

Optimization of Milbemycin Component Ratio by Coordinating Acyl-Coenzyme A Supply Pathways in Streptomyces bingchenggensis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, and Culture Conditions
2.2. Gene Cloning and Plasmid Construction
2.3. Detection of Acyl-CoAs of S. bingchenggensis
2.4. Sequence Analysis
2.5. Time–Course Transcriptome Data Analysis
2.6. Determination of Cell Dry Weight
2.7. Detection of Milbemycins
2.8. Statistical Analysis
3. Results
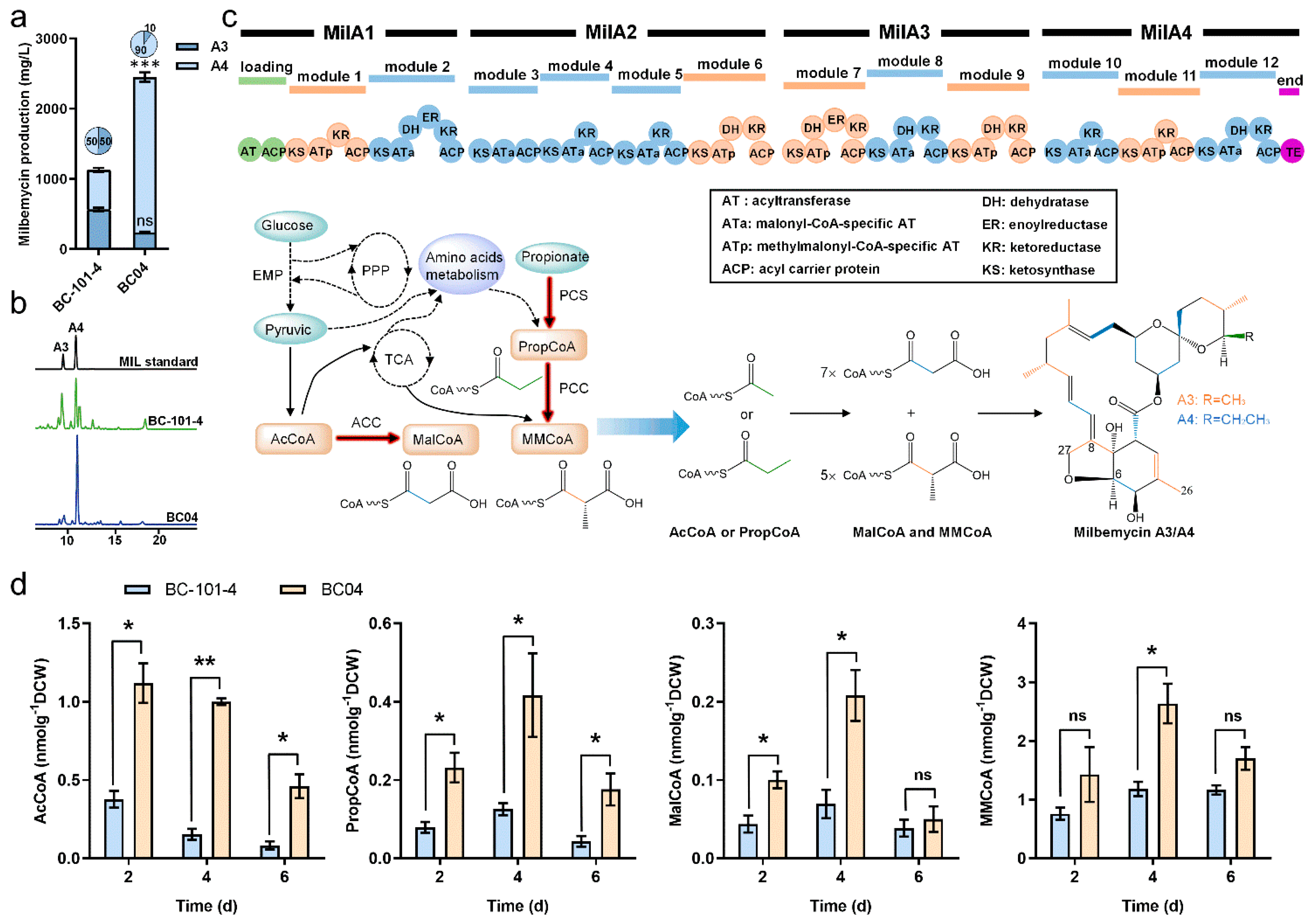
3.1. Analysis of Key Precursors of Milbemycin Biosynthesis
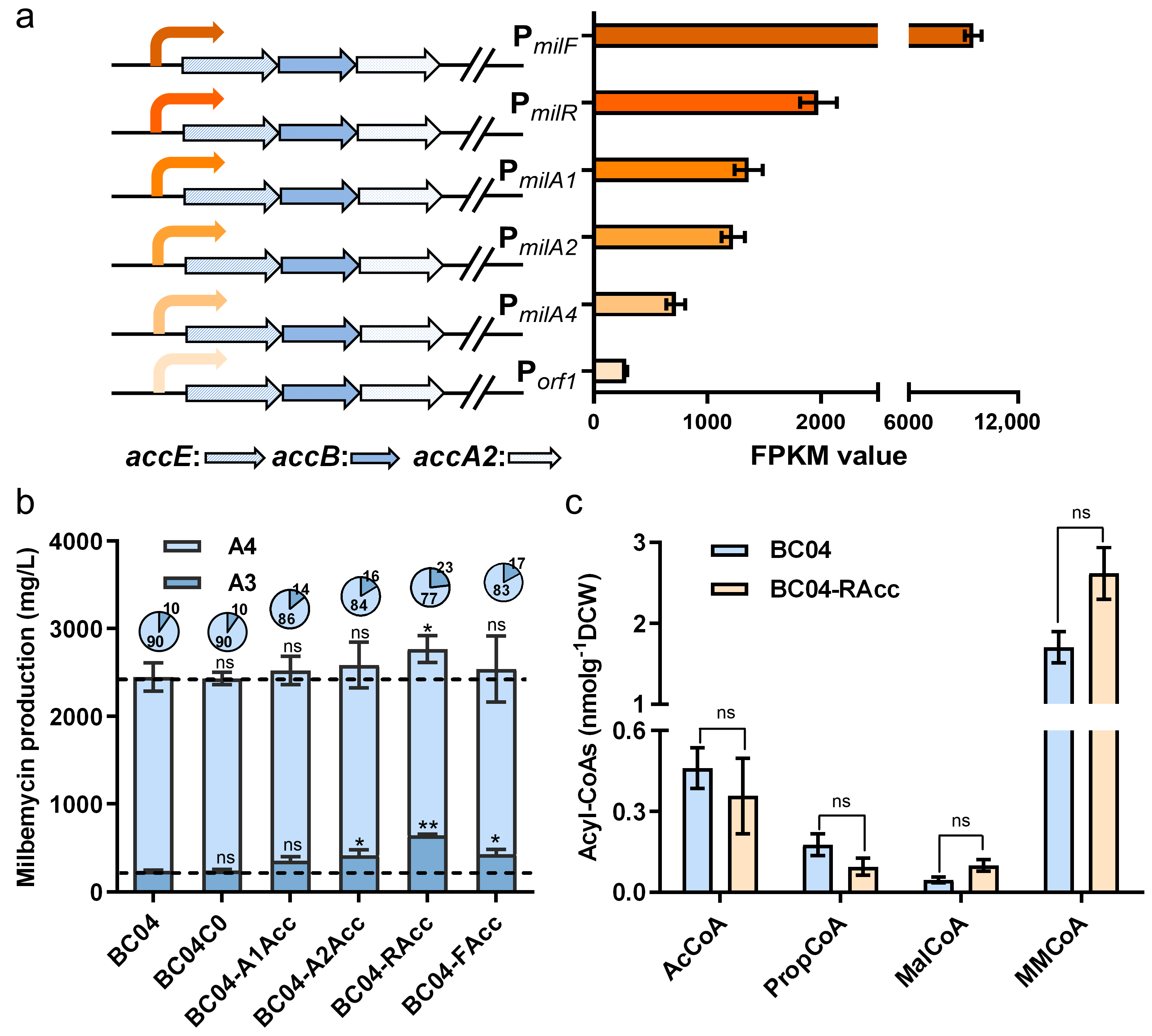
3.2. Fine-Tuning of MalCoA Precursor Supply
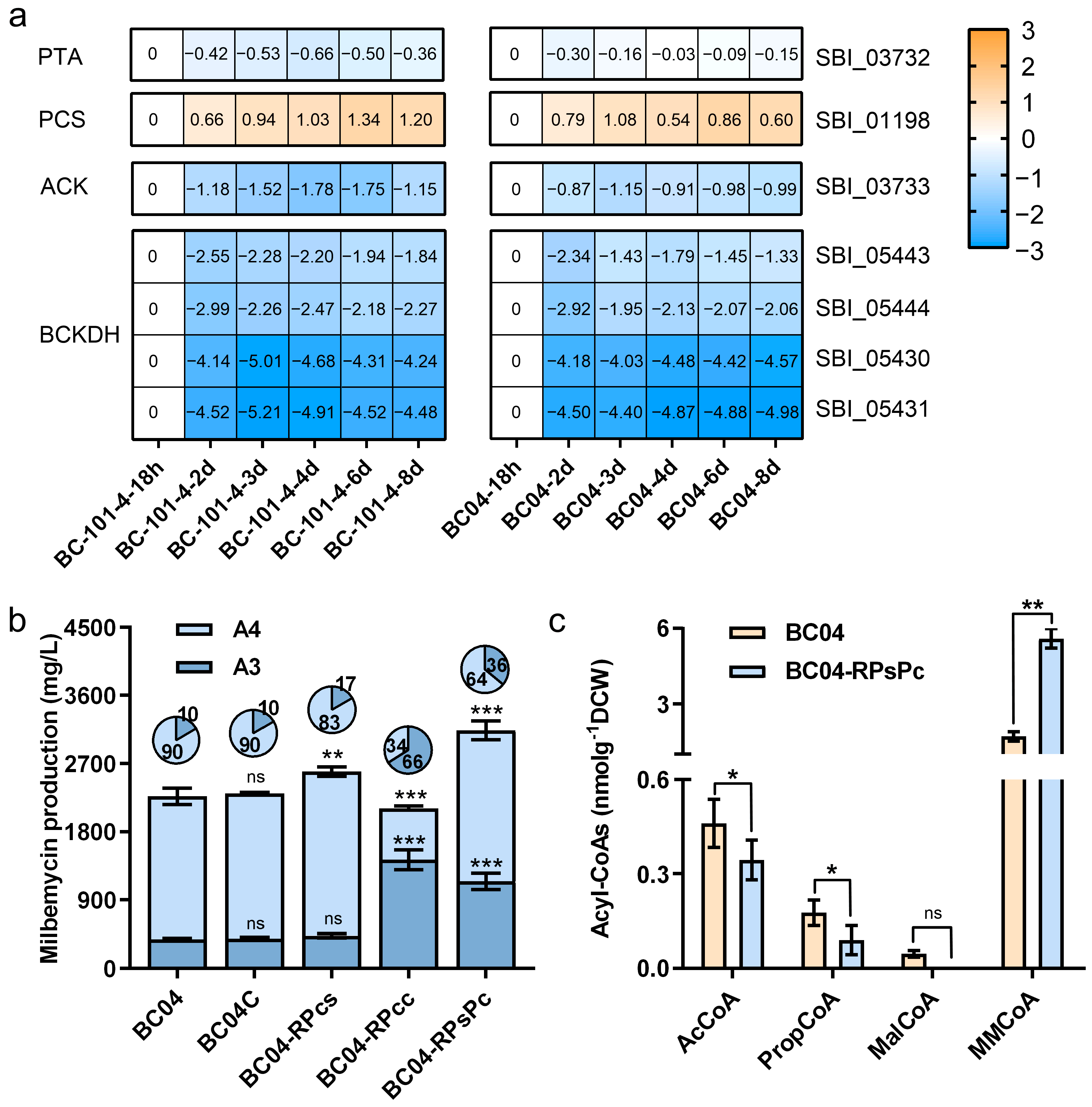
3.3. Engineering the PropCoA and MMCoA Supply Pathway
3.4. Fine-Tuning of PCC Expression Level
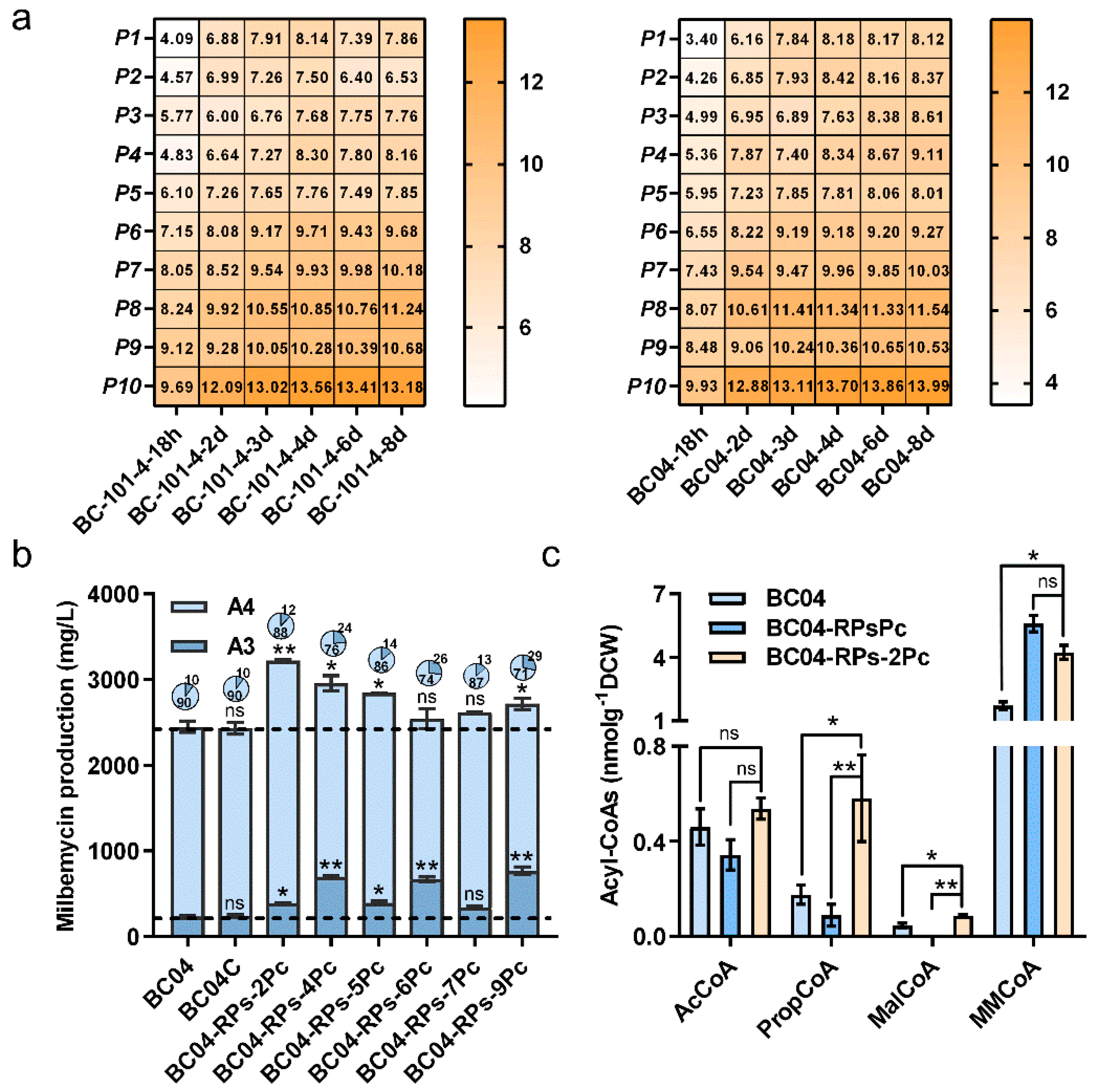
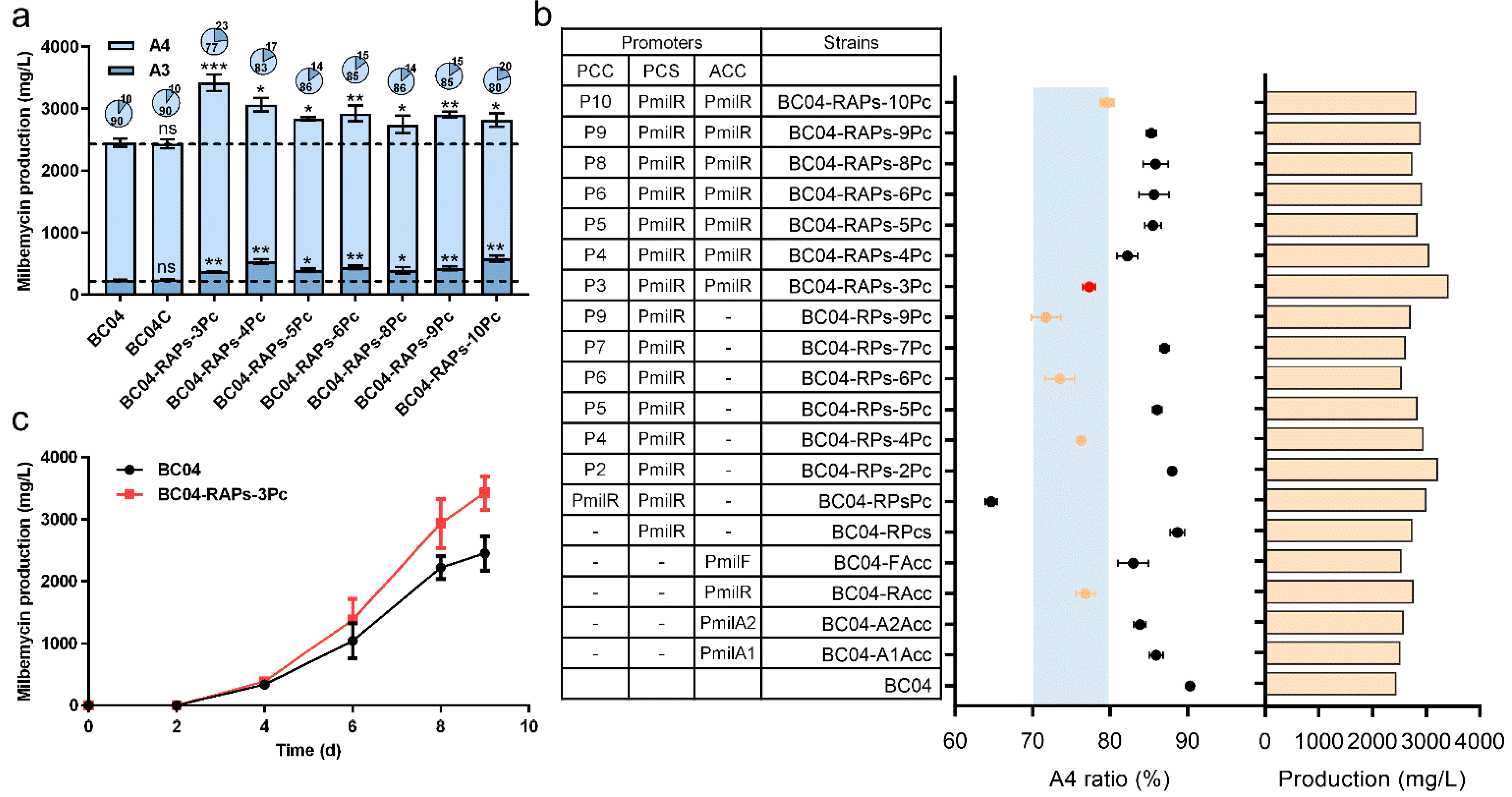
3.5. System Modulation of Precursor Supply for Improved Ratio of Milbemycin A3 and A4
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.; Tan, H. Molecular regulation of antibiotic biosynthesis in Streptomyces. Microbiol. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, B.; Tan, G.Y.; Ouyang, L.M.; Qiu, S.; Wang, W.; Xiang, W.; Zhang, L. Polyketide pesticides from actinomycetes. Curr. Opin. Biotechnol. 2021, 69, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, H.; Nonomiya, T.; Usami, M.; Ohta, T.; Omura, S. Organization of the biosynthetic gene cluster for the polyketide anthelmintic macrolide avermectin in Streptomyces avermitilis. Proc. Natl. Acad. Sci. USA 1999, 96, 9509–9514. [Google Scholar] [CrossRef] [PubMed]
- Hertweck, C. The biosynthetic logic of polyketide diversity. Angew. Chem. Int. Ed. Engl. 2009, 48, 4688–4716. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zhang, X.; Jiang, M.; Bai, L. Enhanced salinomycin production by adjusting the supply of polyketide extender units in Streptomyces albus. Metab. Eng. 2016, 35, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Takiguchi, Y.; Mishima, H.; Okuda, M.; Terao, M.; Aoki, A.; Fukuda, R. Milbemycins, a new family of macrolide antibiotics: Fermentation, isolation and physico-chemical properties. J. Antibiot. 1980, 33, 1120–1127. [Google Scholar] [CrossRef]
- Kim, M.S.; Cho, W.J.; Song, M.C.; Park, S.W.; Kim, K.; Kim, E.; Lee, N.; Nam, S.J.; Oh, K.H.; Yoon, Y.J. Engineered biosynthesis of milbemycins in the avermectin high-producing strain Streptomyces avermitilis. Microb. Cell Factories 2017, 16, 9. [Google Scholar] [CrossRef]
- Campbell, W.C. History of avermectin and ivermectin, with notes on the history of other macrocyclic lactone antiparasitic agents. Curr. Pharm. Biotechnol. 2012, 13, 853–865. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, X.; Ji, M.; Cheng, J.; Zhu, G. Design, synthesis, and biological activities of milbemycin analogues. J. Agric. Food Chem. 2011, 59, 4836–4850. [Google Scholar] [CrossRef]
- Zhang, Y.; He, H.; Liu, H.; Wang, H.; Wang, X.; Xiang, W. Characterization of a pathway-specific activator of milbemycin biosynthesis and improved milbemycin production by its overexpression in Streptomyces bingchenggensis. Microb. Cell Factories 2016, 15, 152. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Xia, H. Recent advances in the research of milbemycin biosynthesis and regulation as well as strategies for strain improvement. Arch. Microbiol. 2021, 203, 5849–5857. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Xiang, W. Improvement of milbemycin-producing Streptomyces bingchenggensis by rational screening of ultraviolet- and chemically induced mutants. World J. Microbiol. Biotechnol. 2009, 25, 1051–1056. [Google Scholar] [CrossRef]
- Xiang, W.; Wang, J.; Wang, X.; Zhang, J. Two new beta-class milbemycins from Streptomyces bingchenggensis: Fermentation, isolation, structure elucidation and biological properties. J. Antibiot. 2007, 60, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, J.; Zhang, Y.; Zhang, B.; Liu, C.; He, H.; Wang, X.; Xiang, W. Combined application of plasma mutagenesis and gene engineering leads to 5-oxomilbemycins A3/A4 as main components from Streptomyces bingchenggensis. Appl. Microbiol. Biotechnol. 2014, 98, 9703–9712. [Google Scholar] [CrossRef]
- He, H.; Ye, L.; Li, C.; Wang, H.; Guo, X.; Wang, X.; Zhang, Y.; Xiang, W. SbbR/SbbA, an Important ArpA/AfsA-Like System, Regulates Milbemycin Production in Streptomyces bingchenggensis. Front. Microbiol. 2018, 9, 1064. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Y.; Li, S.; Ye, L.; Wang, X.; Xiang, W. SspH, a Novel HATPase Family Regulator, Controls Antibiotic Biosynthesis in Streptomyces. Antibiotics 2022, 11, 538. [Google Scholar] [CrossRef]
- Ye, L.; Zhang, Y.; Li, S.; He, H.; Ai, G.; Wang, X.; Xiang, W. Transcriptome-guided identification of a four-component system, SbrH1-R, that modulates milbemycin biosynthesis by influencing gene cluster expression, precursor supply, and antibiotic efflux. Synth. Syst. Biotechnol. 2022, 7, 705–717. [Google Scholar] [CrossRef]
- Jin, P.; Li, S.; Zhang, Y.; Chu, L.; He, H.; Dong, Z.; Xiang, W. Mining and fine-tuning sugar uptake system for titer improvement of milbemycins in Streptomyces bingchenggensis. Synth. Syst. Biotechnol. 2020, 5, 214–221. [Google Scholar] [CrossRef]
- Chu, L.; Li, S.; Dong, Z.; Zhang, Y.; Jin, P.; Ye, L.; Wang, X.; Xiang, W. Mining and engineering exporters for titer improvement of macrolide biopesticides in Streptomyces. Microb. Biotechnol. 2022, 15, 1120–1132. [Google Scholar] [CrossRef]
- Sambrock, J.; Russel, D.J.I. Molecular Cloning: A Laboratory Manual; Cold Spring Harbour Laboratory Press: New York, NY, USA, 2001; Volume 49, pp. 895–909. [Google Scholar]
- Keiser, T.; Bipp, M.; Buttner, M.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; The John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Bierman, M.; Logan, R.; O’Brien, K.; Seno, E.T.; Rao, R.N.; Schoner, B.E. Plasmid cloning vectors for the conjugal transfer of DNA from Escherichia coli to Streptomyces spp. Gene 1992, 116, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Peyraud, R.; Kiefer, P.; Christen, P.; Massou, S.; Portais, J.C.; Vorholt, J.A. Demonstration of the ethylmalonyl-CoA pathway by using 13C metabolomics. Proc. Natl. Acad. Sci. USA 2009, 106, 4846–4851. [Google Scholar] [CrossRef] [PubMed]
- Vance, D.E.; Vance, J.E. Biochemistry of Lipids, Lipoproteins and Membranes; Elsevier Science: New York, NY, USA, 2002. [Google Scholar]
- Ryu, Y.G.; Butler, M.J.; Chater, K.F.; Lee, K.J. Engineering of primary carbohydrate metabolism for increased production of actinorhodin in Streptomyces coelicolor. Appl. Environ. Microbiol. 2006, 72, 7132–7139. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Li, Y.; Guan, H.; Li, J.; Li, D.; Zhang, J.; Tan, H. Component Optimization of Neomycin Biosynthesis via the Reconstitution of a Combinatorial Mini-Gene-Cluster in Streptomyces fradiae. ACS Synth. Biol. 2020, 9, 2493–2501. [Google Scholar] [CrossRef] [PubMed]
- Siegl, T.; Tokovenko, B.; Myronovskyi, M.; Luzhetskyy, A. Design, construction and characterisation of a synthetic promoter library for fine-tuned gene expression in actinomycetes. Metab. Eng. 2013, 19, 98–106. [Google Scholar] [CrossRef]
- Cropp, A.; Chen, S.; Liu, H.; Zhang, W.; Reynolds, K.A. Genetic approaches for controlling ratios of related polyketide products in fermentation processes. J. Ind. Microbiol. Biotechnol. 2001, 27, 368–377. [Google Scholar] [CrossRef]
- Milke, L.; Marienhagen, J. Engineering intracellular malonyl-CoA availability in microbial hosts and its impact on polyketide and fatty acid synthesis. Appl. Microbiol. Biotechnol. 2020, 104, 6057–6065. [Google Scholar] [CrossRef]
- Zabala, D.; Braña, A.F.; Flórez, A.B.; Salas, J.A.; Méndez, C. Engineering precursor metabolite pools for increasing production of antitumor mithramycins in Streptomyces argillaceus. Metab. Eng. 2013, 20, 187–197. [Google Scholar] [CrossRef]
- Gao, Q.; Tan, G.Y.; Xia, X.; Zhang, L. Learn from microbial intelligence for avermectins overproduction. Curr. Opin. Biotechnol. 2017, 48, 251–257. [Google Scholar] [CrossRef]
- Bilyk, O.; Luzhetskyy, A. Metabolic engineering of natural product biosynthesis in actinobacteria. Curr. Opin. Biotechnol. 2016, 42, 98–107. [Google Scholar] [CrossRef]
- Mo, S.; Lee, S.K.; Jin, Y.; Oh, C.; Suh, J.W. Application of a combined approach involving classical random mutagenesis and metabolic engineering to enhance FK506 production in Streptomyces sp. RM7011. Appl. Microbiol. Biotechnol. 2013, 97, 3053–3062. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.S.; Yoo, Y.J.; Park, J.W.; Park, S.R.; Han, A.R.; Ban, Y.H.; Kim, E.J.; Kim, E.; Yoon, Y.J. A combined approach of classical mutagenesis and rational metabolic engineering improves rapamycin biosynthesis and provides insights into methylmalonyl-CoA precursor supply pathway in Streptomyces hygroscopicus ATCC 29253. Appl. Microbiol. Biotechnol. 2011, 91, 1389–1397. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Jin, P.; Dong, Z.; Zhang, Y.; Xiang, W.; Li, S. Optimization of Milbemycin Component Ratio by Coordinating Acyl-Coenzyme A Supply Pathways in Streptomyces bingchenggensis. Fermentation 2023, 9, 555. https://doi.org/10.3390/fermentation9060555
Yang X, Jin P, Dong Z, Zhang Y, Xiang W, Li S. Optimization of Milbemycin Component Ratio by Coordinating Acyl-Coenzyme A Supply Pathways in Streptomyces bingchenggensis. Fermentation. 2023; 9(6):555. https://doi.org/10.3390/fermentation9060555
Chicago/Turabian StyleYang, Xue, Pinjiao Jin, Zhuoxu Dong, Yanyan Zhang, Wensheng Xiang, and Shanshan Li. 2023. "Optimization of Milbemycin Component Ratio by Coordinating Acyl-Coenzyme A Supply Pathways in Streptomyces bingchenggensis" Fermentation 9, no. 6: 555. https://doi.org/10.3390/fermentation9060555
APA StyleYang, X., Jin, P., Dong, Z., Zhang, Y., Xiang, W., & Li, S. (2023). Optimization of Milbemycin Component Ratio by Coordinating Acyl-Coenzyme A Supply Pathways in Streptomyces bingchenggensis. Fermentation, 9(6), 555. https://doi.org/10.3390/fermentation9060555