
Transcription Analysis of the Acid Tolerance Mechanism of Pichia kudriavzevii NBRC1279 and NBRC1664


Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Media
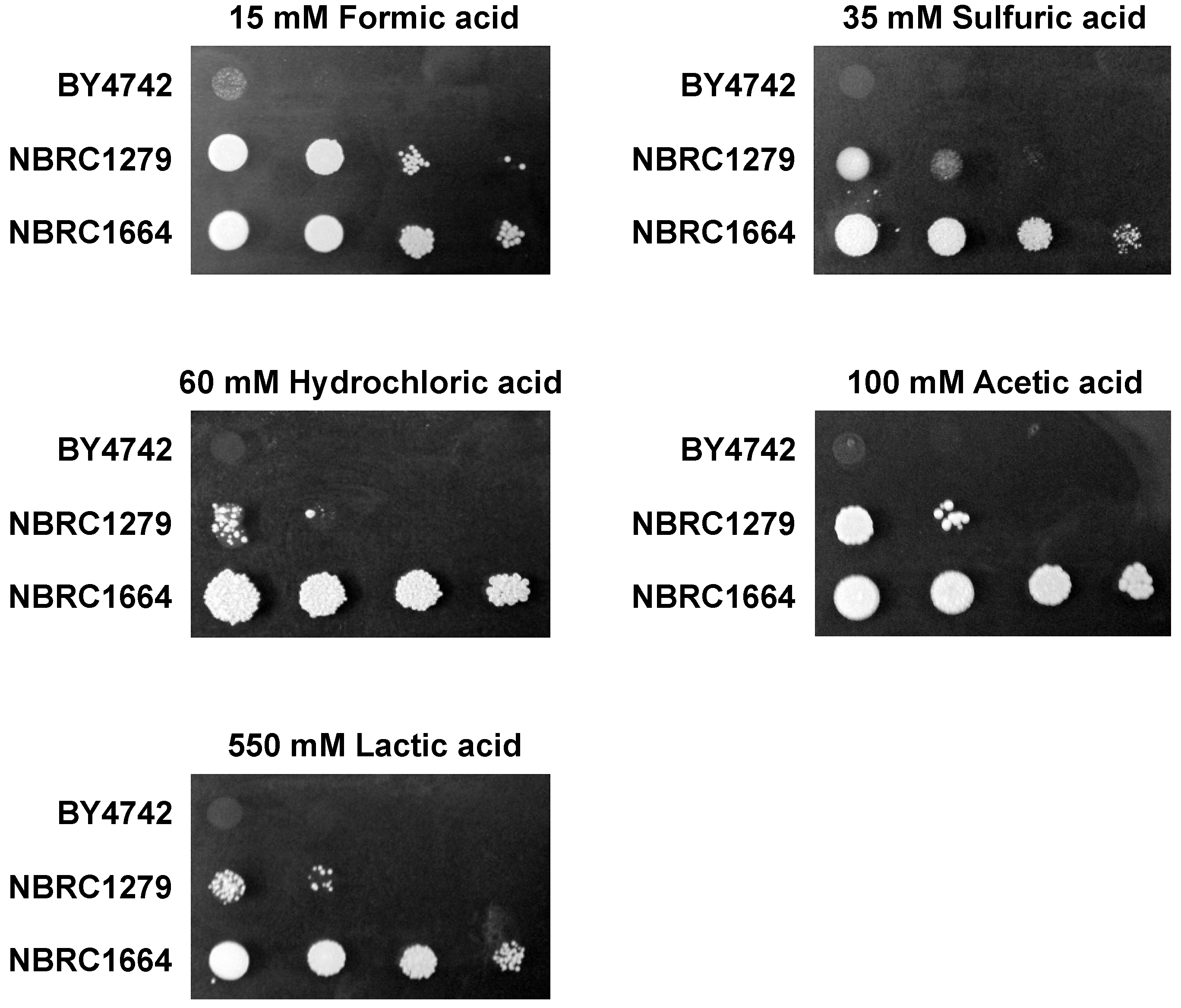
2.2. Spot Assay
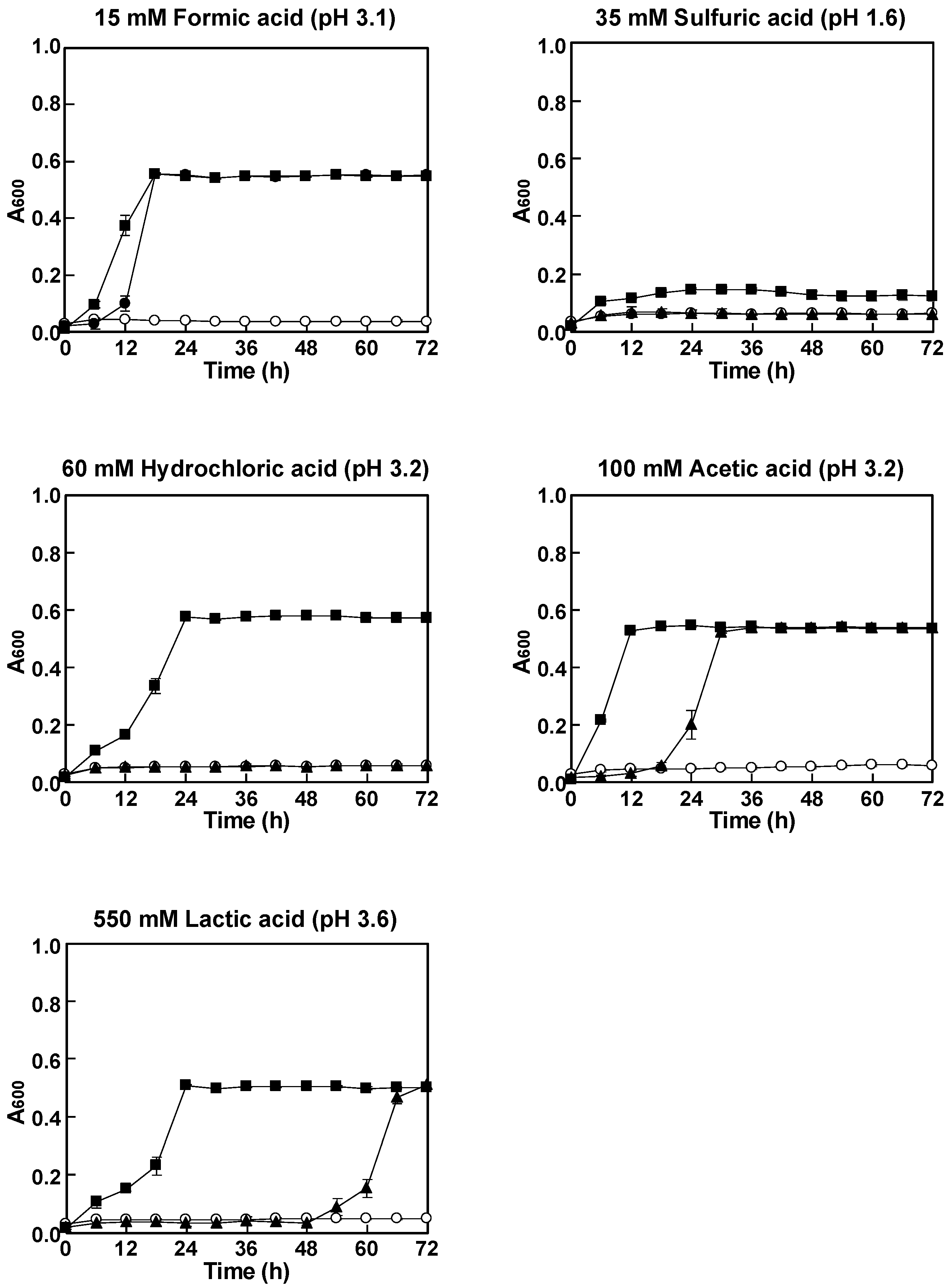
2.3. Aerobic Growth Experiment
2.4. RNA Preparation
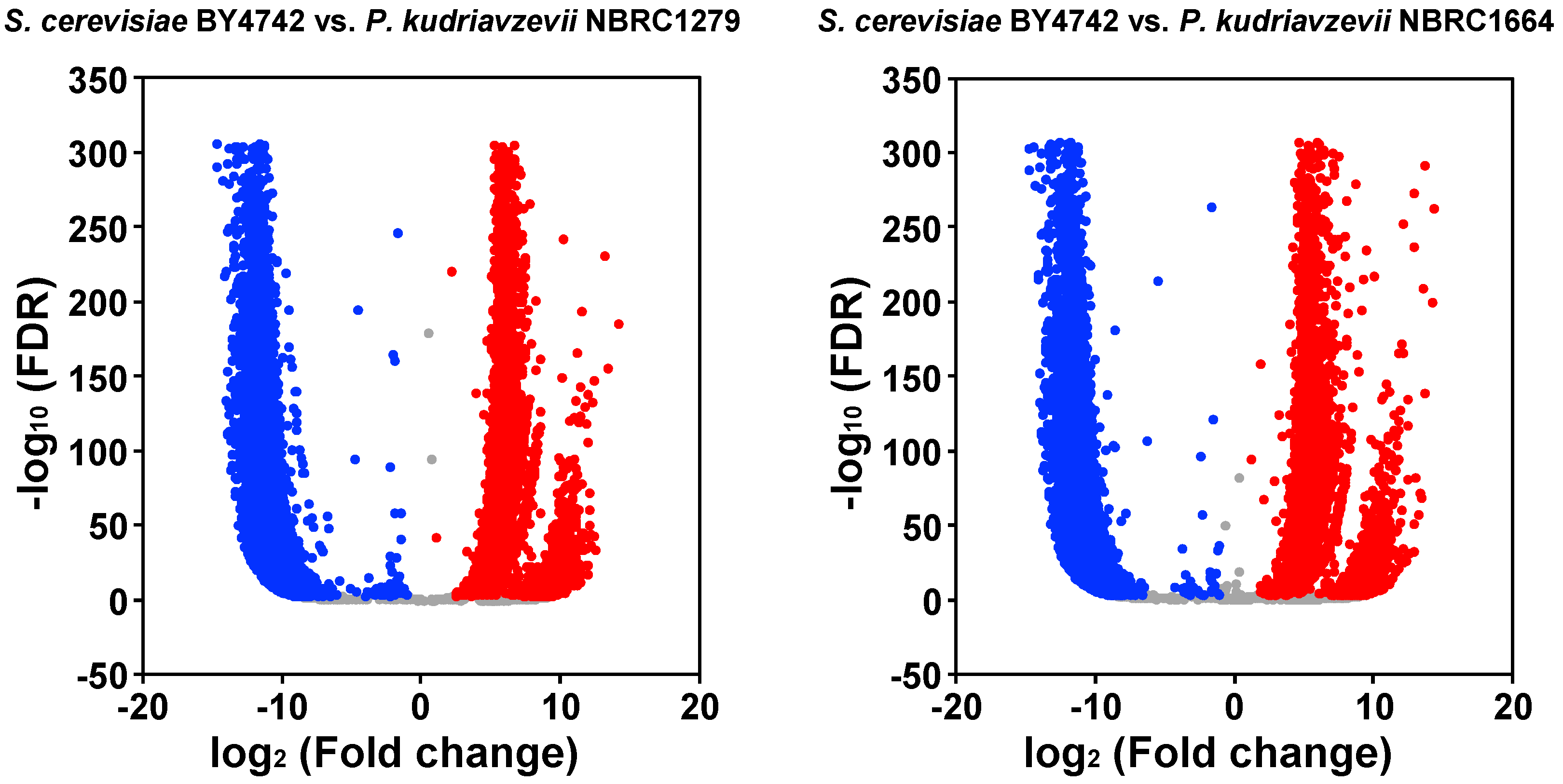
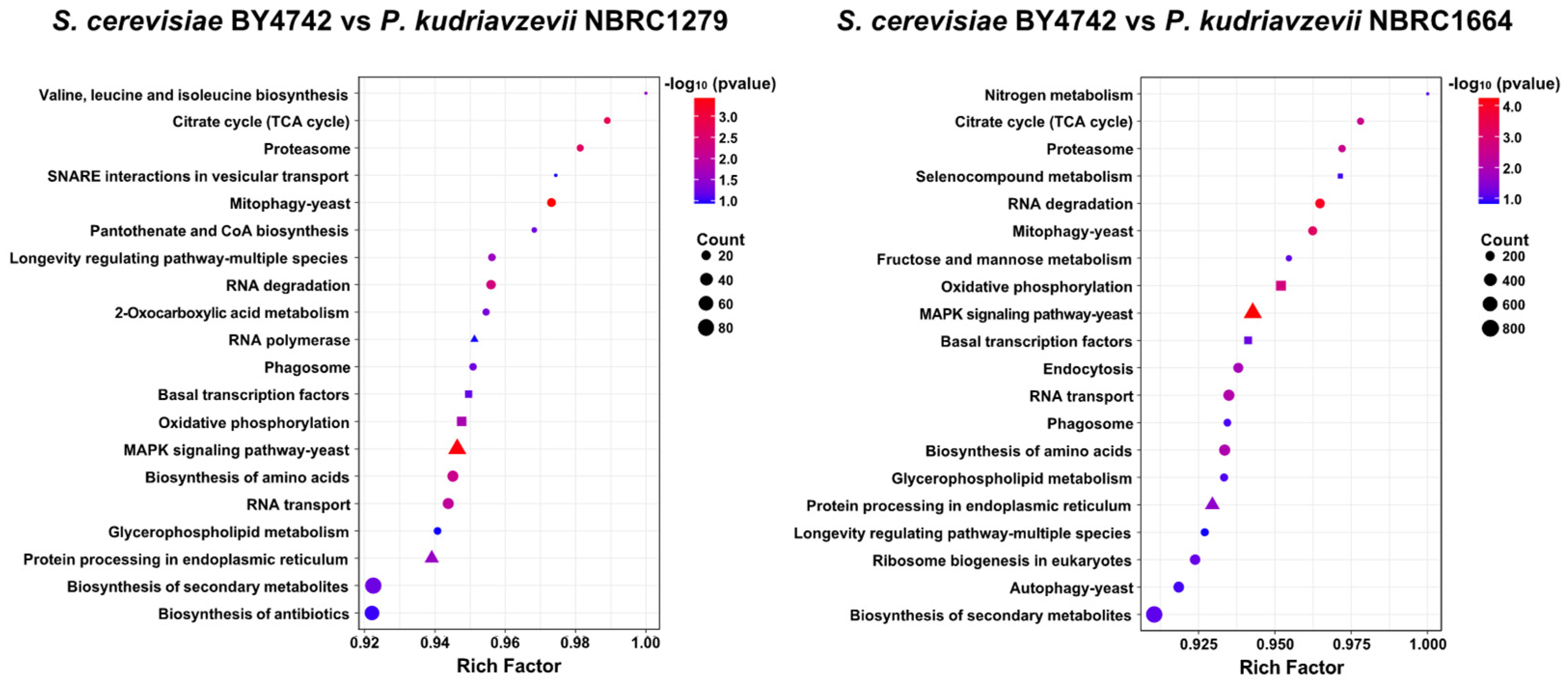
2.5. Transcriptome Analysis
3. Results and Discussion
3.1. Effect of Different Acids on Growth
3.2. Transcriptome Analysis of Yeast Strains under Hydrochloric Acid Stress
3.3. Involvement of the HOG Pathway in Acid Stress Tolerance
3.4. Effect of Gpd1 Expression on Acid Tolerance
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tse, T.J.; Wiens, D.J.; Reaney, M.J.T. Production of Bioethanol—A Review of Factors Affecting Ethanol Yield. Fermentation 2021, 7, 268. [Google Scholar] [CrossRef]
- Robak, K.; Balcerek, M. Review of second generation bioethanol production from residual biomass. Food Technol. Biotechnol. 2018, 56, 174–187. [Google Scholar] [CrossRef]
- Agbor, V.B.; Cicek, N.; Sparling, R.; Berlin, A.; Levin, D.B. Biomass pretreatment: Fundamentals toward application. Biotechnol. Adv. 2011, 29, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Matsushika, A.; Inoue, H.; Murakami, K.; Takimura, O.; Sawayama, S. Bioethanol production performance of five recombinant strains of laboratory and industrial xylose-fermenting Saccharomyces cerevisiae. Bioresour. Technol. 2009, 100, 2392–2398. [Google Scholar] [CrossRef] [PubMed]
- Matsushika, A.; Watanabe, S.; Kodaki, T.; Makino, K.; Inoue, H.; Murakami, K.; Takimura, O.; Sawayama, S. Expression of protein engineered NADP+-dependent xylitol dehydrogenase increases ethanol production from xylose in recombinant Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2008, 81, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Murakami, K.; Endo, T.; Fujimoto, S.; Minowa, T.; Matsushika, A.; Yano, S.; Sawayama, S. Bench-scale bioethanol production from eucalyptus by high solid saccharification and glucose/xylose fermentation method. Bioprocess Biosyst. Eng. 2014, 37, 749–754. [Google Scholar] [CrossRef]
- Akita, H.; Goshima, T.; Suzuki, T.; Itoiri, Y.; Kimura, Z.; Matsushika, A. Application of Pichia kudriavzevii NBRC1279 and NBRC1664 to simultaneous saccharification and fermentation for bioethanol production. Fermentation 2021, 7, 83. [Google Scholar] [CrossRef]
- Akita, H.; Yusoff, M.Z.M.; Fujimoto, S. Preparation of oil palm empty fruit bunch hydrolysate. Fermentation 2021, 7, 81. [Google Scholar] [CrossRef]
- Matsushika, A.; Negi, K.; Suzuki, T.; Goshima, T.; Hoshino, T. Identification and characterization of a novel Issatchenkia orientalis GPI-anchored protein, IoGas1, required for resistance to low pH and salt stress. PLoS ONE 2016, 11, e0161888. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, Z.; Li, R.; Miao, Y.; Weng, P.; Wang, L. Integrated transcriptomic and proteomic analysis of the acetic acid stress in Issatchenkia orientalis. J. Food Biochem. 2020, 44, e13203. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Fu, Y.; Deng, N.; Xu, Y. Transcriptional profiling reveals adaptive response and tolerance to lactic acid stress in Pichia kudriavzevii. Foods 2022, 11, 2725. [Google Scholar] [CrossRef] [PubMed]
- Matsushika, A.; Oguri, E.; Sawayama, S. Evolutionary adaptation of recombinant shochu yeast for improved xylose utilization. J. Biosci. Bioeng. 2010, 110, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Matsushika, A.; Suzuki, T.; Goshima, T.; Hoshino, T. Evaluation of Saccharomyces cerevisiae GAS1 with respect to its involvement in tolerance to low pH and salt stress. J. Biosci. Bioeng. 2017, 124, 164–170. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Soneson, C.; Delorenzi, M. A comparison of methods for differential expression analysis of RNA-seq data. BMC Bioinform. 2013, 14, 91. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Chen, R.E.; Thorner, J. Function and regulation in MAPK signaling pathways: Lessons learned from the yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta 2007, 1773, 1311–1340. [Google Scholar] [CrossRef]
- Furukawa, K.; Hohmann, S. Synthetic biology: Lessons from engineering yeast MAPK signalling pathways. Mol. Microbiol. 2013, 88, 5–19. [Google Scholar] [CrossRef]
- Sakihama, Y.; Hasunuma, T.; Kondo, A. Improved ethanol production from xylose in the presence of acetic acid by the overexpression of the HAA1 gene in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2015, 119, 297–302. [Google Scholar] [CrossRef]
- Inaba, T.; Watanabe, D.; Yoshiyama, Y.; Tanaka, K.; Ogawa, J.; Takagi, H.; Shimoi, H.; Shima, J. An organic acid-tolerant HAA1-overexpression mutant of an industrial bioethanol strain of Saccharomyces cerevisiae and its application to the production of bioethanol from sugarcane molasses. AMB Express 2013, 3, 74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.M.; Zhao, X.Q.; Cheng, C.; Bai, F.W. Improved growth and ethanol fermentation of Saccharomyces cerevisiae in the presence of acetic acid by overexpression of SET5 and PPR1. Biotechnol. J. 2015, 10, 1903–1911. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, B.M.; Barrio, E.; Querol, A.; Pérez-Torrado, R. Enhanced enzymatic activity of glycerol-3-phosphate dehydrogenase from the cryophilic Saccharomyces kudriavzevii. PLoS ONE 2014, 9, e87290. [Google Scholar] [CrossRef]
- Remize, F.; Roustan, J.L.; Sablayrolles, J.M.; Barre, P.; Dequin, S. Glycerol overproduction by engineered Saccharomyces cerevisiae wine yeast strains leads to substantial changes in by-product formation and to a stimulation of fermentation rate in stationary phase. Appl. Environ. Microbiol. 1999, 65, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Cambon, B.; Monteil, V.; Remize, F.; Camarasa, C.; Dequin, S. Effects of GPD1 overexpression in Saccharomyces cerevisiae commercial wine yeast strains lacking ALD6 genes. Appl. Environ. Microbiol. 2006, 72, 4688–4694. [Google Scholar] [CrossRef]
- Michnick, S.; Roustan, J.L.; Remize, F.; Barre, P.; Dequin, S. Modulation of glycerol and ethanol yields during alcoholic fermentation in Saccharomyces cerevisiae strains overexpressed or disrupted for GPD1 encoding glycerol 3-phosphate dehydrogenase. Yeast 1997, 13, 783–793. [Google Scholar] [CrossRef]
- Fujii, T.; Inoue, H.; Ishikawa, K.; Hoshino, T. Deletion analysis of GH7 endoglucanase gene (cel7B) promoter region in a Talaromyces cellulolyticus ligD-disrupted strain. Appl. Biochem. Biotechnol. 2017, 183, 1516–1525. [Google Scholar] [CrossRef]
- Yuzawa, T.; Shirai, T.; Orishimo, R.; Kawai, K.; Kondo, A.; Hirasawa, T. 13C-metabolic flux analysis in glycerol-assimilating strains of Saccharomyces cerevisiae. J. Gen. Appl. Microbiol. 2021, 67, 142–149. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast | 15 mM Formic Acid | 35 mM Sulfuric Acid | 60 mM Hydrochloric Acid | 100 mM Acetic Acid | 550 mM Lactic Acid |
|---|---|---|---|---|---|
| S. cerevisiae BY4742 | 0.0028 | 0.0080 | 0.0040 | 0.0064 | 0.0010 |
| P. kudriavzevii NBRC1279 | 0.243 | 0.0049 | 0.00570 | 0.161 | 0.138 |
| P. kudriavzevii NBRC1664 | 0.288 | 0.0177 | 0.0945 | 0.520 | 0.132 |
| S. cerevisiae BY4742 vs. P. kudriavzevii NBRC1279 | S. cerevisiae BY4742 vs. P. kudriavzevii NBRC1664 | ||||
|---|---|---|---|---|---|
| Gene | Description | Regulation | log2 (Fold Change) | Regulation | log2 (Fold Change) |
| Hkr1 | Signaling mucin HKR1 | Up | 7.27 | Up | 4.73 |
| Sho1 | Osmosensor SHO1 | Up | 4.75 | Up | 3.39 |
| Opy2 | Protein OPY2 | Down | −11.5 | Down | −11.5 |
| Cdc42 | Cell division control protein 42 | Down | −11.3 | Down | −11.3 |
| Cdc24 | Cell division control protein 24 | Down | −10.3 | Down | −11.2 |
| Ste20 | p21-Activated kinase1 | Up | 5.53 | Up | 4.97 |
| Cla4 | Serine/threonine-protein kinase CLA4 | Up | 6.07 | Down | −10.4 |
| Ste11 | Mitogen-activated protein kinase kinase kinase | Up | 5.85 | Up | 5.01 |
| Ste50 | Protein STE50 | Up | 5.21 | Up | 4.43 |
| Pbs2 | Mitogen-activated protein kinase kinase | Up | 10.1 | Up | 9.06 |
| Hog1 | Mitogen-activated protein kinase HOG1 | Up | 6.55 | Up | 5.97 |
| Sln1 | Sensor histidine kinase SLN1 | Up | 5.49 | Up | 4.75 |
| Ypd1 | Phosphorelay intermediate protein YPD1 | Up | 5.54 | Up | 4.60 |
| Ssk1 | Mitogen-activated protein kinase kinase kinase SSK1 | Up | 7.38 | Up | 6.56 |
| Ssk2 | Mitogen-activated protein kinase kinase kinase SSK2 | Up | 6.46 | Up | 5.68 |
| Ssk22 | Mitogen-activated protein kinase kinase kinase SSK22 | Down | −10.3 | Down | −10.3 |
| Ptp2 | Tyrosine protein phosphatase PTP2 | Down | −13.0 | Down | −13.0 |
| Ptp3 | Tyrosine protein phosphatase PTP3 | Down | −9.93 | Down | −9.93 |
| Hot1 | High-osmolarity-induced transcription protein 1 | Down | −10.1 | Down | −10.1 |
| Smp1 | Transcription factor SMP1 | Up | 6.38 | Up | 5.51 |
| Gpd1 | NAD+-dependent glycerol-3-phosphate dehydrogenase | Up | 10.4 | Up | 13.1 |
| S. cerevisiae BY4742 vs. P. kudriavzevii NBRC1279 | S. cerevisiae BY4742 vs. P. kudriavzevii NBRC1664 | ||||
|---|---|---|---|---|---|
| Gene | Description | Regulation | log2 (Fold Change) | Regulation | log2 (Fold Change) |
| HAA1 | Transcriptional activator | Down | −11.9 | Down | −11.9 |
| PPR1 | Transcriptional activator | Down | −10.3 | Down | −9.1 |
| SET1 | Histone methyltransferase | Down | −10.2 | Down | −10.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akita, H.; Matsushika, A. Transcription Analysis of the Acid Tolerance Mechanism of Pichia kudriavzevii NBRC1279 and NBRC1664. Fermentation 2023, 9, 559. https://doi.org/10.3390/fermentation9060559
Akita H, Matsushika A. Transcription Analysis of the Acid Tolerance Mechanism of Pichia kudriavzevii NBRC1279 and NBRC1664. Fermentation. 2023; 9(6):559. https://doi.org/10.3390/fermentation9060559
Chicago/Turabian StyleAkita, Hironaga, and Akinori Matsushika. 2023. "Transcription Analysis of the Acid Tolerance Mechanism of Pichia kudriavzevii NBRC1279 and NBRC1664" Fermentation 9, no. 6: 559. https://doi.org/10.3390/fermentation9060559
APA StyleAkita, H., & Matsushika, A. (2023). Transcription Analysis of the Acid Tolerance Mechanism of Pichia kudriavzevii NBRC1279 and NBRC1664. Fermentation, 9(6), 559. https://doi.org/10.3390/fermentation9060559