type and European-type grapes. Euphytica 2014, 196, 63–76. [Google Scholar] [CrossRef]
- Liang, Z.; Sang, M.; Ma, A.; Zhao, S.; Zhong, G.-Y.; Li, S. Inheritance of sugar and acid contents in the ripe berries of a tetraploid × diploid grape cross population. Euphytica 2011, 182, 251–259. [Google Scholar] [CrossRef]
- Bolt, S.; Cook, D.R.; Ford, C.M. l-Tartaric acid synthesis from vitamin C in higher plants. Proc. Natl. Acad. Sci. USA 2006, 103, 5608–5613. [Google Scholar]
- Ruffner, H.P.; Hawker, J.S.; Hale, C.R. Temperature and enzymic control of malate metabolism in berries of Vitis vinifera. Phytochemistry 1976, 15, 1877–1880. [Google Scholar] [CrossRef]
- Topalovic, A.; Mikulic-Petkovsek, M. Changes in sugars, organic acids, and phenolics of grape berries of cultivar Cardinal during ripening. J. Food Agric. Environ. 2010, 8, 223–227. [Google Scholar]
- Aires, A.; Carvalho, R.; Matos, M.; Carnide, V.; Silva, A.P.; Gonçalves, B. Variation of chemical constituents, antioxidant activity, and endogenous plant hormones throughout different ripening stages of highbush blueberry (Vaccinium corymbosum L.) cultivars produced in the center of Portugal. Sci. Hortic. 2017, 41, e12414. [Google Scholar]
- Cantín, C.M.; Fidelibus, M.W.; Crisosto, C.H. Application of abscisic acid (ABA) at veraison advanced red color development and maintained postharvest quality of ‘Crimson Seedless’ grapes. Postharvest Biol. Technol. 2007, 46, 237–241. [Google Scholar] [CrossRef]
- Gagné, S.; Cluzet, S.; Mérillon, J.-M.; Gény, L. ABA initiates anthocyanin production in grape cell cultures. J. Plant Growth Regul. 2011, 30, 1–10. [Google Scholar] [CrossRef]
- Wheeler, S.; Loveys, B.; Ford, C.; Davies, C. The relationship between the expression of abscisic acid biosynthesis genes, accumulation of abscisic acid and the promotion of Vitis vinifera L. berry ripening by abscisic acid. Aust. J. Grape Wine Res. 2009, 15, 195–204. [Google Scholar] [CrossRef]
- Berli, F.J.; Fanzone, M.; Piccoli, P.; Bottini, R. Solar UV-B and ABA are involved in phenol metabolism of Vitis vinifera L., increasing biosynthesis of berry skin polyphenols. Sci. Hortic. 2011, 59, 4874–4884. [Google Scholar] [CrossRef]
- He, H.; Liang, G.; Lu, S.; Wang, P.; Liu, T.; Ma, Z.; Zuo, C.; Sun, X.; Chen, B.; Mao, J. Genome-Wide Identification and Expression Analysis of GA2ox, GA3ox, and GA20ox Are Related to Gibberellin Oxidase Genes in Grape (Vitis vinifera L.). Genes 2019, 10, 680. [Google Scholar] [CrossRef]
- Koyama, R.; Roberto, S.R.; de Souza, R.T.; Borges, W.F.S.; Anderson, M.; Waterhouse, A.L.; Cantu, D.; Fidelibus, M.W.; Blanco-Ulate, B. Exogenous abscisic acid promotes anthocyanin biosynthesis and increased expression of flavonoid synthesis genes in Vitis vinifera × Vitis labrusca table grapes in a subtropical region. Front. Plant Sci. 2018, 9, 323. [Google Scholar] [CrossRef]
- Gouthu, S.; Deluc, L.G. Timing of ripening initiation in grape berries and its relationship to seed content and pericarp auxin levels. BMC Plant Biol. 2015, 15, 46. [Google Scholar] [CrossRef]
- Bottcher, C.; Harvey, K.; Forde, C.G.; Boss, P.K.; Davies, C. Auxin treatment of pre-veraison grape (Vitis vinifera L.) berries both delays ripening and increases the synchronicity of sugar accumulation. Aust. J. Grape Wine Res. 2010, 17, 1–8. [Google Scholar] [CrossRef]
- Bagchi, D.; Bagchi, M.; Stohs, S.J.; Das, D.K.; Ray, S.D.; Kuszynski, C.A.; Joshi, S.S.; Pruess, H.G. Free radicals and grape seed proanthocyanidin extract: Importance in human health and disease prevention. Sci. Hortic. 2000, 148, 187–197. [Google Scholar] [CrossRef]
- Gris, E.F.; Mattivi, F.; Ferreira, E.A.; Vrhovsek, U.; Filho, D.W.; Pedrosa, R.C.; Bordignon-Luiz, M.T. Stilbenes and tyrosol as target compounds in the assessment of antioxidant and hypolipidemic activity of Vitis vinifera red wines from Southern Brazil. J. Agric. Food Chem. 2011, 59, 7954–7961. [Google Scholar] [CrossRef]
- Burin, V.M.; Ferreira-Lima, N.E.; Panceri, C.P.; Bordignon-Luiz, M.T. Bioactive compounds and antioxidant activity of Vitis vinifera and Vitis labrusca grapes: Evaluation of different extraction methods. Microchem. J. 2014, 114, 155–163. [Google Scholar] [CrossRef]
- Chang, S.K.; Alasalvar, C.; Shahidi, F. Superfruits: Phytochemicals, antioxidant efficacies, and health effects-A comprehensive review. Crit. Rev. Food Sci. Nutr. 2018, 59, 1580–1604. [Google Scholar] [CrossRef]
- HabibiDastjerd, Z.; Naseri, L.; Amiri, J.; Dolati Baneh, H. Effects of gibberellic-acid and salicylic-acid on phytochemical characteristics, antioxidant capacity and nutrient elements of Bidane-Sefid grape. J. Plant Prod. Res. 2019, 26, 103–120. [Google Scholar]
- Rüffer, M.; Steipe, B.; Zenk, M.H. Evidence against specific binding of salicylic acid to plant catalase. FEBS Lett. 1995, 377, 175–180. [Google Scholar] [CrossRef]
- Demir, H.; Güler, A.; Demir, C. Determination of some antioxidant enzyme activities in fresh and dry grapes (Vitis vinifera L.). Van Med. J. 2020, 27, 150–154. [Google Scholar] [CrossRef]
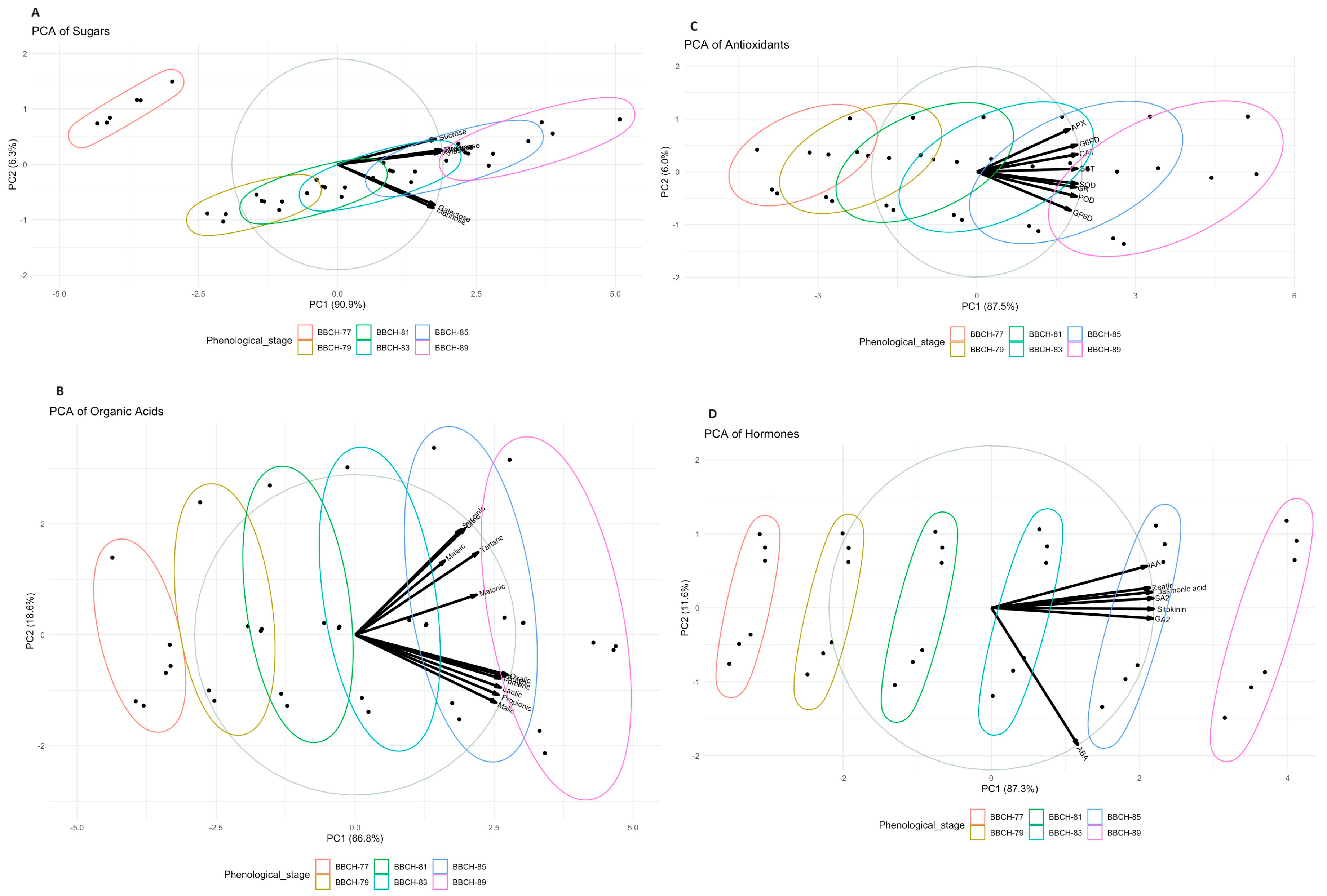
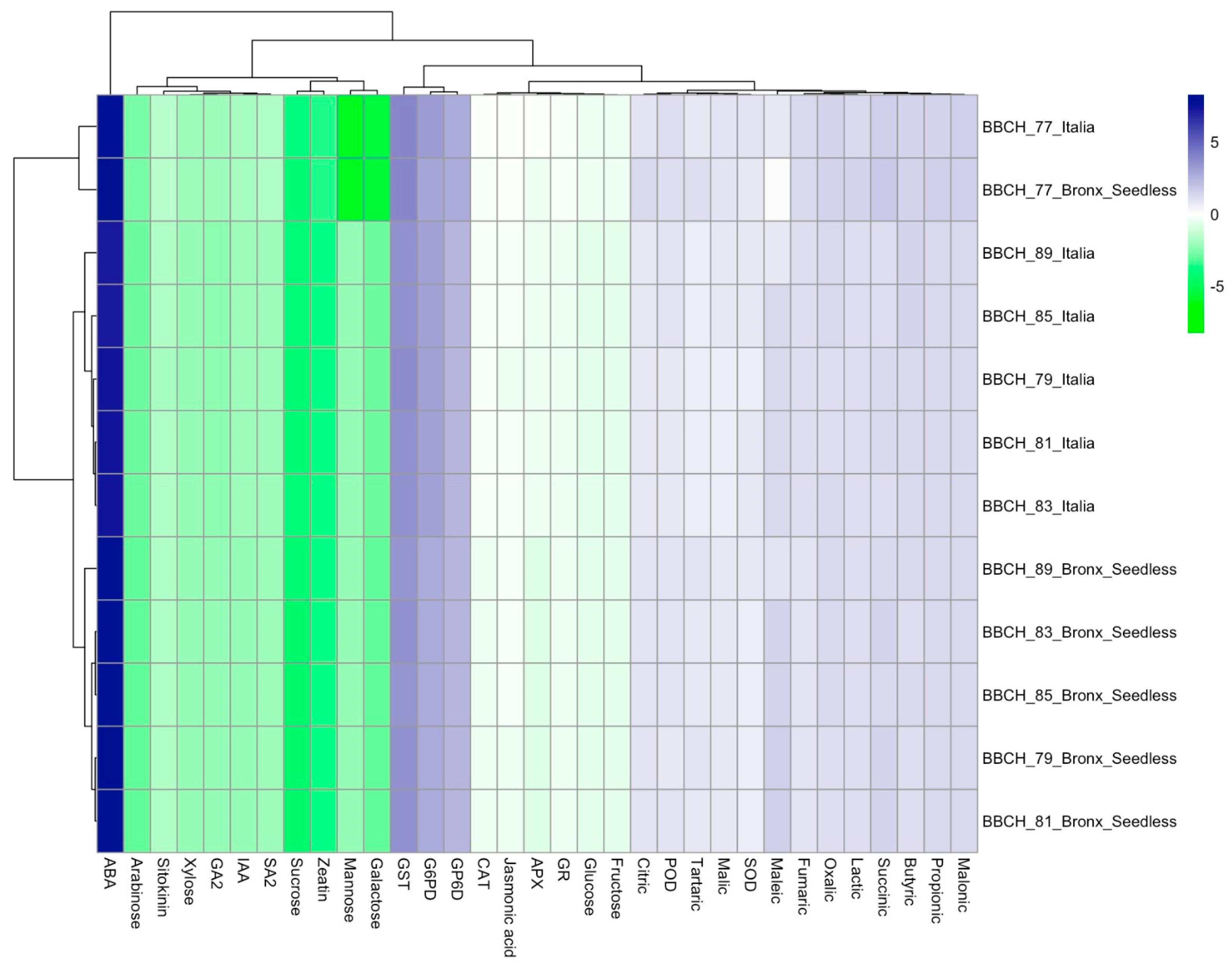

{kind=link}
{kind=link}
| Berry Development Stages | Berry Weight (g/Berry) | Berry Width (mm) | Berry Length (mm) | Total Soluble Solid (TSS) | Titratable Acidity (g/L as Tartaric Acid) (TA) | Maturity Index | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ‘Italia’ | ‘Bronx Seedless’ | ‘Italia’ | ‘Bronx Seedless’ | ‘Italia’ | ‘Bronx Seedless’ | ‘Italia’ | ‘Bronx Seedless’ | ‘Italia’ | ‘Bronx Seedless’ | ‘Italia’ | ‘Bronx Seedless’ | |
| BBCH-77 | 0.70 e | 0.38 f | 12.27 e | 8.97 e | 16.14 f | 11.54 f | 2.90 f | 2.80 f | 34.44 a | 29.52 a | 0.84 f | 0.98 f |
| BBCH-79 | 2.74 d | 1.30 e | 16.35 d | 12.26 d | 20.17 e | 14.77 e | 4.30 e | 4.50 e | 29.32 b | 24.28 b | 1.47 e | 1.85 e |
| BBCH-81 | 4.32 c | 1.84 d | 18.21 c | 13.86 c | 22.13 d | 16.41 d | 9.80 d | 10.10 d | 18.57 c | 15.38 c | 5.28 d | 6.57 d |
| BBCH-83 | 6.21 b | 2.54 c | 19.64 b | 14.79 b | 23.42 c | 17.24 c | 14.10 c | 14.40 c | 10.38 d | 8.59 d | 13.58 c | 16.76 c |
| BBCH-85 | 7.86 ab | 3.02 b | 20.95 ab | 15.82 ab | 25.03 b | 18.51 b | 15.30 b | 15.80 b | 9.21 e | 7.63 e | 16.61 b | 20.71 b |
| BBCH-89 | 8.14 a | 3.61 a | 21.62 a | 16.48 a | 26.45 a | 19.45 a | 16.80 a | 17.40 a | 6.51 f | 5.39 f | 25.81 a | 32.28 a |
| p value | 0.008 * | 0.006 * | 0.009 * | 0.002 * | 0.004 * | 0.008 * | 0.005 * | 0.009 * | 0.008 * | 0.007 * | 0.003 * | 0.003 * |
| Cultivar X (C) | Sucrose | Glucose | Fructose | Mannose | Galactose | Xylone | Arabinose |
|---|---|---|---|---|---|---|---|
| Italia | 0.753 ± 0.01 a | 8.64 ± 0.07 a | 8.67 ± 0.02 a | 2.33 ± 0.04 a | 1.50 ± 0.02 a | 2.64 ± 0.04 a | 1.69 ± 0.03 a |
| Bronx Seedless | 0.595 ± 0.02 b | 7.89 ± 0.08 b | 7.81 ± 0.03 b | 2.15 ± 0.04 b | 1.36 ± 0.01 b | 2.44 ± 0.02 b | 1.52 ± 0.01 b |
| Phenological Stage Y (PS) | |||||||
| BBCH-77 | 0.483 ± 0.02 e | 6.23 ± 0.11 f | 6.21 ± 0.03 f | 0.113 ± 0.07 e | 0.163 ± 0.03 f | 1.92 ± 0.08 e | 1.21 ± 0.03 e |
| BBCH-79 | 0.547 ± 0.03 d | 6.93 ± 0.13 e | 6.91 ± 0.01 e | 2.133 ± 0.03 de | 1.347 ± 0.02 e | 2.13 ± 0.06 de | 1.35 ± 0.01 d |
| BBCH-81 | 0.619 ± 0.01 cd | 7.71 ± 0.15 d | 7.69 ± 0.01 d | 2.372 ± 0.02 cd | 1.498 ± 0.01 d | 2.37 ± 0.08 cd | 1.50 ± 0.03 c |
| BBCH-83 | 0.701 ± 0.02 bc | 8.57 ± 0.13 c | 8.55 ± 0.02 c | 2.638 ± 0.02 bc | 1.666 ± 0.03 c | 2.64 ± 0.02 bc | 1.67 ± 0.01 b |
| BBCH-85 | 0.794 ± 0.03 ab | 9.53 ± 0.12 b | 9.50 ± 0.03 b | 2.933 ± 0.01 b | 1.853 ± 0.02 b | 2.93 ± 0.03 ab | 1.85 ± 0.03 a |
| BBCH-89 | 0.898 ± 0.02 a | 10.60 ± 0.11 a | 10.57 ± 0.04 a | 3.262 ± 0.04 a | 2.060 ± 0.03 a | 3.26 ± 0.08 a | 2.06 ± 0.03 a |
| Significance | |||||||
| C | 3.69 × 10−9 *** | 2.45 × 10−6 *** | <0.0 × 10−16 *** | 0.0094 ** | 9.25 × 10−6 *** | 0.0057 ** | 3.65 × 10−6 *** |
| PS | 4.02 × 10−12 *** | 4.04 × 10−16 *** | <2 × 10−16 *** | <2 × 10−16 *** | <2 × 10−16 *** | 8.51 × 10−11 *** | 9.79 × 10−15 *** |
| C × PS | 0.6169 | 0.9389 | 0.9334 | 0.9000 | 0.9389 | 0.9973 | 0.9449 |
| Cultivar X (C) | Oxalic | Propionic | Tartaric | Butyric | Malonic | Malic | Lactic | Citric | Maleic | Fumaric | Succinic |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Italia | 28.8 ± 0.5 a | 29.6 ± 0.8 | 18.6 ± 0.6 b | 32.0 ± 0.1 a | 32.0 ± 1.3 | 20.3 ± 0.3 | 26.6 ± 0.5 | 21.2 ± 1.2 | 27.4 ± 1.2 | 27.2 0.3 a | 29.0 ± 1.2 |
| Bronx Seedless | 27.1 ± 0.4 b | 28.9 ± 0.8 | 21.3 ± 0.3 a | 26.7 ± 0.3 b | 29.5 ± 1.6 | 29.7 ± 0.8 | 26.0 ± 0.4 | 23.5 ± 1.2 | 28.4 ± 1.1 | 23.0 ± 0.6 b | 31.1 ± 1.1 |
| Phenological stage Y (PS) | |||||||||||
| BBCH-77 | 20.0 ± 0.2 e | 22.1 ± 1.4 d | 15.1 ± 1.2 d | 21.0 ± 1.1 e | 23.2 ± 2.8 b | 14.4 ± 1.4 d | 19.8 ± 0.6 e | 16.9 ± 2.2 b | 11.0 ± 2.1 c | 19.0 ± 1.1 d | 24.6 ± 1.3 c |
| BBCH-79 | 22.7 ± 0.3 de | 24.5 ± 1.3 cd | 16.7 ± 1.3 cd | 23.8 ± 1.1 de | 25.8 ± 2.8 b | 16.3 ± 1.4 cd | 22.1 ± 0.3 de | 18.8 ± 2.1 ab | 27.3 ± 2.1 bc | 21.1 ± 1.4 cd | 26.5 ± 1.9 bc |
| BBCH-81 | 25.7 ± 0.3 bc | 27.3 ± 1.5 cd | 18.6 ± 1.2 bcd | 27.0 ± 1.1 cd | 28.7 ± 2.8 ab | 18.4 ± 1.4 bcd | 24.5 ± 0.9 cd | 20.9 ± 1.1 ab | 30.4 ± 2.1 ab | 23.4 ± 1.3 bc | 28.7 ± 1.1 abc |
| BBCH-83 | 29.1 ± 0.4 c | 30.3 ± 1.6 bc | 20.7 ± 1.3 abc | 30.5 ± 1.1 bc | 31.9 ± 2.8 ab | 20.8 ± 1.4 abc | 27.3 ± 0.8 bc | 23.2 ± 1.3 ab | 33.8 ± 2.1 ab | 26.1 ± 1.2 ab | 31.0 ± 1.4 abc |
| BBCH-85 | 32.9 ± 0.5 b | 33.7 ± 1.7 ab | 23.0 ± 1.3 ab | 34.5 ± 1.1 ab | 35.4 ± 2.8 ab | 23.6 ± 1.4 ab | 30.3 ± 0.7 ab | 25.8 ± 2.1 a | 37.6 ± 2.1 a | 29.0 ± 1.1 ab | 33.4 ± 1.9 ab |
| BBCH-89 | 37.3 ± 0.8 a | 37.5 ± 1.2 a | 25.6 ± 1.1 a | 39.1 ± 1.1 a | 39.4 ± 2.8 a | 26.7 ± 1.4 a | 33.7 ± 0.9 a | 28.7 ± 2.3 a | 27.3 ± 2.1 bc | 32.2 ± 1.2 a | 36.1 ± 1.8 a |
| Significance | |||||||||||
| C | 0.0184 * | 0.5771 | 0.0083 ** | 5.23 × 10−6 *** | 0.2828 | 0.5845 | 0.5226 | 0.2075 | 0.5712 | 0.0001 *** | 0.1979 |
| PS | 4.46 × 10−13 *** | 6.85 × 10−7 *** | 5.43 × 10−16 *** | 1.61 × 10−10 *** | 0.0043 ** | 1.71 × 10−5 *** | 2.69 × 10−9 *** | 0.0057 ** | 1.15 × 10−7 *** | 1.41 × 10−7 *** | 0.0034 ** |
| C × PS | 0.9977 | 1.000 | 0.9979 | 0.9079 | 0.9999 | 1.000 | 1.0000 | 0.9999 | 0.7599 | 0.9841 | 0.9999 |
| Cultivar X (C) | GR (nmol g−1) | GST (nmol g−1) | G6PD (nmol g−1) | 6GPD (nmol g−1) | CAT (EU g Berry−1) | POD (EU g Berry−1) | SOD (EU g Berry−1) | APX (EU g Berry−1) |
|---|---|---|---|---|---|---|---|---|
| Italia | 9.36 ± 0.11 a | 153 ± 2 a | 102.6 ± 1.31 a | 70.3 ± 1.7 a | 10.75 ± 0.10 a | 22.9 ± 0.4 | 20.9 ± 0.3 a | 10.0 ± 0.1 a |
| Bronx Seedless | 8.67 ± 0.18 b | 134 ± 1 b | 77.6 ± 1.34 b | 64.6 ± 1.8 b | 8.89 ± 0.13 b | 21.7 ± 0.3 | 18.5 ± 0.2 b | 7.3 ± 0.2 b |
| Phenological stage Y (PS) | ||||||||
| BBCH-77 | 6.80 ± 0.32 e | 117 ± 3.12 e | 67.3 ± 2.3 e | 51.2 ± 3.3 d | 7.20 ± 0.21 e | 16.0 ± 0.3 e | 13.8 ± 0.5 e | 6.55 ± 0.33 d |
| BBCH-79 | 7.56 ± 0.12 de | 127 ± 3.32 de | 75.1 ± 2.1 d | 56.8 ± 2.1 cd | 8.09 ± 0.33 d | 18.1 ± 0.1 d | 15.7 ± 04 d | 7.28 ± 0.42 cd |
| BBCH-81 | 8.41 ± 0.32 cd | 137 ± 3.41 cd | 83.8 ± 2.2 c | 63.0 ± 4.1 bc | 9.08 ± 0.21 c | 20.5 ± 0.3 c | 18.0 ± 0.3 c | 8.10 ± 0.12 bc |
| BBCH-83 | 9.35 ± 0.33 bc | 148 ± 3.22 bc | 93.5 ± 2.4 bc | 70.0 ± 2.1 ab | 10.20 ± 0.33 b | 23.2 ± 0.5 bc | 20.5 ± 0.4 b | 9.00 ± 0.32 ab |
| BBCH-85 | 10.40 ± 0.21 ab | 160 ± 3.23 ab | 104.4 ± 2.3 ab | 77.7 ± 3.3 ab | 11.46 ± 0.22 ab | 26.3 ± 0.6 ab | 23.5 ± 0.2 a | 10.01 ± 0.32 ab |
| BBCH-89 | 11.56 ± 0.21 a | 173 ± 3.13 a | 116.5 ± 2.1 a | 86.2 ± 2.1 a | 12.86 ± 0.21 a | 29.7 ± 0.6 a | 26.8 ± 0.3 a | 11.13 ± 0.22 a |
| Significance | ||||||||
| C | 0.0126 * | 1.1134 *** | 1.50 × 10−12 *** | 0.0385 * | 8.18 × 10−10 *** | 0.0556 | 3.85 × 10−6 *** | 2.69 × 10−8 *** |
| PS | 6.05 × 10−10 *** | 2.51 × 10−10 *** | 4.28 × 10−13 *** | 3.3 × 10−7 *** | 2.67 × 10−14 *** | 1.09 × 10−12 *** | 6.64 × 10−15 *** | 4.09 × 10−7 *** |
| C × PS | 0.9985 | 0.9809 | 0.3270 | 0.9995 | 0.6026 | 0.9996 | 0.8717 | 0.8277 |
| Cultivar X (C) | IAA | ABA | GA3 | SA | Cytokinin | Zeatin | Jasmonic Acid |
|---|---|---|---|---|---|---|---|
| Italia | 2.97 ± 0.01 a | 2016 ± 21.8 b | 2.56 ± 0.03 | 3.09 ± 0.00 a | 3.72 ± 0.00 a | 0.994 ± 0.01 a | 10.52 ± 0.10 a |
| Bronx Seedless | 2.60 ± 0.02 b | 2801 ± 23.8 a | 2.52 ± 0.01 | 2.92 ± 0.01 b | 3.60 ± 0.03 b | 0.909 ± 0.02 b | 9.67 ± 0.11 b |
| Phenological stage Y (PS) | |||||||
| BBCH-77 | 2.10 ± 0.04 f | 1970 ± 42.6 e | 1.89 ± 0.01 f | 2.28 ± 0.03 f | 2.68 ± 0.03 f | 0.683 ± 0.03 f | 7.06 ± 0.17 f |
| BBCH-79 | 2.34 ± 0.04 e | 2127 ± 41.6 d | 2.12 ± 0.03 e | 2.53 ± 0.01 e | 3.01 ± 0.01 e | 0.773 ± 0.02 e | 8.06 ± 0.13 e |
| BBCH-81 | 2.60 ± 0.04 d | 2297 ± 43.6 c | 2.36 ± 0.01 d | 2.81 ± 0.03 d | 3.38 ± 0.03 d | 0.875 ± 0.03 d | 9.21 ± 0.12 d |
| BBCH-83 | 2.89 ± 0.04 c | 2481 ± 41.6 bc | 2.64 ± 0.03 c | 3.12 ± 0.01 c | 3.80 ± 0.02 c | 0.990 ± 0.04 c | 10.52 ± 0.27 c |
| BBCH-85 | 3.21 ± 0.04 b | 2680 ± 45.6 ab | 2.94 ± 0.04 b | 3.47 ± 0.03 b | 4.27 ± 0.04 b | 1.121 ± 0.01 b | 12.01 ± 0.13 b |
| BBCH-89 | 3.57 ± 0.04 a | 2894 ± 33.6 a | 3.29 ± 0.02 a | 3.85 ± 0.02 a | 4.79 ± 0.02 a | 1.269 ± 0.04 a | 13.72 ± 0.17 a |
| Significance | |||||||
| C | 1.11 × 10−10 *** | <2 × 10−16 *** | 0.1247 | 4.14 × 10−14 *** | 5.27 × 10−9 *** | 3.36 × 10−5 *** | 4.49 × 10−6 *** |
| PS | <2 × 10−16 *** | 8.62 × 10−13 *** | <2 × 10−16 *** | <2 × 10−16 *** | <2 × 10−16 *** | 6.23 × 10−16 *** | <2 × 10−16 *** |
| C × PS | 0.5898 | 0.9998 | 1.000 | 0.9212 | 0.9696 | 0.9460 | 0.8767 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yilmaz, T.; Ates, F.; Turan, M.; Hatterman-Valenti, H.; Kaya, O. Dynamics of Sugars, Organic Acids, Hormones, and Antioxidants in Grape Varieties ‘Italia’ and ‘Bronx Seedless’ during Berry Development and Ripening. Horticulturae 2024, 10, 229. https://doi.org/10.3390/horticulturae10030229
Yilmaz T, Ates F, Turan M, Hatterman-Valenti H, Kaya O. Dynamics of Sugars, Organic Acids, Hormones, and Antioxidants in Grape Varieties ‘Italia’ and ‘Bronx Seedless’ during Berry Development and Ripening. Horticulturae. 2024; 10(3):229. https://doi.org/10.3390/horticulturae10030229
Chicago/Turabian StyleYilmaz, Turhan, Fadime Ates, Metin Turan, Harlene Hatterman-Valenti, and Ozkan Kaya. 2024. "Dynamics of Sugars, Organic Acids, Hormones, and Antioxidants in Grape Varieties ‘Italia’ and ‘Bronx Seedless’ during Berry Development and Ripening" Horticulturae 10, no. 3: 229. https://doi.org/10.3390/horticulturae10030229
APA StyleYilmaz, T., Ates, F., Turan, M., Hatterman-Valenti, H., & Kaya, O. (2024). Dynamics of Sugars, Organic Acids, Hormones, and Antioxidants in Grape Varieties ‘Italia’ and ‘Bronx Seedless’ during Berry Development and Ripening. Horticulturae, 10(3), 229. https://doi.org/10.3390/horticulturae10030229