

Transcription Factor MdPLT1 Involved Adventitious Root Initiation in Apple Rootstocks


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
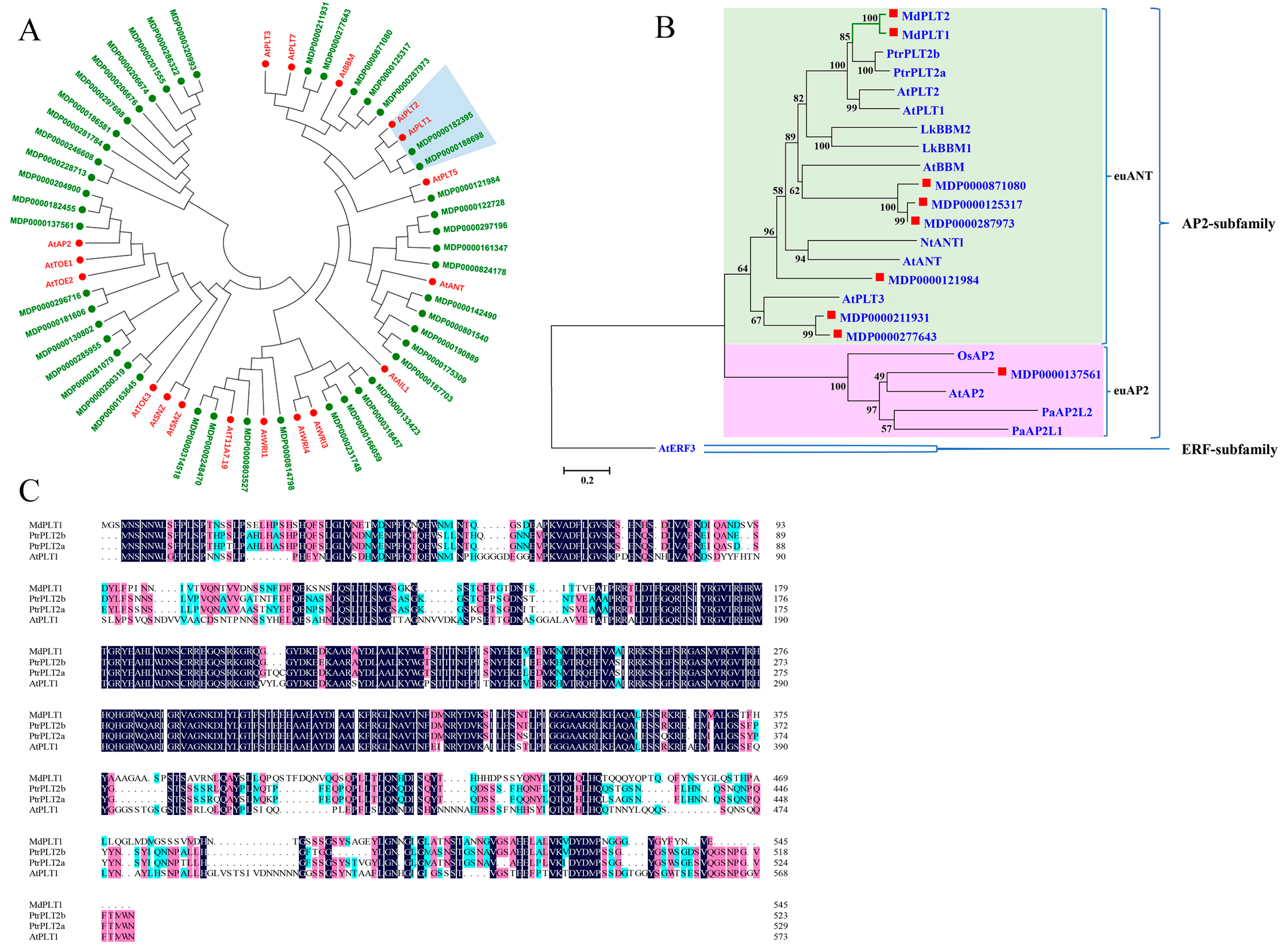
2.2. Construction of Phylogenetic Tree and Multiple Sequence Alignment Analysis
2.3. Extraction of Total RNA and Analysis of MdPLT1 Expression Patterns
2.4. Cloning of the MdPLT1 Gene
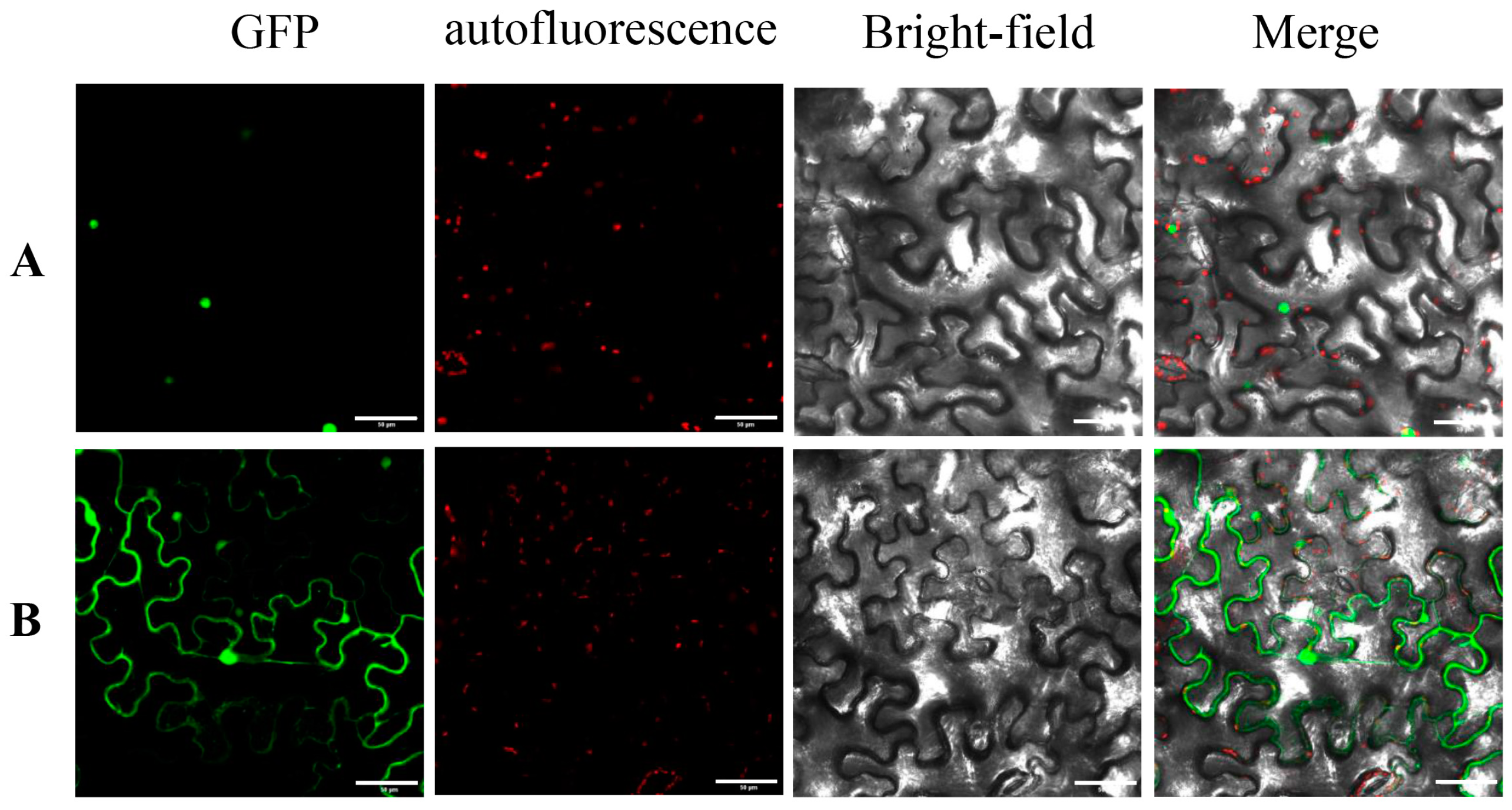
2.5. Construction of MdPLT1-GFP Fusion Expression Vector and Subcellular Localization
2.6. Construction of MdPLT1 Overexpression Vector and Genetic Transformation of Nicotiana benthamiana
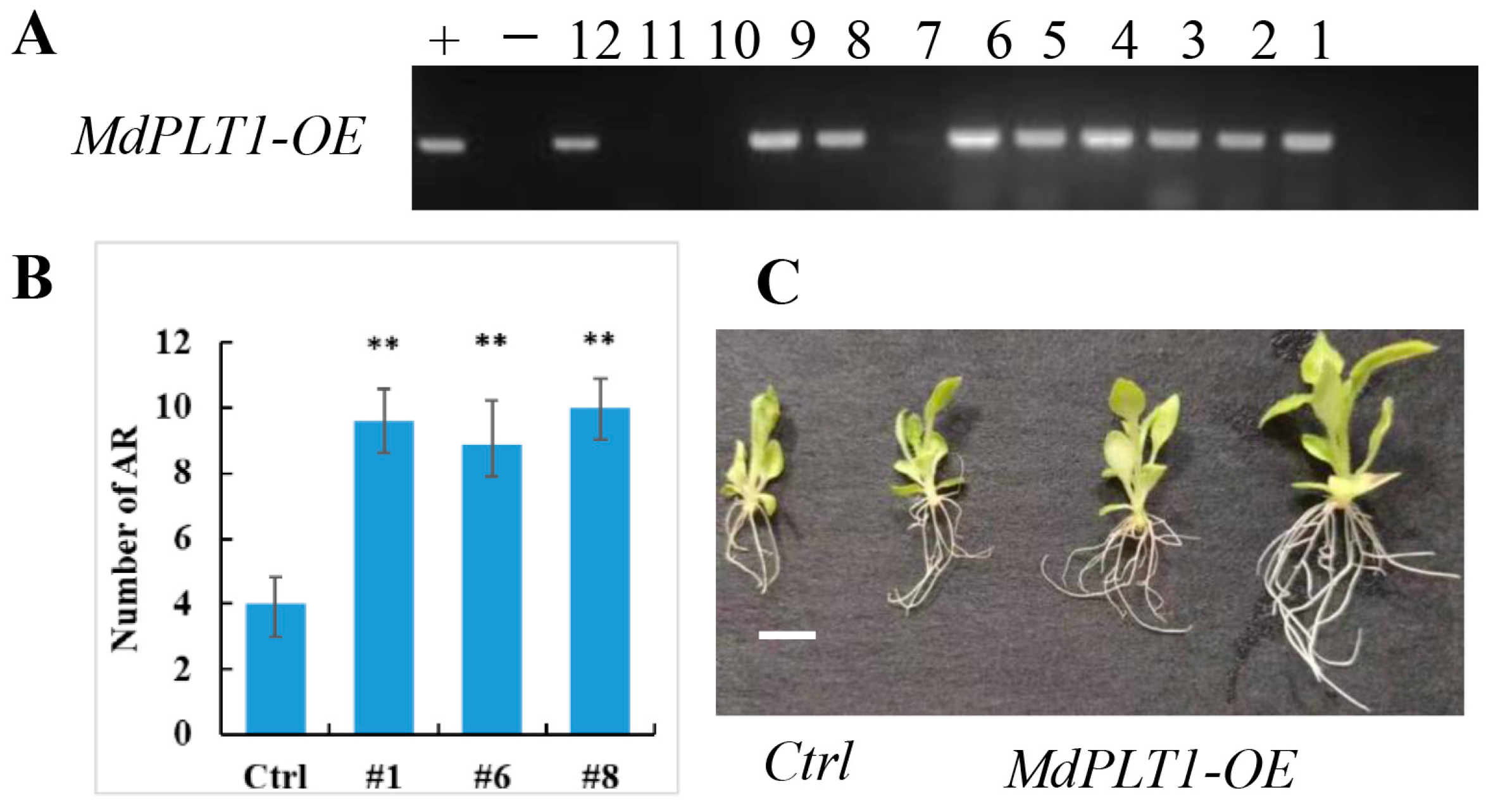
2.7. Identification and Phenotypic Analysis of Transgenic Nicotiana benthamiana with the MdPLT1 Gene
2.8. Relative Expression Analysis of NtPINs and NtYUCCA Gene Families
3. Results
3.1. Identification and Evolutionary Analysis of the Apple PLT1 Gene
3.2. Expression Pattern Analysis of MdPLT1 during Adventitious Root Formation
3.3. Cloning of MdPLT1 Gene
3.4. Subcellular Localization Identification of MdPLT1 Protein
3.5. Tobacco Genetic Transformation and Phenotypic Analysis of MdPLT1
3.6. Relative Expression Analysis of NtPINs and NtYUCCAs Gene Families
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teixeira da Silva, J.A.; Gulyás, A.; Magyar-Tábori, K.; Wang, M.R.; Wang, Q.C.; Dobránszki, J. In vitro tissue culture of apple and other Malus species: Recent advances and applications. Planta 2019, 249, 975–1006. [Google Scholar] [CrossRef]
- Mao, J.; Niu, C.; Li, K.; Chen, S.; Tahir, M.M.; Han, M.; Zhang, D. Melatonin promotes adventitious root formation in apple by promoting the function of MdWOX11. BMC Plant Biol. 2020, 20, 536. [Google Scholar] [CrossRef] [PubMed]
- Naija, S.; Elloumi, N.; Jbir, N.; Ammar, S.; Kevers, C. Anatomical and biochemical changes during adventitious rooting of apple rootstocks MM 106 cultured in vitro. Comptes Rendus Biol. 2008, 331, 518–525. [Google Scholar] [CrossRef]
- Sun, L.; Zhu, Z. The Molecular Basis of Age-Modulated Plant De Novo Root Regeneration Decline in Arabidopsis thaliana. Plant Cell Physiol. 2021, 62, 3–7. [Google Scholar] [CrossRef]
- Rovere, F.D.; Fattorini, L.; Ronzan, M.; Falasca, G.; Altamura, M.M. The quiescent center and the stem cell niche in the adventitious roots of Arabidopsis thaliana. Plant Signal. Behav. 2016, 11, e1176660. [Google Scholar] [CrossRef] [PubMed]
- Singh, Z.; Singh, H.; Garg, T.; Mushahary, K.K.K.; Yadav, S.R. Genetic and Hormonal Blueprint of Shoot-Borne Adventitious Root Development in Rice and Maize. Plant Cell Physiol. 2023, 63, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Sauter, M. Polar Auxin Transport Determines Adventitious Root Emergence and Growth in Rice. Front. Plant Sci. 2019, 10, 444. [Google Scholar] [CrossRef] [PubMed]
- Toscano-Morales, R.; Xoconostle-Cázares, B.; Martínez-Navarro, A.C.; Ruiz-Medrano, R. AtTCTP2 mRNA and protein movement correlates with formation of adventitious roots in tobacco. Plant Signal. Behav. 2016, 11, e1071003. [Google Scholar] [CrossRef]
- Ahkami, A.H. Systems biology of root development in Populus: Review and perspectives. Plant Sci. 2023, 335, 111818. [Google Scholar] [CrossRef]
- Liu, S.; Yang, C.; Wu, L.; Cai, H.; Li, H.; Xu, M. The peu-miR160a-PeARF17.1/PeARF17.2 module participates in the adventitious root development of poplar. Plant Biotechnol. J. 2020, 18, 457–469. [Google Scholar] [CrossRef]
- Wang, H.; Xie, Y.; Liu, W.; Tao, G.; Sun, C.; Sun, X.; Zhang, S. Transcription factor LkWOX4 is involved in adventitious root development in Larix kaempferi. Gene 2020, 758, 144942. [Google Scholar] [CrossRef] [PubMed]
- Tao, G.Y.; Xie, Y.H.; Li, W.F.; Li, K.P.; Sun, C.; Wang, H.M.; Sun, X.M. LkARF7 and LkARF19 overexpression promote adventitious root formation in a heterologous poplar model by positively regulating LkBBM1. Commun. Biol. 2023, 6, 372. [Google Scholar] [CrossRef] [PubMed]
- You, C.X.; Zhao, Q.; Wang, X.F.; Xie, X.B.; Feng, X.M.; Zhao, L.L.; Shu, H.R.; Hao, Y.J. A dsRNA-binding protein MdDRB1 associated with miRNA biogenesis modifies adventitious rooting and tree architecture in apple. Plant Biotechnol. J. 2014, 12, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Smolka, A.; Welander, M.; Olsson, P.; Holefors, A.; Zhu, L.H.J.P.S. Involvement of the ARRO-1 gene in adventitious root formation in apple. Plant Sci. 2009, 177, 710–715. [Google Scholar] [CrossRef]
- Ahkami, A.H.; Lischewski, S.; Haensch, K.T.; Porfirova, S.; Hofmann, J.; Rolletschek, H.; Melzer, M.; Franken, P.; Hause, B.; Druege, U.; et al. Molecular physiology of adventitious root formation in Petunia hybrida cuttings: Involvement of wound response and primary metabolism. New Phytol. 2009, 181, 613–625. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Li, Y.; Huang, K.; Cheng, Z.M. Auxin regulation and MdPIN expression during adventitious root initiation in apple cuttings. Hortic. Res. 2020, 7, 143. [Google Scholar] [CrossRef] [PubMed]
- Bai, T.; Dong, Z.; Zheng, X.; Song, S.; Jiao, J.; Wang, M.; Song, C. Auxin and Its Interaction With Ethylene Control Adventitious Root Formation and Development in Apple Rootstock. Front. Plant Sci. 2020, 11, 574881. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Zhang, D.; Meng, Y.; Li, K.; Wang, H.; Han, M. Inhibition of adventitious root development in apple rootstocks by cytokinin is based on its suppression of adventitious root primordia formation. Physiol. Plant 2019, 166, 663–676. [Google Scholar] [CrossRef]
- Tahir, M.M.; Li, S.; Mao, J.; Liu, Y.; Zhang, D.J.S.H. High nitrate inhibited adventitious roots formation in apple rootstock by altering hormonal contents and miRNAs expression profiles. Sci. Hortic. 2021, 286, 110230. [Google Scholar] [CrossRef]
- Zhang, X.; Tahir, M.M.; Li, S.; Mao, J.; Nawaz, M.A.; Liu, Y.; Li, K.; Xing, L.; Niu, J.; Zhang, D. Transcriptome analysis reveals the inhibitory nature of high nitrate during adventitious roots formation in the apple rootstock. Physiol. Plant 2021, 173, 867–882. [Google Scholar] [CrossRef]
- Li, K.; Liang, Y.; Xing, L.; Mao, J.; Liu, Z.; Dong, F.; Meng, Y.; Han, M.; Zhao, C.; Bao, L.; et al. Transcriptome Analysis Reveals Multiple Hormones, Wounding and Sugar Signaling Pathways Mediate Adventitious Root Formation in Apple Rootstock. Int. J. Mol. Sci. 2018, 19, 2201. [Google Scholar] [CrossRef]
- Li, K.; Wei, Y.H.; Wang, R.H.; Mao, J.P.; Tian, H.Y.; Chen, S.Y.; Li, S.H.; Tahir, M.M.; Zhang, D. Mdm-MIR393b-mediated adventitious root formation by targeted regulation of MdTIR1A expression and weakened sensitivity to auxin in apple rootstock. Plant Sci. 2021, 308, 110909. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Fan, S.; Li, K.; Meng, Y.; Mao, J.; Han, M.; Zhao, C.; Bao, L.; Zhang, D. iTRAQ-Based Proteomic Analysis Reveals Potential Regulation Networks of IBA-Induced Adventitious Root Formation in Apple. Int. J. Mol. Sci. 2018, 19, 667. [Google Scholar] [CrossRef] [PubMed]
- Aida, M.; Beis, D.; Heidstra, R.; Willemsen, V.; Blilou, I.; Galinha, C.; Nussaume, L.; Noh, Y.S.; Amasino, R.; Scheres, B. The PLETHORA genes mediate patterning of the Arabidopsis root stem cell niche. Cell 2004, 119, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Galinha, C.; Hofhuis, H.; Luijten, M.; Willemsen, V.; Blilou, I.; Heidstra, R.; Scheres, B. PLETHORA proteins as dose-dependent master regulators of Arabidopsis root development. Nature 2007, 449, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Ercoli, M.F.; Ferela, A.; Debernardi, J.M.; Perrone, A.P.; Rodriguez, R.E.; Palatnik, J.F. GIF Transcriptional Coregulators Control Root Meristem Homeostasis. Plant Cell 2018, 30, 347–359. [Google Scholar] [CrossRef] [PubMed]
- An, Z.; Liu, Y.; Ou, Y.; Li, J.; Zhang, B.; Sun, D.; Sun, Y.; Tang, W. Regulation of the stability of RGF1 receptor by the ubiquitin-specific proteases UBP12/UBP13 is critical for root meristem maintenance. Proc. Natl. Acad. Sci. USA 2018, 115, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Salvi, E.; Rutten, J.P.; Di Mambro, R.; Polverari, L.; Licursi, V.; Negri, R.; Dello Ioio, R.; Sabatini, S.; Ten Tusscher, K. A Self-Organized PLT/Auxin/ARR-B Network Controls the Dynamics of Root Zonation Development in Arabidopsis thaliana. Dev. Cell 2020, 53, 431–443.e423. [Google Scholar] [CrossRef]
- Liu, G.; Yang, W.; Zhang, X.; Peng, T.; Zou, Y.; Zhang, T.; Wang, H.; Liu, X.; Tao, L.Z. Cystathionine beta-lyase is crucial for embryo patterning and the maintenance of root stem cell niche in Arabidopsis. Plant J. 2019, 99, 536–555. [Google Scholar] [CrossRef]
- Mao, J.; Niu, C.; Li, K.; Fan, L.; Liu, Z.; Li, S.; Ma, D.; Tahir, M.M.; Xing, L.; Zhao, C.; et al. Cytokinin-responsive MdTCP17 interacts with MdWOX11 to repress adventitious root primordium formation in apple rootstocks. Plant Cell 2023, 35, 1202–1221. [Google Scholar] [CrossRef]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, G.Q.; Kang, H.H.; Zhou, S.M.; Wang, W. TaPUB1, a Putative E3 Ligase Gene from Wheat, Enhances Salt Stress Tolerance in Transgenic Nicotiana benthamiana. Plant Cell Physiol. 2017, 58, 1673–1688. [Google Scholar] [CrossRef]
- Hu, X.; Xu, L. Transcription Factors WOX11/12 Directly Activate WOX5/7 to Promote Root Primordia Initiation and Organogenesis. Plant Physiol. 2016, 172, 2363–2373. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Friml, J. Auxin regulates distal stem cell differentiation in Arabidopsis roots. Proc. Natl. Acad. Sci. USA 2010, 107, 12046–12051. [Google Scholar] [CrossRef] [PubMed]
- Krizek, B.A.; Blakley, I.C.; Ho, Y.Y.; Freese, N.; Loraine, A.E. The Arabidopsis transcription factor AINTEGUMENTA orchestrates patterning genes and auxin signaling in the establishment of floral growth and form. Plant J. 2020, 103, 752–768. [Google Scholar] [CrossRef]
- Zhang, H.; Guo, L.; Li, Y.; Zhao, D.; Liu, L.; Chang, W.; Zhang, K.; Zheng, Y.; Hou, J.; Fu, C.; et al. TOP1α fine-tunes TOR-PLT2 to maintain root tip homeostasis in response to sugars. Nat. Plants 2022, 8, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, L.; Bao, L.; Long, T.; Chen, C.; Xu, J. Clonging and expression analysis of two PLT genes promoters in rice. J. Huazhong Agric. Univ. 2012, 31, 147–151. [Google Scholar]
- Rigal, A.; Yordanov, Y.S.; Perrone, I.; Karlberg, A.; Tisserant, E.; Bellini, C.; Busov, V.B.; Martin, F.; Kohler, A.; Bhalerao, R.; et al. The AINTEGUMENTA LIKE1 homeotic transcription factor PtAIL1 controls the formation of adventitious root primordia in poplar. Plant Physiol. 2012, 160, 1996–2006. [Google Scholar] [CrossRef]
- Han, X.L.; Liu, K.; Yuan, G.P.; He, S.S.; Cong, P.H.; Zhang, C.X. Genome-wide identification and characterization of AINTEGUMENTA-LIKE (AIL) family genes in apple (Malus domestica Borkh.). Genomics 2022, 114, 110313–110323. [Google Scholar] [CrossRef]
- Xu, X.Z.; Li, X.; Zhang, X.Z.; Wu, T.; Wang, Y.; Xu, X.F.; Han, Z.H. High miR156 expression is required for auxin-induced adventitious root formation via MxSPL26 independent of PINs and ARFs in Malus xiaojinensis. Front. Plant Sci. 2017, 8, 1059–1076. [Google Scholar] [CrossRef]
- Xiong, F.; Zhang, B.K.; Liu, H.H.; Wei, G.; Wu, J.H.; Wu, Y.N.; Zhang, Y.; Li, S. Transcriptional Regulation of PLETHORA1 in the Root Meristem Through an Importin and Its Two Antagonistic Cargos. Plant Cell 2020, 32, 3812–3824. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Ba, G.; Uwamungu, J.Y.; Ma, W.; Yang, L. Transcription Factor MdPLT1 Involved Adventitious Root Initiation in Apple Rootstocks. Horticulturae 2024, 10, 64. https://doi.org/10.3390/horticulturae10010064
Wang H, Ba G, Uwamungu JY, Ma W, Yang L. Transcription Factor MdPLT1 Involved Adventitious Root Initiation in Apple Rootstocks. Horticulturae. 2024; 10(1):64. https://doi.org/10.3390/horticulturae10010064
Chicago/Turabian StyleWang, Hongming, Guang Ba, Jean Yves Uwamungu, Wenjuan Ma, and Linna Yang. 2024. "Transcription Factor MdPLT1 Involved Adventitious Root Initiation in Apple Rootstocks" Horticulturae 10, no. 1: 64. https://doi.org/10.3390/horticulturae10010064
APA StyleWang, H., Ba, G., Uwamungu, J. Y., Ma, W., & Yang, L. (2024). Transcription Factor MdPLT1 Involved Adventitious Root Initiation in Apple Rootstocks. Horticulturae, 10(1), 64. https://doi.org/10.3390/horticulturae10010064