

Monofilament Shading Nets Improved Water Use Efficiency on High-Temperature Days in Grapevines Subjected to Hyperarid Conditions

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Study and Treatments
2.2. Plant Measurements
2.3. Statistical Analysis
3. Results
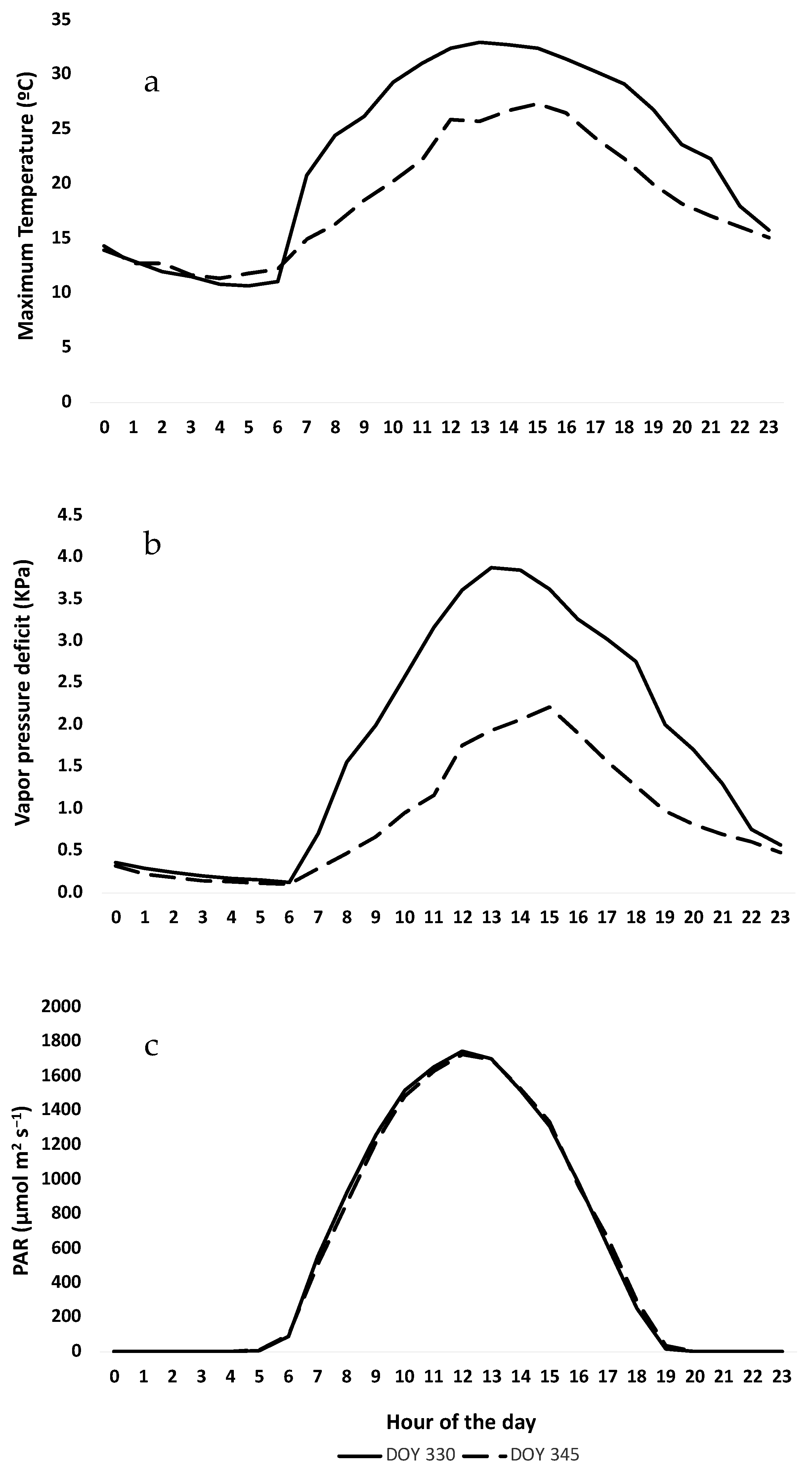
3.1. Climatic Measured Variables
3.2. Effects of Shading Nets and Day of Measurement on Gas Exchange and Water Status of Vines
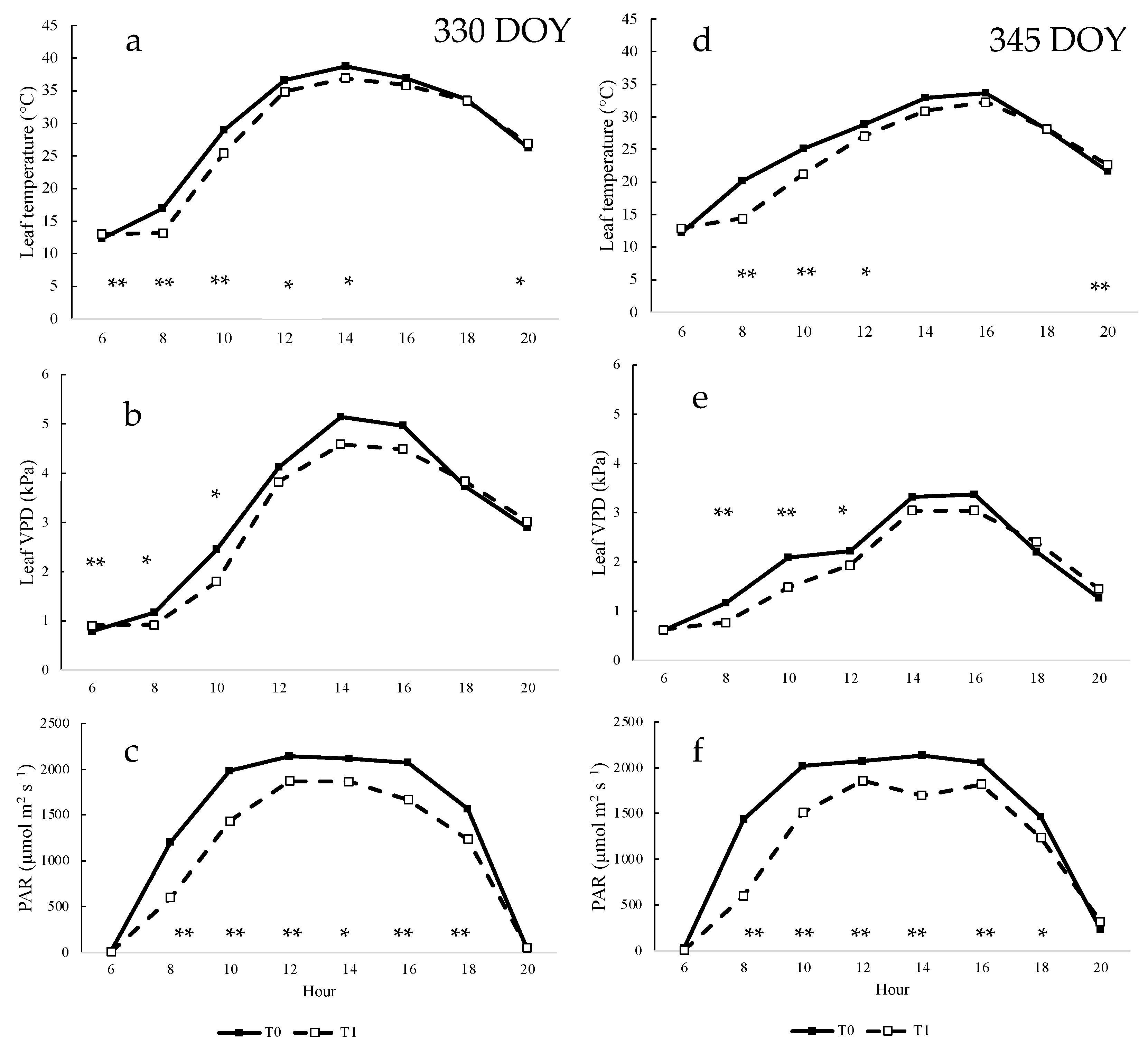
3.3. Effects of Shading Nets on Each Day of Measurement on Gas Exchange and Water Status of Vines
3.4. Daily Measurements of Ecophysiological Responses of Vines
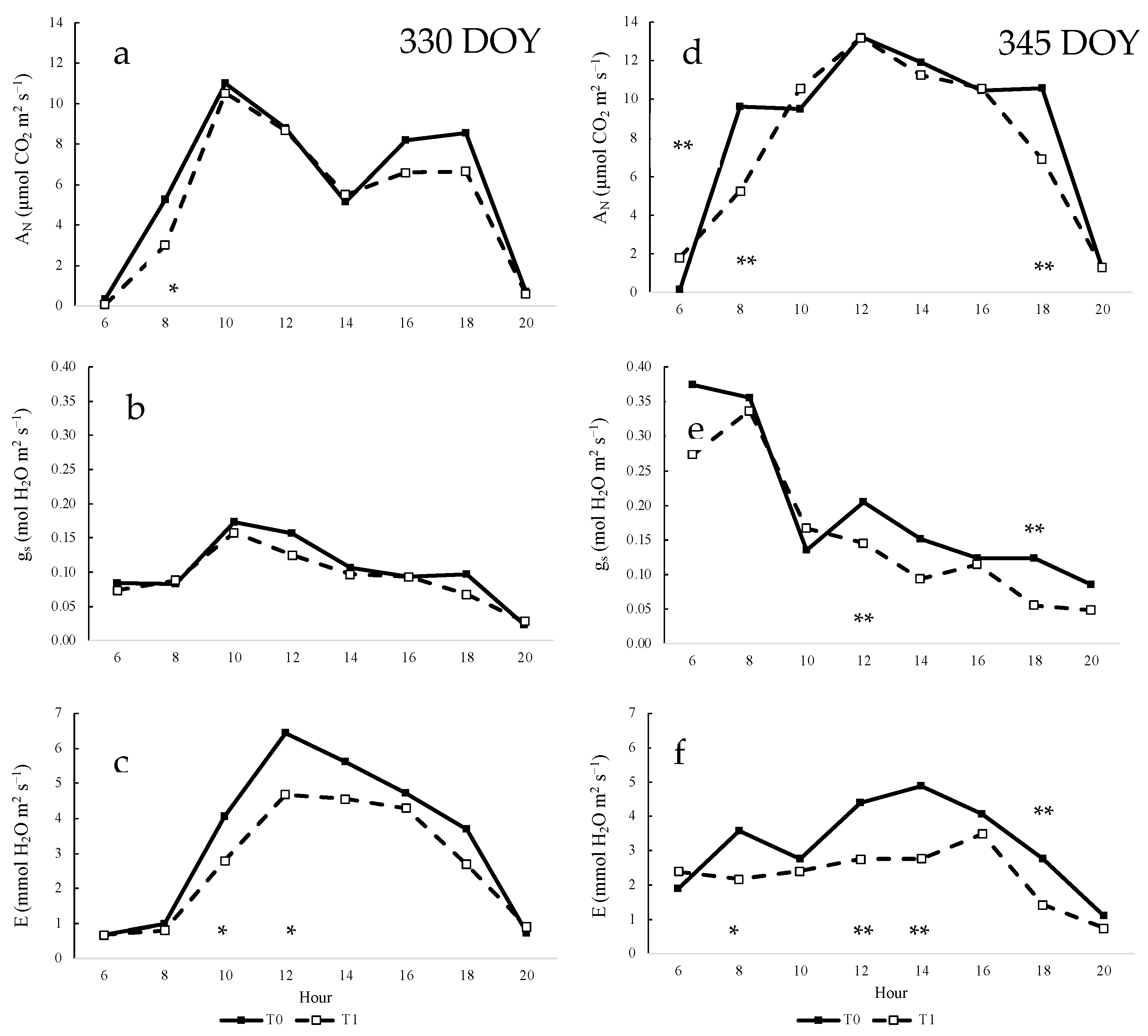
3.5. Daily Measurements of Gas Exchange Parameters of Vines
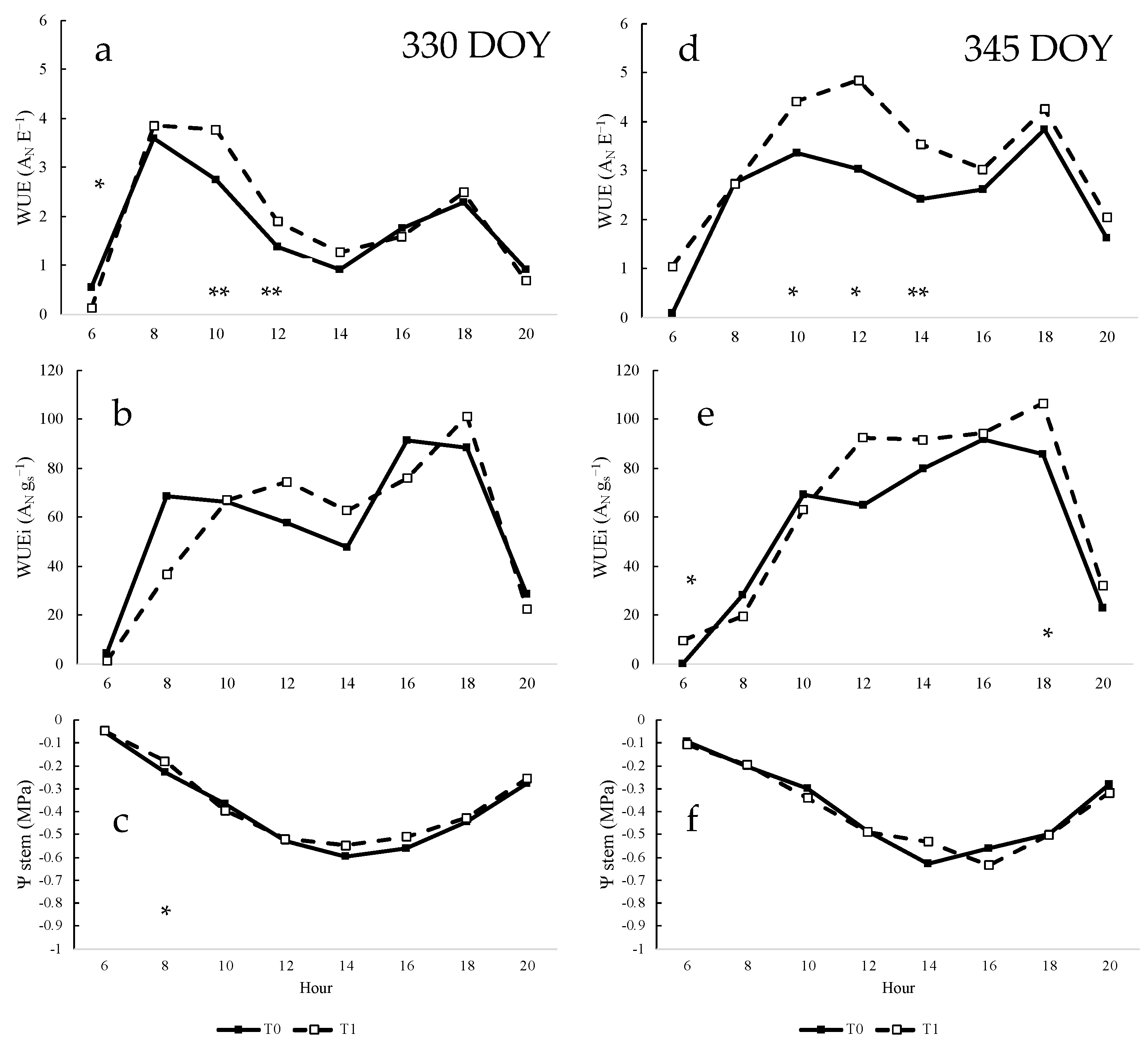
3.6. Daily Measurements of Water Status Parameters of Vines
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zúñiga-Espinoza, C.; Aspillaga, C.; Ferreyra, R.; Selles, G. Response of Table Grape to Irrigation Water in the Aconcagua Valley, Chile. Agronomy 2015, 5, 405–417. [Google Scholar] [CrossRef]
- Uquillas, C.; Torres, E.; Ibacache, A.; Defilippi, B.G. ‘Iniagrape-One’, a New Chilean Table Grape Cultivar. HortScience 2013, 48, 501–503. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Zheng, W.; Martínez de Toda, F. Current Viticultural Techniques to Mitigate the Effects of Global Warming on Grape and Wine Quality: A Comprehensive Review. Food Res. Int. 2021, 139, 109946. [Google Scholar] [CrossRef] [PubMed]
- Verdugo-Vásquez, N.; Orrego, R.; Gutiérrez-Gamboa, G.; Reyes, M.; Zurita-Silva, A.; Balbontín, C.; Gaete, N.; Salazar-Parra, C. Climate Trends and Variability in the Chilean Viticultural Production Zones during 1985–2015. OENO One 2023, 57, 345–362. [Google Scholar] [CrossRef]
- Stolpe, N.; Undurraga, P. Long Term Climatic Trends in Chile and Effects on Soil Moisture and Temperature Regimes. Chil. J. Agric. Res. 2016, 76, 487–496. [Google Scholar] [CrossRef]
- Villalobos-Soublett, E.; Gutiérrez-Gamboa, G.; Balbontín, C.; Zurita-Silva, A.; Ibacache, A.; Verdugo-Vásquez, N. Effect of Shading Nets on Yield, Leaf Biomass and Petiole Nutrients of a Muscat of Alexandria Vineyard Growing under Hyper-Arid Conditions. Horticulturae 2021, 7, 445. [Google Scholar] [CrossRef]
- Venios, X.; Korkas, E.; Nisiotou, A.; Banilas, G. Grapevine Responses to Heat Stress and Global Warming. Plants 2020, 9, 1754. [Google Scholar] [CrossRef]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Response to High Temperature Stress. J. Photochem. Photobiol. B 2014, 137, 116–126. [Google Scholar] [CrossRef]
- Ye, Q.; Yu, J.; Zhang, Z.; Hou, L.; Liu, X. VvBAP1, a Grape C2 Domain Protein, Plays a Positive Regulatory Role Under Heat Stress. Front. Plant Sci. 2020, 11, 544374. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Pérez-Donoso, A.G.; Pou-Mir, A.; Acevedo-Opazo, C.; Valdés-Gómez, H. Hydric Behaviour and Gas Exchange in Different Grapevine Varieties (Vitis Vinifera L.) from the Maule Valley (Chile). South Afr. J. Enol. Vitic. 2019, 40, 181–191. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, B.H.; Hao, Y.; Yang, R.; Wang, Y.A. Effects of Short-Term Heat Stress on PSII and Subsequent Recovery for Senescent Leaves of Vitis Vinifera L. Cv. Red Globe. J. Integr. Agric. 2018, 17, 2683–2693. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Greer, D.H.; Hutton, R.J.; Landsberg, J.J. Does Night-Time Transpiration Contribute to Anisohydric Behaviour in a Vitis Vinifera Cultivar? J. Exp. Bot. 2009, 60, 3763. [Google Scholar] [CrossRef]
- Blanco, V.; Zoffoli, J.P.; Ayala, M. High Tunnel Cultivation of Sweet Cherry (Prunus Avium L.): Physiological and Production Variables. Sci. Hortic. 2019, 251, 108–117. [Google Scholar] [CrossRef]
- Guastella, D.; Sigwebela, M.; Suarez, E.; Stubbs, O.; Acevedo, J.; Engelbrecht, G. Effect of Photo-Selective Shade Nets on Pollination Process and Nut Development of Corylus Avellana L. Front. Plant Sci. 2020, 11, 602766. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Gamboa, G.; Pszczólkowski, P.; Cañón, P.; Taquichiri, M.; Peñarrieta, J.M. UV-B Radiation as a Factor That Deserves Further Research in Bolivian Viticulture: A Review. S. Afr. J. Enol. Vitic. 2021, 42, 201–212. [Google Scholar] [CrossRef]
- Naulleau, A.; Gary, C.; Prévot, L.; Hossard, L. Evaluating Strategies for Adaptation to Climate Change in Grapevine Production—A Systematic Review. Front. Plant Sci. 2021, 11, 607859. [Google Scholar] [CrossRef] [PubMed]
- Lobos, G.A.; Acevedo-Opazo, C.; Guajardo-Moreno, A.; Valdés-Gómez, H.; Taylor, J.A.; Laurie, V.F. Effects of Kaolin-Based Particle Film and Fruit Zone Netting on Cabernet Sauvignon Grapevine Physiology and Fruit Quality. OENO One 2015, 49, 137–144. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Chen, C.C.L.; Brillante, L.; Kurtural, S.K. Partial Solar Radiation Exclusion with Color Shade Nets Reduces the Degradation of Organic Acids and Flavonoids of Grape Berry (Vitis Vinifera L.). J. Agric. Food Chem. 2017, 65, 10693–10702. [Google Scholar] [CrossRef] [PubMed]
- Gowdy, M.; Suter, B.; Pieri, P.; Marguerit, E.; Irvine, A.D.; Gambetta, G.A.; van Leeuwen, C. Variety-Specific Response of Bulk Stomatal Conductance of Grapevine Canopies to Changes in Net Radiation, Atmospheric Demand, and Drought Stress. OENO One 2022, 56, 205–222. [Google Scholar] [CrossRef]
- Kukal, M.S.; Irmak, S. Nocturnal Transpiration in Field Crops: Implications for Temporal Aggregation and Diurnal Weighing of Vapor Pressure Deficit. Agric. Water Manag. 2022, 266, 107578. [Google Scholar] [CrossRef]
- Zufferey, V.; Spring, J.L.; Verdenal, T.; Dienes, A.; Belcher, S.; Lorenzini, F.; Koestel, C.; Rösti, J.; Gindro, K.; Spangenberg, J.; et al. The Influence of Water Stress on Plant Hydraulics, Gas Exchange, Berry Composition and Quality of Pinot Noir Wines in Switzerland. OENO One 2017, 51, 37–57. [Google Scholar] [CrossRef]
- Greer, D.H.; Weston, C. Heat Stress Affects Flowering, Berry Growth, Sugar Accumulation and Photosynthesis of Vitis Vinifera Cv. Semillon Grapevines Grown in a Controlled Environment. Funct. Plant Biol. 2010, 37, 206–214. [Google Scholar] [CrossRef]
- Greer, D.H.; Weedon, M.M. Modelling Photosynthetic Responses to Temperature of Grapevine (Vitis Vinifera Cv. Semillon) Leaves on Vines Grown in a Hot Climate. Plant Cell Environ. 2012, 35, 1050–1064. [Google Scholar] [CrossRef] [PubMed]
- Greer, D.H. Photon Flux Density and Temperature-Dependent Responses of Photosynthesis and Photosystem II Performance of Apple Leaves Grown in Field Conditions. Funct. Plant Biol. 2015, 42, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Verdugo-Vásquez, N.; Gutiérrez-Gamboa, G.; Díaz-Gálvez, I.; Ibacache, A.; Zurita-Silva, A. Modifications Induced by Rootstocks on Yield, Vigor and Nutritional Status on Vitis Vinifera Cv Syrah under Hyper-Arid Conditions in Northern Chile. Agronomy 2021, 11, 979. [Google Scholar] [CrossRef]
- Flexas, J.; Galmés, J.; Gallé, A.; Gulías, J.; Pou, A.; Ribas-Carbo, M.; Tomàs, M.; Medrano, H. Improving Water Use Efficiency in Grapevines: Potential Physiological Targets for Biotechnological Improvement. Aust. J. Grape Wine Res. 2010, 16, 106–121. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Dold, C. Water-Use Efficiency: Advances and Challenges in a Changing Climate. Front. Plant Sci. 2019, 10, 429990. [Google Scholar] [CrossRef] [PubMed]
- Medrano, H.; Tomás, M.; Martorell, S.; Escalona, J.M.; Pou, A.; Fuentes, S.; Flexas, J.; Bota, J. Improving Water Use Efficiency of Vineyards in Semi-Arid Regions. A Review. Agron. Sustain. Dev. 2015, 35, 499–517. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, X.; Chen, L.; Jia, G. Whole-Plant Instantaneous and Short-Term Water-Use Efficiency in Response to Soil Water Content and CO2 Concentration. Plant Soil 2019, 444, 281–298. [Google Scholar] [CrossRef]
- Cartechini, A.; Palliotti, A. Effect of Shading on Vine Morphology and Productivity and Leaf Gas Exchange Characteristics in Grapevines in the Field. Am. J. Enol. Vitic. 1995, 46, 227–234. [Google Scholar] [CrossRef]
- Agarie, S. Physiological Roles of Silicon in Photosynthesis and Dry Matter Production in Rice Plants I. Effects of Silicon and Shading Treatments. Jpn. J. Crop Sci. 1992, 61, 200–206. [Google Scholar] [CrossRef]
- Pallotti, L.; Silvestroni, O.; Dottori, E.; Lattanzi, T.; Lanari, V. Effects of Shading Nets as a Form of Adaptation to Climate Change on Grapes Production: A Review. OENO One 2023, 57, 467–476. [Google Scholar] [CrossRef]
- Medrano, H.; Tomás, M.; Martorell, S.; Flexas, J.; Hernández, E.; Rosselló, J.; Pou, A.; Escalona, J.M.; Bota, J. From Leaf to Whole-Plant Water Use Efficiency (WUE) in Complex Canopies: Limitations of Leaf WUE as a Selection Target. Crop J. 2015, 3, 220–228. [Google Scholar] [CrossRef]
- Fuentes, S.; De Bei, R.; Collins, M.J.; Escalona, J.M.; Medrano, H.; Tyerman, S. Night-Time Responses to Water Supply in Grapevines (Vitis Vinifera L.) under Deficit Irrigation and Partial Root-Zone Drying. Agric. Water Manag. 2014, 138, 1–9. [Google Scholar] [CrossRef]
- Escalona, J.M.; Fuentes, S.; Tomás, M.; Martorell, S.; Flexas, J.; Medrano, H. Responses of Leaf Night Transpiration to Drought Stress in Vitis Vinifera L. Agric. Water Manag. 2013, 118, 50–58. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AN (µmol CO2 m−2 s−1) | gs (mol H2O m−2 s−1) | E (mmol m−2 s−1) | WUEi (AN gs−1) | WUE (AN E−1) | ψStem (MPa) | ψLeaf (MPa) | |
|---|---|---|---|---|---|---|---|
| Day of the year (DOY) | |||||||
| 330 DOY | 5.61 a | 0.097 a | 3.02 a | 55.96 a | 1.99 a | −0.37 a | −0.67 a |
| 345 DOY | 8.13 b | 0.174 b | 2.72 a | 62.23 a | 3.04 b | −0.39 a | −0.60 a |
| Significance | 0.004 | 0.000 | 0.310 | 0.176 | 0.002 | 0.650 | 0.200 |
| Treatment (T) | |||||||
| T0 Control | 7.17 a | 0.148 a | 3.27 b | 56.02 a | 2.24 a | −0.38 a | −0.65 a |
| T1 Shading net | 6.57 a | 0.123 a | 2.47 a | 62.16 a | 2.78 a | −0.38 a | −0.62 a |
| Significance | 0.700 | 0.120 | 0.007 | 0.390 | 0.070 | 0.830 | 0.630 |
| Interaction (DOY x T) | 0.500 | 0.320 | 0.570 | 0.360 | 0.120 | 0.640 | 0.580 |
| Treatment | AN (µmol CO2 m−2 s−1) | gs (mol H2O m−2 s−1) | E (mmol m−2 s−1) | WUEi (AN gs−1) | WUE (AN E−1) | ψStem (MPa) | ψLeaf (MPa) | |
|---|---|---|---|---|---|---|---|---|
| 330 DOY | T0 Control | 6.00 a | 0.10 a | 3.37 a | 56.70 a | 2.02 a | −0.38 a | −0.70 a |
| T1 Shading net | 5.21 a | 0.09 a | 2.67 a | 55.22 a | 1.96 a | −0.36 a | −0.64 a | |
| Significance | 0.52 | 0.36 | 0.17 | 0.88 | 0.77 | 0.63 | 0.51 | |
| 345 DOY | T0 Control | 8.33 a | 0.19 a | 3.18 b | 55.35 a | 2.47 a | −0.38 a | −0.60 a |
| T1 Shading net | 7.92 a | 0.15 a | 2.26 a | 69.10 a | 3.60 b | −0.39 a | −0.60 a | |
| Significance | 0.570 | 0.190 | 0.004 | 0.140 | 0.016 | 0.860 | 0.950 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Gamboa, G.; Villalobos-Soublett, E.; Garrido-Salinas, M.; Verdugo-Vásquez, N. Monofilament Shading Nets Improved Water Use Efficiency on High-Temperature Days in Grapevines Subjected to Hyperarid Conditions. Horticulturae 2024, 10, 176. https://doi.org/10.3390/horticulturae10020176
Gutiérrez-Gamboa G, Villalobos-Soublett E, Garrido-Salinas M, Verdugo-Vásquez N. Monofilament Shading Nets Improved Water Use Efficiency on High-Temperature Days in Grapevines Subjected to Hyperarid Conditions. Horticulturae. 2024; 10(2):176. https://doi.org/10.3390/horticulturae10020176
Chicago/Turabian StyleGutiérrez-Gamboa, Gastón, Emilio Villalobos-Soublett, Marco Garrido-Salinas, and Nicolás Verdugo-Vásquez. 2024. "Monofilament Shading Nets Improved Water Use Efficiency on High-Temperature Days in Grapevines Subjected to Hyperarid Conditions" Horticulturae 10, no. 2: 176. https://doi.org/10.3390/horticulturae10020176
APA StyleGutiérrez-Gamboa, G., Villalobos-Soublett, E., Garrido-Salinas, M., & Verdugo-Vásquez, N. (2024). Monofilament Shading Nets Improved Water Use Efficiency on High-Temperature Days in Grapevines Subjected to Hyperarid Conditions. Horticulturae, 10(2), 176. https://doi.org/10.3390/horticulturae10020176