
Production of Liquid Biofuel Precursors: Optimization and Regulation of Lipase Fermentation and Its Application in Plant Oil Hydrolysis Process

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. GC Method for Determining Free Fatty Acids
2.3. Shake Flask Fermentation
2.3.1. The Impact of the Citric Acid Addition in a Shaking Flask
2.3.2. The Impact of the Types and Amounts of Amino Acids in a Shaking Flask
2.4. Fermentation Process of 5 L Fermentation Tank
2.4.1. The Impact of the Citric Acid Addition in a 5 L Fermentation Tank
2.4.2. The Impact of the Carbon Source Automatic Replenishment Scheme in a 5 L Fermentation Tank
2.4.3. The Impact of the Compound Amino Acids in a 5 L Fermentation Tank
2.4.4. Multibatch Stable Fermentation in a 5 L Fermentation Tank
2.5. Lipase Hydrolysis Technology of Soybean Oil
2.5.1. The Impact of the Material Flow Rate on the Production of Free Fatty Acids
2.5.2. The Impact of the Reaction Temperature on the Production of Free Fatty Acids
2.5.3. The Impact of the Water Content on the Production of Free Fatty Acids
2.5.4. The Impact of the Lipase Fermentation Broth Dosage on the Production of Free Fatty Acids
2.5.5. The Impact of the Reuse of Lipase Fermentation Broth on the Production of Free Fatty Acids
3. Results and Discussion
3.1. Adjusting the Medium Formula and Optimizing Cultivation Conditions
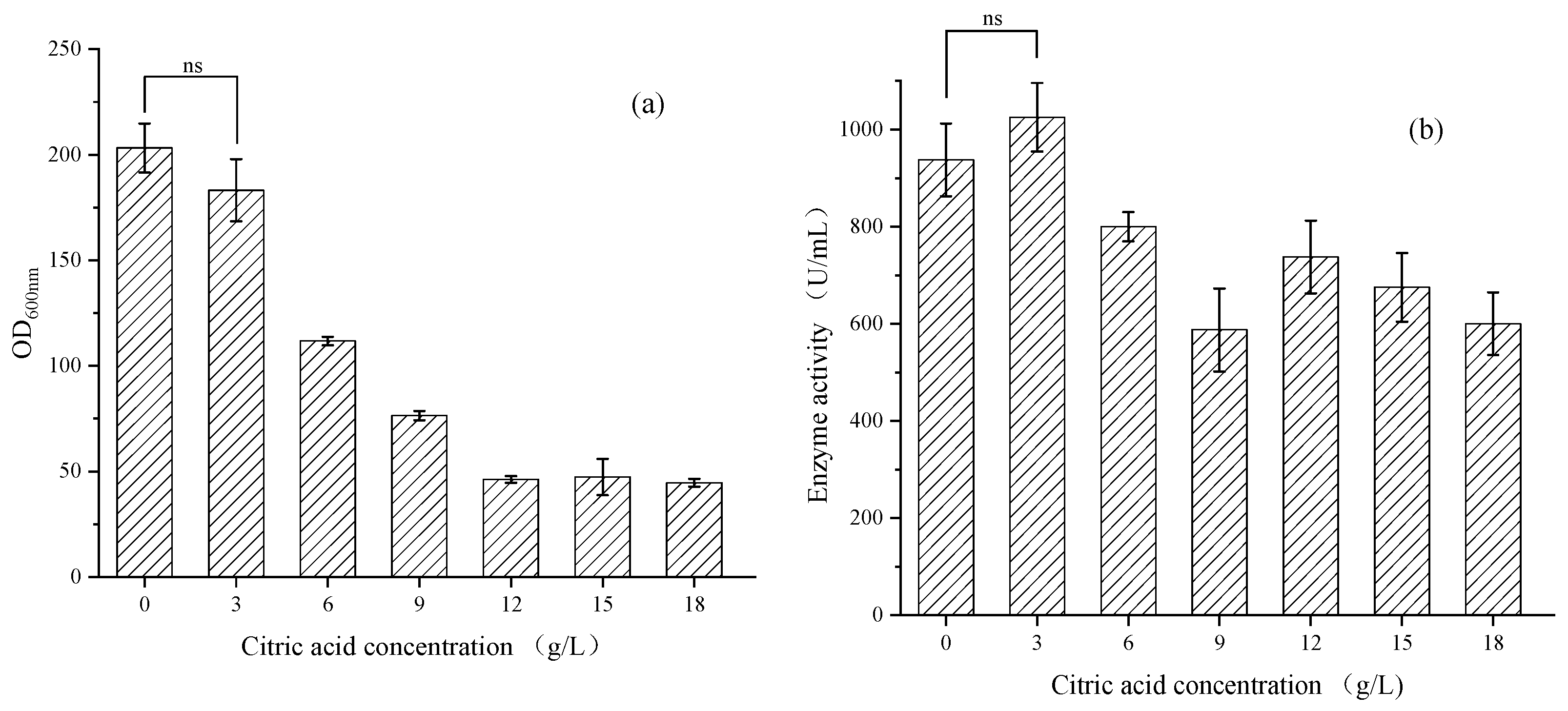
3.1.1. Optimization of Citric Acid Addition in a Shaking Flask
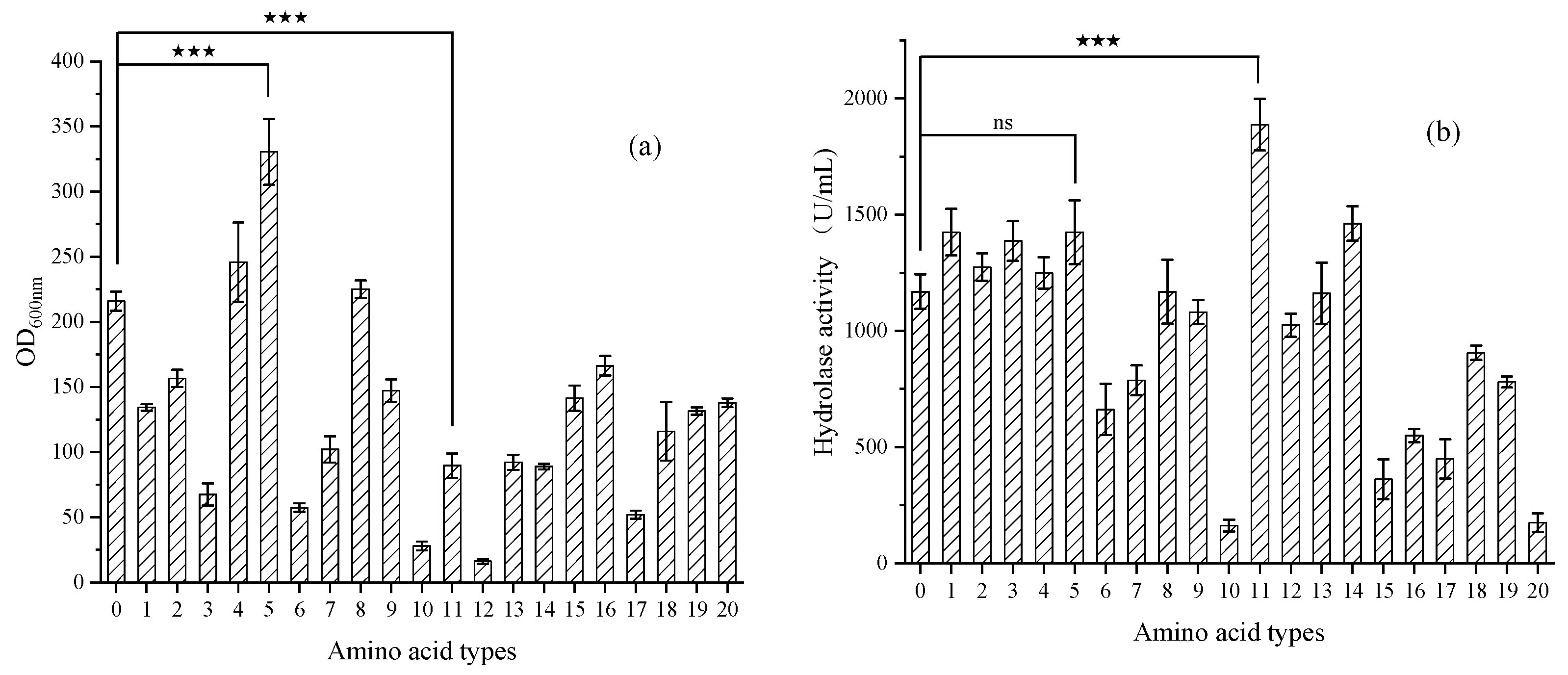
3.1.2. Amino Acid Screening and Optimization of Composite Amino Acids
Pre-Experiment for Screening 20 Amino Acids
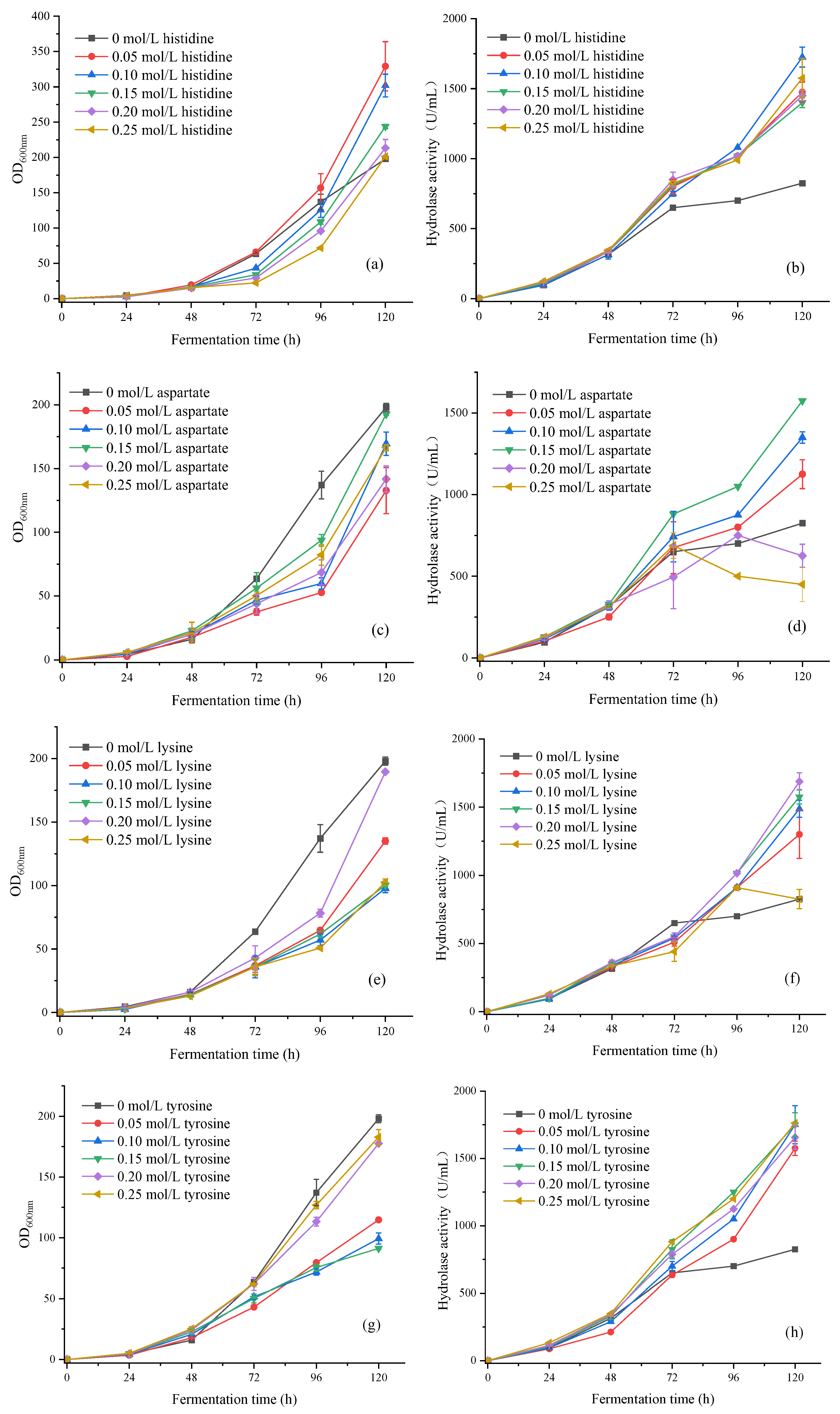
Optimization Experiment of Four Amino Acid Concentration Gradients
Composite Optimization of Two Amino Acids
Optimization of Three Composite Amino Acids
Optimization of Four Compound Amino Acids
3.2. Process Optimization of Y. lipolytica in a 5 L Fermentation Tank
3.2.1. Optimization of Citric Acid Addition in a 5 L Fermentation Tank
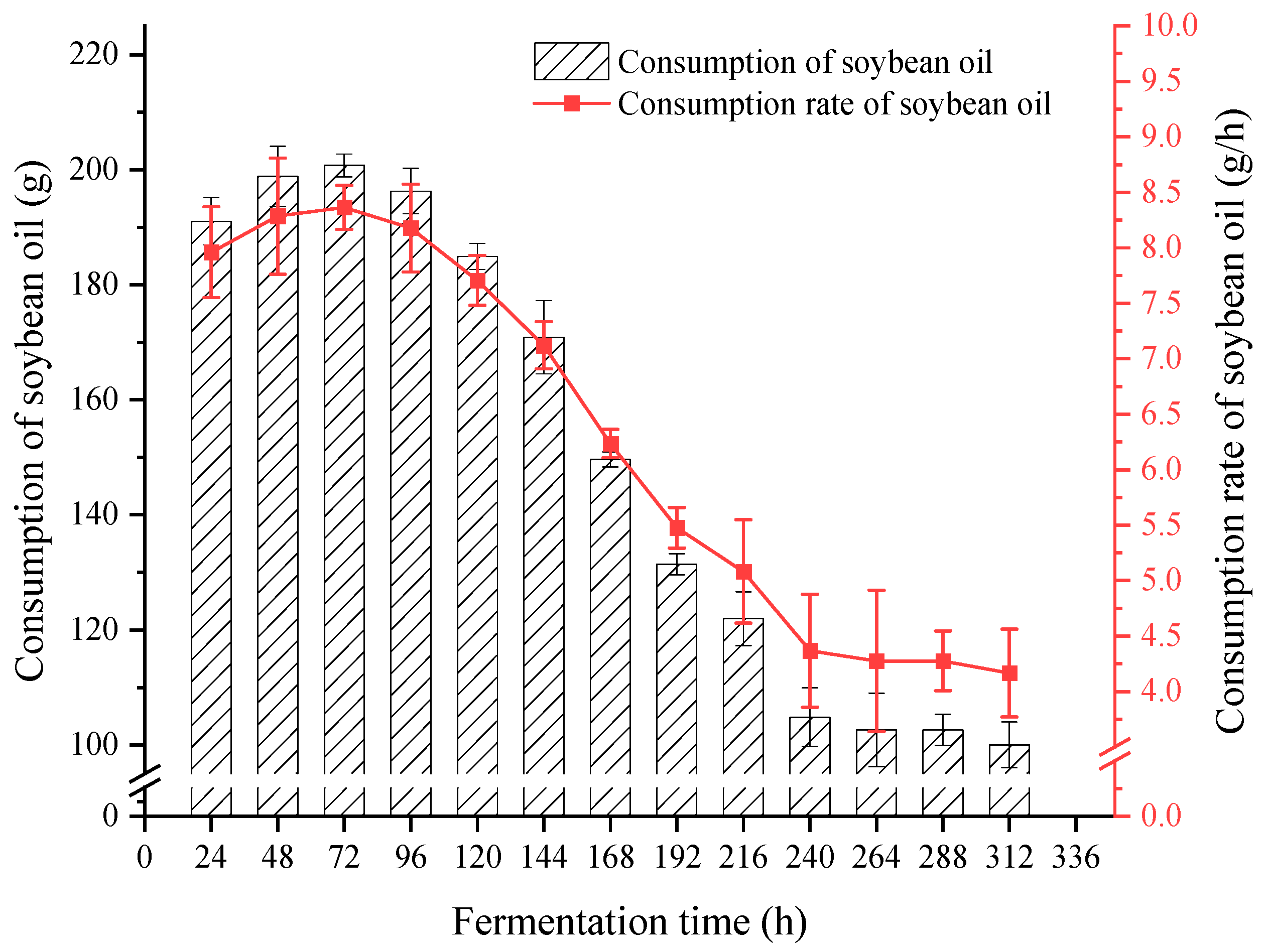
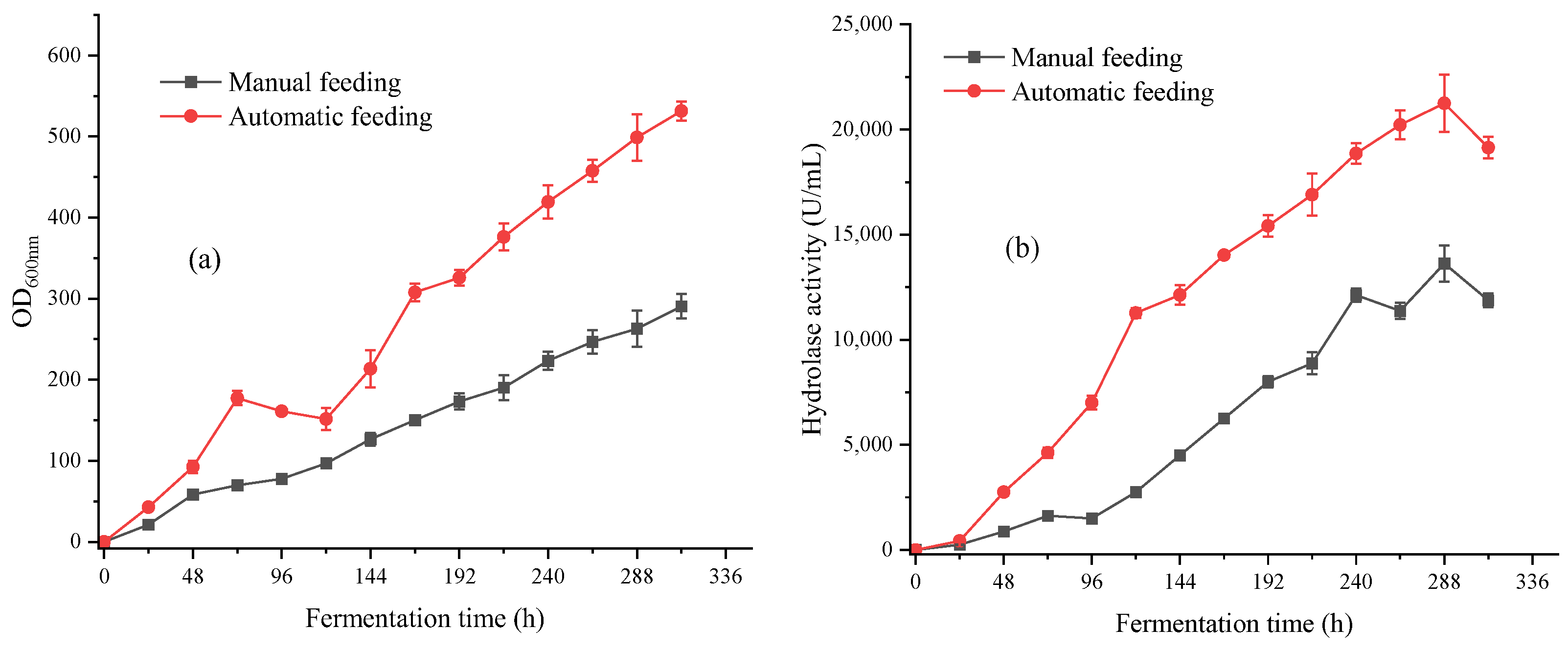
3.2.2. Design of Carbon Source Automatic Replenishment Scheme in a 5 L Fermentation Tank
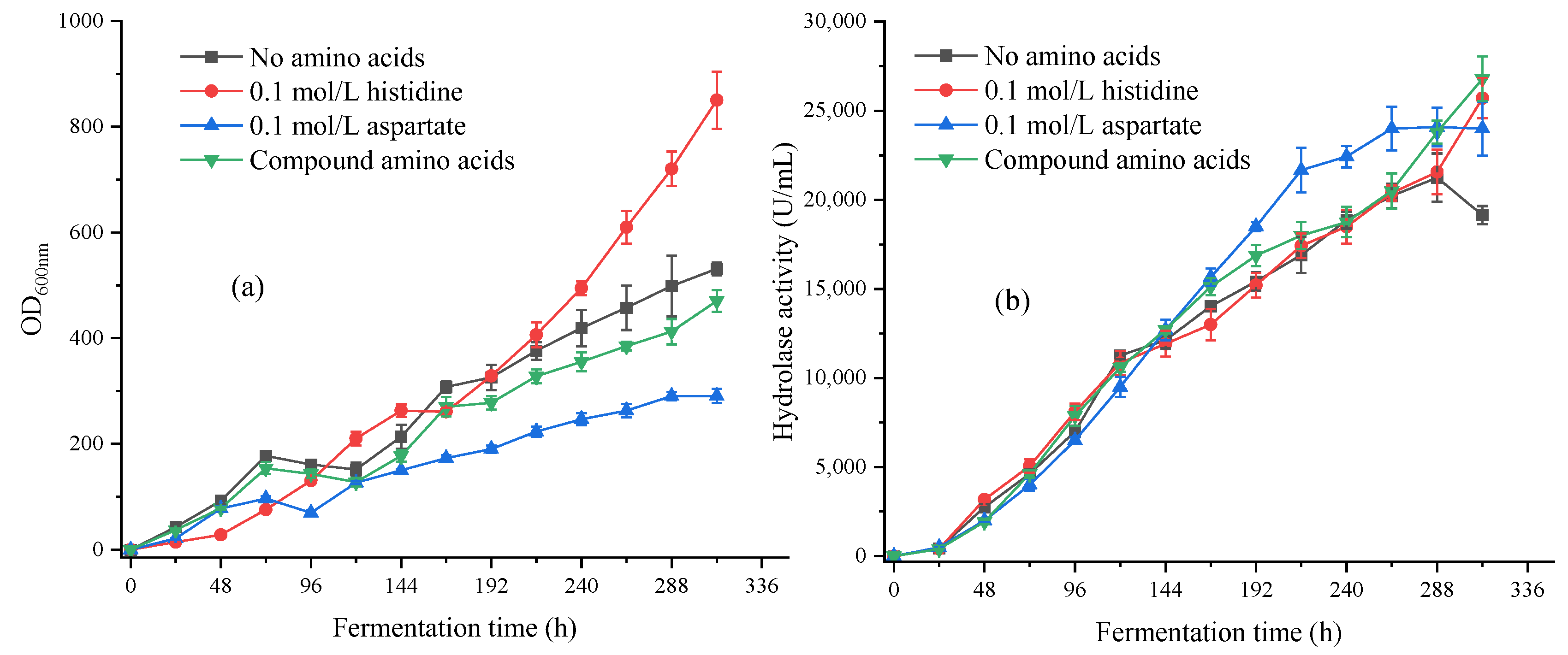
3.2.3. Optimization of Compound Amino Acids in a 5 L Fermentation Tank
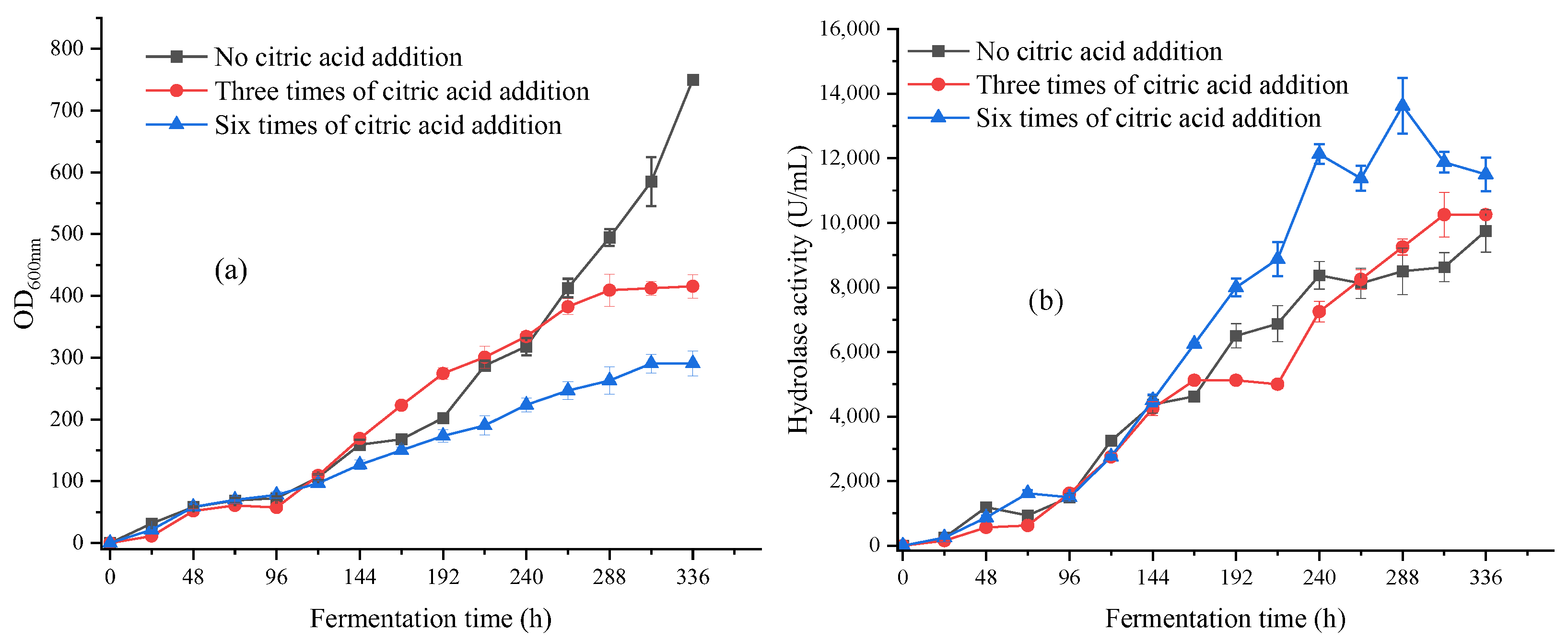
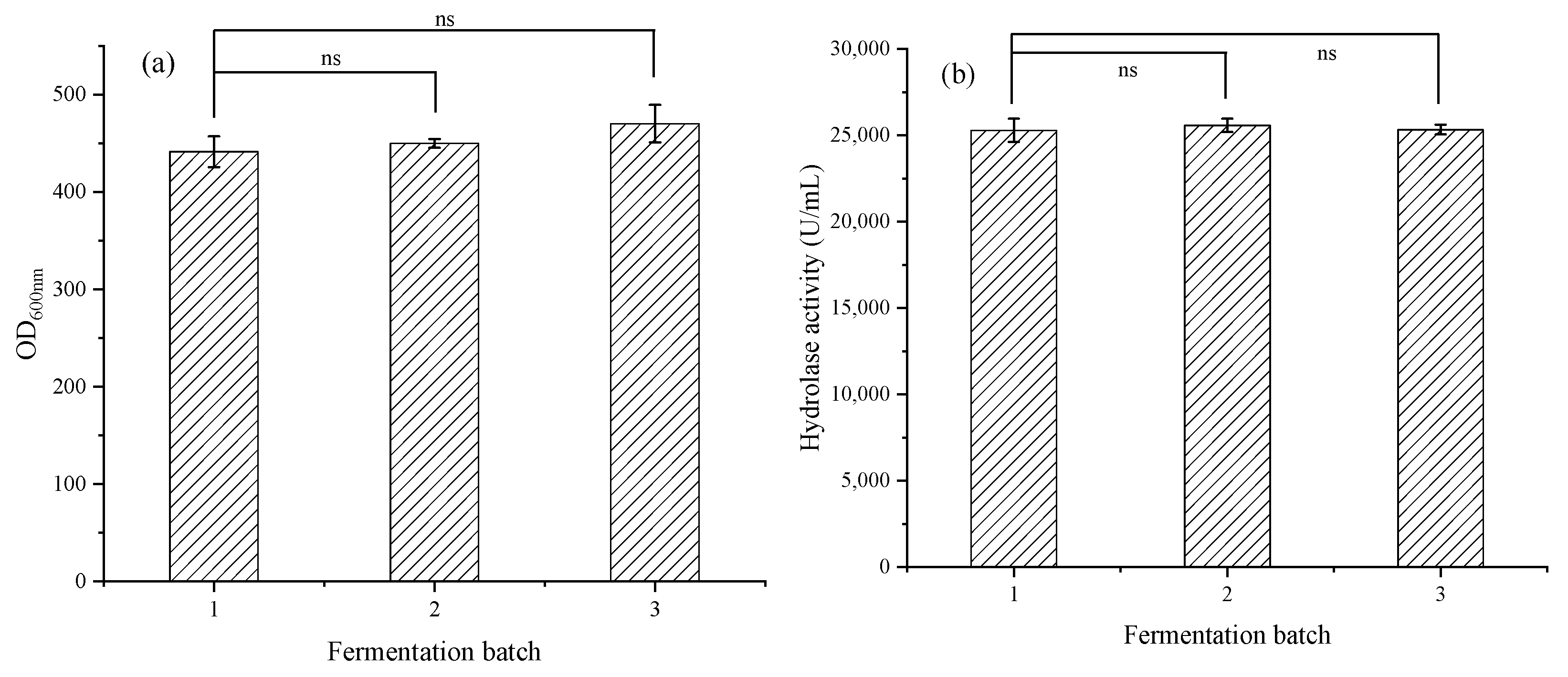
3.2.4. Multibatch Stable Fermentation
3.3. Optimization of Production Process for Free Fatty Acids
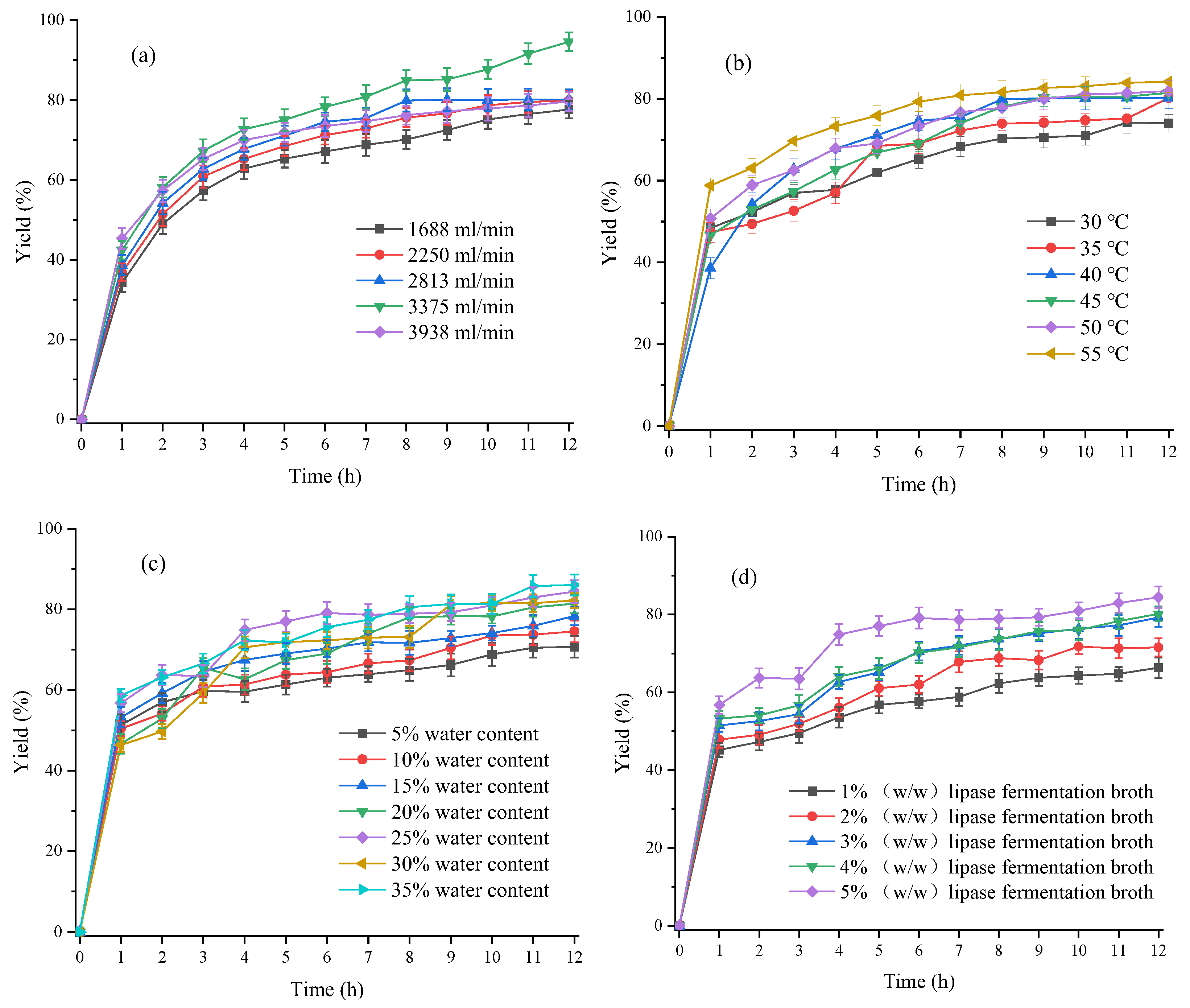
3.3.1. Optimization of Material Flow Rate in Free Fatty Acids Production Process
3.3.2. Optimization of Reaction Temperature in the Production Process of Free Fatty Acids
3.3.3. Optimization of Water Content in the Production Process of Free Fatty Acids
3.3.4. Optimization of Lipase Fermentation Broth Dosage in the Production Process of Free Fatty Acids
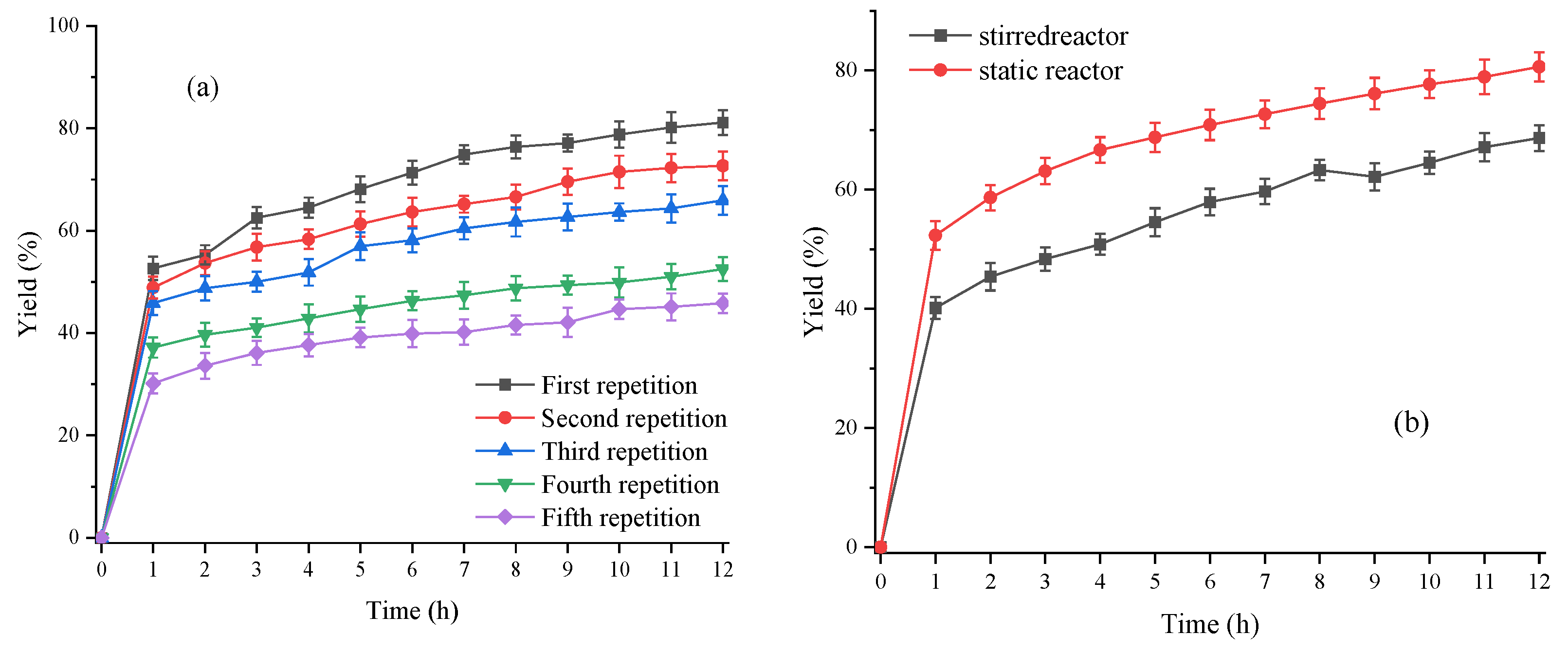
3.3.5. Reuse of Lipase Fermentation Broth
3.3.6. Comparison of Fatty Acid Production in Stirred Reactors and Static Mixers
4. Limitation and Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Paliwal, M. Natural Gas Is It the Alternate Fuel? Sol. Q. 2017, 5, 46–51. [Google Scholar]
- Garba, M.D.; Usman, M.; Khan, S.; Shehzad, F.; Galadima, A.; Ehsan, M.F.; Ghanem, A.S.; Humayun, M. CO2 towards fuels: A review of catalytic conversion of carbon dioxide to hydrocarbons. J. Environ. Chem. Eng. 2021, 9, 104756. [Google Scholar] [CrossRef]
- Arora, N.K.; Mishra, I. United Nations Sustainable Development Goals 2030 and environmental sustainability: Race against time. Environ. Sustain. 2019, 2, 339–342. [Google Scholar] [CrossRef]
- Park, H.Y.; Han, K.; Kim, H.H.; Park, S.; Jang, J.; Yu, G.S.; Ko, J.H. Comparisons of combustion characteristics between bioliquid and heavy fuel oil combustion in a 0.7 MWth pilot furnace and a 75 MWe utility boiler. Energy 2020, 192, 116557. [Google Scholar] [CrossRef]
- Sandesh, K.; Ujwal, P. Trends and perspectives of liquid biofuel—Process and industrial viability. Energy Convers. Manag. X 2021, 10, 100075. [Google Scholar] [CrossRef]
- Løkke, S.; Løkke, S.; Aramendia, E.; Malskær, J. A review of public opinion on liquid biofuels in the EU: Current knowledge and future challenges. Biomass Bioenergy 2021, 150, 106094. [Google Scholar] [CrossRef]
- Sarkar, A.; Das, P.; Laskar, I.B.; Vadivel, S.; Puzari, A.; Paul, B. Parkia speciosa: A basic heterogeneous catalyst for production of soybean oil-based biodiesel. Fuel 2023, 348, 128537. [Google Scholar] [CrossRef]
- Naeem, A.; Zaman, S.; Farooq, M.; Khan, I.W.; Ghazi, Z.A.; Saeed, T.; Hamayun, M. Biodiesel production from waste cooking oil employing natural bentonite supported heterogeneous catalyst: Waste to biodiesel. Korean J. Chem. Eng. 2022, 39, 1450–1459. [Google Scholar] [CrossRef]
- Reena Singh, R.; Bux, F.; Sharma, Y.C. Optimization of biodiesel synthesis from microalgal (Spirulina platensis) oil by using a novel heterogeneous catalyst, β-strontium silicate (β-Sr2SiO4). Fuel 2020, 280, 118312. [Google Scholar] [CrossRef]
- Murty, V.R.; Bhat, J.; Muniswaran, P.K.A. Hydrolysis of oils by using immobilized lipase enzyme: A review. Biotechnol. Bioprocess Eng. 2002, 7, 57–66. [Google Scholar] [CrossRef]
- Souza, G.P.R.; Correia, T.B.A.; Reis, W.S.M.; Bredda, E.H.; Da Rós, P.C.M.; Pereira, E.B. Enzymatic Hydrolysis of Waste Cooking Oil by Lipase Catalysis: Simplex Mixture Design Optimization. Catal. Lett. 2023, 153, 689–697. [Google Scholar] [CrossRef]
- Yulianto, M.E.; Amalia, R.; Paramita, V.; Nisa, Q.A.K. Preliminary study of auto catalytic palm oil hydrolysis into fatty acid through hydrothermalysis process. J. Phys. Conf. Ser. 2020, 1524, 012085. [Google Scholar] [CrossRef]
- Avelar, M.H.M.; Cassimiroa, D.M.J.; Santos, K.C.; Domingues, R.C.C.; De Castro, H.F.; Mendes, A.A. Hydrolysis of vegetable oils catalyzed by lipase extract powder from dormant castor bean seeds. Ind. Crop. Prod. 2013, 44, 452–458. [Google Scholar] [CrossRef]
- Salgado, C.A.; Isabela, A.; Vanetti, M.D. Microbial lipases: Propitious biocatalysts for the food industry. Food Biosci. 2022, 45, 101509. [Google Scholar] [CrossRef]
- Tongs, M.S. Ectoenzymes of streptococci. J. Am. Med. Assoc. 1919, 73, 1277–1279. [Google Scholar] [CrossRef]
- Mohammed, A.B.A.; Hegazy, A.E.; Salah, A. Predigested high-fat meats based on Lactobacillus fermentum lipase enzyme immobilized on silver-alginate nanoparticle matrix. Appl. Nanosci. 2023, 13, 641–649. [Google Scholar] [CrossRef]
- Jonathan, A.A.; Irina, B. Engineering of Yarrowia lipolytica for terpenoid production. Metab. Eng. Commun. 2022, 15, e00213. [Google Scholar]
- Cokun, G.; Plak, Z.; Yldz, N.; Mehmetolu, L. Immobilization of Candida antarctica Lipase on Nanomaterials and Investigation of the Enzyme Activity and Enantioselectivity. Appl. Biochem. Biotech. 2021, 193, 430–445. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Zhang, X.H. Evolution of Subfamily I.1 Lipases in Pseudomonas aeruginosa. Curr. Microbiol. 2021, 78, 3494–3504. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich-Wyder, M.T.; Arias-Roth, E.; Jakob, E. Cheese yeasts. Yeast 2019, 36, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, J.B.; Cornelison, C.; Crow, S.A. CANDIDA|Yarrowia lipolytica (Candida lipolytica)—ScienceDirect. In Encyclopedia of Food Microbiology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 374–378. [Google Scholar]
- Gonçalves, F.A.G.; Colen, G.; Takahashi, J.A. Yarrowia lipolytica and Its Multiple Applications in the Biotechnological Industry. Sci. World J. 2014, 2014, 476207. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Cui, Z.H.; Zhu, R.S.; Chen, C.J.; Song, S.Y.; Song, J.T.; Wang, M.; Tan, T.W. Design and development of a new static mixing bioreactor for enzymatic bioprocess: Application in biodiesel production. Renew. Energy 2022, 197, 922–931. [Google Scholar] [CrossRef]
- Park, P.W.; Goins, R.E. In Situ Preparation of Fatty Acid Methyl Esters for Analysis of Fatty Acid Composition in Foods. J. Food Sci. 1994, 59, 1262–1266. [Google Scholar] [CrossRef]
- Rocha, T.N.; Carballares, D.; Guimaraes, J.R.; Rocha-Martin, J.; Tardioli, P.W.; Gonçalves, L.R.B.; Fernandez-Lafuente, R. Determination of immobilized lipase stability depends on the substrate and activity determination condition: Stress inactivations and optimal temperature as biocatalysts stability indicators. Sustain. Chem. Pharm. 2022, 29, 100823. [Google Scholar] [CrossRef]
- Lv, L.L.; Dai, L.M.; Du, W.; Liu, D.H. Effect of water on lipase NS81006-catalyzed alcoholysis for biodiesel production. Process Biochem. 2017, 58, 239–244. [Google Scholar] [CrossRef]
- Zhou, X.N.; Zhang, W.; Zhao, L.W.; Gao, S.; Liu, T.Y.; Yu, D.Y. Immobilization of lipase in chitosan-mesoporous silica material and pore size adjustment. Int. J. Biol. Macromol. 2023, 235, 123789. [Google Scholar] [CrossRef]
- Guebara, S.A.B.; Ract, J.N.R.; Vitolo, M. Conversion of Caprylic Acid into Mono-, Di- and Triglycerides Using Immobilized Lipase. Arab. J. Sci. Eng. 2018, 43, 3631–3637. [Google Scholar] [CrossRef]
- Tan, T.W.; Zhang, M.; Wang, B.W.; Ying, C.H.; Deng, L. Screening of high lipase producing Candida sp. and production of lipase by fermentation. Process Biochem. 2003, 39, 459–465. [Google Scholar] [CrossRef]
- He, Y.Q.; Tan, T.W. Use of response surface methodology to optimize culture medium for production of lipase with Candida sp. 99–125. J. Mol. Catal. B-Enzym. 2006, 43, 9–14. [Google Scholar] [CrossRef]
- Nie, K.L.; Wang, M.; Zhang, X.; Hu, W.J.; Liu, L.; Wang, F.; Deng, L.; Tan, T.W. Additives improve the enzymatic synthesis of biodiesel from waste oil in a solvent free system. Fuel 2015, 146, 13–19. [Google Scholar] [CrossRef]
- Cao, H.; Wang, M.; Deng, L.; Liu, L.; Schwaneberg, U.; Tan, T.W.; Wang, F.; Nie, K.L. Sugar-Improved Enzymatic Synthesis of Biodiesel with Yarrowia lipolytica Lipase 2. Energy Fuel 2017, 31, 6248–6256. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid Molecular Formula | Types of Fatty Acids | Composition (wt%) |
|---|---|---|
| C16:0 | Palmitic acid | 11.56 |
| C18:0 | Stearic acid | 2.83 |
| C18:1 | Oleic acid | 20.44 |
| C18:2 | Linoleic acid | 56.15 |
| C18:3 | Linolenic acid | 8.68 |
| C20:0 | Arachidic acid | 0.34 |
| Serial Number | A | B | OD600nm | Hydrolase Activity (U/mL) |
|---|---|---|---|---|
| Histidine (mol/L) | Aspartate (mol/L) | |||
| 1 | 0.05 | 0 | 329.18 ± 34.90 | 1475.0 ± 70.71 |
| 2 | 0.05 | 0.03 | 113.13 ± 4.42 | 1520.0 ± 167.94 |
| 3 | 0.05 | 0.06 | 166.88 ± 5.83 | 1957.5 ± 20.89 |
| 4 | 0.05 | 0.09 | 179.40 ± 1.70 | 1867.5 ± 68.10 |
| 5 | 0.05 | 0.12 | 116.25 ± 4.95 | 1710.0 ± 29.64 |
| 6 | 0.05 | 0.15 | 104.00 ± 3.54 | 1665.0 ± 25.68 |
| 7 | 0.1 | 0 | 301.88 ± 16.09 | 1725.0 ± 58.96 |
| 8 | 0.1 | 0.03 | 200.00 ± 3.98 | 1912.5 ± 127.28 |
| 9 | 0.1 | 0.06 | 165.63 ± 13.26 | 1822.5 ± 20.64 |
| 10 | 0.1 | 0.09 | 133.13 ± 8.31 | 1710.0 ± 95.46 |
| 11 | 0.1 | 0.12 | 96.50 ± 1.06 | 1417.5 ± 63.64 |
| 12 | 0.1 | 0.15 | 108.00 ± 5.30 | 1507.5 ± 25.98 |
| 13 | 0.15 | 0 | 243.95 ± 2.97 | 1400.0 ± 35.36 |
| 14 | 0.15 | 0.03 | 173.00 ± 3.18 | 1665.0 ± 30.58 |
| 15 | 0.15 | 0.06 | 140.00 ± 1.58 | 1665.0 ± 98.59 |
| 16 | 0.15 | 0.09 | 117.13 ± 7.25 | 1620.0 ± 31.82 |
| 17 | 0.15 | 0.12 | 128.88 ± 4.54 | 1462.5 ± 65.49 |
| 18 | 0.15 | 0.15 | 118.88 ± 1.59 | 1507.5 ± 60.98 |
| Serial Number | A | B | OD600nm | Hydrolase Activity (U/mL) | |
|---|---|---|---|---|---|
| Histidine (mol/L) | Aspartate (mol/L) | Lysine (mol/L) | |||
| 1 | 0.05 | 0.06 | 0 | 166.88 ± 5.83 | 1957.50 ± 20.89 |
| 2 | 0.05 | 0.06 | 0.03 | 86.25 ± 0.67 | 1656.25 ± 128.21 |
| 3 | 0.05 | 0.06 | 0.06 | 86.50 ± 0.34 | 1750.00 ± 93.60 |
| 4 | 0.05 | 0.06 | 0.09 | 107.88 ± 0.84 | 2242.50 ± 140.51 |
| 5 | 0.05 | 0.06 | 0.12 | 95.25 ± 2.79 | 2031.25 ± 126.88 |
| 6 | 0.05 | 0.09 | 0 | 179.40 ± 1.70 | 1867.50 ± 68.10 |
| 7 | 0.05 | 0.09 | 0.03 | 139.50 ± 2.77 | 1906.25 ± 43.22 |
| 8 | 0.05 | 0.09 | 0.06 | 31.25 ± 0.69 | 1125.00 ± 175.01 |
| 9 | 0.05 | 0.09 | 0.09 | 37.50 ± 1.37 | 1312.50 ± 84.76 |
| 10 | 0.05 | 0.09 | 0.12 | 77.38 ± 0.86 | 1375.00 ± 86.71 |
| 11 | 0.1 | 0.03 | 0 | 200.00 ± 3.98 | 1912.50 ± 127.28 |
| 12 | 0.1 | 0.03 | 0.03 | 188.75 ± 0.35 | 2117.50 ± 112.94 |
| 13 | 0.1 | 0.03 | 0.06 | 151.50 ± 1.03 | 2031.25 ± 42.21 |
| 14 | 0.1 | 0.03 | 0.09 | 61.38 ± 1.26 | 1843.75 ± 44.81 |
| 15 | 0.1 | 0.03 | 0.12 | 113.13 ± 1.66 | 2093.75 ± 45.65 |
| 16 | 0.1 | 0.06 | 0 | 165.63 ± 13.26 | 1822.5 ± 20.64 |
| 17 | 0.1 | 0.06 | 0.03 | 143.63 ± 0.93 | 2127.50 ± 79.77 |
| 18 | 0.1 | 0.06 | 0.06 | 124.50 ± 2.87 | 1906.25 ± 46.09 |
| 19 | 0.1 | 0.06 | 0.09 | 112.13 ± 0.54 | 1906.25 ± 45.61 |
| 20 | 0.1 | 0.06 | 0.12 | 111.13 ± 5.96 | 1875.00 ± 18.07 |
| Serial Number | A | B | OD600nm | Hydrolase Activity (U/mL) | ||
|---|---|---|---|---|---|---|
| Histidine (mol/L) | Aspartate (mol/L) | Lysine (mol/L) | Tyrosine (mol/L) | |||
| 1 | 0.05 | 0.06 | 0.09 | 0 | 107.88 ± 0.84 | 2242.5 ± 140.51 |
| 2 | 0.05 | 0.06 | 0.09 | 0.03 | 86.25 ± 0.69 | 1957.5 ± 30.36 |
| 3 | 0.05 | 0.06 | 0.09 | 0.06 | 86.50 ± 0.34 | 1755.0 ± 44.14 |
| 4 | 0.05 | 0.06 | 0.09 | 0.09 | 107.88 ± 0.85 | 1777.5 ± 90.59 |
| 5 | 0.05 | 0.06 | 0.09 | 0.12 | 95.25 ± 2.70 | 1800.0 ± 58.90 |
| 6 | 0.1 | 0.03 | 0.03 | 0 | 188.75 ± 0.35 | 2117.5 ± 112.94 |
| 7 | 0.1 | 0.03 | 0.03 | 0.03 | 139.50 ± 2.69 | 1890.0 ± 62.24 |
| 8 | 0.1 | 0.03 | 0.03 | 0.06 | 31.25 ± 0.68 | 1552.5 ± 155.12 |
| 9 | 0.1 | 0.03 | 0.03 | 0.09 | 37.50 ± 1.37 | 1687.5 ± 30.87 |
| 10 | 0.1 | 0.03 | 0.03 | 0.12 | 77.38 ± 0.85 | 1710.0 ± 25.85 |
| 11 | 0.1 | 0.06 | 0.03 | 0 | 151.50 ± 1.03 | 2031.25 ± 42.21 |
| 12 | 0.1 | 0.06 | 0.03 | 0.03 | 188.75 ± 0.34 | 1800.0 ± 124.86 |
| 13 | 0.1 | 0.06 | 0.03 | 0.06 | 151.50 ± 1.03 | 1732.5 ± 30.77 |
| 14 | 0.1 | 0.06 | 0.03 | 0.09 | 61.38 ± 1.29 | 1665.0 ± 64.59 |
| 15 | 0.1 | 0.06 | 0.03 | 0.12 | 113.13 ± 1.62 | 1642.5 ± 99.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.; Li, H.; Zhu, R.; Wang, M.; Tan, T. Production of Liquid Biofuel Precursors: Optimization and Regulation of Lipase Fermentation and Its Application in Plant Oil Hydrolysis Process. Fermentation 2023, 9, 708. https://doi.org/10.3390/fermentation9080708
Huang S, Li H, Zhu R, Wang M, Tan T. Production of Liquid Biofuel Precursors: Optimization and Regulation of Lipase Fermentation and Its Application in Plant Oil Hydrolysis Process. Fermentation. 2023; 9(8):708. https://doi.org/10.3390/fermentation9080708
Chicago/Turabian StyleHuang, Shuai, Hao Li, Ruisong Zhu, Meng Wang, and Tianwei Tan. 2023. "Production of Liquid Biofuel Precursors: Optimization and Regulation of Lipase Fermentation and Its Application in Plant Oil Hydrolysis Process" Fermentation 9, no. 8: 708. https://doi.org/10.3390/fermentation9080708
APA StyleHuang, S., Li, H., Zhu, R., Wang, M., & Tan, T. (2023). Production of Liquid Biofuel Precursors: Optimization and Regulation of Lipase Fermentation and Its Application in Plant Oil Hydrolysis Process. Fermentation, 9(8), 708. https://doi.org/10.3390/fermentation9080708