
Effects of Oligosaccharide Fermentation on Canine Gut Microbiota and Fermentation Metabolites in an In Vitro Fecal Fermentation Model

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Canine Feces Collection and Inoculum Preparation
2.3. In Vitro Fermentation
2.4. pH and Ultraviolet (UV) Measure
2.5. Microbial Analysis
2.6. SCFAs and Lactic Acid Quantitative Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. pH and Total Bacterial Growth
3.2. Gut Microbial Diversity
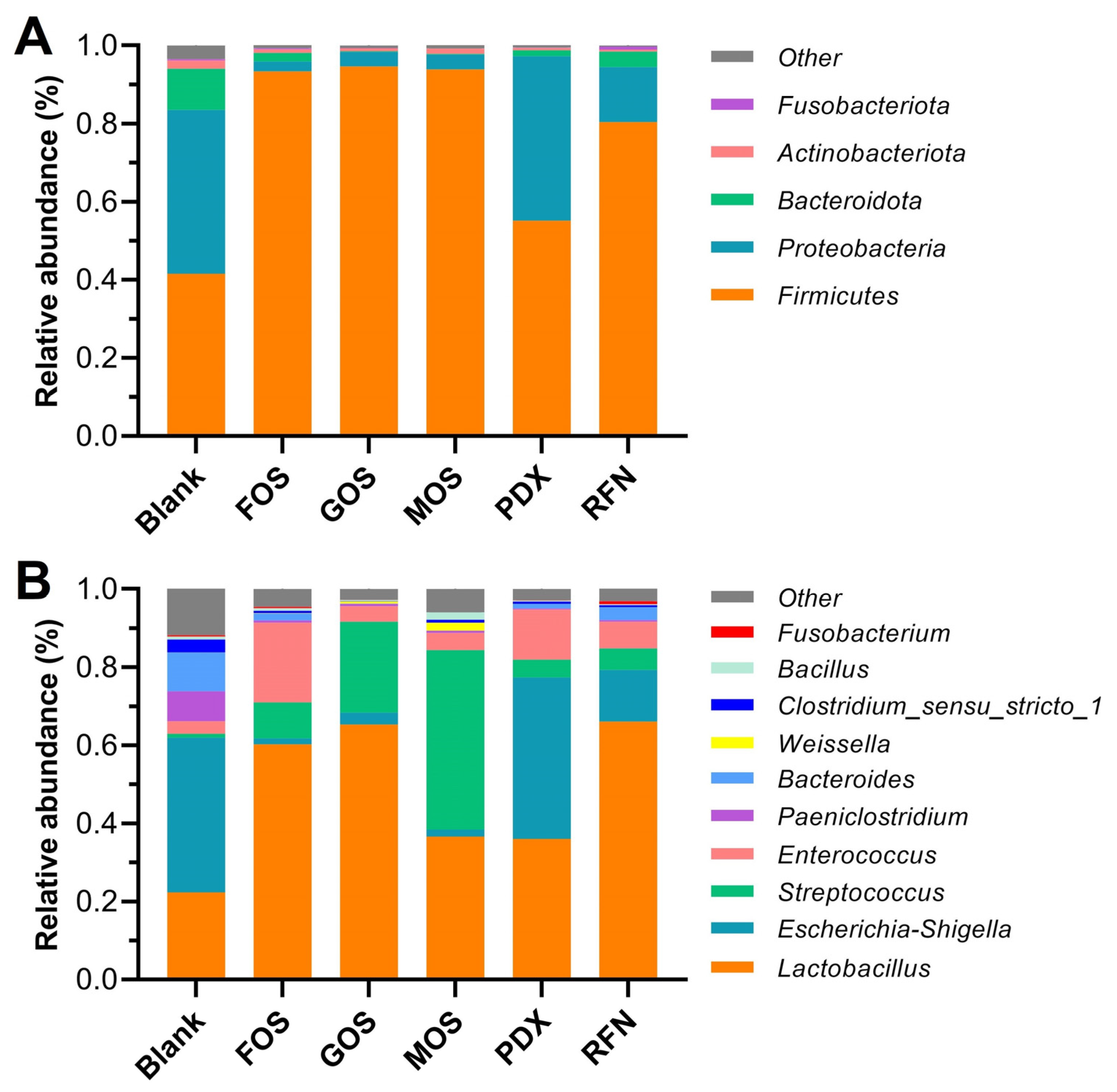
3.3. Microbial Composition
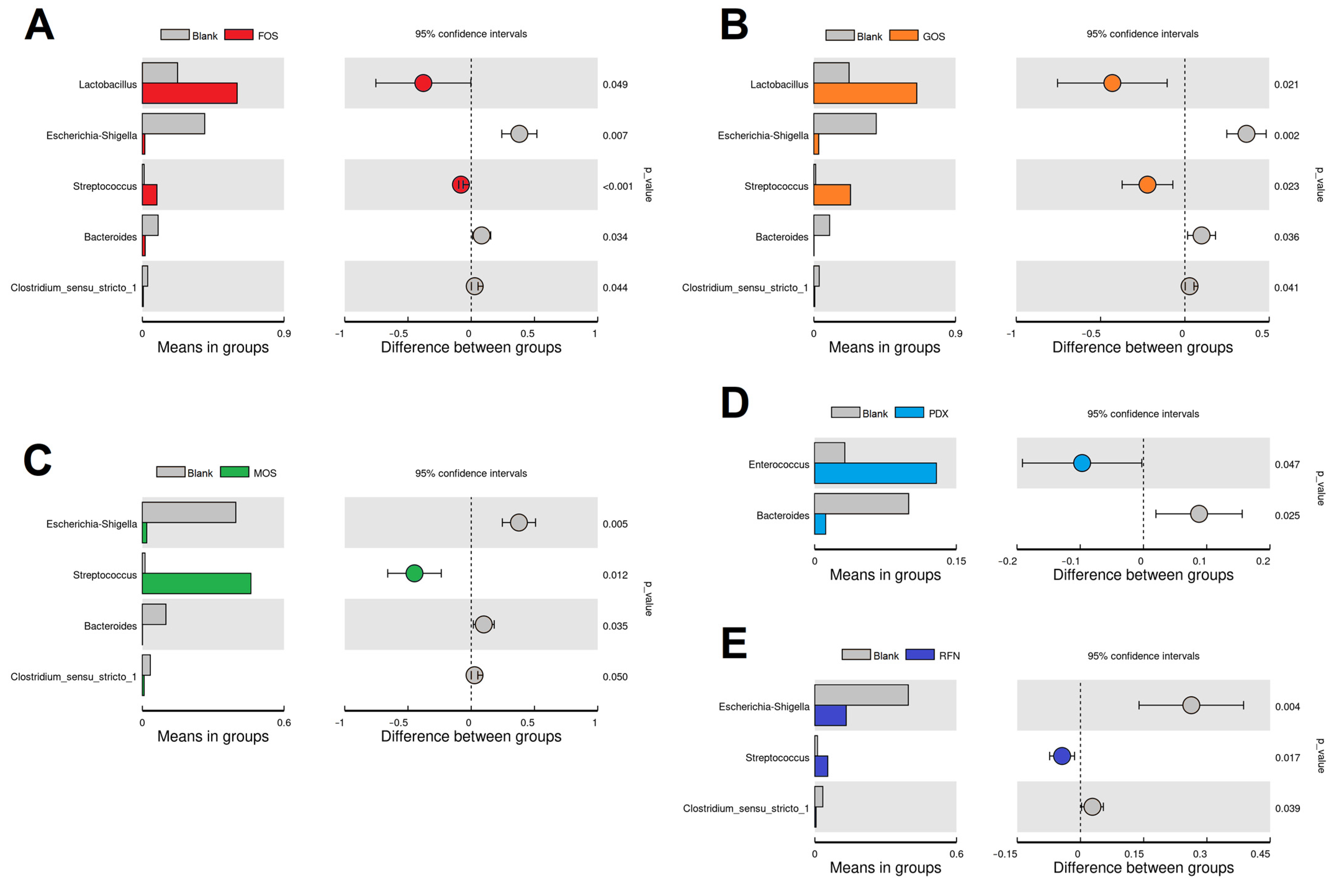
3.4. Linear Discriminant Analysis (LDA)
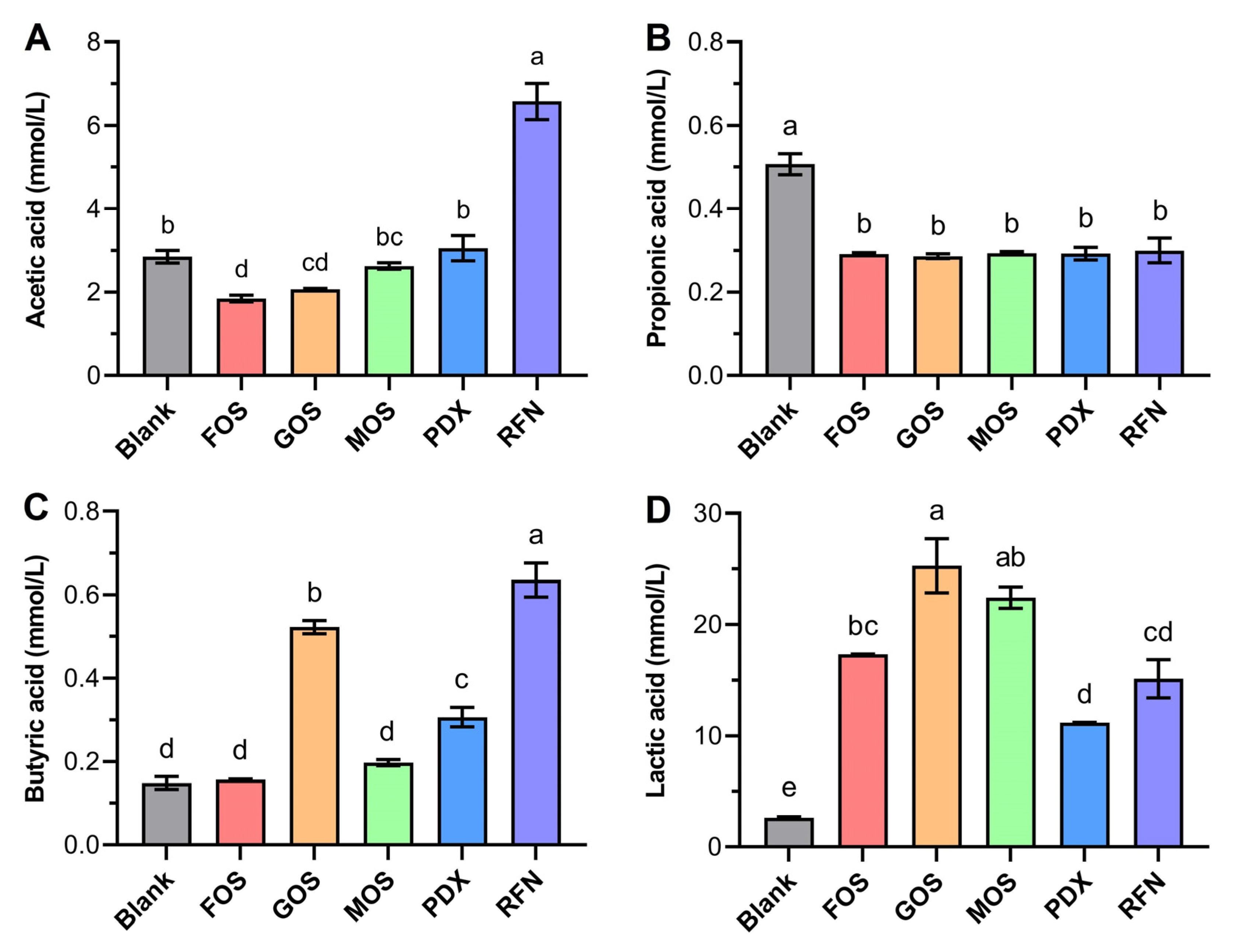
3.5. Influences on SCFA Production
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deschamps, C.; Denis, S.; Humbert, D.; Zentek, J.; Priymenko, N.; Apper, E.; Blanquet-Diot, S. In vitro models of the canine digestive tract as an alternative to in vivo assays: Advances and current challenges. Altex 2022, 39, 235–257. [Google Scholar] [CrossRef]
- Duysburgh, C.; Ossieur, W.P.; De Paepe, K.; Van den Abbeele, P.; Vichez-Vargas, R.; Vital, M.; Pieper, D.H.; Van de Wiele, T.; Hesta, M.; Possemiers, S.; et al. Development and validation of the Simulator of the Canine Intestinal Microbial Ecosystem (SCIME)1. J. Anim. Sci. 2020, 98, skz357. [Google Scholar] [CrossRef]
- Kim, H.S.S.; Aldrich, C.G. 111 Organic Matter Disappearance and Production of Short- and Branched-Chain Fatty Acids from Whole Soybeans and Selected Fiber Sources Used in pet Foods by a Canine in Vitro Fermentation Model. J. Anim. Sci. 2022, 100, 51–52. [Google Scholar] [CrossRef]
- Kim, H.S.; Titgemeyer, E.C.; Aldrich, C.G. Evaluation of Fermentability of Whole Soybeans and Soybean Oligosaccharides by a Canine In Vitro Fermentation Model. Fermentation 2023, 9, 414. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef]
- Grześkowiak, Ł.; Endo, A.; Beasley, S.; Salminen, S.; Microbiota and probiotics in canine and feline welfare. Anaerobe 2015, 34, 14–23. [Google Scholar] [CrossRef]
- Patra, A.K. Responses of feeding prebiotics on nutrient digestibility, faecal microbiota composition and short-chain fatty acid concentrations in dogs: A meta-analysis. Animal 2011, 5, 1743–1750. [Google Scholar] [CrossRef]
- Belà, B.; Coman, M.M.; Verdenelli, M.C.; Bianchi, C.; Pignataro, G.; Fiorini, D.; Silvi, S. In vitro fermentation of Cucumis sativus fructus extract by canine gut microbiota in combination with two probiotic strains. J. Funct. Foods 2019, 63, 103585. [Google Scholar] [CrossRef]
- Tian, J.; Wang, X.; Zhang, X.; Chen, X.; Rui, X.; Zhang, Q.; Dong, M.; Li, W. Simulated digestion and fecal fermentation behaviors of exopolysaccharides from Paecilomyces cicadae TJJ1213 and its effects on human gut microbiota. Int. J. Biol. Macromol. 2021, 188, 833–843. [Google Scholar] [CrossRef]
- Ogué-Bon, E.; Khoo, C.; Hoyles, L.; McCartney, A.L.; Gibson, G.R.; Rastall, R.A. In vitro fermentation of rice bran combined with Lactobacillus acidophilus 14 150B or Bifidobacterium longum 05 by the canine faecal microbiota. FEMS Microbiol. Ecol. 2011, 75, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Ogué-Bon, E.; Khoo, C.; McCartney, A.L.; Gibson, G.R.; Rastall, R.A. In vitro effects of synbiotic fermentation on the canine faecal microbiota. FEMS Microbiol. Ecol. 2010, 73, 587–600. [Google Scholar] [CrossRef][Green Version]
- Ma, G.; Xu, Q.; Du, H.; Muinde Kimatu, B.; Su, A.; Yang, W.; Hu, Q.; Xiao, H. Characterization of polysaccharide from Pleurotus eryngii during simulated gastrointestinal digestion and fermentation. Food Chem. 2022, 370, 131303. [Google Scholar] [CrossRef]
- Koziolek, M.; Grimm, M.; Bollmann, T.; Schäfer, K.J.; Blattner, S.M.; Lotz, R.; Boeck, G.; Weitschies, W. Characterization of the GI transit conditions in Beagle dogs with a telemetric motility capsule. Eur. J. Pharm. Biopharm. 2019, 136, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Vierbaum, L.; Eisenhauer, L.; Vahjen, W.; Zentek, J. In vitro evaluation of the effects of Yucca schidigera and inulin on the fermentation potential of the faecal microbiota of dogs fed diets with low or high protein concentrations. Arch. Anim. Nutr. 2019, 73, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J. Fiber and Prebiotics: Mechanisms and Health Benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cui, X.; Duan, M.; Ai, C.; Song, S.; Chen, X. In vitro fermentation of κ-carrageenan oligosaccharides by human gut microbiota and its inflammatory effect on HT29 cells. J. Funct. Foods 2019, 59, 80–91. [Google Scholar] [CrossRef]
- Wu, D.-T.; Nie, X.-R.; Gan, R.-Y.; Guo, H.; Fu, Y.; Yuan, Q.; Zhang, Q.; Qin, W. In vitro digestion and fecal fermentation behaviors of a pectic polysaccharide from okra (Abelmoschus esculentus) and its impacts on human gut microbiota. Food Hydrocoll. 2021, 114, 106577. [Google Scholar] [CrossRef]
- Li, X.; Guo, R.; Wu, X.; Liu, X.; Ai, L.; Sheng, Y.; Song, Z.; Wu, Y. Dynamic digestion of tamarind seed polysaccharide: Indigestibility in gastrointestinal simulations and gut microbiota changes in vitro. Carbohydr. Polym. 2020, 239, 116194. [Google Scholar] [CrossRef]
- Yao, H.; Wang, L.; Tang, X.; Yang, Z.; Li, H.; Sun, C.; Wu, X.; Xu, D. Two novel polysaccharides from Solanum nigrum L. exert potential prebiotic effects in an in vitro fermentation model. Int. J. Biol. Macromol. 2020, 159, 648–658. [Google Scholar] [CrossRef]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef]
- Vázquez-Baeza, Y.; Hyde, E.R.; Suchodolski, J.S.; Knight, R. Dog and human inflammatory bowel disease rely on overlapping yet distinct dysbiosis networks. Nat. Microbiol. 2016, 1, 16177. [Google Scholar] [CrossRef]
- Pilla, R.; Suchodolski, J.S. The Role of the Canine Gut Microbiome and Metabolome in Health and Gastrointestinal Disease. Front. Vet. Sci. 2019, 6, 498. [Google Scholar] [CrossRef]
- Salas-Mani, A.; Jeusette, I.; Castillo, I.; Manuelian, C.L.; Lionnet, C.; Iraculis, N.; Sanchez, N.; Fernández, S.; Vilaseca, L.; Torre, C. Fecal microbiota composition changes after a BW loss diet in Beagle dogs. J. Anim. Sci. 2018, 96, 3102–3111. [Google Scholar] [CrossRef]
- Bermudez Sanchez, S.; Pilla, R.; Sarawichitr, B.; Gramenzi, A.; Marsilio, F.; Steiner, J.M.; Lidbury, J.A.; Woods, G.R.T.; German, A.J.; Suchodolski, J.S. Fecal microbiota in client-owned obese dogs changes after weight loss with a high-fiber-high-protein diet. PeerJ 2020, 8, e9706. [Google Scholar] [CrossRef]
- Apper, E.; Privet, L.; Taminiau, B.; Le Bourgot, C.; Svilar, L.; Martin, J.-C.; Diez, M. Relationships between Gut Microbiota, Metabolome, Body Weight, and Glucose Homeostasis of Obese Dogs Fed with Diets Differing in Prebiotic and Protein Content. Microorganisms 2020, 8, 513. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Lanerie, D.J.; Dowd, S.E.; Paddock, C.G.; Grützner, N.; Steiner, J.M.; Ivanek, R.; Suchodolski, J.S. Effect of a multi-species synbiotic formulation on fecal bacterial microbiota of healthy cats and dogs as evaluated by pyrosequencing. FEMS Microbiol. Ecol. 2011, 78, 542–554. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, F.; Hou, Q.; Huang, W.; Liu, Y.; Zhang, H.; Sun, Z. Metagenomic analysis revealed beneficial effects of probiotics in improving the composition and function of the gut microbiota in dogs with diarrhoea. Food Funct. 2019, 10, 2618–2629. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Zhang, X.; Aweya, J.J.; Huang, Z.-X.; Kang, Z.-Y.; Bai, Z.-H.; Li, K.-H.; He, X.-T.; Liu, Y.; Chen, X.-Q.; Cheong, K.-L. In vitro fermentation of Gracilaria lemaneiformis sulfated polysaccharides and its agaro-oligosaccharides by human fecal inocula and its impact on microbiota. Carbohydr. Polym. 2020, 234, 115894. [Google Scholar] [CrossRef]
- Rose, L.; Rose, J.; Gosling, S.; Holmes, M. Efficacy of a Probiotic-Prebiotic Supplement on Incidence of Diarrhea in a Dog Shelter: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Vet. Intern. Med. 2017, 31, 377–382. [Google Scholar] [CrossRef]
- Rossi, G.; Pengo, G.; Caldin, M.; Palumbo Piccionello, A.; Steiner, J.M.; Cohen, N.D.; Jergens, A.E.; Suchodolski, J.S. Comparison of Microbiological, Histological, and Immunomodulatory Parameters in Response to Treatment with Either Combination Therapy with Prednisone and Metronidazole or Probiotic VSL#3 Strains in Dogs with Idiopathic Inflammatory Bowel Disease. PLoS ONE 2014, 9, e94699. [Google Scholar] [CrossRef]
- Makielski, K.; Cullen, J.; O’Connor, A.; Jergens, A.E. Narrative review of therapies for chronic enteropathies in dogs and cats. J. Vet. Intern. Med. 2019, 33, 11–22. [Google Scholar] [CrossRef]
- Beloshapka, A.N.; Dowd, S.E.; Suchodolski, J.S.; Steiner, J.M.; Duclos, L.; Swanson, K.S. Fecal microbial communities of healthy adult dogs fed raw meat-based diets with or without inulin or yeast cell wall extracts as assessed by 454 pyrosequencing. FEMS Microbiol. Ecol. 2013, 84, 532–541. [Google Scholar] [CrossRef]
- Wang, Y.; Ablimit, N.; Zhang, Y.; Li, J.; Wang, X.; Liu, J.; Miao, T.; Wu, L.; Wang, H.; Wang, Z.; et al. Novel β-mannanase/GLP-1 fusion peptide high effectively ameliorates obesity in a mouse model by modifying balance of gut microbiota. Int. J. Biol. Macromol. 2021, 191, 753–763. [Google Scholar] [CrossRef]
- Sowah, S.A.; Milanese, A.; Schübel, R.; Wirbel, J.; Kartal, E.; Johnson, T.S.; Hirche, F.; Grafetstätter, M.; Nonnenmacher, T.; Kirsten, R.; et al. Calorie restriction improves metabolic state independently of gut microbiome composition: A randomized dietary intervention trial. Genome Med. 2022, 14, 30. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Li, H.; Lane, J.A.; Chen, J.; Lu, Z.; Wang, H.; Dhital, S.; Fu, X.; Huang, Q.; Liu, F.; Zhang, B. In vitro fermentation of human milk oligosaccharides by individual Bifidobacterium longum-dominant infant fecal inocula. Carbohydr. Polym. 2022, 287, 119322. [Google Scholar] [CrossRef]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate mediates a microbiome–brain–β-cell axis to promote metabolic syndrome. Nature 2016, 534, 213–217. [Google Scholar] [CrossRef]
- Hong, Y.-H.; Nishimura, Y.; Hishikawa, D.; Tsuzuki, H.; Miyahara, H.; Gotoh, C.; Choi, K.-C.; Feng, D.D.; Chen, C.; Lee, H.-G.; et al. Acetate and Propionate Short Chain Fatty Acids Stimulate Adipogenesis via GPCR43. Endocrinology 2005, 146, 5092–5099. [Google Scholar] [CrossRef]
- Li, J.; Pang, B.; Yan, X.; Shang, X.; Hu, X.; Shi, J. Prebiotic properties of different polysaccharide fractions from Artemisia sphaerocephala Krasch seeds evaluated by simulated digestion and in vitro fermentation by human fecal microbiota. Int. J. Biol. Macromol. 2020, 162, 414–424. [Google Scholar] [CrossRef]
- Al-Lahham, S.H.; Peppelenbosch, M.P.; Roelofsen, H.; Vonk, R.J.; Venema, K. Biological effects of propionic acid in humans; metabolism, potential applications and underlying mechanisms. Biochim. Biophys. Acta 2010, 1801, 1175–1183. [Google Scholar] [CrossRef]
- Biagi, G.; Cipollini, I.; Zaghini, G. In vitro fermentation of different sources of soluble fiber by dog faecal inoculum. Vet. Res. Commun. 2008, 32 (Suppl. 1), 335–337. [Google Scholar] [CrossRef]
- Donadelli, R.A.; Titgemeyer, E.C.; Aldrich, C.G. Organic matter disappearance and production of short- and branched-chain fatty acids from selected fiber sources used in pet foods by a canine in vitro fermentation model1. J. Anim. Sci. 2019, 97, 4532–4539. [Google Scholar] [CrossRef]
- Fu, X.; Cao, C.; Ren, B.; Zhang, B.; Huang, Q.; Li, C. Structural characterization and in vitro fermentation of a novel polysaccharide from Sargassum thunbergii and its impact on gut microbiota. Carbohydr. Polym. 2018, 183, 230–239. [Google Scholar] [CrossRef]
- Bai, J.; Li, T.; Zhang, W.; Fan, M.; Qian, H.; Li, Y.; Wang, L. Systematic assessment of oat β-glucan catabolism during in vitro digestion and fermentation. Food Chem. 2021, 348, 129116. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Diet |
|---|---|
| Dry matter, % | 90.2 |
| Crude protein, % of dry matter | 26.0 |
| Acid hydrolyzed ether extract, % of dry matter | 11.0 |
| Total dietary fiber, % of dry matter | 5.0 |
| Ash, % of dry matter | 10.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Ding, Z.; Chen, X.; Wen, M.; Wang, Q.; Wang, Z. Effects of Oligosaccharide Fermentation on Canine Gut Microbiota and Fermentation Metabolites in an In Vitro Fecal Fermentation Model. Fermentation 2023, 9, 722. https://doi.org/10.3390/fermentation9080722
Zhang Y, Ding Z, Chen X, Wen M, Wang Q, Wang Z. Effects of Oligosaccharide Fermentation on Canine Gut Microbiota and Fermentation Metabolites in an In Vitro Fecal Fermentation Model. Fermentation. 2023; 9(8):722. https://doi.org/10.3390/fermentation9080722
Chicago/Turabian StyleZhang, Yu, Zhuang Ding, Xiaoyu Chen, Min Wen, Qingpeng Wang, and Zhengping Wang. 2023. "Effects of Oligosaccharide Fermentation on Canine Gut Microbiota and Fermentation Metabolites in an In Vitro Fecal Fermentation Model" Fermentation 9, no. 8: 722. https://doi.org/10.3390/fermentation9080722
APA StyleZhang, Y., Ding, Z., Chen, X., Wen, M., Wang, Q., & Wang, Z. (2023). Effects of Oligosaccharide Fermentation on Canine Gut Microbiota and Fermentation Metabolites in an In Vitro Fecal Fermentation Model. Fermentation, 9(8), 722. https://doi.org/10.3390/fermentation9080722