
Growth, Triterpene Glycosides, and Antioxidant Activities of Centella asiatica L. Urban Grown in a Controlled Environment with Different Nutrient Solution Formulations and LED Light Intensities


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Establishment of Growth Conditions
2.2.1. Experiment I, Effect of Different NSFs
2.2.2. Experiment II, Effect of LED Light Intensities
2.3. Plant Growth Measurements
2.4. Sample Preparation and Extraction
2.5. Determination of Triterpene Glycoside Content
2.6. Determination of Total Phenolic Content (TPC), Total Flavonoid Content (TFC), and Antioxidant Activities
2.7. Statistical Analysis
3. Results
3.1. Effect of NSFs on Growth, Triterpene Glycoside, Antioxidants, and Antioxidant Activity of C. asiatica Grown in a Greenhouse
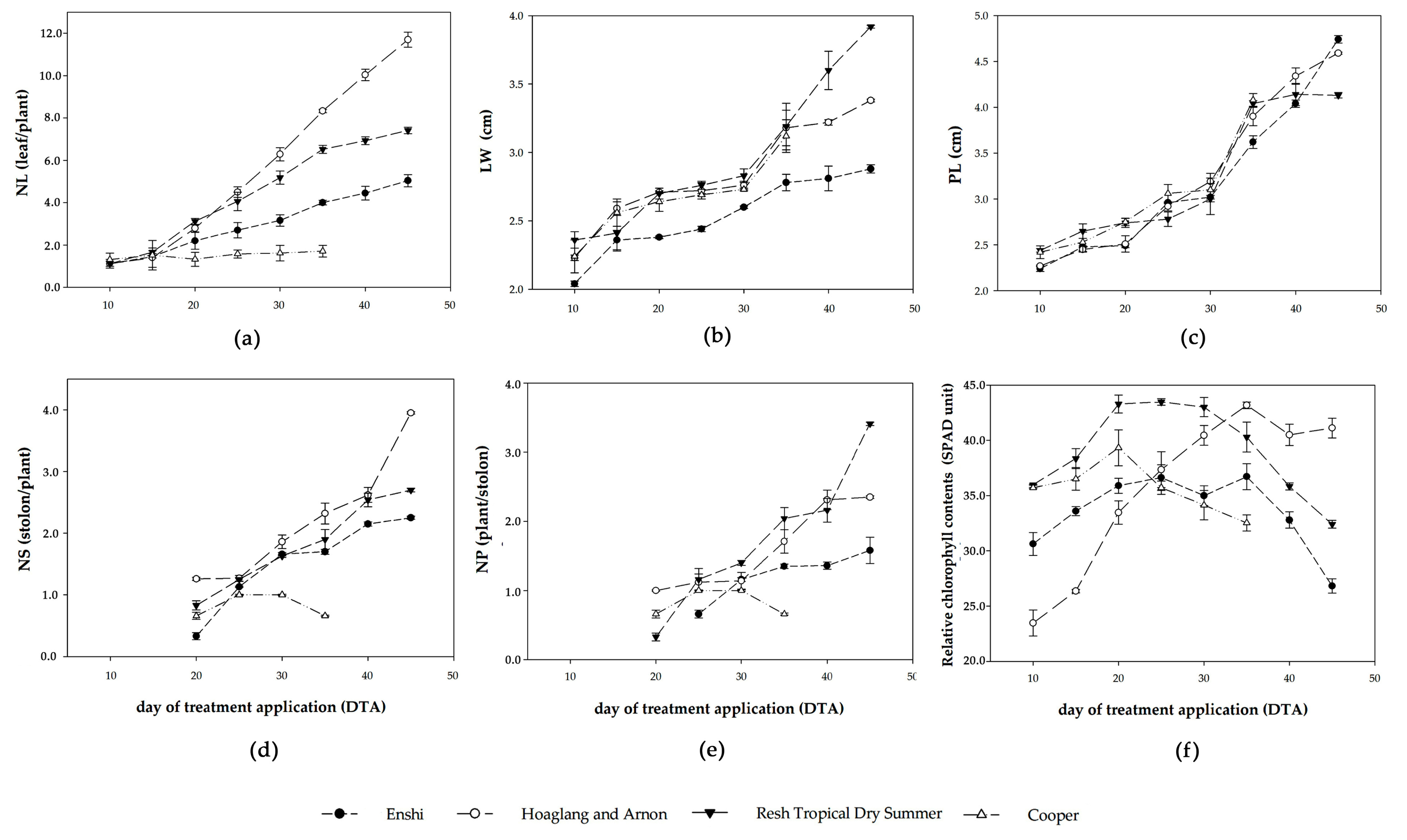
3.1.1. Growth Performance
3.1.2. Triterpene Glycoside, Antioxidants, and Antioxidant Activity
3.2. Effect of LED Light Intensities on Growth, Triterpene Glycoside, Antioxidants, and Antioxidant Activity of C. asiatica Grown in a Controlled Environment
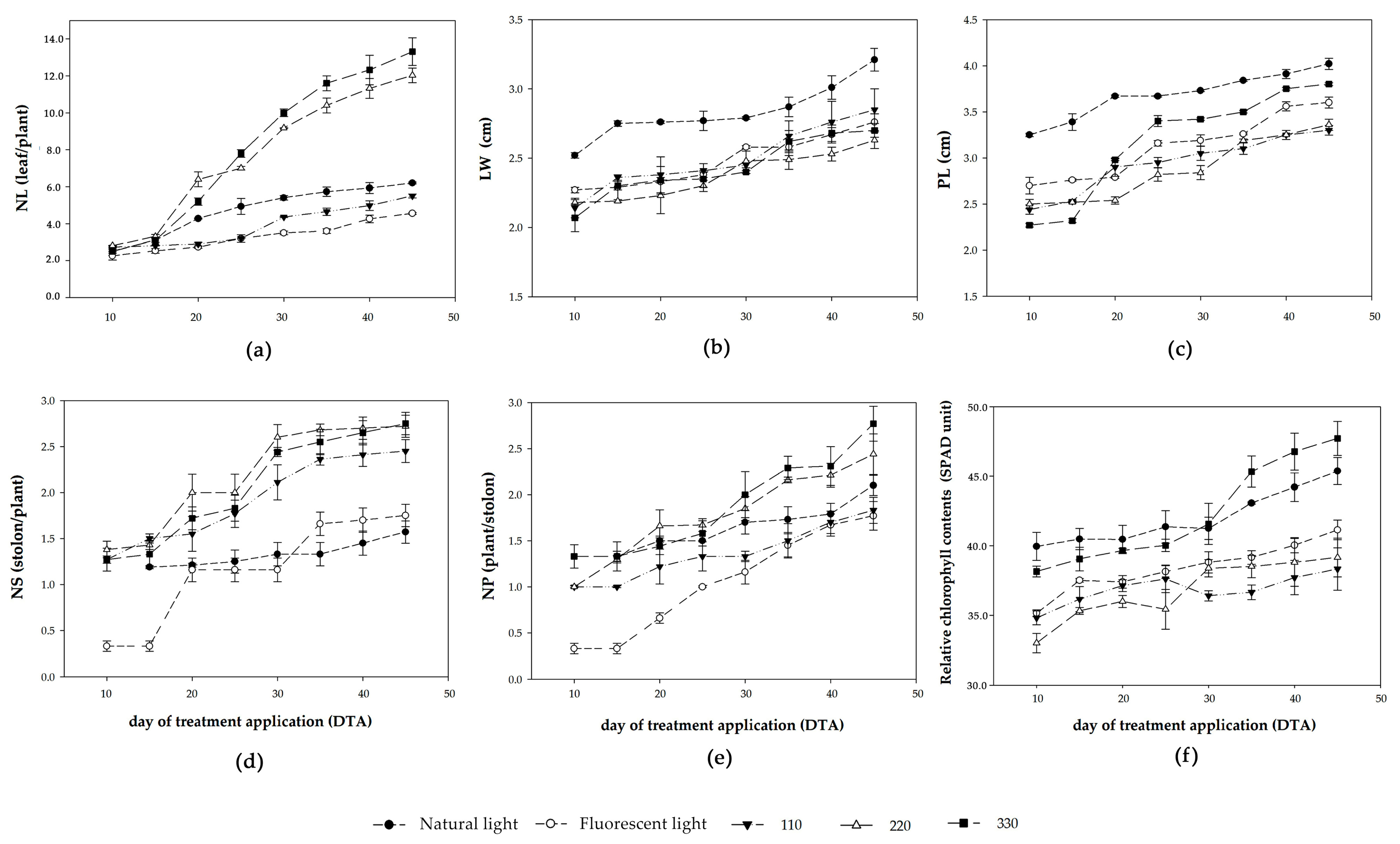
3.2.1. Growth Performance
3.2.2. Triterpene Glycoside, Antioxidants, and Antioxidant Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arshad, M.S.; Khan, U.; Sadiq, A.; Khalid, W.; Hussain, M.; Yasmeen, A.; Asghar, Z.; Rehana, H. Coronavirus disease (COVID-19) and immunity booster green foods: A mini review. Food Sci. Nutr. 2020, 8, 3971–3976. [Google Scholar] [CrossRef] [PubMed]
- Fortune Business Insights. Available online: https://www.fortunebusinessinsights.com/herbal-medicine-market-106320 (accessed on 1 December 2023).
- Gomez, L.A. Growth evaluation and proximate analysis of Gotu Kola (Centella asiatica L.) in response to organic growing media combinations. Asian J. Fundam. Appl. Sci. 2021, 2, 65–76. [Google Scholar]
- Orhan, I.E.; Loizzo, M.R.; Khan, M.T.H. Therapeutic approaches to neuroprotective activity by complementary and alternative medicines. Evid.-Based Complement. Altern. Med. 2012, 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Maulidiani, A.F.; Khatib, A.; Shaari, K.; Lajis, N.H. Chemical characterization and antioxidant activity of three medicinal Apiaceae species. Ind. Crops Prod. 2014, 55, 238–247. [Google Scholar] [CrossRef]
- Sun, B.; Wu, L.; Wu, Y.; Zhang, C.; Qin, L.; Hayashi, M.; Kudo, M.; Gao, M.; Liu, T. Therapeutic potential of Centella asiatica and its triterpenes: A review. Front. Pharmacol. 2020, 11, 568032. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Kharsyntiew, B.; Sharma, P.; Sahoo, U.K.; Sarangi, P.K.; Prus, P.; Imbrea, F. The effect of production and post-harvest processing practices on quality attributes in Centella asiatica (L.) Urban—A review. Agronomy 2023, 13, 1999. [Google Scholar] [CrossRef]
- Hashim, P.; Sidek, H.J.; Helan, M.; Sabery, A.; Palanisamy, U.D.; Ilham, M. Triterpene composition and bioactivities of Centella asiatica. Molecules 2011, 16, 1310–1322. [Google Scholar] [CrossRef]
- Long, H.S.; Stander, M.A.; Van Wyk, B.-E. Notes on the occurrence and significance of triterpenoids (asiaticoside and related compounds) and caffeoylquinic acids in Centella species. S. Afr. J. Bot. 2012, 82, 53–59. [Google Scholar] [CrossRef]
- Prasad, A.; Pragadheesh, V.S.; Mathur, A.; Srivastava, N.; Singh, M.; Mathur, A. Growth and centelloside production in hydroponically established medicinal plant-Centella asiatica (L.). Ind. Crops Prod. 2012, 35, 309–312. [Google Scholar] [CrossRef]
- Müller, V.; Lankes, C.; Zimmermann, B.F.; Noga, G.; Hunsche, M. Centelloside accumulation in leaves of Centella asiatica is determined by resource partitioning between primary and secondary metabolism while influenced by supply levels of either nitrogen, phosphorus or potassium. J. Plant Physiol. 2013, 170, 1165–1175. [Google Scholar] [CrossRef]
- Niu, G.; Masabni, J. Plant production in controlled environments. Horticulturae 2018, 4, 28. [Google Scholar] [CrossRef]
- Folta, K.M. Breeding new varieties for controlled environments. Plant Biol. 2019, 1, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Reza, M.N.; Chowdhury, M.; Chung, S.-O.; Choi, I.-S. A review on effect of ambient environment factors and monitoring technology for plant factory. Precis. Agric. Sci. Technol. 2021, 3, 83–98. [Google Scholar]
- Walters, K.J.; Currey, C.J. Effects of nutrient solution concentration and daily light integral on growth and nutrient concentration of several basil species in hydroponic production. HortScience 2018, 53, 1319–1325. [Google Scholar] [CrossRef]
- De Pascale, S.; Maggio, A.; Orsini, F.; Barbieri, G. Nutrients influence on ready to eat sweet basil quality. Acta Hortic. 2006, 718, 523–530. [Google Scholar] [CrossRef]
- Kilinc, S.S.; Ertan, E.; Seferoğlu, S. Effects of different nutrient solution formulations on morphological and biochemical characteristics of nursery fig trees grown in substrate culture. Sci. Hortic. 2007, 113, 20–27. [Google Scholar] [CrossRef]
- Şirin, U. Effects of different nutrient solution formulations on yield and cut flower quality of gerbera (Gerbera jamesonii) grown in soilless culture system. Afr. J. Agric. Res. 2011, 6, 4910–4919. [Google Scholar]
- Devkota, A.; Dall’Acqua, S.; Comai, S.; Innocenti, G.; Jha, P.K. Centella asiatica (L.) urban from Nepal: Quali-quantitative analysis of samples from several sites, selection of high terpene containing populations for cultivation. Biochem. Syst. Ecol. 2010, 38, 12. [Google Scholar] [CrossRef]
- Shawon, M.R.A.; Azad, M.O.K.; Ryu, B.R.; Na, J.K.; Choi, K.Y. The electrical conductivity of nutrient solution influenced the growth, centellosides content and gene expression of Centella asiatica in a hydroponic system. Agriculture 2023, 13, 2236. [Google Scholar] [CrossRef]
- Fu, Y.; Li, H.Y.; Yu, J.; Liu, H.; Cao, Z.y.; Manukovsky, N.S.; Liu, H. Interaction effects of light intensity and nitrogen concentration on growth, photosynthetic characteristics and quality of lettuce (Lactuca sativa L. Var. youmaicai). Sci. Hortic. 2017, 214, 51–57. [Google Scholar] [CrossRef]
- Gao, W.; He, D.; Ji, F.; Zhang, S.; Zheng, J. Effects of daily light integral and LED spectrum on growth and nutritional quality of hydroponic spinach. Agronomy 2020, 10, 1082. [Google Scholar] [CrossRef]
- Sutulienė, R.; Laužikė, K.; Pukas, T.; Samuolienė, G. Effect of light intensity on the growth and antioxidant activity of sweet basil and lettuce. Plants 2022, 11, 1709. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Yang, S.; Xu, J.; Wang, H.; Zhang, Y.; Cui, J.; Zhang, H.; Jin, H.; Lu, P.; He, L.; et al. Effects of light intensity on growth and quality of lettuce and spinach cultivars in a plant factory. Plants 2023, 12, 3337. [Google Scholar] [CrossRef]
- Veremeichik, G.N.; Grigorchuk, V.P.; Makhazen, D.S.; Subbotin, E.P.; Kholin, A.S.; Subbotina, N.I.; Bulgakov, D.V.; Kulchin, Y.N.; Bulgakov, V.P. High production of flavonols and anthocyanins in Eruca sativa (Mill) Thell plants at high artificial LED light intensities. Food Chem. 2023, 408, 135216. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaityte, A.; Jankauskiene, J.; Virsile, A.; Sirtautaus, R.; Novickovas, A.; Sakalauskaite, J.; Duchovskis, P. LED irradiance level affects growth and nutritional quality of Brassica microgreens. Cent. Eur. J. Biol. 2013, 8, 1241–1249. [Google Scholar] [CrossRef]
- Harakotr, B.; Srijunteuk, S.; Rithichai, P.; Tabunhan, S. Effects of Light-Emitting Diode light irradiance levels on yield, antioxidants and antioxidant capacities of indigenous vegetable microgreens. Sci. Technol. Asia 2019, 24, 59–66. [Google Scholar]
- Jones-Baumgardt, C.; Llewellyn, D.; Zheng, Y. Different microgreen genotypes have unique growth and yield responses to intensity of supplemental PAR from Light-emitting Diodes during winter greenhouse production in Southern Ontario, Canada. HortScience 2020, 55, 156–163. [Google Scholar] [CrossRef]
- Borbély, P.; Gasperl, A.; Pálmai, T.; Ahres, M.; Asghar, M.A.; Galiba, G.; Müller, M.; Kocsy, G. Light intensity- and spectrum-dependent redox regulation of plant metabolism. Antioxidants 2022, 11, 1311. [Google Scholar] [CrossRef]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130243. [Google Scholar] [CrossRef]
- Sritongkul, J.; Srilaong, V.; Uthairatanakij, A.; Kanlayanarat, S.; Chalermglin, P. Effect of light intensity on chemical composition of asiatic pennywort (Centella asiatica L. Urban). Acta Hortic. 2009, 837, 87–93. [Google Scholar] [CrossRef]
- Li, K.; Chen, J.; Wei, Q.; Li, Q.; Lei, N. Effects of transgenerational plasticity on morphological and physiological properties of stoloniferous herb Centella asiatica subjected to high/low light. Front. Plant Sci. 2018, 9, 1640. [Google Scholar] [CrossRef] [PubMed]
- Müller, V.; Albert, A.; Winkler, J.B.; Lankes, C.; Noga, G.; Hunsche, M. Ecologically relevant UV-B dose combined with high PAR intensity distinctly affect plant growth and accumulation of secondary metabolites in leaves of Centella asiatica L. Urban. J. Photochem. Photobiol. B Biol. 2013, 127, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Song, J.W.; Bhandari, S.R.; Shin, Y.K.; Lee, J.G. The influence of red and blue light ratios on growth performance, secondary metabolites, and antioxidant activities of Centella asiatica (L.) Urban. Horticulturae 2022, 8, 601. [Google Scholar] [CrossRef]
- Srithongkul, J.; Kanlayanarat, S.; Srilaong, V.; Uthairatanakij, A.; Chalermglin, P. Effects of light intensity on growth and accumulation of triterpenoids in three accessions of Asiatic pennywort (Centella asiatica (L.) Urb.). J. Food Agric. Environ. 2011, 9, 360–363. [Google Scholar]
- Hori, H. Gravel Culture of Vegetables and Ornamentals; Yokendo: Tokyo, Japan, 1966; pp. 60–79. [Google Scholar]
- Hoagland, D.R.; Arnon, D.L. The water culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 12. [Google Scholar]
- Resh, M.H. Hydroponics for Food Production; Woodbridge Press Publishing Company: Santa Barbara, CA, USA, 1978; 277p. [Google Scholar]
- Cooper, A.J. Crop production in recirculating nutrient solution. Sci. Hortic. 1975, 3, 251–258. [Google Scholar] [CrossRef]
- Tamulaitis, G.; Duchovskis, P.; Bliznikas, Z.; Breive, K.; Ulinskaite, R.; Brazaityte, A.; Novičkovas, A.; Žukauskas, A. High power light-emitting diode based facility for plant cultivation. J. Phys. D Appl. Phys. 2005, 38, 3182–3187. [Google Scholar] [CrossRef]
- Jaiarree, N. Biological Activities of Dioscorea birmanica Prain and Burkill Extract and Its Active Ingredients. Ph.D Thesis, Thammasat University, Pathum Thani, Thailand, 2010. [Google Scholar]
- Hengjumrut, P.; Anukunwithaya, T.; Tantisira, M.H.; Tantisira, B.; Khemawoot, P. Comparative pharmacokinetics between madecassoside and asiaticoside presented in a standardised extract of Centella asiatica, ECa 233 and their respective pure compound given separately in rats. Xenobiotica 2018, 48, 18–27. [Google Scholar] [CrossRef]
- Folin, O.; Ciocalteu, V. On tyrosine and tryptophan determinations in proteins. J. Biol. Chem. 1927, 73, 627–650. [Google Scholar] [CrossRef]
- Kubola, J.; Siriamornpun, S.; Meeso, N. Phytochemicals, vitamin C and sugar content of Thai wild fruits. Food Chem. 2011, 126, 972–981. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Leb. Wiss Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Kolega, S.; Miras-Moreno, B.; Buffagni, V.; Lucini, L.; Valentinuzzi, F.; Maver, M.; Mimmo, T.; Trevisan, M.; Pii, Y.; Cesco, S. Nutraceutical profiles of two hydroponically grown sweet basil cultivars as affected by the composition of the nutrient solution and the inoculation with Azospirillum brasilense. Front. Plant Sci. 2020, 11, 596000. [Google Scholar] [CrossRef] [PubMed]
- Priyanka, S.P.; Sujatha, S.; Smitha, G.R.; Suryanarayana, M.A.; Kalaivanan, D. Biomass accumulation, bioactive compounds and nutrient uptake in Centella asiatica (L.) in relation to organic nutrition in open-field and shade. Ind. Crops Prod. 2022, 176, 114352. [Google Scholar] [CrossRef]
- Jin, D.; Su, X.; Li, Y.; Shi, M.; Yang, B.; Wan, W.; Wen, X.; Yang, S.; Ding, X.; Zou, J. Effect of red and blue light on cucumber seedlings grown in a plant factory. Horticulturae 2023, 9, 124. [Google Scholar] [CrossRef]
- Resh, H.M. Hydroponic Food Production: A Definitive Guidebook for the Advanced Home Gardener and the Commercial Hydroponic Grower, 7th ed.; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Ma, L.; Zhang, J.; Ren, R.; Fan, B.; Hou, L.; Li, J. Effects of different organic nutrient solution formulations and supplementation on tomato fruit quality and aromatic volatiles. Arch. Agron. Soil Sci. 2020, 67, 563–575. [Google Scholar] [CrossRef]
- Prasad, A.; Yadav, K.S.; Yadav, N.P.; Mathur, A.; Sreedhar, R.V.; Lal, R.K.; Mathur, A.K. Biomass and centellosides production in two elite Centella asiatica germplasms from India in response to seasonal variation. Ind. Crops Prod. 2016, 94, 711–720. [Google Scholar] [CrossRef]
- Tan, S.C.; Bhattamisra, S.K.; Chellappan, D.K.; Candasamy, M. Actions and therapeutic potential of madecassoside and other major constituents of Centella asiatica: A review. Appl. Sci. 2021, 11, 8475. [Google Scholar] [CrossRef]
- Puttarak, P.; Panichayupakaranant, P. Factors affecting the content of pentacyclic triterpenes in Centella asiatica raw materials. Pharm. Biol. 2012, 50, 1508–1512. [Google Scholar] [CrossRef]
- Thimmappa, R.; Geisler, K.; Louveau, T.; O’Maille, P.; Osbourn, A. Triterpene biosynthesis in plants. Annu. Rev. Plant Biol. 2014, 65, 225–257. [Google Scholar] [CrossRef]
- Vinolina, N.S.; Sigalingging, R. Growth and secondary metabolites production of Centella asiatica (L.) Urb. cultivated at different phosphate application rates in acid soil. Trends Sci. 2022, 19, 5820. [Google Scholar] [CrossRef]
- Bryant, J.P.; Chapin, F.S.; Klein, D.R. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 1983, 40, 357–368. [Google Scholar] [CrossRef]
- Herms, D.A.; Mattson, W.J. The Dilemma of plants: To grow or defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef]
- Juszczuk, I.M.; Wiktorowska, A.; Malusá, E.; Rychter, A.M. Changes in the concentration of phenolic compounds and exudation induced by phosphate deficiency in bean plants (Phaseolus vulgaris L.). Plant Soil 2004, 267, 41–49. [Google Scholar] [CrossRef]
- Sakamoto, M.; Komatsu, Y.; Suzuki, T. Nutrient deficiency affects the growth and nitrate concentration of hydroponic radish. Horticulturae 2021, 7, 525. [Google Scholar] [CrossRef]
- Fu, W.; Li, P.; Wu, Y. Effects of different light intensities on chlorophyll fluorescence characteristics and yield in lettuce. Sci. Hortic. 2012, 135, 45–51. [Google Scholar] [CrossRef]
- Dong, C.; Fu, Y.; Liu, C.; Liu, H. Low light intensity effects on the growth, photosynthetic characteristics, antioxidant capacity, yield and quality of wheat (Triticum aestivum L.) at different growth stages in BLSS. Adv. Space Res. 2014, 53, 1557–1566. [Google Scholar] [CrossRef]
- Wu, X.; Khan, R.; Gao, H.; Liu, H.; Zhang, J.; Ma, X. Low light alters the photosynthesis process in cigar tobacco via modulation of the chlorophyll content, chlorophyll fluorescence, and gene expression. Agriculture 2021, 11, 755. [Google Scholar] [CrossRef]
- Yamuangmorn, S.; Jumrus, S.; Jamjod, S.; Sringarm, K.; Arjin, C.; Prom-u-thai, C. Responses of purple rice variety to light intensities and soil zinc application on plant growth, yield and bioactive compounds synthesis. J. Cereal Sci. 2022, 106, 103495. [Google Scholar] [CrossRef]
- Karimi, E.; Jaafar, H.Z.; Ghasemzadeh, A.; Ibrahim, M.H. Light intensity effects on production and antioxidant activity of flavonoids and phenolic compounds in leaves, stems and roots of three varieties of Labisia pumila Benth. Aust. J. Crop Sci. 2013, 7, 1016–1023. [Google Scholar]
- Kumari, R.; Singh, S.; Agrawal, S.B. Effects of supplemental ultraviolet-B radiation on growth and physiology of Acorus calamus L. (sweet flag). Acta Biol. Crac. Ser. Bot. 2009, 51, 19–27. [Google Scholar]
- Pennisi, G.; Orsini, F.; Landolfo, M.; Pistillo, A.; Crepaldi, A.; Nicola, S.; Fernández, J.A.; Marcelis, L.F.M.; Gainquinto, G. Optimal photoperiod for indoor cultivation of leafy vegetables and herbs. Eur. J. Hortic. Sci. 2020, 85, 329–338. [Google Scholar] [CrossRef]
- Nawae, W.; Yoocha, T.; Narong, N.; Paemanee, A.; Ketngamkum, Y.; Romyanon, K.; Toojinda, T.; Tangphatsornruang, S.; Pootakham, W. Transcriptome sequencing revealed the influence of blue light on the expression levels of light-stress response genes in Centella asiatica. PLoS ONE 2021, 16, e0260468. [Google Scholar] [CrossRef] [PubMed]
- Taulavuori, K.; Hyöky, V.; Oksanen, J.A.; Taulavuori, E.; Julkunen-Tiitto, R. Species-specific differences in synthesis of flavonoids and phenolic acids under increasing periods of enhanced blue light. Environ. Exp. Bot. 2016, 121, 145–150. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| NSFs | Fresh Weight (g/Plant) | Dry Weight (g/Plant) |
|---|---|---|
| Enshi | 1.02 ± 0.00 c 1/ | 0.12 ± 0.00 b |
| Hoagland and Arnon | 2.10 ± 0.14 a | 0.21 ± 0.00 a |
| Resh Tropical Dry Summer | 1.36 ± 0.25 b | 0.13 ± 0.02 b |
| C.V. (%) | 11.18 | 7.95 |
| NSFs | Triterpene Glycoside (mg/g DW) | TPC | TFC | Antioxidant Activities (mg TE/g DW) | ||
|---|---|---|---|---|---|---|
| Asiaticoside | Madecassoside | (mg GAE/g DW) | (mg QE/g DW) | DPPH | ABTS | |
| Enshi | 4.80 ± 0.01 b 1/ | 11.07 ± 0.80 b | 40.26 ± 1.53 c | 20.07 ± 1.12 c | 39.69 ± 0.27 b | 41.72 ± 0.82 b |
| Hoagland and Arnon | 0.71 ± 0.07 c | 3.70 ± 0.44 c | 52.02 ± 0.81 b | 29.19 ± 0.91 b | 18.57 ± 0.41 c | 32.64 ± 1.24 c |
| Resh Tropical Dry Summer | 5.39 ± 0.37 a | 26.78 ± 0.97 a | 59.17 ± 0.80 a | 32.54 ± 0.64 a | 42.17 ± 1.03 a | 49.09 ± 0.30 a |
| C.V. (%) | 6.02 | 5.57 | 2.19 | 3.34 | 1.98 | 2.13 |
| Light Intensity (µmol/m2/s PPFD) | Fresh Weight (g/Plant) | Dry Weight (g/Plant) |
|---|---|---|
| Natural light | 1.25 ± 0.00 c 1/ | 0.12 ± 0.01 c |
| Fluorescent light | 0.56 ± 0.00 d | 0.04 ± 0.00 d |
| 110 | 1.14 ± 0.01 c | 0.11 ± 0.00 c |
| 220 | 1.54 ± 0.11 b | 0.15 ± 0.00 b |
| 330 | 1.97 ± 0.09 a | 0.22 ± 0.00 a |
| C.V. (%) | 5.46 | 2.73 |
| Light Intensity (µmol/m2/s PPFD) | Triterpene Glycosides (mg/g DW) | TPC | TFC | Antioxidant Activities (mg TE/g DW) | ||
|---|---|---|---|---|---|---|
| Asiaticoside | Madecassoside | (mg GAE/g DW) | (mg QE/g DW) | DPPH | ABTS | |
| Natural light | 3.37 ± 0.22 c 1/ | 27.96 ± 0.23 b | 59.50 ± 0.89 c | 33.44 ± 0.41 c | 41.65 ± 0.96 b | 46.97 ± 0.25 b |
| Fluorescent light | 1.03 ± 0.05 e | 19.51 ± 1.05 d | 34.56 ± 0.22 d | 18.48 ± 0.67 d | 31.87 ± 0.12 d | 31.65 ± 0.52 e |
| 110 | 1.49 ± 0.17 d | 18.51 ± 0.69 c | 34.69 ± 1.11 d | 17.49 ± 0.54 d | 34.79 ± 1.12 c | 38.05 ± 0.84 c |
| 220 | 5.01 ± 0.13 b | 26.93 ± 0.11 b | 60.98 ± 0.77 b | 35.07 ± 1.11 b | 40.81 ± 1.18 b | 36.17 ± 1.00 d |
| 330 | 8.12 ± 0.20 a | 31.94 ± 0.68 a | 65.97 ± 0.21 a | 43.06 ± 0.48 a | 51.37 ± 0.96 a | 54.32 ± 0.61 a |
| C.V. (%) | 4.35 | 2.62 | 1.44 | 2.33 | 2.38 | 1.68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harakotr, B.; Charoensup, L.; Rithichai, P.; Jirakiattikul, Y. Growth, Triterpene Glycosides, and Antioxidant Activities of Centella asiatica L. Urban Grown in a Controlled Environment with Different Nutrient Solution Formulations and LED Light Intensities. Horticulturae 2024, 10, 71. https://doi.org/10.3390/horticulturae10010071
Harakotr B, Charoensup L, Rithichai P, Jirakiattikul Y. Growth, Triterpene Glycosides, and Antioxidant Activities of Centella asiatica L. Urban Grown in a Controlled Environment with Different Nutrient Solution Formulations and LED Light Intensities. Horticulturae. 2024; 10(1):71. https://doi.org/10.3390/horticulturae10010071
Chicago/Turabian StyleHarakotr, Bhornchai, Lalita Charoensup, Panumart Rithichai, and Yaowapha Jirakiattikul. 2024. "Growth, Triterpene Glycosides, and Antioxidant Activities of Centella asiatica L. Urban Grown in a Controlled Environment with Different Nutrient Solution Formulations and LED Light Intensities" Horticulturae 10, no. 1: 71. https://doi.org/10.3390/horticulturae10010071
APA StyleHarakotr, B., Charoensup, L., Rithichai, P., & Jirakiattikul, Y. (2024). Growth, Triterpene Glycosides, and Antioxidant Activities of Centella asiatica L. Urban Grown in a Controlled Environment with Different Nutrient Solution Formulations and LED Light Intensities. Horticulturae, 10(1), 71. https://doi.org/10.3390/horticulturae10010071