

Silver Nanoparticles and Biostimulants Affect Chemical Constituents, Total Phenolics, Antioxidants, and Potential Antimicrobial Activities of Santolina chamaecyparissus

 , , ,
, , ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Biostimulants Treatments
2.3. Quantifying Hydrogen Peroxide (H2O2) Levels
2.4. Quantification of Chlorophylls and Carotenoids
2.5. Antibacterial Activity
| Organism | Accession Number |
| Gram-negative bacteria | |
| Escherichia coli | ATCC 25922 |
| Pseudomonas aeruoginosa | ATCC 27853 |
| Gram-positive bacteria | |
| Bacillus subtilis | ATCC 6633 |
| Staphylococcus aureus | ATCC 43300 |
| Bacillus cereus | ATCC 11778 |
2.6. Sample Preparation for Antioxidant Determination
2.7. Determination of Total Phenolic Content
2.8. Determination of Antioxidant Activity
2.9. Reverse Phase High-Performance Liquid Chromatography (RP-HPLC) Analysis for Phenolic Compounds
2.10. Statistical Analysis
3. Results
3.1. Chlorophylls and Carotenoids Content of S. chamaecyparissus Leaves
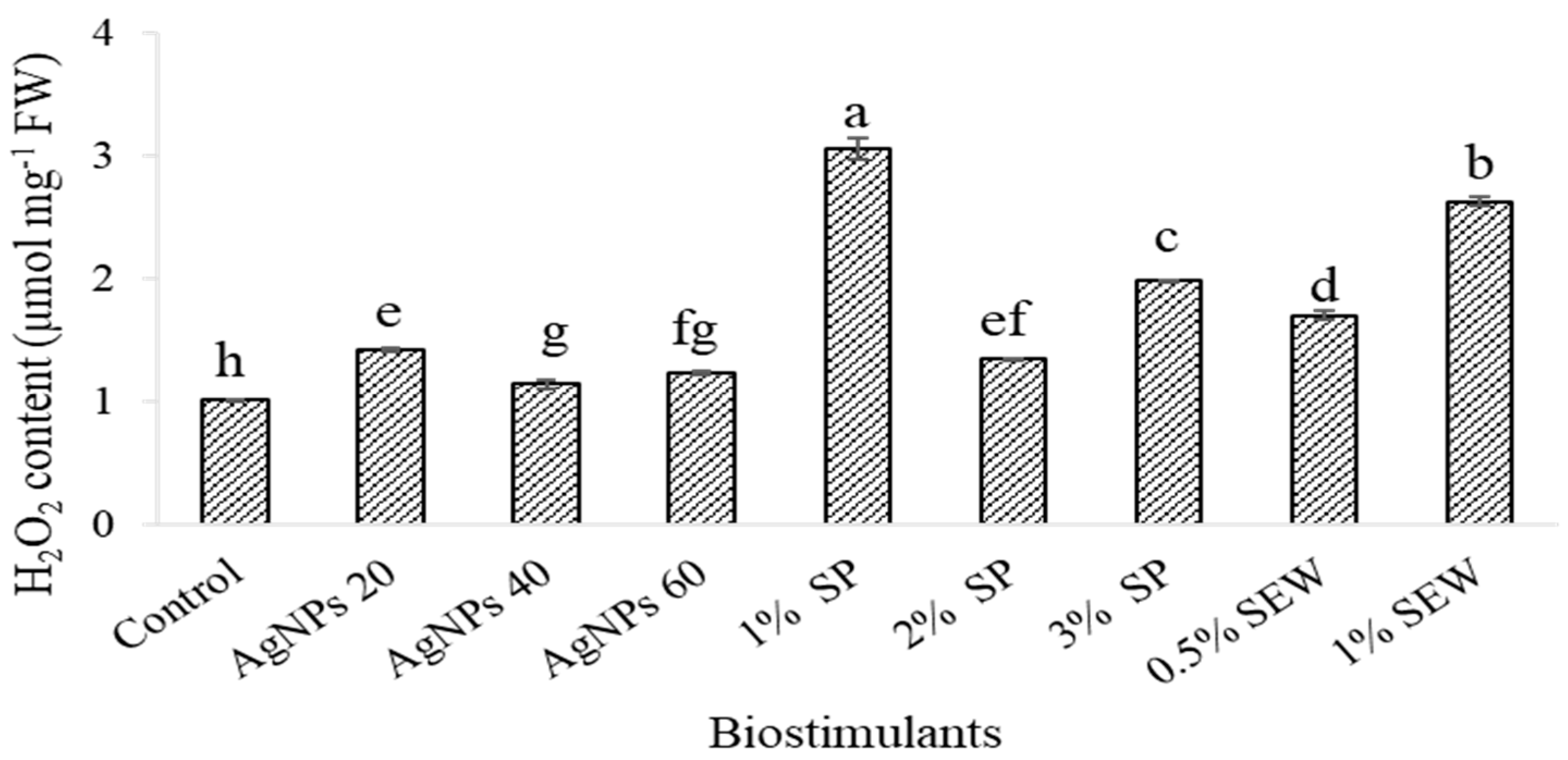
3.2. H2O2 Content in Leaves of S. chamaecyparissus
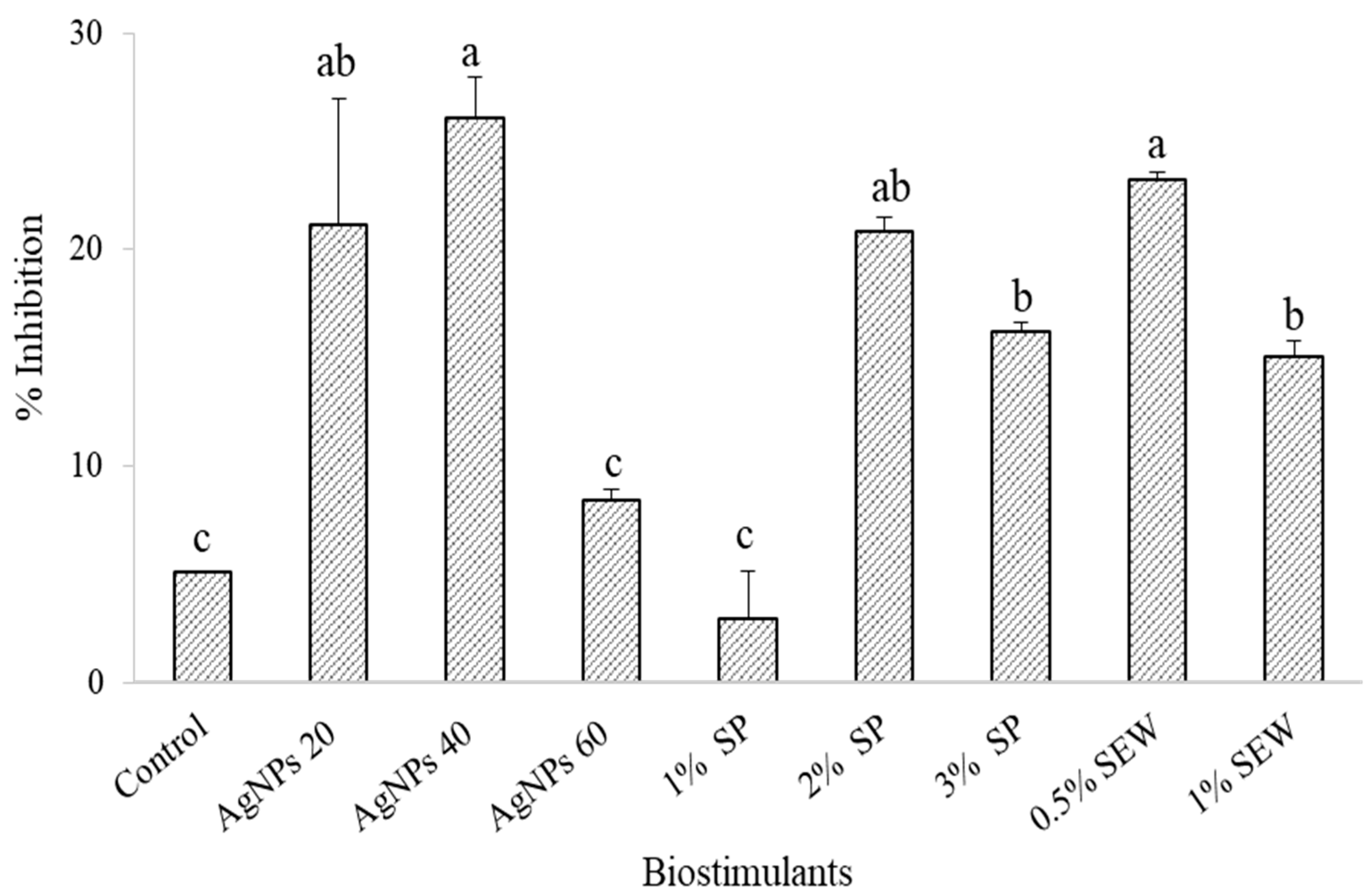
3.3. Chemical Profile and Biological Activity
3.4. Potential Antimicrobial Effects
3.5. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Akerreta, S.; Cavero, R.Y.; López, V.; Calvo, M.I. Analyzing factors that influence the folk use and phytonomy of 18 medicinal plants in Navarra. J. Ethnobiol. Ethnomed. 2007, 3, 16. [Google Scholar] [CrossRef] [PubMed]
- Elsharkawy, E.R. Anticancer effect and seasonal variation in oil constituents of Santolina chamaecyparissu. Chem. Mater. Res. 2014, 6, 85–91. [Google Scholar]
- Ali, A.; Ali, A.; Warsi, M.H.; Ahmad, W.; Tahir, A. Chemical characterization, antidiabetic and anticancer activities of Santolina chamaecyparissus. Saudi J. Biol. Sci. 2021, 28, 4575–4580. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Tahar, D.; Soumia, K.; Dahmane, D.; Mohamed, T.; Lamari, L.; Chabane, C.; Farida, R. Chemical composition, antioxidant and antimicrobial activities of the essential oil of Santolina chamaecyparissus L. of Algeria. J. Coast. Life Med. 2015, 3, 220–227. [Google Scholar]
- Bouriche, H.; Moussaoui, S.; Meziti, H.; Senator, A. Anti-inflammatory activity of methanolic extract of Santolina chamaecyparissus. Acta Hortic. 2013, 1098, 23–30. [Google Scholar] [CrossRef]
- Djeddi, S.; Djebile, K.; Hadjbourega, G.; Achour, Z.; Argyropoulou, C.; Skaltsa, H. In vitro Antimicrobial Properties and Chemical Composition of Santolina chamaecyparissus Essential Oil from Algeria. Nat. Prod. Commun. 2012, 7, 937–940. [Google Scholar] [CrossRef]
- Niu, L.L.; Qin, Q.P.; Wang, L.T.; Gai, Q.Y.; Jiao, J.; Zhao, C.J.; Fu, Y.J. Chemical profiling of volatile components of micropropagated Santolina chamaecyparissus L. Ind. Crops Prod. 2019, 137, 162–170. [Google Scholar] [CrossRef]
- Ahuja, A.; Bakshi, S.K.; Sharma, K.S.; Thappa, R.K.; Agarwal, S.G.; Kichlu, S.K.; Paul, R.; Kaul, M.K. Production of volatile terpenes by proliferating shoots and micropropagated plants of Santolina chamaecyparissus L. (cotton lavender). Flavour Fragr. J. 2005, 20, 403–406. [Google Scholar] [CrossRef]
- Bölek, S.; Tosya, F.; Akçura, S. Effects of Santolina chamaecyparissus essential oil on rheological, thermal and antioxidative properties of dark chocolate. Int. J. Gastron. Food Sci. 2022, 27, 100481. [Google Scholar] [CrossRef]
- de Elguea-Culebras, G.O.; Bourbon, A.I.; Costa, M.J.; Muñoz-Tebar, N.; Carmona, M.; Molina, A.; Sánchez-Vioque, R.; Isabel Berruga, M.; Vicente Vicente, A.A. Optimization of a chitosan solution as potential carrier for the incorporation of Santolina chamaecyparissus L. solid by-product in an edible vegetal coating on ‘Manchego’cheese. Food Hydrocoll. 2019, 89, 272–282. [Google Scholar] [CrossRef]
- Azevedo, T.; Faustino-Rocha, A.I.; Barros, L.; Finimundy, T.C.; Matos, M.; Oliveira, P.A. Santolina chamaecyparissus L.: A Brief Overview of Its Medicinal Properties. Med. Sci. Forum 2023, 21, 8. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, A.H.; Miao, J.H.; Sun, H.; Han, Y.; Yan, G.L.; Wu, F.F.; Wang, X.J. Metabolomics biotechnology, applications, and future trends: A systematic review. RSC Adv. 2019, 9, 37245–37257. [Google Scholar] [CrossRef] [PubMed]
- Hefferon, K.L. Crops With Improved Nutritional Content Though Agricultural Biotechnology. In Plant Micronutrient Use Efficiency-Molecular and Genomic Perspectives in Crop Plants; Hossain, A.M., Kamiya, T., Burritt, D.J., Tran, L.-S.P., Fujiwara, T., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 279–294. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Toward a Sustainable Agriculture Through Plant Biostimulants: From Experimental Data to Practical Applications. Agronomy 2020, 10, 1461. [Google Scholar] [CrossRef]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant Biostimulants: Importance of the Quality and Yield of Horticultural Crops and the Improvement of Plant Tolerance to Abiotic Stress—A Review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Biostimulants in agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Godlewska, K.; Michalak, I.; Pacyga, P.; Baśladyńska, S.; Chojnacka, K. Potential applications of cyanobacteria: Spirulina platensis filtrates and homogenates in agriculture. World J. Microbiol. Biotechnol. 2019, 35, 80. [Google Scholar] [CrossRef]
- Al-Ramamneh, E.A.-D.M.; Ghrair, A.M.; Shakya, A.K.; Alsharafa, K.Y.; Al-Ismail, K.; Al-Qaraleh, S.Y.; Mojski, J.; Naik, R.R. Efficacy of Sterculia diversifolia Leaf Extracts: Volatile Compounds, Antioxidant and Anti-Inflammatory Activity, and Green Synthesis of Potential Antibacterial Silver Nanoparticles. Plants 2022, 11, 2492. [Google Scholar] [CrossRef]
- Salachna, P.; Byczyńska, A.; Zawadzińska, A.; Piechocki, R.; Mizielińska, M. Stimulatory Effect of Silver Nanoparticles on the Growth and Flowering of Potted Oriental Lilies. Agronomy 2019, 9, 610. [Google Scholar] [CrossRef]
- Rabiza-Świder, J.; Skutnik, E.; Jędrzejuk, A.; Rochala-Wojciechowska, J. Nanosilver and sucrose delay the senescence of cut snapdragon flowers. Postharvest Biol. Technol. 2020, 165, 111165. [Google Scholar] [CrossRef]
- Christou, A.; Manganaris, G.A.; Papadopoulos, I.; Fotopoulos, V. Hydrogen sulfide induces systemic tolerance to salinity and non-ionic osmotic stress in strawberry plants through modification of reactive species biosynthesis and transcriptional regulation of multiple defence pathways. J. Exp. Bot. 2013, 64, 1953–1966. [Google Scholar] [CrossRef]
- Porra, R.J. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynth Res. 2002, 73, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Hazra, B.; Mandal, N.; Chaudhuri, T.K. Assessment of the antioxidant and free radical scavenging activities of methanolic extract of Diplazium esculentum. Int. J. Food Prop. 2013, 16, 1351–1370. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L.W.T. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Alli, I.; Ereifej, K.; Alhamad, M.; Al-Tawaha, A.R.; Rababah, T. Optimisation, characterisation and quantification of phenolic compounds in olive cake. Food Chem. 2010, 123, 117–122. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Al-Rabadi, G.J.; Tranchant, C.C.; Almajwal, A.; Alli, I. Occurrence, types, properties and interactions of phenolic compounds with other food constituents in oil-bearing plants. Crit. Rev. Food Sci. Nutr. 2018, 58, 3209–3218. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, A. Brief overview of the application of silver nanoparticles to improve growth of crop plants. IET Nanobiotechnology 2018, 12, 701–705. [Google Scholar] [CrossRef] [PubMed]
- Arahou, F.; Lijassi, I.; Wahby, A.; Rhazi, L.; Arahou, M.; Wahby, I. Spirulina-Based Biostimulants for Sustainable Agriculture: Yield Improvement and Market Trends. BioEnergy Res. 2022, 16, 1401–1416. [Google Scholar] [CrossRef]
- Asadi, M.; Rasouli, F.; Amini, T.; Hassanpouraghdam, M.B.; Souri, S.; Skrovankova, S.; Mlcek, J.; Ercisli, S. Improvement of Photosynthetic Pigment Characteristics, Mineral Content, and Antioxidant Activity of Lettuce (Lactuca sativa L.) by Arbuscular Mycorrhizal Fungus and Seaweed Extract Foliar Application. Agronomy 2022, 12, 1943. [Google Scholar] [CrossRef]
- Deolu-Ajayi, A.O.; van der Meer, I.M.; Van der Werf, A.; Karlova, R. The power of seaweeds as plant biostimulants to boost crop production under abiotic stress. Plant Cell Environ. 2022, 45, 2537–2553. [Google Scholar] [CrossRef]
- Nair, P.M.G.; Chung, I.M. Physiological and molecular level effects of silver nanoparticles exposure in rice (Oryza sativa L.) seedlings. Chemosphere 2014, 112, 105–113. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. Impacts of Silver Nanoparticles on Plants: A Focus on the Phytotoxicity and Underlying Mechanism. Int. J. Mol. Sci. 2019, 20, 1003. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, G.; Vijver, M.G.; Bosker, T.; Peijnenburg, W.J. Foliar versus root exposure of AgNPs to lettuce: Phytotoxicity, antioxidant responses and internal translocation. Environ. Pollut. 2020, 261, 114117. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.N.; Gupta, D.; Mehta, V.K.; Kumar, S. Volatile constituents of the essential oil of Santolina chamaecyparissus Linn. from the southern hills of India. J. Essent. Oil Res. 2001, 13, 234–235. [Google Scholar] [CrossRef]
- Fan, D.; Hodges, D.M.; Critchley, A.T.; Prithiviraj, B. A commercial extract of brown macroalga (Ascophyllum nodosum) affects yield and the nutritional quality of spinach in vitro. Commun. Soil Sci. Plant Anal. 2013, 44, 1873–1884. [Google Scholar] [CrossRef]
- Wafaa, A.E.A.; Hendawy, S.F.; Hamed, E.S.; Toaima, W.I.M. Effect of planting dates, organic fertilization and foliar spray of algae extract on productivity of Dutch fennel plants under Sinai conditions. J. Med. Plants Stud. 2017, 5, 327–334. [Google Scholar]
- Amer, H.M.; Marrez, D.A.; Salama, A.B.; Wahba, H.E.; Khalid, K.A. Growth and chemical constituents of cardoon plant in response to foliar application of various algal extracts. Biocatal. Agric. Biotechnol. 2019, 21, 101336. [Google Scholar] [CrossRef]
- Maswada, H.F.; Mazrou, Y.S.; Elzaawely, A.A.; Eldein, S.M.A. Nanomaterials. Effective tools for field and horticultural crops to cope with drought stress: A review. Span. J. Agric. Res. 2020, 18, 15. [Google Scholar] [CrossRef]
- Soleas, G.J.; Tomlinson, G.; Diamandis, E.P.; Goldberg, D.M. Relative contributions of polyphenolic constituents to the antioxidant status of wines: Development of a predictive model. J. Agric. Food Chem. 1997, 45, 3995–4003. [Google Scholar] [CrossRef]
- Farag, M.A.; Abou Zeid, A.H.; Hamed, M.A.; Kandeel, Z.; El-Rafie, H.M.; El-Akad, R.H. Metabolomic fingerprint classification of Brachychiton acerifolius organs via UPLC-qTOF-PDA-MS analysis and chemometrics. Nat. Prod. Res. 2015, 29, 116–124. [Google Scholar] [CrossRef]
- Irawaty, W.; Ayucitra, A. Assessment on antioxidant and in vitro antidiabetes activities of different fractions of Citrus hystrix peel. Int. Food Res. J. 2018, 25, 2467–2477. [Google Scholar]
- Shrivastava, S.; Bera, T.; Roy, A.; Singh, G.; Ramachandrarao, P.; Dash, D. Characterization of enhanced antibacterial effects of novel silver nanoparticles. Nanotechnology 2007, 18, 225103. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | AgNPs, 20 | AgNPs, 40 | AgNPs, 60 | 1% SP | 2% SP | 3% SP | 0.5% SEW | 1% SEW | |
|---|---|---|---|---|---|---|---|---|---|
| Chl a/Chl b | 3.51 ± 0.28 | 2.56 ± 0.06 | 2.24 ± 0.15 | 1.67 ± 0.05 | 2.25 ± 0.12 | 10.56 ± 1.86 | 3.14 ± 0.09 | 2.06 ± 0.21 | 2.87 ± 0.16 |
| Chl/Caro | 4.52 ± 0.46 | 4.02 ± 0.12 | 4.09 ± 0.13 | 4.66 ± 0.20 | 4.49 ± 0.12 | 3.50 ± 0.19 | 2.71 ± 0.05 | 4.03 ± 0.37 | 4.09 ± 0.08 |
| Standard/Sample | Control | AgNPs, 20 | AgNPs, 40 | AgNPs, 60 | 1% SP | 2% SP | 3% SP | 0.5% SEW | 1% SEW |
|---|---|---|---|---|---|---|---|---|---|
| Gallic acid | 14.06 ± 1.12 | ND | ND | 14.16 ± 0.17 | 10.09 ± 0.01 | 2.74 ± 0.08 | 12.87 ± 0.22 | ND | ND |
| 3,4-Dihydroxyphethyl alcohol | ND | ND | ND | ND | 10.09 ± 0.004 | 35.99 ± 0.46 | 63.25 ± 0.74 | 50.34 ± 1.42 | 46.42 ± 5.54 |
| Catechin | ND | ND | ND | 49.24 ± 0.24 | ND | 19.78 ± 0.48 | 38.16 ± 0.22 | ND | ND |
| 2-Hydroxyphenethyl alcohol | ND | ND | ND | 24.20 ± 0.47 | 14.31 ± 0.62 | 7.44 ± 0.17 | 37.73 ± 1.11 | 18.52 ± 1.60 | 18.11 ± 1.59 |
| Chlorogenic acid | ND | ND | ND | ND | ND | 14.35 ± 0.26 | 29.59 ± 0.42 | ND | ND |
| Vanillic acid | ND | ND | ND | ND | ND | ND | 18.03 ± 0.40 | ND | 5.61 ± 1.64 |
| Epicatechin | ND | ND | ND | 14.89 ± 0.19 | ND | 9.89 ± 0.24 | 36.36 ± 0.68 | ND | ND |
| Caffeic acid | ND | ND | ND | 17.87 ± 0.09 | ND | 23.26 ± 0.21 | 17.14 ± 0.41 | 13.07 ± 2.09 | ND |
| Syringic acid | ND | ND | ND | ND | ND | 1.13 ± 0.07 | 6.74 ± 0.21 | ND | ND |
| P-coumaric acid | ND | ND | ND | 24.42 ± 0.31 | 15.95 ± 1.25 | 15.74 ± 0.14 | 38.83 ± 0.84 | 18.66 ± 0.00 | 18.24 ± 1.64 |
| Sinapic acid | ND | ND | ND | ND | 45.03 ± 0.23 | ND | ND | ND | ND |
| Ferulic acid | ND | ND | ND | 16.27 ± 0.12 | 16.27 ± 0.75 | 3.07 ± 0.09 | 32.11 ± 1.10 | 14.44± 1.53 | 16.19 ± 1.71 |
| Rutin | ND | ND | ND | ND | 22.71 ± 2.05 | 263.22 ± 0.12 | ND | ND | 18.77 ± 0.00 |
| Rosmarinic acid | ND | ND | ND | 65.72 ± 0.38 | 55.82 ± 1.15 | ND | 37.99 ± 0.66 | ND | 57.58 ± 2.06 |
| Quercetin | 3.87 ± 1.13 | ND | ND | 121.92 ± 0.17 | 47.15 ± 0.74 | 2.66 ± 0.09 | 187.26 ± 0.66 | 105.25 ± 1.63 | 49.651 ± 2.06 |
| Thymol | 23.62 ± 0.94 | ND | 13.11 ± 0.11 | 106.74 ± 0.38 | 76.55 ± 0.74 | 2.66 ± 0.10 | 208.65 ± 0.43 | 91.82 ± 2.04 | 85.39 ± 1.64 |
| Conc. (µg/mL) | B. cereus | B. subtilis | S. aureus | E. coli | P. aeruginosa | |
|---|---|---|---|---|---|---|
| Control | 300 | NA | NA | NA | 11.33 ± 0.94 | NA |
| 500 | NA | NA | NA | 19.33 ± 1.25 | NA | |
| AgNPs, 20 | 300 | NA | NA | NA | NA | NA |
| 500 | NA | NA | NA | NA | NA | |
| AgNPs, 40 | 300 | 10.67 ± 1.15 | NA | NA | 20.33 ± 2.05 | NA |
| 500 | 13.0 ± 0.00 | NA | NA | 24.67 ± 1.69 | NA | |
| AgNPs, 60 | 300 | 20.0 ± 2.00 | NA | NA | 22± 0 | NA |
| 500 | 23.33 ± 1.15 | NA | NA | 28.33 ± 1.69 | NA | |
| 1% SP | 300 | 25.66 ± 0.57 | NA | NA | NA | NA |
| 500 | 28.0 ± 0.00 | NA | NA | NA | NA | |
| 2% SP | 300 | 16 ± 2.00 | NA | NA | NA | NA |
| 500 | 20.33 ± 1.15 | NA | NA | NA | NA | |
| 3% SP | 300 | NA | NA | NA | NA | NA |
| 500 | NA | NA | NA | NA | NA | |
| 0.5% SEW | 300 | NA | NA | NA | NA | NA |
| 500 | NA | NA | NA | NA | NA | |
| 1% SEW | 300 | NA | NA | NA | 17.66 ± 0.94 | NA |
| 500 | NA | NA | NA | 21.33 ± 0.94 | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Ramamneh, E.A.-D.; Alsharafa, K.Y.; Rababah, T.; Rahahleh, R.J.; Al-Rimawi, F.; Shakya, A.K.; Ghrair, A.M.; Alu’datt, M.H.; Alnawafleh, M.K. Silver Nanoparticles and Biostimulants Affect Chemical Constituents, Total Phenolics, Antioxidants, and Potential Antimicrobial Activities of Santolina chamaecyparissus. Horticulturae 2024, 10, 26. https://doi.org/10.3390/horticulturae10010026
Al-Ramamneh EA-D, Alsharafa KY, Rababah T, Rahahleh RJ, Al-Rimawi F, Shakya AK, Ghrair AM, Alu’datt MH, Alnawafleh MK. Silver Nanoparticles and Biostimulants Affect Chemical Constituents, Total Phenolics, Antioxidants, and Potential Antimicrobial Activities of Santolina chamaecyparissus. Horticulturae. 2024; 10(1):26. https://doi.org/10.3390/horticulturae10010026
Chicago/Turabian StyleAl-Ramamneh, Ezz Al-Dein, Khalid Y. Alsharafa, Taha Rababah, Razan J. Rahahleh, Fuad Al-Rimawi, Ashok K. Shakya, Ayoup M. Ghrair, Muhammad H. Alu’datt, and Moayad K. Alnawafleh. 2024. "Silver Nanoparticles and Biostimulants Affect Chemical Constituents, Total Phenolics, Antioxidants, and Potential Antimicrobial Activities of Santolina chamaecyparissus" Horticulturae 10, no. 1: 26. https://doi.org/10.3390/horticulturae10010026
APA StyleAl-Ramamneh, E. A.-D., Alsharafa, K. Y., Rababah, T., Rahahleh, R. J., Al-Rimawi, F., Shakya, A. K., Ghrair, A. M., Alu’datt, M. H., & Alnawafleh, M. K. (2024). Silver Nanoparticles and Biostimulants Affect Chemical Constituents, Total Phenolics, Antioxidants, and Potential Antimicrobial Activities of Santolina chamaecyparissus. Horticulturae, 10(1), 26. https://doi.org/10.3390/horticulturae10010026