
Effects of Light Intensity and Photoperiod on Morphological Development and Photosynthetic Characteristics of Coriander


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
2.3. Sample Collection and Measurements
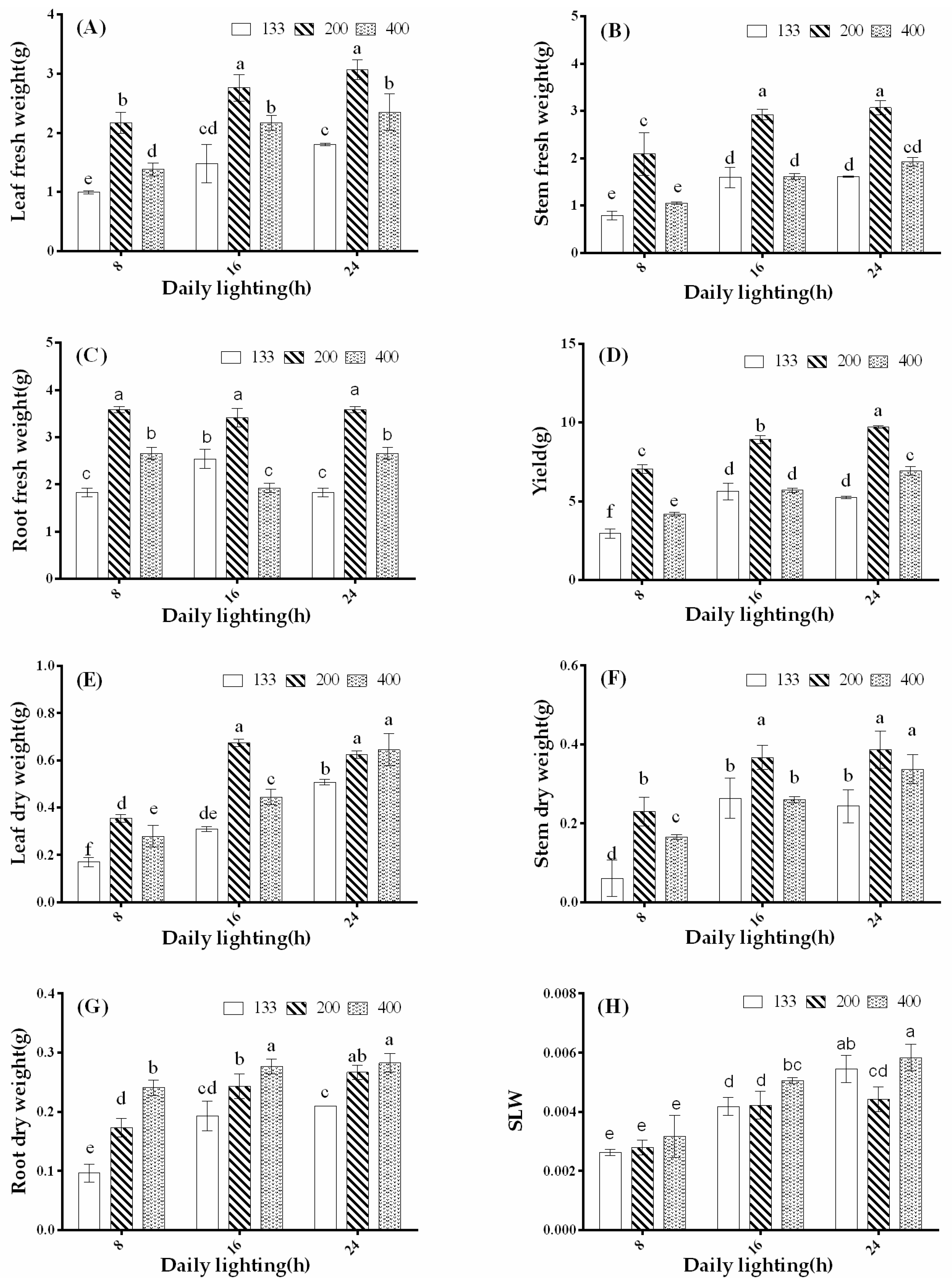
2.3.1. Morphological Characteristics and Yield Measurements
2.3.2. Photosynthetic Pigment Content
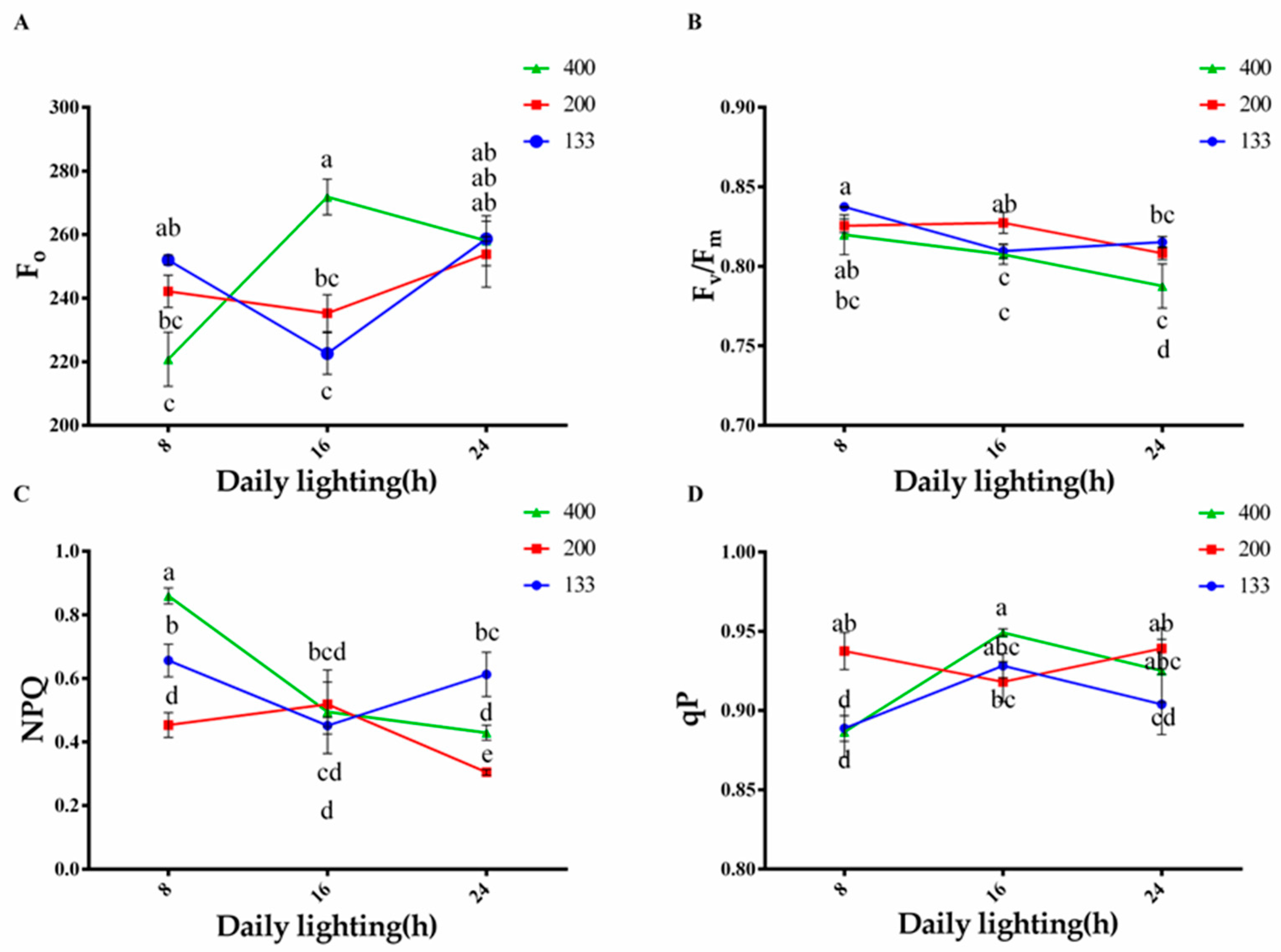
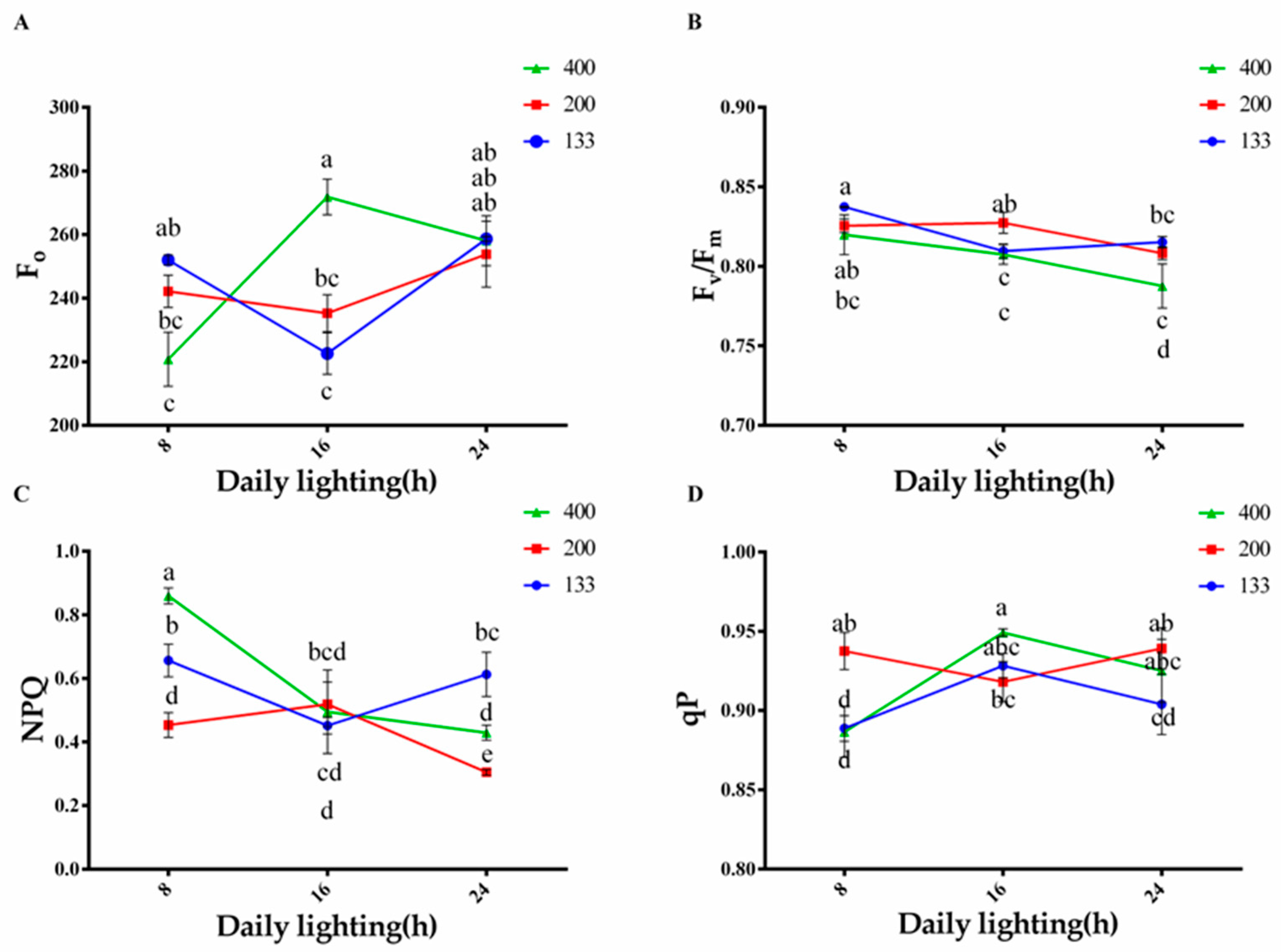
2.3.3. Chlorophyll Fluorescence
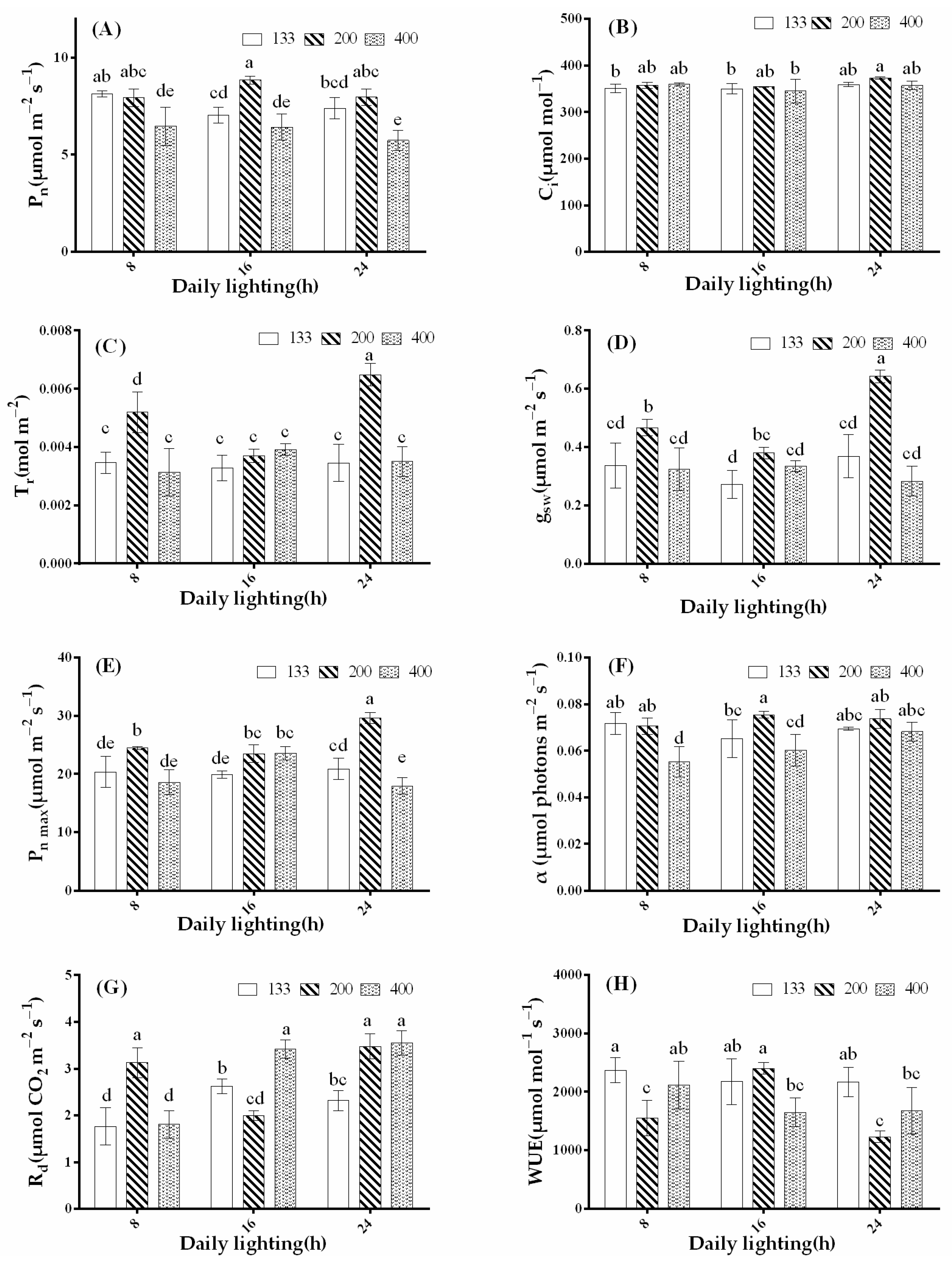
2.3.4. Photosynthetic Parameters
2.3.5. Stomata
2.3.6. Electric Energy Use Efficiency
2.3.7. Data Analysis
3. Results
3.1. Effects of Light Intensity and Photoperiod on Plant Morphology and Biomass
3.2. Effects of Light Intensity and Photoperiod on Plant Chlorophyll Content
3.3. Effects of Light Intensity and Photoperiod on Chlorophyll Fluorescence Content in Plants
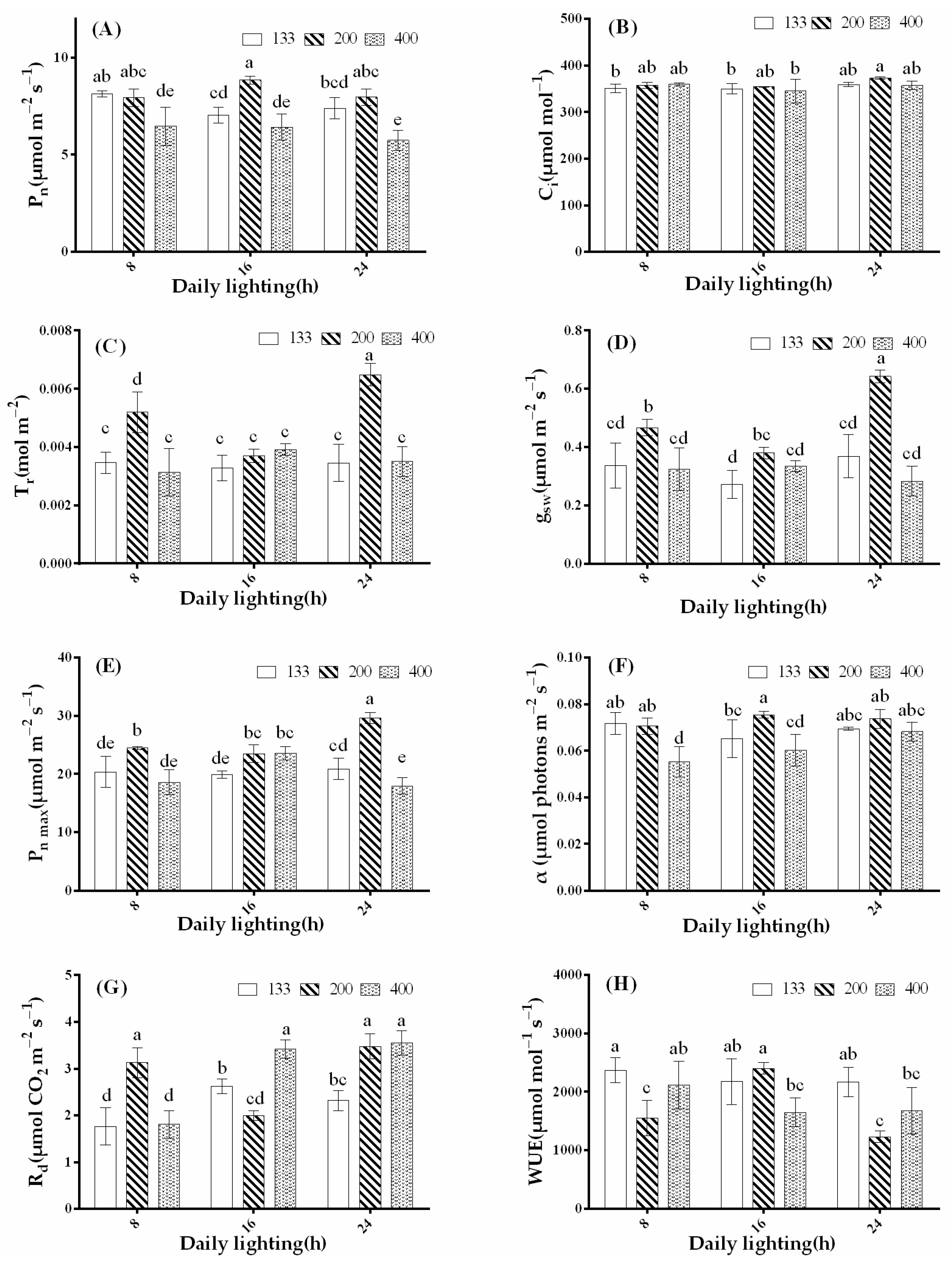
3.4. Effects of Light Intensity and Photoperiod on Plant Photosynthetic Characteristics
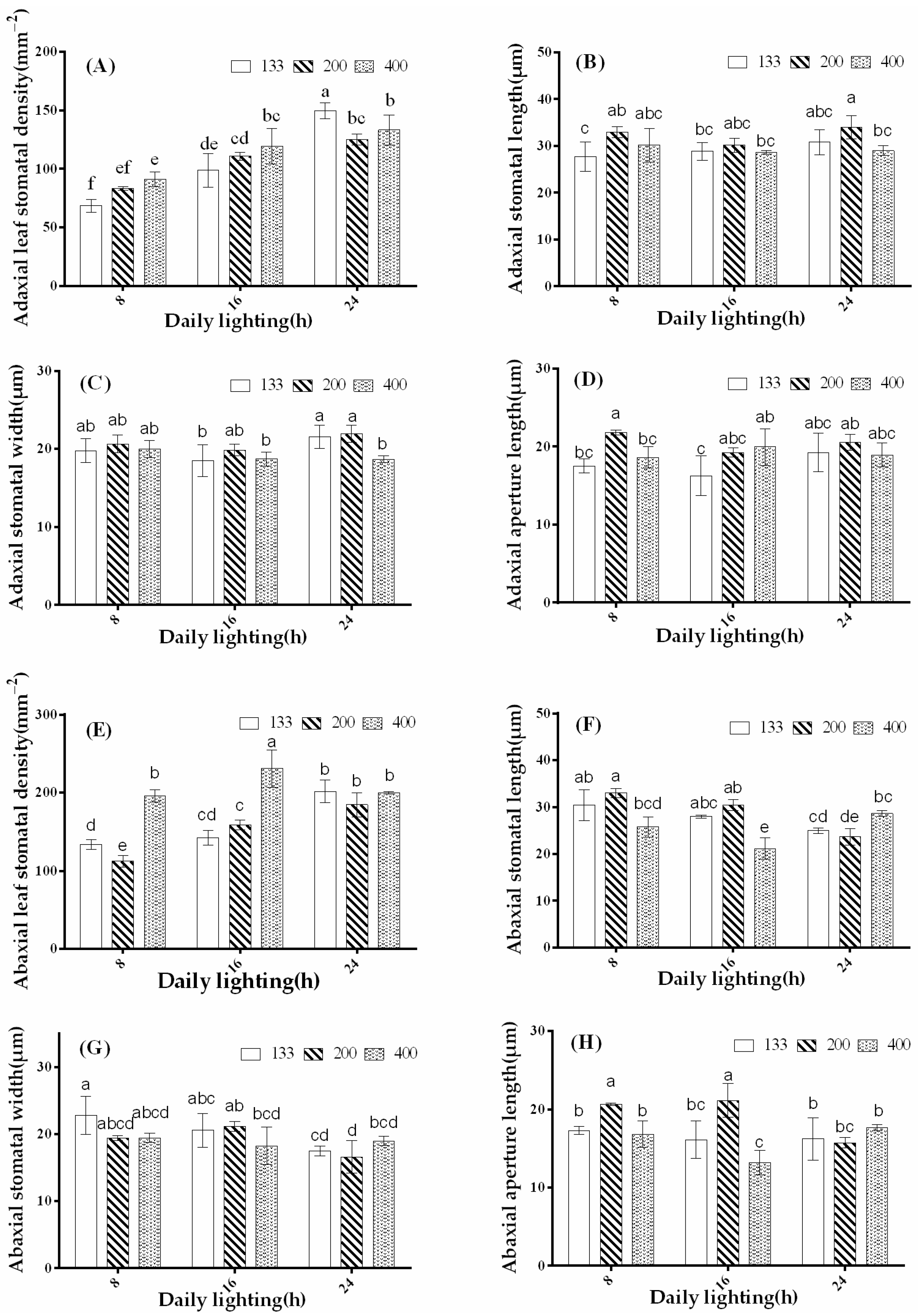
3.5. The Effects of Light Intensity and Photoperiod on Stomatal Development in Coriander Leaves
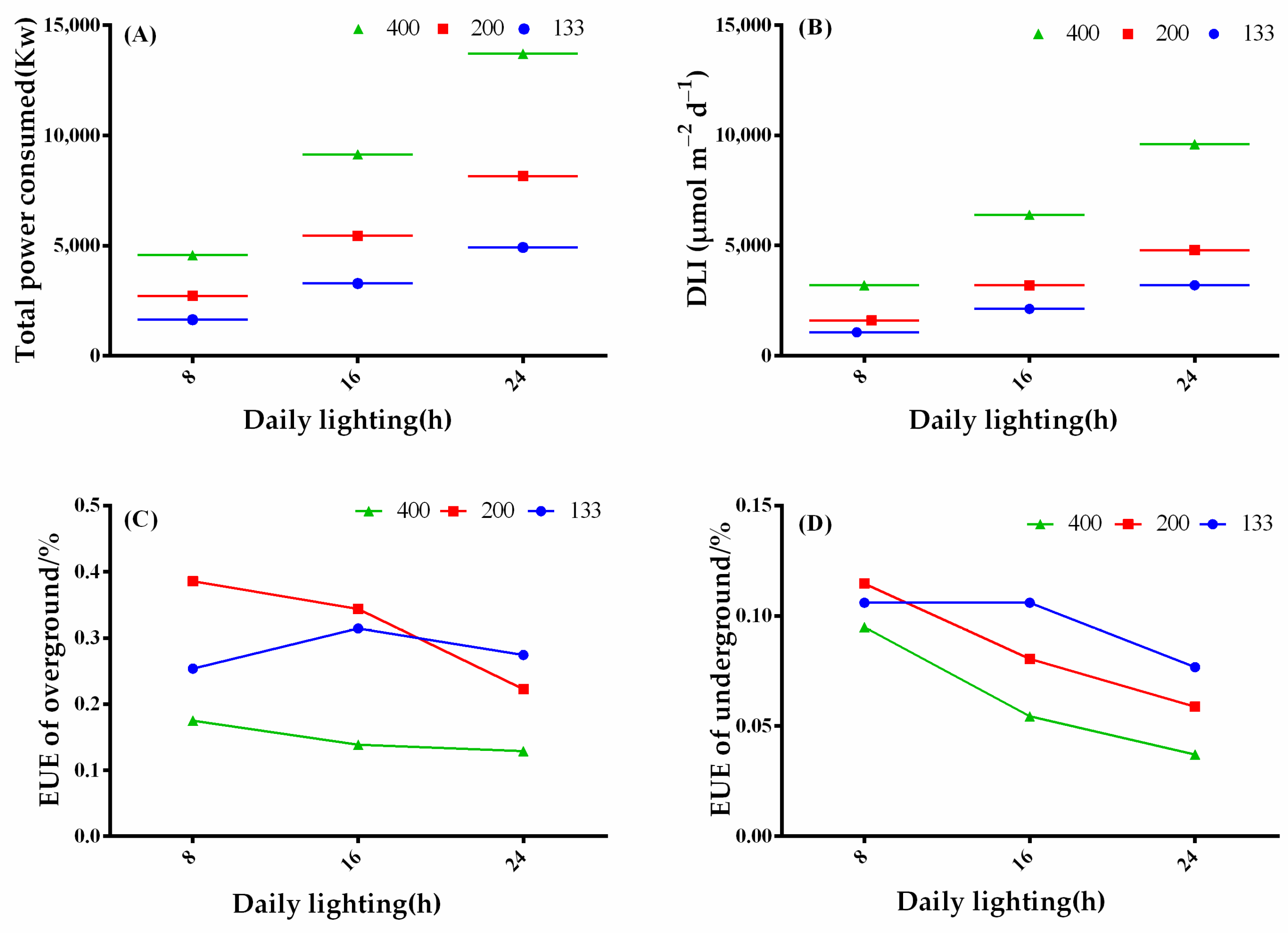
3.6. The Influence of Light Intensity and Photoperiod on Plant Energy Utilization Efficiency
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mandal, S.; Mandal, M. Coriander (Coriandrum sativum L.) essential oil: Chemistry and biological activity. Asian Pac. J. Trop. Biomed. 2015, 5, 421–428. [Google Scholar] [CrossRef]
- Laribi, B.; Kouki, K.; M'Hamdi, M.; Bettaieb, T. Coriander (Coriandrum sativum L.) and its bioactive constituents. Fitoterapia 2015, 103, 9–26. [Google Scholar] [CrossRef]
- Nadeem, M.; Muhammad Anjum, F.; Issa Khan, M.; Tehseen, S.; El-Ghorab, A.; Iqbal Sultan, J. Nutritional and medicinal aspects of coriander (Coriandrum sativum L.) A review. Br. Food J. 2013, 115, 743–755. [Google Scholar] [CrossRef]
- Al-Snafi, A.E. A review on chemical constituents and pharmacological activities of Coriandrum sativum. IOSR J. Pharm. 2016, 6, 17–42. [Google Scholar] [CrossRef]
- Duarte, A.; Luís, Â.; Oleastro, M.; Domingues, F.C. Antioxidant properties of coriander essential oil and linalool and their potential to control Campylobacter spp. Food Control 2016, 61, 115–122. [Google Scholar] [CrossRef]
- Prachayasittikul, V.; Prachayasittikul, S.; Ruchirawat, S.; Prachayasittikul, V. Coriander (Coriandrum sativum): A promising functional food toward the well-being. Food Res. Int. 2018, 105, 305–323. [Google Scholar] [CrossRef]
- Khare, M.; Tiwari, S.; Sharma, Y. Disease problems in the cultivation of coriander (Coriandrum sativum L.) and their management leading to production of high quality pathogen free seed. Int. J. Seed Spices 2017, 7, 1–7. [Google Scholar]
- Swisher Grimm, K.; Crosslin, J.; Cooper, W.; Frost, K.; du Toit, L.J.; Wohleb, C.H. First report of curly top of Coriandrum sativum caused by Beet curly top virus in the Columbia Basin of Washington State. Plant Dis. 2021, 105, 3313. [Google Scholar] [CrossRef]
- Masada, K.; Hosni, K.; Taarit, M.B.; Hammami, M.; Marzouk, B. Effects of crop season and maturity stages on the yield and composition of essential oil of coriander (Coriandrum sativum L.) fruit. Med. Aromat. Plant Sci. Biotechnol. 2012, 6, 115–122. [Google Scholar]
- Msaada, K.; Hosni, K.; Taarit, M.B.; Hammami, M.; Marzouk, B. Effects of growing region and maturity stages on oil yield and fatty acid composition of coriander (Coriandrum sativum L.) fruit. Sci. Hortic. 2009, 120, 525–531. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Pickett, K.M.; Caldwell, C.D.; Pincock, J.A.; Roberts, J.C.; Mapplebeck, L. Cultivar and sowing date effects on seed yield and oil composition of coriander in Atlantic Canada. Ind. Crops Prod. 2008, 28, 88–94. [Google Scholar] [CrossRef]
- Khodadadi, M.; Dehghani, H.; Javaran, M.J.; Christopher, J.T. Fruit yield, fatty and essential oils content genetics in coriander. Ind. Crops Prod. 2016, 94, 72–81. [Google Scholar] [CrossRef]
- Mikhail, A.F.; Pereboom, M.; Utsi, L.; Hawker, J.; Lighthill, J.; Aird, H.; Swindlehurst, M.; Greig, D.R.; Jenkins, C.; Godbole, G. Utility of whole-genome sequencing during an investigation of multiple foodborne outbreaks of Shigella sonnei. Epidemiol. Infect. 2021, 149, e71. [Google Scholar] [CrossRef]
- Mishra, B.K.; Dubey, P.; Aishwath, O.; Kant, K.; Sharma, Y.; Vishal, M. Effect of plant growth promoting rhizobacteria on coriander (Coriandrum sativum) growth and yield under semi-arid condition of India. Indian J. Agric. Sci 2017, 87, 607–612. [Google Scholar] [CrossRef]
- Szempliński, W.; Nowak, J.; Jankowski, K.J. Coriander (Coriandrum sativum L.) response to different levels of agronomic factors in Poland. Ind. Crops Prod. 2018, 122, 456–464. [Google Scholar] [CrossRef]
- Li, X.; Liang, T.; Liu, H. How plants coordinate their development in response to light and temperature signals. Plant Cell 2022, 34, 955–966. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Raschke, K. Effect of light quality on stomatal opening in leaves of Xanthium strumarium L. Plant Physiol. 1981, 68, 1170–1174. [Google Scholar] [CrossRef]
- Shibaeva, T.G.; Sherudilo, E.G.; Rubaeva, A.A.; Titov, A.F. Continuous LED lighting enhances yield and nutritional value of four genotypes of Brassicaceae microgreens. Plants 2022, 11, 176. [Google Scholar] [CrossRef]
- Zhang, X.; He, D.; Niu, G.; Yan, Z.; Song, J. Effects of environment lighting on the growth, photosynthesis, and quality of hydroponic lettuce in a plant factory. Int. J. Agric. Biol. Eng. 2018, 11, 33–40. [Google Scholar] [CrossRef]
- Dolezal, J.; Jandova, V.; Macek, M.; Liancourt, P. Contrasting biomass allocation responses across ontogeny and stress gradients reveal plant adaptations to drought and cold. Funct. Ecol. 2021, 35, 32–42. [Google Scholar] [CrossRef]
- Ye, Z.P.; Suggett, D.J.; Robakowski, P.; Kang, H.J. A mechanistic model for the photosynthesis–light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef]
- Savvides, A.; Fanourakis, D.; van Ieperen, W. Co-ordination of hydraulic and stomatal conductances across light qualities in cucumber leaves. J. Exp. Bot. 2012, 63, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Ding, J.; Qin, G.; Wang, Y.; Xi, L.; Yang, J.; Li, J.; Zhang, J.; Zou, Z. Interaction of supplementary light and CO2 enrichment improves growth, photosynthesis, yield, and quality of tomato in autumn through spring greenhouse production. HortScience 2019, 54, 246–252. [Google Scholar] [CrossRef]
- Tang, W.; Guo, H.; Baskin, C.C.; Xiong, W.; Yang, C.; Li, Z.; Song, H.; Wang, T.; Yin, J.; Wu, X. Effect of light intensity on morphology, photosynthesis and carbon metabolism of alfalfa (Medicago sativa) seedlings. Plants 2022, 11, 1688. [Google Scholar] [CrossRef]
- Bellstaedt, J.; Trenner, J.; Lippmann, R.; Poeschl, Y.; Zhang, X.; Friml, J.; Quint, M.; Delker, C. A mobile auxin signal connects temperature sensing in cotyledons with growth responses in hypocotyls. Plant Physiol. 2019, 180, 757–766. [Google Scholar] [CrossRef]
- Jeon, K.S.; Song, K.S.; Choi, K.S.; Kim, C.H.; Park, Y.B.; Kim, J.J. Growth and photosynthetic characteristics of Atractylodes japonica by light controls and leaf mold treatment in forest farming. Korean J. Med. Crop Sci. 2015, 23, 161–167. [Google Scholar] [CrossRef]
- Pan, T.; Wang, Y.; Wang, L.; Ding, J.; Cao, Y.; Qin, G.; Yan, L.; Xi, L.; Zhang, J.; Zou, Z. Increased CO2 and light intensity regulate growth and leaf gas exchange in tomato. Physiol. Plant. 2020, 168, 694–708. [Google Scholar] [CrossRef]
- Kircher, S.; Schopfer, P. Photosynthetic sucrose acts as cotyledon-derived long-distance signal to control root growth during early seedling development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 11217–11221. [Google Scholar] [CrossRef]
- Zheng, J.; Ji, F.; He, D.; Niu, G. Effect of light intensity on rooting and growth of hydroponic strawberry runner plants in a LED plant factory. Agronomy 2019, 9, 875. [Google Scholar] [CrossRef]
- Costigan, S.E.; Warnasooriya, S.N.; Humphries, B.A.; Montgomery, B.L. Root-localized phytochrome chromophore synthesis is required for photoregulation of root elongation and impacts root sensitivity to jasmonic acid in Arabidopsis. Plant Physiol. 2011, 157, 1138–1150. [Google Scholar] [CrossRef]
- Swarup, K.; Benková, E.; Swarup, R.; Casimiro, I.; Péret, B.; Yang, Y.; Parry, G.; Nielsen, E.; De Smet, I.; Vanneste, S. The auxin influx carrier LAX3 promotes lateral root emergence. Nat. Cell Biol. 2008, 10, 946–954. [Google Scholar] [CrossRef]
- Zha, L.; Zhang, Y.; Liu, W. Dynamic responses of ascorbate pool and metabolism in lettuce to long-term continuous light provided by red and blue LEDs. Environ. Exp. Bot. 2019, 163, 15–23. [Google Scholar] [CrossRef]
- Shao, M.; Liu, W.; Zhou, C.; Wang, Q.; Li, B. Alternation of temporally overlapped red and blue light under continuous irradiation affected yield, antioxidant capacity and nutritional quality of purple-leaf lettuce. Sci. Hortic. 2022, 295, 110864. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Ač, A.; Marek, M.V.; Kalina, J.; Urban, O. Differences in pigment composition, photosynthetic rates and chlorophyll fluorescence images of sun and shade leaves of four tree species. Plant Physiol. Biochem. 2007, 45, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Pham, D.M.; Chun, C. Growth and leaf injury in tomato plants under continuous light at different settings of constant and diurnally varied photosynthetic photon flux densities. Sci. Hortic. 2020, 269, 109347. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Pang, J.; Huo, Z. Effect of low light on photosynthesis and membrane-lipid peroxidation of Cucumis sativus seedling. Acta Agric. Univ. Henanensis 1998, 32, 68–72. [Google Scholar]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.K.; Huemmrich, K.F.; Middleton, E.M.; Ward, L.A.; Julitta, T.; Daughtry, C.S.; Burkart, A.; Russ, A.L.; Kustas, W.P. Diurnal and seasonal variations in chlorophyll fluorescence associated with photosynthesis at leaf and canopy scales. Remote Sens. 2019, 11, 488. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Van Wittenberghe, S.; Alonso, L.; Verrelst, J.; Moreno, J.; Samson, R. Bidirectional sun-induced chlorophyll fluorescence emission is influenced by leaf structure and light scattering properties—A bottom-up approach. Remote Sens. Environ. 2015, 158, 169–179. [Google Scholar] [CrossRef]
- Paunov, M.; Koleva, L.; Vassilev, A.; Vangronsveld, J.; Goltsev, V. Effects of different metals on photosynthesis: Cadmium and zinc affect chlorophyll fluorescence in durum wheat. Int. J. Mol. Sci. 2018, 19, 787. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Li, P.; Wu, Y. Effects of different light intensities on chlorophyll fluorescence characteristics and yield in lettuce. Sci. Hortic. 2012, 135, 45–51. [Google Scholar] [CrossRef]
- Souza, R.; Machado, E.; Silva, J.; Lagôa, A.; Silveira, J. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ. Exp. Bot. 2004, 51, 45–56. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Magney, T.S.; Barnes, M.L.; Yang, X. On the covariation of chlorophyll fluorescence and photosynthesis across scales. Geophys. Res. Lett. 2020, 47, e2020GL091098. [Google Scholar] [CrossRef]
- Roelfsema, M.R.G.; Hanstein, S.; Felle, H.H.; Hedrich, R. CO2 provides an intermediate link in the red light response of guard cells. Plant J. 2002, 32, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.S.; Driscoll, S.P.; Olmos, E.; Harbinson, J.; Arrabaça, M.C.; Foyer, C.H. Adaxial/abaxial specification in the regulation of photosynthesis and stomatal opening with respect to light orientation and growth with CO2 enrichment in the C4 species Paspalum dilatatum. New Phytol. 2008, 177, 186–198. [Google Scholar] [CrossRef]
- Wang, Y.; Noguchi, K.; Terashima, I. Photosynthesis-dependent and-independent responses of stomata to blue, red and green monochromatic light: Differences between the normally oriented and inverted leaves of sunflower. Plant Cell Physiol. 2011, 52, 479–489. [Google Scholar] [CrossRef]
- Urban, J.; Ingwers, M.W.; McGuire, M.A.; Teskey, R.O. Increase in leaf temperature opens stomata and decouples net photosynthesis from stomatal conductance in Pinus taeda and Populus deltoides x nigra. J. Exp. Bot. 2017, 68, 1757–1767. [Google Scholar] [CrossRef]
- Hanba, Y.; Kogami, H.; Terashima, I. The effect of growth irradiance on leaf anatomy and photosynthesis in Acer species differing in light demand. Plant Cell Environ. 2002, 25, 1021–1030. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Photoperiod × Light Intensity | Plant Height /cm | Stem Diameter/mm | Leaf Area /cm2 | Branching Number |
|---|---|---|---|---|
| h × μmol·m−2·s−1 | ||||
| 8 × 133 | 5.80 ± 0.38 d | 4.64 ± 0.43 e | 70.19 ± 7.14 de | 9.10 ± 1.14 cd |
| 16 × 133 | 8.24 ± 0.31 c | 5.95 ± 0.76 d | 74.51 ± 7.99 e | 8.60 ± 055 d |
| 24 × 133 | 8.30 ± 0.27 c | 5.67 ± 0.30 d | 93.17 ± 5.54 cde | 8.4 ± 0.55 d |
| 8 × 200 | 12.20 ± 0.45 a | 7.83 ± 0.57 c | 140.36 ± 8.47 ab | 11.60 ± 0.55 b |
| 16 × 200 | 11.00 ± 1.0 b | 9.31 ± 0.52 a | 160.63 ± 16.82 a | 14.00 ± 0.71 a |
| 24 × 200 | 12.68 ± 1.01 a | 8.26 ± 0.09 bc | 156.74 ± 10.63 a | 10.40 ± 0.89 bc |
| 8 × 400 | 5.50 ± 0.35 d | 5.46 ± 0.31 d | 111.00 ± 14.73 bcd | 10.80 ± 0.84 b |
| 16 × 400 | 8.50 ± 0.87 c | 8.84 ± 0.90 ab | 87.93 ± 5.99 cde | 10.40 ± 0.55 bc |
| 24 × 400 | 8.50 ± 0.35 c | 6.20 ± 0.56 d | 120.25 ± 18.35 bc | 10.9 ± 0.65 b |
| Photoperiod × Light Intensity | Chlorophyll a /mg·g−1 | Chlorophyll b /mg·g−1 | Carotenoid /mg·g−1 | Total Chlorophyll /mg·g−1 |
|---|---|---|---|---|
| /h × μmol·m−2·s−1 | ||||
| 8 × 133 | 0.89 ± 0.07 ab | 0.24 ± 0.01 ab | 0.19 ± 0.01 abc | 1.14 ± 0.09 ab |
| 16 × 133 | 0.94 ± 0.10 a | 0.22 ± 0.03 bc | 0.21 ± 0.02 a | 1.16 ± 0.13 ab |
| 24 × 133 | 0.77 ± 0.07 bcd | 0.19 ± 0.01 de | 0.18 ± 0.02 bc | 0.96 ± 0.08 c |
| 8 × 200 | 0.95 ± 0.15 a | 0.24 ± 0.04 ab | 0.20 ± 0.03 ab | 1.19 ± 0.19 a |
| 16 × 200 | 0.80 ± 0.11 bc | 0.21 ± 0.03 cd | 0.17 ± 0.02 a | 1.01 ± 0.15 bc |
| 24 × 200 | 0.97 ± 0.06 a | 0.26 ± 0.02 a | 0.21 ± 0.02 a | 1.23 ± 0.08 a |
| 8 × 400 | 0.74 ± 0.07 cd | 0.17 ± 0.01 e | 0.17 ± 0.02 bc | 0.91 ± 0.09 cd |
| 16 × 400 | 0.78 ± 0.05 bcd | 0.18 ± 0.01 de | 0.19 ± 0.02 ab | 0.96 ± 0.05 c |
| 24 × 400 | 0.65 ± 0.12 d | 0.14 ± 0.02 f | 0.16 ± 0.02 c | 0.79 ± 0.14 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Gao, Q.; Ji, G.; Wang, J.; Ding, Y.; Wang, S. Effects of Light Intensity and Photoperiod on Morphological Development and Photosynthetic Characteristics of Coriander. Horticulturae 2024, 10, 215. https://doi.org/10.3390/horticulturae10030215
Wang F, Gao Q, Ji G, Wang J, Ding Y, Wang S. Effects of Light Intensity and Photoperiod on Morphological Development and Photosynthetic Characteristics of Coriander. Horticulturae. 2024; 10(3):215. https://doi.org/10.3390/horticulturae10030215
Chicago/Turabian StyleWang, Fang, Qi Gao, Guangsi Ji, Jingxuan Wang, Yifeng Ding, and Sen Wang. 2024. "Effects of Light Intensity and Photoperiod on Morphological Development and Photosynthetic Characteristics of Coriander" Horticulturae 10, no. 3: 215. https://doi.org/10.3390/horticulturae10030215
APA StyleWang, F., Gao, Q., Ji, G., Wang, J., Ding, Y., & Wang, S. (2024). Effects of Light Intensity and Photoperiod on Morphological Development and Photosynthetic Characteristics of Coriander. Horticulturae, 10(3), 215. https://doi.org/10.3390/horticulturae10030215