
Biosynthesis of Gamma-Aminobutyric Acid by Engineered Clostridium tyrobutyricum Co-Overexpressing Glutamate Decarboxylase and Class I Heat Shock Protein

Abstract
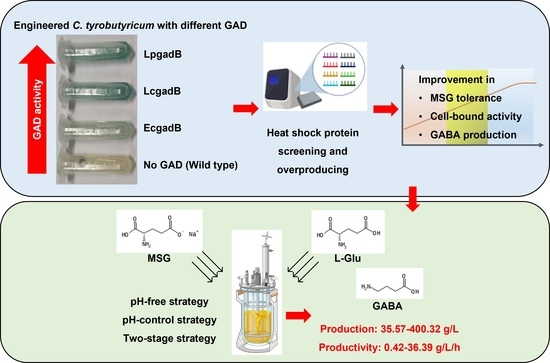
1. Introduction
2. Materials and Methods
2.1. Organism, Plasmids, and Culture Conditions
2.2. Construction of the Engineered C. tyrobutyricum
2.3. Quantitative Real-Time PCR and Relative-Gene-Expression Analysis
2.4. GAD-Enzyme-Activity and Cell-Bound-Activity Analysis
2.5. MSG-Tolerance Test
2.6. Fermentation Experiments
3. Results
3.1. Amino-Acid-Sequence Analysis and Enzymatic-Property Comparison of Putative GADs
3.2. Production of GABA by Engineered C. tyrobutyricum
3.3. Enhanced MSG Tolerance and GABA Production by Class I Heat-Shock Protein GroESL Overexpression
3.4. Production of GABA in Fed-Batch Fermentation
3.5. Improvement of GABA Biosynthesis by Two-Stage Strategy Using Glutamate Acid (L-Glu) as Substrate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luo, H.; Liu, Z.; Xie, F.; Bilal, M.; Liu, L.; Yang, R.; Wang, Z. Microbial production of gamma-aminobutyric acid: Applications, state-of-the-art achievements, and future perspectives. Crit. Rev. Biotechnol. 2021, 41, 491–512. [Google Scholar] [CrossRef] [PubMed]
- Pannerchelvan, S.; Rios-Solis, L.; Wong, F.W.F.; Zaidan, U.H.; Wasoh, H.; Mohamed, M.S.; Tan, J.S.; Mohamad, R.; Halim, M. Strategies for improvement of gamma-aminobutyric acid (GABA) biosynthesis via lactic acid bacteria (LAB) fermentation. Food Funct. 2023. [Google Scholar] [CrossRef] [PubMed]
- Heli, Z.; Hongyu, C.; Dapeng, B.; Yee Shin, T.; Yejun, Z.; Xi, Z.; Yingying, W. Recent advances of γ-aminobutyric acid: Physiological and immunity function, enrichment, and metabolic pathway. Front. Nutr. 2022, 9, 1076223. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Ma, Y.; Wei, Z.-Z.; Yuan, W.-X.; Li, Y.-l.; Zhang, C.-H.; Xue, X.-T.; Zhou, H.-J. Determination and Comparison of γ-Aminobutyric Acid (GABA) Content in Pu-erh and Other Types of Chinese Tea. J. Agric. Food Chem. 2011, 59, 3641–3648. [Google Scholar] [CrossRef]
- Koubaa, M.; Delbecq, F.; Roohinejad, S.; Mallikarjunan, K. Gamma-Aminobutyric Acid. In Encyclopedia of Food Chemistry; Melton, L., Shahidi, F., Varelis, P., Eds.; Academic Press: Oxford, UK, 2019; pp. 528–534. [Google Scholar]
- Boonstra, E.; de Kleijn, R.; Colzato, L.S.; Alkemade, A.; Forstmann, B.U.; Nieuwenhuis, S. Neurotransmitters as food supplements: The effects of GABA on brain and behavior. Front. Psychol. 2015, 6, 1520. [Google Scholar] [CrossRef]
- Sarasa, S.B.; Mahendran, R.; Muthusamy, G.; Thankappan, B.; Selta, D.R.F.; Angayarkanni, J. A Brief Review on the Non-protein Amino Acid, Gamma-amino Butyric Acid (GABA): Its Production and Role in Microbes. Curr. Microbiol. 2020, 77, 534–544. [Google Scholar] [CrossRef]
- Zhang, R.; Yang, T.; Rao, Z.; Sun, H.; Xu, M.; Zhang, X.; Xu, Z.; Yang, S. Efficient one-step preparation of γ-aminobutyric acid from glucose without an exogenous cofactor by the designed Corynebacterium glutamicum. Green Chem. 2014, 16, 4190–4197. [Google Scholar] [CrossRef]
- Soma, Y.; Fujiwara, Y.; Nakagawa, T.; Tsuruno, K.; Hanai, T. Reconstruction of a metabolic regulatory network in Escherichia coli for purposeful switching from cell growth mode to production mode in direct GABA fermentation from glucose. Metab. Eng. 2017, 43, 54–63. [Google Scholar] [CrossRef]
- Lyu, C.J.; Liu, L.; Huang, J.; Zhao, W.R.; Hu, S.; Mei, L.H.; Yao, S.J. Biosynthesis of gamma-aminobutyrate by engineered Lactobacillus brevis cells immobilized in gellan gum gel beads. J. Biosci. Bioeng. 2019, 128, 123–128. [Google Scholar] [CrossRef]
- Cui, Y.; Miao, K.; Niyaphorn, S.; Qu, X. Production of Gamma-Aminobutyric Acid from Lactic Acid Bacteria: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 995. [Google Scholar] [CrossRef]
- Gong, L.; Ren, C.; Xu, Y. GlnR Negatively Regulates Glutamate-Dependent Acid Resistance in Lactobacillus brevis. Appl. Environ. Microbiol. 2020, 86, e02615–e02619. [Google Scholar] [CrossRef]
- Li, H.; Qiu, T.; Gao, D.; Cao, Y. Medium optimization for production of gamma-aminobutyric acid by Lactobacillus brevis NCL912. Amino Acids 2010, 38, 1439–1445. [Google Scholar] [CrossRef]
- Lyu, C.; Yao, L.; Zhu, Q.; Mei, J.; Cao, Y.; Hu, S.; Zhao, W.; Huang, J.; Mei, L.; Yao, S.; et al. Reconstruction of the glutamate decarboxylase system in Lactococcus lactis for biosynthesis of food-grade gamma-aminobutyric acid. Appl. Microbiol. Biotechnol. 2021, 105, 4127–4140. [Google Scholar] [CrossRef]
- Wei, L.; Zhao, J.; Wang, Y.; Gao, J.; Du, M.; Zhang, Y.; Xu, N.; Du, H.; Ju, J.; Liu, Q.; et al. Engineering of Corynebacterium glutamicum for high-level gamma-aminobutyric acid production from glycerol by dynamic metabolic control. Metab. Eng. 2022, 69, 134–146. [Google Scholar] [CrossRef]
- Yang, X.; Ke, C.; Zhu, J.; Wang, Y.; Zeng, W.; Huang, J. Enhanced productivity of gamma-amino butyric acid by cascade modifications of a whole-cell biocatalyst. Appl. Microbiol. Biotechnol. 2018, 102, 3623–3633. [Google Scholar] [CrossRef]
- Gong, L.; Ren, C.; Xu, Y. Deciphering the crucial roles of transcriptional regulator GadR on gamma-aminobutyric acid production and acid resistance in Lactobacillus brevis. Microb. Cell Fact. 2019, 18, 108. [Google Scholar] [CrossRef]
- Lyu, C.-J.; Fei, J.-Y.; Yan, J.-P.; Xu, Q.-C.; Mei, J.-Q.; Yue, H.-Y.; Chen, L.-L.; Mei, L.-H.; Huang, J.; Yao, S.-J. Improvement of γ-aminobutyrate biosynthesis by genetically engineered Lactococcus lactis. Biochem. Eng. J. 2020, 157, 107525. [Google Scholar] [CrossRef]
- Altaib, H.; Kozakai, T.; Badr, Y.; Nakao, H.; El-Nouby, M.A.M.; Yanase, E.; Nomura, I.; Suzuki, T. Cell factory for gamma-aminobutyric acid (GABA) production using Bifidobacterium adolescentis. Microb. Cell Fact. 2022, 21, 33. [Google Scholar] [CrossRef]
- Lee, K.W.; Shim, J.M.; Yao, Z.; Kim, J.A.; Kim, H.J.; Kim, J.H. Characterization of a glutamate decarboxylase (GAD) from Enterococcus avium M5 isolated from Jeotgal, a Korean fermented seafood. J. Microbiol. Biotechnol. 2017, 27, 1216–1222. [Google Scholar] [CrossRef]
- Yang, S.-Y.; Lin, Q.; Lu, Z.-X.; Lü, F.-X.; Bie, X.-M.; Zou, X.-K.; Sun, L.-J. Characterization of a novel glutamate decarboxylase from Streptococcus salivarius ssp. thermophilus Y2. J. Chem. Technol. Biotechnol. 2008, 83, 855–861. [Google Scholar] [CrossRef]
- Cataldo, P.G.; Villegas, J.M.; Savoy de Giori, G.; Saavedra, L.; Hebert, E.M. Enhancement of gamma-aminobutyric acid (GABA) production by Lactobacillus brevis CRL 2013 based on carbohydrate fermentation. Int. J. Food Microbiol. 2020, 333, 108792. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, S.S.; Gonzalez, R. Anaerobic fermentation of glycerol: A path to economic viability for the biofuels industry. Curr. Opin. Biotechnol. 2007, 18, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Faraone, I.; Giavalisco, M.; Parente, E.; Lela, L.; Storti, L.V.; Ricciardi, A. The Production of γ-Aminobutyric Acid from Free and Immobilized Cells of Levilactobacillus brevis Cultivated in Anaerobic and Aerobic Conditions. Microorganisms 2022, 10, 2184. [Google Scholar] [CrossRef] [PubMed]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. γ-Aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Fu, H.; Yang, H.K.; Xu, W.; Wang, J.; Yang, S.T. Butyric acid: Applications and recent advances in its bioproduction. Biotechnol. Adv. 2018, 36, 2101–2117. [Google Scholar] [CrossRef]
- Bao, T.; Feng, J.; Jiang, W.; Fu, H.; Wang, J.; Yang, S.T. Recent advances in n-butanol and butyrate production using engineered Clostridium tyrobutyricum. World J. Microbiol. Biotechnol. 2020, 36, 138. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, H.; Feng, J.; Yang, L.; Luo, K.; Fu, H.; Wang, J. De novo biosynthesis of butyl butyrate in engineered Clostridium tyrobutyricum. Metab. Eng. 2023, 77, 64–75. [Google Scholar] [CrossRef]
- Suo, Y.; Luo, S.; Zhang, Y.; Liao, Z.; Wang, J. Enhanced butyric acid tolerance and production by Class I heat shock protein-overproducing Clostridium tyrobutyricum ATCC 25755. J. Ind. Microbiol. Biotechnol. 2017, 44, 1145–1156. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, J.; Liang, S.; Cai, J.; Xu, Z. Control and optimization of Clostridium tyrobutyricum ATCC 25755 adhesion into fibrous matrix in a fibrous bed bioreactor. Appl. Biochem. Biotechnol. 2011, 165, 98–108. [Google Scholar] [CrossRef]
- Williams, D.R.; Young, D.I.; Young, M. Conjugative plasmid transfer from Escherichia coli to Clostridium acetobutylicum. J. Gen. Microbiol. 1990, 136, 819–826. [Google Scholar] [CrossRef]
- Liu, Q.; Cheng, H.; Ma, X.; Xu, N.; Liu, J.; Ma, Y. Expression, characterization and mutagenesis of a novel glutamate decarboxylase from Bacillus megaterium. Biotechnol. Lett. 2016, 38, 1107–1113. [Google Scholar] [CrossRef]
- Lyu, C.; Zhao, W.; Peng, C.; Hu, S.; Fang, H.; Hua, Y.; Yao, S.; Huang, J.; Mei, L. Exploring the contributions of two glutamate decarboxylase isozymes in Lactobacillus brevis to acid resistance and gamma-aminobutyric acid production. Microb. Cell Fact. 2018, 17, 180. [Google Scholar] [CrossRef]
- De Biase, D.; Pennacchietti, E. Glutamate decarboxylase-dependent acid resistance in orally acquired bacteria: Function, distribution and biomedical implications of the gadBC operon. Mol. Microbiol. 2012, 86, 770–786. [Google Scholar] [CrossRef]
- Le Vo, T.D.; Kim, T.W.; Hong, S.H. Effects of glutamate decarboxylase and gamma-aminobutyric acid (GABA) transporter on the bioconversion of GABA in engineered Escherichia coli. Bioprocess Biosyst. Eng. 2012, 35, 645–650. [Google Scholar] [CrossRef]
- Suo, Y.; Fu, H.; Ren, M.; Liao, Z.; Ma, Y.; Wang, J. Enhanced butyric acid production in Clostridium tyrobutyricum by overexpression of rate-limiting enzymes in the Embden-Meyerhof-Parnas pathway. J. Biotechnol. 2018, 272, 14–21. [Google Scholar] [CrossRef]
- Shi, X.; Chang, C.; Ma, S.; Cheng, Y.; Zhang, J.; Gao, Q. Efficient bioconversion of L-glutamate to gamma-aminobutyric acid by Lactobacillus brevis resting cells. J. Ind. Microbiol. Biotechnol. 2017, 44, 697–704. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, X.; Fu, J.; Wang, S.; Chen, Y.; Chang, K.; Li, H. Substrate sustained release-based high efficacy biosynthesis of GABA by Lactobacillus brevis NCL912. Microb. Cell Fact. 2018, 17, 80. [Google Scholar] [CrossRef]
- Lyu, C.J.; Zhao, W.R.; Hu, S.; Huang, J.; Lu, T.; Jin, Z.H.; Mei, L.H.; Yao, S.J. Physiology-Oriented Engineering Strategy to Improve Gamma-Aminobutyrate Production in Lactobacillus brevis. J. Agric. Food Chem. 2017, 65, 858–866. [Google Scholar] [CrossRef]
- Capitani, G.; De Biase, D.; Aurizi, C.; Gut, H.; Bossa, F.; Grütter, M.G. Crystal structure and functional analysis of Escherichia coli glutamate decarboxylase. EMBO J. 2003, 22, 4027–4037. [Google Scholar] [CrossRef]
- Ke, C.; Yang, X.; Rao, H.; Zeng, W.; Hu, M.; Tao, Y.; Huang, J. Whole-cell conversion of l-glutamic acid into gamma-aminobutyric acid by metabolically engineered Escherichia coli. Springerplus 2016, 5, 591. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M.; Nakajima, I.; Fujita, Y.; Kobayashi, M.; Kimoto, H.; Suzuki, I.; Aso, H. Lactococcus lactis contains only one glutamate decarboxylase gene. Microbiology 1999, 145, 1375–1380. [Google Scholar] [CrossRef] [PubMed]
- Selby, K.; Lindstrom, M.; Somervuo, P.; Heap, J.T.; Minton, N.P.; Korkeala, H. Important role of class I heat shock genes hrcA and dnaK in the heat shock response and the response to pH and NaCl stress of group I Clostridium botulinum strain ATCC 3502. Appl. Environ. Microbiol. 2011, 77, 2823–2830. [Google Scholar] [CrossRef] [PubMed]
- Heap, J.T.; Pennington, O.J.; Cartman, S.T.; Minton, N.P. A modular system for Clostridium shuttle plasmids. J. Microbiol. Methods 2009, 78, 79–85. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sugar Concentration (g/L) | Initial OD600 of Bioconversion Stage | Titer (g/L) | Productivity (g/L/h) | Molar Yield (%) |
|---|---|---|---|---|
| 60 | 24.40 | 277.96 | 5.67/16.35 * | 67.40 |
| 90 | 34.40 | 400.32 | 9.31/36.39 * | 97.07 |
| 120 | 33.4 | 400.55 | 7.85/36.41 * | 97.13 |
| Microorganism | Engineering/Process Strategy | Substrate | Operation Mode and Strategy | Titer (g/L) | Productivity (g/L/h) | References |
|---|---|---|---|---|---|---|
| E. coli BW25113 | ↓lacI, ↓gabT, ↓sucA, ↓aceA (IPTG based dynamic control) | Glucose | Batch, aerobic | 4.8 | 0.15 | [9] |
| E. coli BW25113 | +Lactococcus lactis GAD mutation, removing C-plug of GadC, ↑groESL, ΔgabT | L-Glu | Batch, bioconversion, aerobic | 308.26 | 44.04 * | [16] |
| Lactobacillus brevis NCL912 | Wild type | L-Glu | Batch, aerobic | 205 | 4.8 | [38] |
| Lb. brevis CGMCC1306 | ΔF0F1-ATPase | MSG | Batch, pH-control, aerobic | 43.65 | 0.9 | [39] |
| L. brevis ATCC367 | ΔglnR | MSG | Fed-batch, pH-control, aerobic | 284.7 | 3.95 | [12] |
| Bifidobacterium. adolescentis JCM 1275 | ↑gadBC | MSG | Fed-batch, pH-control, anaerobic | 43.99 | 0.73 | [19] |
| C. glutamicum | +L. plantarum GAD, ΔdapA, ΔargB, ΔproA | Glucose | Fed-batch, pH-control, aerobic | 70.6 | 1 | [8] |
| C. tyrobutyricum ATCC25755 | +L. lactis GAD, ↑groESL | MSG | Fed-batch, pH-free, anerobic | 35.57 | 0.42 | This work |
| C. tyrobutyricum ATCC25755 | +L. lactis GAD, ↑groESL | MSG | Fed-batch, pH-control, anerobic | 122.34 | 1.46 | This work |
| C. tyrobutyricum ATCC25755 | +L. lactis GAD, ↑groESL | L-Glu | Fed-batch, two stage, anerobic | 400.32 | 9.31/36.39 * | This work |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Guo, X.; Dai, K.; Feng, J.; Zhou, T.; Fu, H.; Wang, J. Biosynthesis of Gamma-Aminobutyric Acid by Engineered Clostridium tyrobutyricum Co-Overexpressing Glutamate Decarboxylase and Class I Heat Shock Protein. Fermentation 2023, 9, 445. https://doi.org/10.3390/fermentation9050445
Liu Z, Guo X, Dai K, Feng J, Zhou T, Fu H, Wang J. Biosynthesis of Gamma-Aminobutyric Acid by Engineered Clostridium tyrobutyricum Co-Overexpressing Glutamate Decarboxylase and Class I Heat Shock Protein. Fermentation. 2023; 9(5):445. https://doi.org/10.3390/fermentation9050445
Chicago/Turabian StyleLiu, Ziyao, Xiaolong Guo, Kaiqun Dai, Jun Feng, Tiantian Zhou, Hongxin Fu, and Jufang Wang. 2023. "Biosynthesis of Gamma-Aminobutyric Acid by Engineered Clostridium tyrobutyricum Co-Overexpressing Glutamate Decarboxylase and Class I Heat Shock Protein" Fermentation 9, no. 5: 445. https://doi.org/10.3390/fermentation9050445
APA StyleLiu, Z., Guo, X., Dai, K., Feng, J., Zhou, T., Fu, H., & Wang, J. (2023). Biosynthesis of Gamma-Aminobutyric Acid by Engineered Clostridium tyrobutyricum Co-Overexpressing Glutamate Decarboxylase and Class I Heat Shock Protein. Fermentation, 9(5), 445. https://doi.org/10.3390/fermentation9050445