Abstract
Ploidy breeding is one of the important approaches for persimmon (Diospyros kaki Thunb.) genetic improvement, and vegetative growth of seedlings is the basis for subsequent fruit development. Therefore, the physiological characteristics and transcriptional differences of seedling growth traits in different ploidy persimmon germplasm were studied in this study, which provided a theoretical basis for fruit evaluation and breeding of new polyploid persimmon varieties. Nonuploid and its full-sib hexaploid seedlings obtained from endosperm culture were used as materials. By observing plant phenotype, leaf tissue section, endogenous hormone content, and transcriptome sequencing, the phenotype and physiological characteristics of different ploidy Persimmon seedlings were compared, as well as the differences in transcription levels. (1) Compared with hexaploid seedlings, the nonuploid were more robust and compact, and the leaves were obviously thicker. The cell size of leaf veins and parenchyma were significantly different between the different ploidy plants. (2) The contents of Salicylic Acid (SA), Jasmonic Acid (JA), gibberellin A1 (GA1), gibberellin A3 (GA3) and Indole-3-acetic acid (IAA) in nonuploid leaves were significantly higher than those in hexaploid leaves, while the contents of cytokinin trans-zeanoside (Tzt) and dihydrozeanoside (DZR), N6-isopentenyladenine (iP) and Jasmonoyl-L-isoleucine (JA-ILE) in nonuploid leaves were significantly lower than those in hexaploid leaves. (3) A total of 5796 differentially expressed genes were identified in nonuploid and hexaploid leaves. These differentially expressed genes were mainly related to photosynthesis, plant-pathogen interaction, etc. Among them, YUCCA genes, GA3ox genes, and IPT genes related to hormone synthesis were significantly differentially expressed in the nonuploid and hexaploid leaves. It is speculated that it may be the key regulatory gene that leads to the difference in IAA, gibberellin (GA), and indolepropionic acid (IPA) levels in the nonuploid and the hexaploid. The growth traits of the new Persimmon germplasms with different ploidy were significantly different. The nonuploid plants were shorter and more compact, and the leaves were larger and thicker. These traits were closely related to the content of endogenous hormones, and the balance of endogenous hormones was affected by gene expression. In addition, based on the biological processes involved in hormones and differentially expressed genes, it is speculated that the nonuploid may be superior to the hexaploid in terms of resistance.
1. Introduction
Diospyros kaki Thunb. belongs to the genus Diospyros of the Ebenaceae family and is one of the important economic forest species [1]. Persimmon cultivars are mostly hexaploid (2n = 6x = 90), and very few are nonuploid (2n = 9x = 135), such as the famous Japanese varieties ‘Hiratanenashi’ and ‘Fukuoka K. 1 Gou-Akiou’ [2]. The nonuploid of persimmon is actually an intra-specific triploid, which is theoretically formed by the fusion of an unreduced gamete (2n) and a haploid gamete (n). The fruit of nonuploid persimmon is naturally seedless, which gives it a significant advantage in fresh food and processing [3]. Nonuploid cultivars bred by natural selection or artificial creation showed great utilization value and development potential with their excellent comprehensive characteristics, and it also showed that polyploid breeding is an important way for persimmon to create excellent new germplasm.
Polyploid breeding is an efficient breeding technique commonly used in economic forests. Polyploid germplasms often show the characteristics of vigorous growth, huge organs, increased biochemical component content, seedless, and enhanced adaptability to the environment [4]. In many polyploid plants with different ploidy, triploid often combines the ploidy effect and heterozygous effect, and the comprehensive performance is more prominent, which has higher development and utilization value. Eucommia triploids were superior to diploid plants in terms of vegetative growth, leaf traits, photosynthetic rate, and secondary metabolites, among which the average contents of chlorogenic acid, aucubin, geniposide and geniposidic acid in the triploid leaves were 10.20%, 1.34%, 118.22% and 22.04% higher than those in the diploid controls, respectively [5]. The synthetic triploid carambola (Averrhoa carambola L.) was superior to the diploid control in branch number, leaf thickness, and fruit size [6]. Triploid and diploid citrus varieties obtained from ‘Fortune’ mandarin and ‘Ellendale’ tangor hybridization have shown different physiological characteristics. Compared with diploid citrus, triploid citrus has lower stomatal density, larger stomatal size, increased chloroplast number, enhanced photosynthetic capacity and water holding capacity, and showed stronger water stress tolerance [7]. At present, triploid has been widely used in more than 40 kinds of horticultural plant crops, including fruit trees, vegetables, flowers, and other tree species, such as pear, apple, citrus, grape, loquat, beet, watermelon, lily, peony, etc. [8].
Polyploid phenotype and adaptation variation are related to the differential expression of genes. The process of polyploidy is generally accompanied by the loss and rearrangement of a large number of genes or the subfunctionalization and new functionalization of multi-copy genes so that they have stronger genetic diversity and plasticity [9]. Currently, research on gene differential expression caused by plant polyploidization has attracted increasing attention. Triploid poplar has significant advantages in leaf development and photosynthetic characteristics. Based on leaf phenotype and transcriptome data, weighted correlation network analysis (WGCNA) analysis revealed a strong correlation between PpnGRF5-1 and leaf development and net photosynthetic rate (Pn). Ppngrf5-1 can up-regulate the expression of the chlorophyll synthesis gene and down-regulate the transcription of chlorophyll degrading enzyme [10]. The fruit of triploid Siraitia grosvenorii was smaller than that of diploid fruits. Transcriptome analysis showed that genes related to the biosynthesis of auxin, gibberellin, and cytokinin were significantly down-regulated in triploid fruits, which is consistent with the determination trend of corresponding hormone content, indicating that the down-regulation of genes related to the biosynthesis of auxin, gibberellin and cytokinin in triploid fruits may lead to the phenomenon of smaller fruits [11]. The hormone content in tomato autotetraploid is 30% lower than that in diploid [12]. A study on the hormone content of three different ploidy cucumbers found that the levels of IAA, IPA, and Zeatin riboside (ZR) in allotetraploid cucumbers were lower than those in diploid hybrids, while the levels of IAA and IPA in allotriploid cucumbers were between tetraploid and diploid, and the ZR content was lower than both [13]. Studying the gene expression and regulatory pattern variations in polyploidy and analyzing the molecular mechanisms of polyploid trait advantages has important guiding significance for creating excellent polyploid germplasm. At present, although there have been reports on nonuploid Persimmon varieties, basic physiological and molecular studies on their growth characteristics have not been reported. Therefore, in this study, the mature leaves of nonuploid Persimsima and its full-sib hexaploid were used as materials to observe tissue sections, determine endogenous hormones, and analyze the cytological and physiological basis of phenotypic differences in different ploidy leaves. Further, transcriptome sequencing (RNA-seq) was used to analyze the gene expression differences between the leaves of the nonuploid and the hexaploid, identify the key regulatory genes affecting the growth of leaves and plants, and reveal the differences in transcription levels of different ploidy growth characteristics, laying a foundation for the molecular mechanism analysis of dominant traits in nonuploid persimmon growth.
2. Materials and Methods
2.1. Plant Materials
The experimental materials were the nonuploid and the full-sib hexaploid plants obtained by tissue culture from the endosperm of the immature seeds of D. kaki cv. ‘Fuyu’ × D. kaki cv. ‘Zhongshi male 1’. The parents were planted in the Yuanyang long-term experimental station (Yuanyang Country, Henan Province, China, 34°55′30″–34°56′45″ N, 113°46′24″–113°47′59″ E) of the Research Institute of Non-timber Forestry, Chinese Academy of Forestry. The nonuploid and hexaploid seedlings were planted in the same seedling bag, and the growing environment and tending conditions were the same.
2.2. Observation of Plant Phenotype and Leaf Phenotype
The plant and leaf phenotypes of the nonuploid and hexaploid plants were observed during their vigorous growth period. The height of the plant was measured by vernier calipers, and the length, width, area, and circumference of the mature leaves were analyzed using the leaf area analysis system (Wseen, Hangzhou, China).
2.3. Observation of Leaf Tissue by Paraffin Section
The mature leaves of hexaploid and nonaploid plants were fixed by FAA (70% ethanol/acetic acid/formaldehyde, 90:5:5) for 24 h and dehydrated in the graded ethanol series. The sample was then soaked in paraffin wax at 60 °C three times, each time for 2 h [14]. Sections 5 μm thick were prepared with Leica RM2265 microtome (Leica Microsystems, Nussloch, Germany) and mounted on clean glass slides. Slices were removed from the wax using xylene and ethanol and stained overnight with hematoxylin. After staining with 1% eosin for 20 s, it was dehydrated with ethanol and xylene. The microslides were dried and mounted with a lid. The stained sections were observed and photographed under an optical microscope (Olympus, Tokyo, Japan).
2.4. Determination of Endogenous Hormone Content in Leaves Using HPLC-ESI-MS/MS Method
The mature leaves were frozen in liquid nitrogen and stored at −80 °C for analysis of endogenous hormones. Each sample had three biological replicates. High-performance liquid chromatography-electrospray ionization tandem mass spectrometry (HPLC–ESI–MS/MS) in multiple-reaction monitoring (MRM) mode was used to quantify the endogenous phytohormone levels. Endogenous phytohormone include Indole-3-acetic acid (IAA), Abscisic Acid (ABA), Jasmonic Acid (JA), Jasmonoyl-L-isoleucine (JA-ILE), Salicylic Acid (SA), Zeatin (Zt), Trans-Zeatin riboside (tZR), Trans-zeatin (tZ), N6-isopentenyladenine (iP), N6-isopentenyladenenosine (iPR), gibberellin A1 (GA1), gibberellin A3 (GA3), and gibberellin A4 (GA4). The extraction and purification of hormones are carried out according to the method described by Sun [15]. The HPLC–ESI–MS/MS consisted of an Agilent 1290 HPLC system (Agilent Technologies, Santa Clara, CA, USA) and an AB Qtrap 6500 triple quadrupole mass spectrometer (AB Sciex LLC, Framingham, MA, USA) with an electrospray ionization source. Samples were injected into a poroshell 120 SB-C18 (2.1 × 150, 2.7 um); Column temperature: 30 °C; Mobile phase: A:B = (methanol/0.1% formic acid): (water/0.1% formic acid); Flow rate 0.3 mL/min; The injection volume was 2 μL. Mass spectrum parameters: ionization mode was ESI positive and negative ion mode respectively, scanning type was MRM, gas curtain gas was 15 psi, spray voltage was +4500 V, −4000 V, atomizing gas pressure was 65 psi, auxiliary gas pressure was 70 psi, atomizing temperature was 400 °C. Using MultiQuant (Framingham, MA, USA) software for automatic identification, draw a linear regression standard curve with the mass spectrometry peak area of the analyte as the y-axis and the mass of the analyte as the x-axis. Sample concentration calculation: Substitute the mass spectrometry peak area of the sample analyte into a linear equation to calculate the hormone mass in the sample. All the endogenous hormone content was determined by Nanjing Ruiyuan Biotechnology Co., Ltd. (Nanjing, China).
2.5. Transcriptome Sequencing
The mature leaves were frozen in liquid nitrogen and stored at −80 °C for analysis of transcriptome sequencing. Each sample had three biological replicates. Total RNA was extracted by UNIQ-10 column Trizol total RNA extraction kit (B511321, Shanghai, China), and the integrity and total amount were detected by Agilent 2100 bioanalyzer. Total RNA was used for library construction, and Illumina sequencing was performed after quality inspection through Illumina NovaSeq 6000 (Illumina Inc., San Diego, CA, USA). The raw data obtained by sequencing were filtered to obtain clean data. Sequence alignment using the hexaploid persimmon genome as a reference genome [16]. The Fragment Per Kilobase of exon per million fragments mapped (FPKM) of each gene was calculated, and differential expression analysis between the two samples was performed using DESeq2 software (1.20.0). After correction, p value ≤ 0.05 and |log2-foldchange| ≥ 1 were taken as the threshold of significant differential expression. GO enrichment analysis of differentially expressed genes was performed by clusterProfiler (3.8.1) software, and statistical enrichment of differentially expressed genes in the KEGG pathway was analyzed by clusterProfiler (3.8.1) software.
2.6. Statistical Analysis
The Excel 2019 software is used to calculate the average value and standard deviation. SPSS 23.0 software was used for variance analysis, independent sample T-test (p < 0.05 level), and Excel and GraphPad Prism 6 were used for mapping.
3. Results
3.1. Comparison of Plant Phenotypes and Leaf Tissue Sections between Nonuploid and Hexaploid Persimmons
Compared to hexaploid plants, the nonuploid plants were thicker and more compact, with large and significantly thicker leaves (Figure 1). For 1-year-old seedlings, the average height of the nonuploid plants was 92 mm, while the hexaploid plants were significantly higher than the nonuploid plants, reaching 154 mm (Table 1). There were also significant differences in leaf phenotypes between the nonuploid and hexaploid plants. The average leaf length of hexaploid plants is 49.0 mm, while the leaves of nonuploid plants are longer (67.7 mm). The leaf area of nonuploid plants is 1391 mm2, significantly larger than that of hexaploid plants (1002 mm2) (Table 2). However, there was no significant difference in the width of leaves between the nonuploid and hexaploid plants.
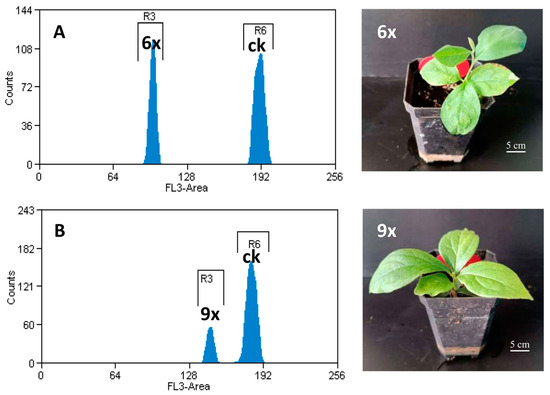
Figure 1.
Flow cytometry results and plant phenotypes of nonuploid and hexaploid seedlings. (A) hexaploidy, (B) nonuploid. The genome size of CK (tobacco) is 4.5 Gb, the genome size of hexaploid is 2.3 Gb, and the genome size of nonuploid is 3.6 Gb.

Table 1.
Comparison of growth and leaf phenotype data of nonuploid and hexaploid plants.
Table 2.
Comparison of leaf vein cell size.
In order to further understand the phenotypic differences between nonuploid and hexaploid leaves, paraffin sections of the leaves were performed. Results as shown in Figure 2a, the veins of the leaves of the nonuploid were significantly different from those of the hexaploid, with the area of the xylem in the nonuploid being 2.5 times that of the hexaploid. In addition, the shape and size of the mesophyll cells were also significantly different between the nonuploid and hexaploid (Figure 2b). The thickness of the epidermal cells in the mesophyll of the nonuploid was 13.6% larger than that of the hexaploid, showing a significant difference (Table 3). The lower epidermis was arranged neatly. Mesophyll cells near the lower epidermis were of different sizes, mostly irregular in shape but neatly arranged, with significant differences. The mesophyll palisade tissue cells of nonuploid leaves were arranged closely, with different shapes and sizes. The palisade tissue cells of hexaploid leaves were loosely arranged cylinders with large cell gaps, and the shape and size of the cells were consistent. In addition, there were significant differences in the mesophyll spongy tissue between the nonuploid and hexaploid leaves. The cells of the nonuploid sponge are large and dense, and the number of cells is about twice that of the hexaploid cells. However, hexaploid sponge tissue cells are small and sparse. There were no significant changes in the mesophyll parenchyma of nonuploid and hexaploid. The mesophyll cells near the main vein were obviously larger than the surrounding mesophyll cells, and the epidermal cells near the main vein were nearly round, while the rest of the epidermal cells were mostly elongated.
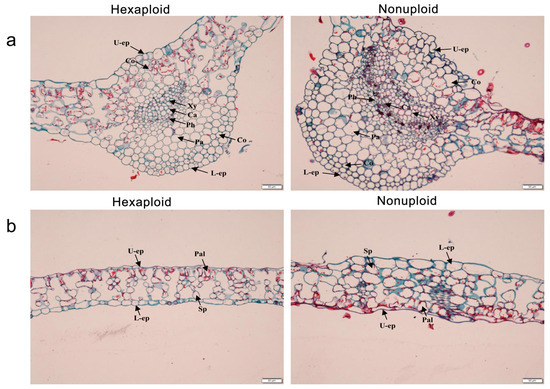
Figure 2.
Anatomical characterization of hexaploid and nonuploid leaves. (a) vein paraffin section (b) mesophyll paraffin section Note: L-ep. Lower epidermal cells; U-ep. Upper epidermal cells; Xy. Xylem; Pa. Parenchyma; Co. Thick angular tissue; Palisade tissue; Sp. Spongy tissue. Pal. palisade cell; Ca. cambium; Ph. phloem.
Table 3.
Comparison of mesophyll cell size.
3.2. Comparative Analysis of Endogenous Hormone Content in Leaves of Nonuploid and Hexaploid Plants
Plant endogenous hormones are trace organic compounds synthesized in plants with signal function, which act on specific protein receptors and regulate plant growth and development. To compare the content of endogenous hormones in hexaploid and nonuploid leaves, Auxin and its precursors (IAA and TAM), abscisic acid (ABA), jasmonic acid and its precursors (JA, MEJA, JA-ILE and OPDA), salicylic acid (SA and MESA), cytokinin (TZR, DZR, T-Zeatin, IP and IPA), and gibberellin (GA1 and GA3) were determined by UHPLC-ESI-MS/MS. Results as shown in Figure 3, there was a significant difference in the content of SA between the nonuploid and the hexaploid, with the average content of 464.98 ng·g−1 in the nonuploid and 94.81 ng·g−1 in the hexaploid, and the SA content in the nonuploid was nearly five times that in the hexaploid. The content of JA was 28.81 ng·g−1 in nonuploid and 6.38 ng·g−1 in hexaploid, which was significantly higher than that in hexaploid (4.5 times). The contents of OPDA, MEJA, GA1, GA3, and IAA in the leaves of the two ploidy plants were lower, but the contents of these hormones in nonuploid were still higher than those in the hexaploid. On the contrary, the content of JA-ILE in hexaploid leaves was 375.54 ng·g−1, which was three times that of nonuploid leaves (122.57 ng·g−1). The content of Tzt and DZR in hexaploid was 52.31 ng·g−1 and 26.46 ng·g−1, respectively, which was also three times that in hexaploid (16.46 ng·g−1 and 8.01 ng·g−1). The content of TAM was 12.15 ng·g−1 in hexaploid and 0.94 ng·g−1 in nonuploid. In addition, the content of ABA, IP, and IPA in hexaploid was slightly higher than that in nonuploid. ZT and IBA were not detected in both samples. In summary, there were significant differences in the contents of multiple hormones between the nonuploid and hexaploid plants, and the contents of SA, JA, GA1, GA3, and IAA in the nonuploid leaves were significantly higher than those in the hexaploid leaves. The contents of Tzt, DZR, and JA-ILE in the leaves of nonuploid were significantly lower than those of hexaploid leaves. It is suggested that the growth characteristics of persimmon with different ploidy may be synergically regulated by various plant hormones.
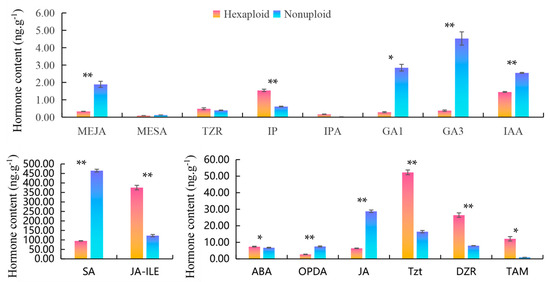
Figure 3.
Comparison of the contents of endogenous hormones in leaves of nonuploid and hexaploid persimmons. Methyl Jasmonate (MEJA), Methyl salicylate (MESA), Trans-Zeatin riboside (TZR) N6-isopentenyladenine (IP), Indolepropionic acid (IPA), Gibberellin A1 (GA1), Gibberellin A3 (GA3), In-dole-3-acetic acid (IAA), Salicylic Acid (SA), Jasmonoyl-L-isoleucine (JA-ILE), Abscisic Acid (ABA), cis(+)-12-oxophytodienoic acid (OPDA), Jasmonic Acid (JA), Trans-zeanoside (Tzt), Dihydrozeanoside (DZR), Tryptamine (TAM). * is p < 0.05 ** is p < 0.01.
3.3. Analysis of Gene Expression Difference between Hexaploid and Nonuploid Persimmons
3.3.1. Identification of Differentially Expressed Genes
According to the screening criteria (padj < 0.05, |log2FoldChange| > 1) for differentially expressed genes, the gene expression variation in the leaves of nonuploid and hexaploid were analyzed. As shown in Figure 4, there were 17,728 genes with no significant difference in expression level between nonuploid and hexaploid persimmon. A total of 5796 differentially expressed genes were screened, among which 2756 genes were up-regulated and 3040 genes were down-regulated in the nonuploid.
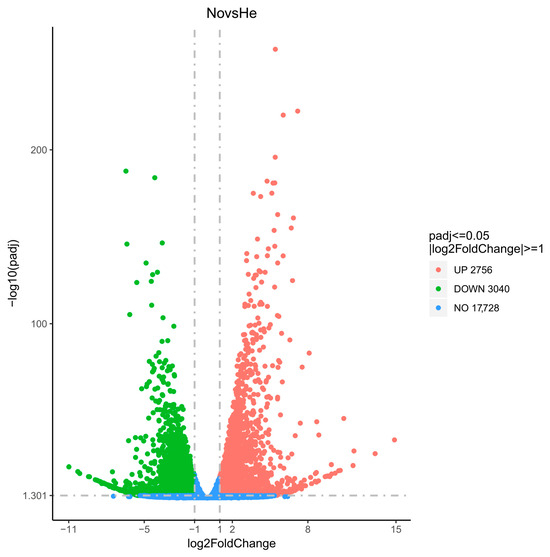
Figure 4.
The volcano plot shows the number of DEGs between nonuploid and hexaploid persimmons.NovsHe is Nonuploid vs. Hexaploid.
3.3.2. GO and KEGG Entichment Analysis of DEGs
GO enrichment analysis of DEGs was performed, padj < 0.05 represented a significant difference. The results were shown in Figure 5: the upregulated differential genes in the nonuploid were mainly enriched in two GO terms: DNA binding transcription factor activity (GO:0003700) and transcription regulator activity (GO:0140110). Down-regulated differential genes in nonuploid have mainly been enriched in several different terms, such as photosynthesis (GO:0015979), thylakoid (GO:0009579), thylakoid part (GO:0044436), photosynthetic membrane (GO:0034357), photosystem (GO:0009521), photosystem II (GO:0009523), photosystem II oxygen-evolving complex (GO:0009654), thylakoid membrane (GO:0042651), extrinsic component of membrane (GO:0019898), oxidoreductase complex (GO:1990204), membrane protein complex (GO:0098796), apoplast (GO:0048046).
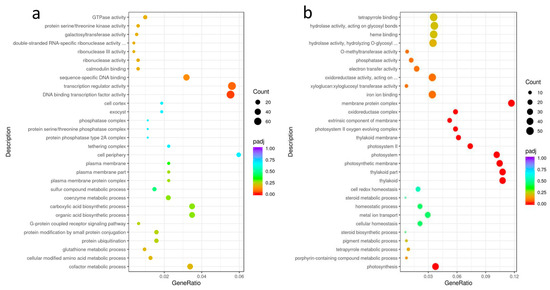
Figure 5.
GO enrichment analysis of DEGs between nonuploid and hexaploid: (a) upregulated DEGs in the nonuploid. (b) downregulated DEGs in the nonuploid.
Furthermore, KEGG enrichment analysis was performed on differentially expressed genes, and the results are shown in Figure 6. Upregulated DEGs in nonuploid were significantly enriched in Plant-pathogen interaction, alpha-linolenic acid metabolism, and Protein processing in plants’ endoplasmic reticulum, Biosynthesis of amino acids, Phenylalanine, tyrosine, and tryptophan biosynthesis, Isoquinoline alkaloid biosynthesis, and Endocytosis. Down-regulated DEGs are mainly enriched in photosynthesis, photosynthesis–antenna proteins, porphyrin metabolism, carbon fixation in photosynthetic organisms, and zeatin biosynthesis.
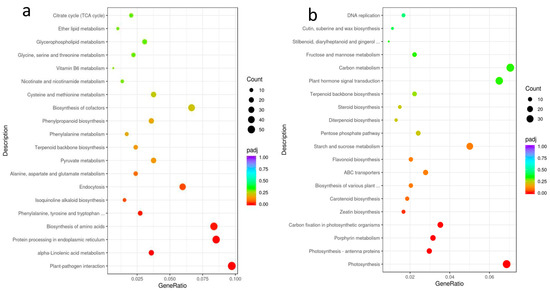
Figure 6.
KEGG enrichment analysis of DEGs between nonuploid and hexaploid: (a) upregulated DEGs in the nonuploid. (b) downregulated DEGs in the nonuploid.
3.3.3. Analysis of DEGs Related to Plant Hormone Biosynthesis
In order to investigate the reasons for the differences in hormone content in the leaves of the nonuploid and hexaploid plants, we focused on analyzing the expression of key genes in the biosynthesis pathways of IAA, GA, and CTK. Auxin biosynthesis pathways include the indole acetaldoxime pathway, indole pyruvate pathway, tryptamine pathway, and indole acetamide pathway. The results showed that in the auxin biosynthesis pathway (Figure 7), the expressions of two monooxygenase genes (YUCCA, evm.TU.contig1398.21 and evm.TU.contig2434.187) of the indole pyruvate pathway in the leaves of the nonuploid were significantly up-regulated, which were 1.77 and 1.79 times higher than those of the hexaploid. The indole pyruvate pathway is the most basic auxin synthesis pathway in plants, and the YUCCA gene is the key enzyme in this pathway. The expression of 2 YUCCA genes in nonuploid was higher than that in hexaploid, which may be related to the significantly higher IAA content in nonuploid than that in hexaploid.
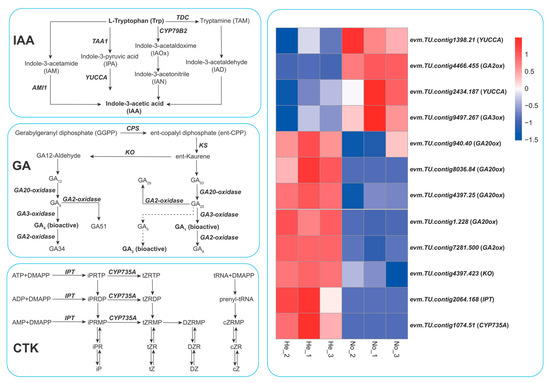
Figure 7.
Analysis of DEGs related to the biosynthesis of auxin, gibberellin, and zeatin.Indole-3-acetic acid (IAA);gibberellin (GA); cytokinin (CTK);Tryptophan aminotransferase of Arabidopsis1 (TAA1); AMIDASE 1 (AMI1); L-tyrosine decarboxylase (TDC); flavin monooxygenase-like proteins (YUCCA); Copalyl pyrophosphate synthase (CPS); Ent-Kaurene Synthase (KS); Ent-kaurene oxidase (KO); isopentenyl transferase (IPT); cytochrome P450 monooxygenase, family 735, subfamily A (CYP735A).
In the gibberellin biosynthesis pathway, GA20ox (evm.TU.contig1.228, evm.TU.contig4397.25, evm.TU.contig8036.84 and evm.TU.contig940.40), KO (evm.TU.contig4397.423) and GA2ox (evm.TU.contig7281.500) genes were found to be downregulated in the nonuploid, while GA2ox (evm.TU.contig4466.455) and GA3ox (evm.TU.contig9497.267) genes were up-regulated. GA3oxis is a key enzyme in the final step of GA synthesis, catalyzing the synthesis of biologically active GA1, GA3, and GA4 using GA9 and GA20 as substrates. The expression level of GA3ox in the nonuploid is 1.44 times higher than that in the hexaploid, which is consistent with the high content of GA1 and GA3 in the nonuploid.
Isopentenyltransferase IPT and cytochrome monooxygenase P450 (CYP735A) are key enzymes in the cytokinin biosynthesis pathway. Transcriptome results showed that IPT (evm.TU.contig2064.168) and CYP735A (evm.TU.contig1074.51) genes were significantly down-regulated in nonuploid, which were 7.37 times and 5.03 times lower than that in hexaploid, respectively. IPT is a rater step synthetase for cytokinin synthesis. The downregulation of the IPT (evm.TU.contig2064.168) gene was consistent with the content of cytokinin in the leaves of the nonuploid and the hexaploid, that is, the content of zeatin iP, tZT, and DZR in the nonuploid was significantly lower than that in the hexaploid.
4. Discussion
Leaf tissue structure can reflect the physiological adaptability of plants to a certain extent, and it can be used as an important basis for germplasm resource evaluation, breeding, and stress resistance research [17]. The leaf characteristics of different ploidy plants were different. The average leaf area and leaf thickness of triploid Eucommia eucommia was 280.36 cm2 and 449.43 μm, 45.8% and 27.8% higher than that of diploid control, respectively. The average leaf size and photosynthetic rate of triploid plants were higher than that of diploid control plants, which was the internal reason for the Eucommia triploid vegetative growth advantage [18]. In this study, the growth of the nonuploid (intra-species triploid) was stronger, the leaf phenotype was thicker than that of the hexaploid leaves, and the tissue section observation also showed obvious changes in leaf veins, and the shape and size of the mesophyric cells was also significantly different. According to leaf and plant phenotype, the nonuploid plant is shorter and stronger, which is more suitable for production application. This result provides a morphological basis for studying the growth characteristics of persimmon polyploids. Plant hormones play an important role in the growth and development of plant organs and the response to environmental stimuli. They regulate the growth, development, and differentiation of plants in terms of cell division and elongation, tissue and organ differentiation, flowering and fruiting, maturation and senescence, dormancy and germination, and in vitro tissue culture, respectively or in coordination [19]. JA is involved in regulating a series of physiological processes related to plant growth and development, such as germination, maturation, aging, and rooting, and is also closely related to the induction of insect resistance in plants as an inducer of plant resistance [20]. In this study, it was found that the JA content in the nonuploid persimmon leaves was significantly higher than that in the hexaploid persimmon. GA is a kind of plant hormone that stimulates cell division and regulates cell wall growth [21]. The contents of GA1 and GA3 in nonuploid persimmon were significantly higher than those in hexaploid persimmon, which may have influenced the growth of nonuploid leaves, making them larger and thicker than hexaploid leaves. ZR, Tzt, and DZR are cytokinins that can promote cell division [22] and lateral shoot growth and stimulate cell differentiation (lateral dominance). The cytokinins in hexaploid persimmons are generally higher than those in nonuploid persimmons, suggesting that hexaploid plants have a stronger ability to develop lateral buds. SA is a new plant endogenous hormone that is involved in regulating physiological processes related to plant growth and development, such as growth, maturation, senescence, etc. It also exhibits strong physiological effects in plant disease resistance, high-temperature resistance, drought resistance, and other adverse stresses [23]. Studies have shown that the use of salicylic acid during the bolting period of narcissus can dwarf the plants and make them compact, while the control group exhibits undesirable characteristics such as elongated plants, loose plant types, and easy lodging [24]. In addition, when treating oriental lilies with salicylic acid solution, it was found that salicylic acid can increase stem diameter and flower bud number [25]. In this study, the content of salicylic acid in the nonuploid persimmon was significantly higher than that in the hexaploid plants, and the plants showed a short and compact shape, which was consistent with the above results. In addition, based on the hormone content, it is speculated that the nonuploid may have higher stress resistance, which requires further verification in the future.
Polyploid plants are produced by chromosome doubling, resulting in a corresponding doubling of the number of alleles at the same locus on the chromosome, which further causes changes in gene expression and subsequently leads to mutations in plant phenotype and physiological state [26]. Transcriptome sequencing is an effective method for studying variations in gene expression levels. According to relevant research reports, compared with diploid cowpeas, tetraploid cowpeas exhibit significant differences in leaves, stem thickness, petals, and pods, and their photosynthetic rate is significantly increased. Transcriptome analysis identified 18 differentially expressed genes related to phenotype and photosynthesis [27]. In this study, through transcriptome sequencing analysis, 5796 differentially expressed genes in the leaves of nonuploid and hexaploid were identified, and the differentially expressed genes were significantly enriched into pathways such as photosynthesis, thylakoids, and photosynthetic system, indicating that there would be significant differences in the intensity of photosynthesis between nonuploid and hexaploid leaves. However, due to limitations in the materials used in this study, the photosynthetic rate has not been measured yet.
Leaf development and growth of plants are jointly regulated by genes, hormones, and the environment. Environmental factors can induce the expression of certain genes related to growth and development, and the balance of intracellular plant hormones is affected by gene expression, while plant hormones can feedback and regulate the expression of other genes, etc., which is a rather complex process [28]. Plant hormones are signal molecules that can produce physiological effects at a small concentration and play an important role in regulating plant growth and development and environmental adaptation. Research has shown that IAA, CTK, and GA promote cell division, stem elongation, and growth, while ET, JA, and SA promote chlorophyll and chloroplast degradation, tissue and organ aging, and response to stress. By comparing the hormone content and gene expression of triploid and diploid poplar leaves, Liao et al. found that the contents of CTK, IAA, and GA were high in triploid leaves, and most of the differentially expressed genes related to signal transduction pathways, such as BR, CTK, IAA, and GA were up-regulated in triploids with vegetative growth advantages [29]. In this study, the IAA content in the leaves of the nonuploid was significantly higher than that of the hexaploid, and the transcriptome found that the expression of two YUCCA genes was significantly increased in the nonuploid, and YUCCA genes are key enzymes in the indole pyruvate pathway. In the gibberellin synthesis pathway, gibberellin GA3-oxidase is the key enzyme in the final step of GA synthesis. GA3ox gene is up-regulated 1.44 times in the leaves of the nonuploid, which is consistent with the result that the contents of GA1 and GA3 in the nonuploid are higher. In addition, isopentenyl transferase IPT and cytochrome monooxygenase P450 CYP735A genes are key enzymes in the cytokinin biosynthesis pathway, and IPT and CYP735A genes are significantly down-regulated in the nonuploid, thus regulating the reduction of IP, tZT, DZR and other hormones in the nonuploid.
5. Conclusions
There were significant differences in growth traits between the nonuploid and hexaploid persimmon. The nonuploid plants were shorter and more compact, and the leaves were larger and thicker. The contents of salicylic acid (SA) and jasmonate JA in the leaves of the nonuploid plants were significantly higher than those of the hexaploid plants, while the contents of zeasin cytokinins were significantly lower than those of the hexaploid plants. The synergistic effects of these hormones affected the plant and leaf phenotypes of the nonuploid plants. The balance of endogenous hormones is affected by gene expression. YUCCA genes, GA3ox genes, and IPT genes related to hormone synthesis are significantly differentially expressed in nonuploid and hexaploid, which may be the key regulatory genes leading to the differences in IAA, GA, and IPA levels in nonuploid and hexaploid. In addition, based on the biological processes involved in hormones and differentially expressed genes, it is speculated that the nonuploid may be superior to the hexaploid in terms of resistance, which requires further study.
Author Contributions
Conceptualization, Y.S. and J.F.; methodology, T.P.; software, Y.W.; validation, T.P., Y.W., W.H., H.L. and P.S.; writing—original draft preparation, T.P. and Y.W.; writing—review and editing, Y.S.; project administration, J.F.; funding acquisition, Y.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key R&D Program of China, grant number 2022YFD2200400 and 2019YFD1001200.
Data Availability Statement
The transcriptome sequencing raw data were deposited in the National Center for Biotechnology Information Sequence Read Archive (NCBI SRA) under the Bioproject ID PRJNA1063545.
Acknowledgments
The processing and analysis of transcriptome data were completed with the help of Novogene.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Han, W.J.; Li, J.R.; Li, H.W.; Liang, Y.Q.; Sun, P.; Fu, J.M. Annual variation of total polyphenol and flavonoid contents in leaves of different species (varieties) of Diospyros. J. China Agric. Univ. 2016, 21, 31–40. [Google Scholar]
- Mei, Y.; Li, S.Z.; Suo, Y.J.; Sun, P.; Han, W.J.; Diao, S.F.; Wang, L.Y.; Li, H.W.; Fu, J.M. Identification of natural 2n pollens in different persimmon germplasms and ascertainment of their induction period. J. China Agric. Univ. 2019, 9, 44–52. [Google Scholar]
- Tong, M.; Kang, Z.X.; Cheng, S.M.; Chen, Y.W. Research Progress in the Genetic Resources of Dispyros kaki. Hubei Agric. Sci. 2008, 08, 960–964. [Google Scholar]
- Yang, X.Y.; Su, H.D.; Zhang, M.Z.; Zhu, G.T.; Cheng, S.F.; Han, F.P.; Huang, S.W. Polyploidization and domestication. Sci. China Life Sci. 2021, 51, 1457–1466. [Google Scholar] [CrossRef]
- Li, Y.; Yang, J.; Song, L.J.; Qi, Q.; Du, K.; Han, Q.; Kang, X.Y. Study of variation in the growth, photosynthesis, and content of secondary metabolites in Eucommia triploids. Trees 2019, 33, 817–826. [Google Scholar] [CrossRef]
- Hu, Y.; Sun, D.; Hu, H.; Zuo, X.; Xia, T.; Xie, J. A comparative study on morphological and fruit quality traits of diploid and polyploid carambola (Averrhoa carambola L.) genotypes. Sci. Hortic. 2021, 277, 109843. [Google Scholar] [CrossRef]
- Lourkisti, R.; Oustric, J.; Quilichini, Y.; Froelicher, Y.; Herbette, S.; Morillon, R.; Berti, L.; Santini, J. Improved response of triploid citrus varieties to water deficit is related to anatomical and cytological properties. Plant Physiol. Biochem. 2021, 162, 762–775. [Google Scholar] [CrossRef] [PubMed]
- Dang, J.B.; Song, Q.; Li, C.; Guo, Q.G.; Liang, G.L. Application Status and Breeding Prospects of Triploid in Horticultural Plant. J. Hortic. 2018, 45, 1813–1830. [Google Scholar]
- Zhang, J.K.; Zeng, P.; Yu, H.; Meng, X.B.; Li, J.Y. De novo domestication of polyploid rice: A novel breeding strategy and future prospects. Chin. Sci. 2021, 51, 1467–1476. [Google Scholar]
- Wu, W.; Liao, T.; Du, K.; Wei, H.; Kang, X. Transcriptome comparison of different ploidy reveals the mechanism of photosynthetic efficiency superiority of triploid poplar. Genomics 2021, 113, 2211–2220. [Google Scholar] [CrossRef]
- Zhao, H. Master of Medicine, Transcriptome Expression Profile Analysis of Triploid Siraitia grosvenorii and Function of 4 Key Enzyme Genes in Glucoside V Biosynthesis. Master’s Thesis, Peking Union Medical College, Beijing, China, 2014. [Google Scholar]
- Gustafson, F.G. Growth Hormone Studies of Some Diploid And Autotetraploid Plants. J. Hered. 1944, 35, 269–272. [Google Scholar] [CrossRef]
- Luo, X.D.; Dai, L.F.; Qian, C.T.; Lou, Q.F.; Chen, J.F. Morphological and Physiological Characteristics in Cucumis Allopolyploids with Different Ploidy Level. Acta Bot. Sin. 2008, 28, 542–546. [Google Scholar]
- Chun, Q.S.; Fa, D.C.; Nian, J.T.; Zhao, L.L.; Wei, M.F.; Xi, L.H. Factors affecting seed set in the crosses between Dendranthema grandiflorum (Ramat.) Kitamura and its wild species. Euphytica 2010, 171, 181–192. [Google Scholar] [CrossRef]
- Sun, J.R.; Du, G.G.; Han, W.J.; Diao, S.F.; Suo, Y.J.; Li, F.D. Endogenous phytohormone profiles in male and female floral buds of the persimmons (Diospyros kaki Thunb.) during development. Sci. Hortic.-Amst. 2017, 218, 213–221. [Google Scholar] [CrossRef]
- Li, H.W.; Sun, P.; Wang, Y.R.; Zhang, Z.R.; Yang, J.; Suo, Y.J.; Han, W.J.; Diao, S.F.; Li, F.D.; Fu, J.M. Allele-aware chromosome-level genome assembly of the autohexaploid Diospyros kaki Thunb. Sci. Data 2023, 10, 270. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.F.; Jing, C.J.; Zhao, X.P.; Wu, X.H. Advances in Application of Plant Leaf Tissue Structure in the Research of Stress Tolerance. Hebei Agric. Sci. 2018, 22, 50–53. [Google Scholar]
- Li, Y. Master of Forestry, Study on Triploid Induction and Variation of Traits in Eucommia ulmoides Oliver. Master’s Thesis, Beijing Forestry University, Beijing, China, 2017. [Google Scholar]
- Perkins-Veazie, P.; Collins, J.K.; Davis, A.R.; Roberts, W. Carotenoid Content of 50 Watermelon Cultivars. J. Agric. Food Chem. 2006, 54, 2593–2597. [Google Scholar] [CrossRef]
- Xu, W.; Yan, S.C. Role of jasmonic acid in plant induced defense. Acta Ecol. Sin. 2005, 25, 2074–2082. [Google Scholar]
- Zhang, G.H.; Zhang, Y.J.; Cong, R.C.; Zhao, Q.; Dong, K.Q.; Gu, R.Z. Advances on Gibberellins Mechanism. Acta Bot. Sin. 2009, 29, 412–419. [Google Scholar]
- Chen, K.L.; Meng, M.; Zhang, P.; Wei, F.R.; Wang, G.L.; Ma, Y.P. Effects of Rainfall Gradients on the Distribution Pattern and Growth of Endogenous Hormones in Different Organs of Robinia pseudoacacia on the Loess Plateau. Soil Water Conserv. Res. 2020, 1, 298–304. [Google Scholar]
- Yin, L.L.; Hou, X.J. The Recent Advances of Salicylic Acid as Signal Molecules of Resistance in Plant. Chin. Agric. Sci. Bull. 2007, 12, 338–342. [Google Scholar]
- Ding, Y.F.; Liu, P. Research progress on stress resistance induced by salicylic acid in plants. Biol. Teach. 2011, 36, 2–4. [Google Scholar]
- Wang, W.Y.; Lin, J.B.; Zou, H.; Wu, L.J.; Huang, W.Q.; Ju, Y.D. Effects of Salicylicacid on the Ideotype and Activities of Antioxidant Enzymes of Narcissus ttazetta L. Var. Chinensis Roem. Chin. Agric. Sci. Bull. 2009, 25, 157–160. [Google Scholar]
- Wang, T.; Chen, M.L.; Liu, L.; Ning, C.L.; Cai, B.H.; Zhang, Z.; Qiao, Y.S. Changes in Genome and Gene Expression During Plant Polyploidization. Acta Bot. 2015, 50, 504–515. [Google Scholar]
- Wang, Q. Master of Forestry, Analysis The Difference of Morphology, Gene Expression Profile and DNA Methylation on Diploid and Tetraploid Cowpea. Master’s Thesis, Jiangxi Agricultural University, Jiangxi, China, 2020. [Google Scholar]
- Huang, H. Recent Progresses from Studies of Leaf Development. Acta Bot. 2003, 20, 416–422. [Google Scholar]
- Liao, T. Master of Forestry, Molecular Mechanism of Vegetative Growth Advantage in Allotriploid Populus spp. (Section Tacamahaca). Master’s Thesis, Beijing Forestry University, Beijing, China, 2017. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).