
The Role of Fermented Dairy Products on Gut Microbiota Composition

 ,
,  ,
,
Abstract
1. Introduction
2. Fermented Milk Products
2.1. Composition of Various Kinds of Milk Used to Produce Fermented Milk Products
2.2. Types of Fermented Milk Products
- 1.
- Products of lactic fermentation, where strains of mesophilic or thermophilic lactic acid bacteria are used (e.g., yogurt).
- 2.
- Products obtained through alcohol-lactic fermentation involving yeast and lactic acid bacteria (e. g., kefir, kumis).
- 3.
- Products with mold growth in addition to the fermentation types above (e.g., viili) [27].
3. Gut Microbiota
3.1. Factors Affecting Variability in Gut Microbiota Composition
- Bacteroides (more represented in enterotype 1),
- Prevotella (more numerous in enterotype 2),
- Ruminococcus (dominant in enterotype 3).
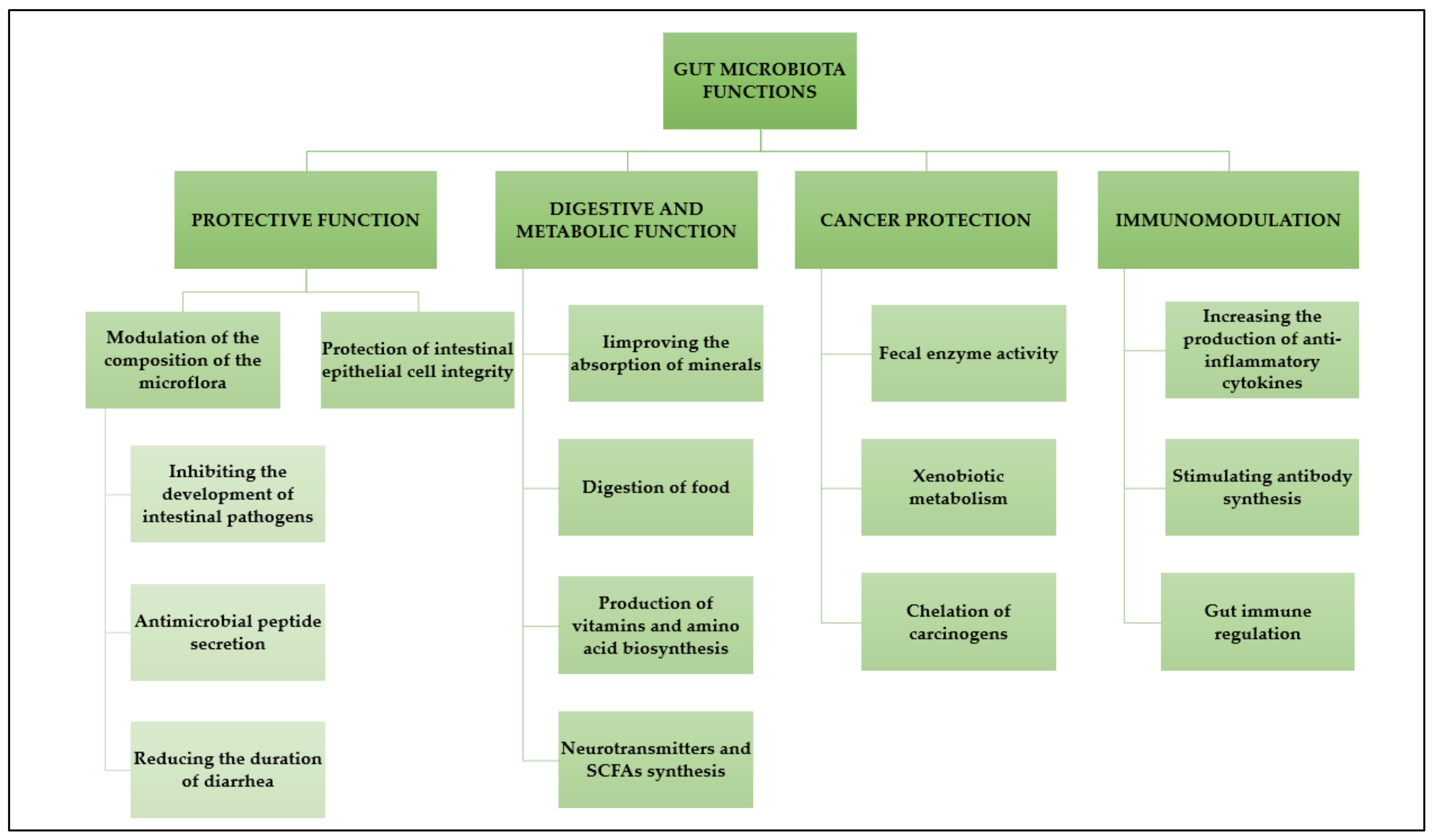
3.2. Main Functions of the Gut Microbiota
- 1.
- Protective function; One of the leading defence mechanisms is the occupation of an ecological niche, which makes it difficult for pathogenic bacteria to reach the intestinal epithelial layer. At the same time, numerous commensal bacteria block receptors are recognized by pathogenic bacteria. An example is Lb. plantarum, which uses mannose receptors for adhesion. The same receptors are necessary for the adhesion of enteropathogenic Escherichia coli strains. Moreover, commensal bacteria compete with pathogens for nutrients and production of compounds with bacteriostatic/bactericidal activity (bacteriocins, organic acids, hydrogen acid, compounds of the lactoperoxidase system, and others), modification of the intestinal environment to make it unfavorable for the development of harmful microorganisms (lowering pH), thereby maintaining the continuity of the gastrointestinal mucosa: stimulating the secretion of mucin “sealing” the intestinal epithelium and production of short-chain fatty acids and polyamines (regeneration of the epithelium and the effect on cell maturation).
- 2.
- Digestive function: Gut microbiota is involved in the digestion of numerous compounds that are otherwise inaccessible to humans, such as cellulose, pectin, or lignin. These compounds are converted into simple sugars or short-chain fatty acids. An interesting example here may be Bifidobacterium longum subs. infantis colonizing the intestines of newborns and breaking down HMO (human milk oligosaccharider) sugars not broken down by human digestive enzymes. However, bacteria provide not only nutrients but also vitamins necessary for humans, such as K, B1, B6, B12, or folic acid.
- 3.
- Immune function and Stimulation of the immune system: Probiotic bacteria do not differ significantly from pathogenic bacteria, and ingredients such as lipopolysaccharide (LPS), peptidoglycan, or lipoteichoic acids are recognized by the TLR (tool-like receptor) in the same way. These receptors are involved in stimulating the immune response by promoting the production of pro-inflammatory cytokines (such as TNF-α or IL-1, 6, 8, 12). The NF-κB transcription factor is also activated, leading to, e.g., production of anti-bacterial proteins (defensins) by enterocytes. Epithelial cells can also produce other anti-bacterial substances, such as lysozyme or phospholipase. Probiotic bacteria have developed several adaptations and interactions with the host organism that allow them to survive and colonize the gastrointestinal tract (e.g., Bifidobacterium longum and Bacteroides thetaiotaomicron together can reduce the expression of genes responsible for fighting gram-positive bacteria. Bifidobacterium bacilli can also inhibit the signal stimulating the production of RegIIIγ lectin, which is a consequence of activation of TLR receptors, and Enterococcus has the ability to induce the expression of genes responsible for the production of IL-10, having an anti-inflammatory effect).
- 4.
- Anti-cancer function: Bacterial enzymes play an important role in carcinogenesis. Probiotic strains can reduce the activity of carcinogens, e.g., the Lb. acidophilus strain causes a decrease in the activity of 1,2-dimethylhydrosine and the Bifidobacterium longum strain reduces the activity of 2-amino-3-methyl-limidazal (4,5-t) choline. Moreover, Lb. casei (LC9018) strains induce immune response mechanisms against cancer cells. In addition, the reduction of hepatic lipogenesis by probiotic strains may be useful in the treatment of cancer. Figure 2 illustrates the functions of the gut microbiota.
3.3. Eubiosis and Dysbiosis
- 1.
- Loss of beneficial organisms (antibiotics),
- 2.
- Excessive growth of potentially harmful organisms (infections, lack of hygiene), and
- 3.
- lLss of overall microbial biodiversity (poor diet).
- food, food additives, and alcohol consumption—unhealthy eating habits negatively affect the composition of the gut microflora and can act as a disease-causing factor impacting metabolic pathways. A high-fat diet and meat are associated with an increased risk of Crohn’s disease (CD) and ulcerative colitis (UC). The risk of inflammatory bowel syndrome (IBS) can be reduced by modulating the structure of the gut microflora and/or its metabolome with a vegetarian diet [95,96,97,98];
- antibiotics and medication—the main consequence of antibiotic treatment is the elimination of sensitive microorganisms (symbiotic bacteria) and the selection and multiplication of dysbiotic bacteria or fungi—primarily pathogenic. This imbalance of the ecosystem can lead to diarrhea due to the pathological proliferation of opportunistic endogenous pathogens, such as Clostridium difficile and vancomycin-resistant enterococci. Moreover, patients treated with antibiotics are more susceptible to infections caused by hexogen pathogens due to the loss of microbiota integrity and barrier function [99],
- age (in people over 70, the number of Bacteroides and Bifidobacterium decreases), gender (the effect of sex hormones), stress (under stress, the bacteria such as Lactobacillus Bacteroides spp. and Clostridium spp. decrease), lifestyle (smoking habits and drug consumption can together contribute to gut dysbiosis),
- gastrointestinal disorders and infections.
4. The Influence of Fermented Milk Products on the Microbiota Composition
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kopp, W. How Western Diet and Lifestyle Drive the Pandemic of Obesity and Civilization Diseases. Diabetes Metab. Syndr. Obes. 2019, 12, 2221–2236. [Google Scholar] [CrossRef]
- Rakhra, V.; Galappaththy, S.L.; Bulchandani, S.; Cabandugama, P.K. Obesity and the Western Diet: How We Got Here. Mol. Med. 2020, 117, 536–538. [Google Scholar]
- Statovci, D.; Aguilera, M.; MacSharry, J.; Melgar, S. The Impact of Western Diet and Nutrients on the Microbiota and Immune Response at Mucosal Interfaces. Front. Immunol. 2017, 8, 838. [Google Scholar] [CrossRef] [PubMed]
- Chiba, M.; Nakane, K.; Komatsu, M. Westernized Diet Is the Most Ubiquitous Environmental Factor in Inflammatory Bowel Disease. Perm. J. 2019, 23, 18–107. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, D.; Hernández-Camba, A.; Ramos, L. Diet and Microbiome in the Beginning of the Sequence of Gut Inflammation. World J. Clin. Cases 2021, 9, 11122–11147. [Google Scholar] [CrossRef]
- González, S.; Fernández-Navarro, T.; Arboleya, S.; de Los Reyes-Gavilán, C.G.; Salazar, N.; Gueimonde, M. Fermented Dairy Foods: Impact on Intestinal Microbiota and Health-Linked Biomarkers. Front. Microbiol. 2019, 10, 1046. [Google Scholar] [CrossRef]
- Leeuwendaal, N.K.; Stanton, C.; O’Toole, P.W.; Beresford, T.P. Fermented Foods, Health and the Gut Microbiome. Nutrients 2022, 14, 1527. [Google Scholar] [CrossRef]
- Szydłowska, A.; Sionek, B. Probiotics and Postbiotics as the Functional Food Components Affecting the Immune Response. Microorganisms 2022, 11, 104. [Google Scholar] [CrossRef]
- Evershed, R.P.; Payne, S.; Sherratt, A.G.; Copley, M.S.; Coolidge, J.; Urem-Kotsu, D.; Kotsakis, K.; Ozdoğan, M.; Ozdoğan, A.E.; Nieuwenhuyse, O.; et al. Earliest Date for Milk Use in the Near East and Southeastern Europe Linked to Cattle Herding. Nature 2008, 455, 528–531. [Google Scholar] [CrossRef]
- Bintsis, T.; Papademas, P. The Evolution of Fermented Milks, from Artisanal to Industrial Products: A Critical Review. Fermentation 2022, 8, 679. [Google Scholar] [CrossRef]
- Sakandar, H.A.; Zhang, H. Curious Case of the History of Fermented Milk: Tangible Evidence. Sci. Bull. 2022, 67, 1625–1627. [Google Scholar] [CrossRef] [PubMed]
- Ségurel, L.; Bon, C. On the Evolution of Lactase Persistence in Humans. Annu. Rev. Genom. Hum. Genet. 2017, 18, 297–319. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Strata, A. Milk, Dairy Products, and Their Functional Effects in Humans: A Narrative Review of Recent Evidence. Adv. Nutr. 2014, 5, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Leciej, D.; Herzig, K.-H.; Thalmann, O. Zoonoses and Their Traces in Ancient Genomes—A Possible Indicator for Ancient Life-Style Changes? JMS 2020, 89, e467. [Google Scholar] [CrossRef]
- Muehlhoff, E.; FAO (Eds.) Milk and Dairy Products in Human Nutrition; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; ISBN 978-92-5-107864-8. [Google Scholar]
- Bezie, A.; Regasa, H. The Role of Starter Culture and Enzymes/ Rennet for Fermented Dairy Products Manufacture—A Review. Nutr. Food Sci. Int. J. 2019, 9, 555756. [Google Scholar]
- Khedkar, C.D.; Kalyankar, S.D.; Deosarkar, S.S. Fermented Foods: Fermented Milks. In Encyclopedia of Food and Health; Elsevier: Amsterdam, The Netherlands, 2016; pp. 661–667. ISBN 978-0-12-384953-3. [Google Scholar]
- Górska-Warsewicz, H.; Rejman, K.; Laskowski, W.; Czeczotko, M. Milk and Dairy Products and Their Nutritional Contribution to the Average Polish Diet. Nutrients 2019, 11, 1771. [Google Scholar] [CrossRef]
- Walther, B.; Guggisberg, D.; Badertscher, R.; Egger, L.; Portmann, R.; Dubois, S.; Haldimann, M.; Kopf-Bolanz, K.; Rhyn, P.; Zoller, O.; et al. Comparison of Nutritional Composition between Plant-Based Drinks and Cow’s Milk. Front. Nutr. 2022, 9, 988707. [Google Scholar] [CrossRef]
- Leischner, C.; Egert, S.; Burkard, M.; Venturelli, S. Potential Protective Protein Components of Cow’s Milk against Certain Tumor Entities. Nutrients 2021, 13, 1974. [Google Scholar] [CrossRef]
- Woźniak, D.; Cichy, W.; Dobrzyńska, M.; Przysławski, J.; Drzymała-Czyż, S. Reasonableness of Enriching Cow’s Milk with Vitamins and Minerals. Foods 2022, 11, 1079. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, F.; Pellegrino, L.; Verduci, E.; Ghiselli, A.; Bernabei, R.; Calvani, R.; Cetin, I.; Giampietro, M.; Perticone, F.; Piretta, L.; et al. Cow’s Milk Consumption and Health: A Health Professional’s Guide. J. Am. Coll. Nutr. 2019, 38, 197–208. [Google Scholar] [CrossRef]
- Weiss, W.P. A 100-Year Review: From Ascorbic Acid to Zinc-Mineral and Vitamin Nutrition of Dairy Cows. J. Dairy Sci. 2017, 100, 10045–10060. [Google Scholar] [CrossRef] [PubMed]
- Haug, A.; Høstmark, A.T.; Harstad, O.M. Bovine Milk in Human Nutrition—A Review. Lipids Health Dis. 2007, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; He, S.; Li, H. Yak Milk. In Milk and Dairy Products in Human Nutrition; Park, Y.W., Haenlein, G.F.W., Eds.; John Wiley & Sons: Oxford, UK, 2013; pp. 627–643. ISBN 978-1-118-53416-8. [Google Scholar]
- Fox, P.F. Introduction | History of Dairy Products and Processes. In Encyclopedia of Dairy Sciences; Elsevier: Amsterdam, The Netherlands, 2011; pp. 12–17. ISBN 978-0-12-374407-4. [Google Scholar]
- Surono, I.S.; Hosono, A. FERMENTED MILKS|Types and Standards of Identity. In Encyclopedia of Dairy Sciences; Elsevier: Amsterdam, The Netherlands, 2011; pp. 470–476. ISBN 978-0-12-374407-4. [Google Scholar]
- Tamime, A. (Ed.) Types of Fermented Milks; Blackwell Publishing Ltd.: Oxford, UK, 2006; pp. 1–8. ISBN 978-0-470-99550-1. [Google Scholar]
- Guarner, F.; Perdigon, G.; Corthier, G.; Salminen, S.; Koletzko, B.; Morelli, L. Should Yoghurt Cultures Be Considered Probiotic? Br. J. Nutr. 2005, 93, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Jomaa, S.A.; Abd El-Wahed, A.; El-Seedi, H.R. The Many Faces of Kefir Fermented Dairy Products: Quality Characteristics, Flavour Chemistry, Nutritional Value, Health Benefits, and Safety. Nutrients 2020, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Toba, T.; Kotani, T.; Adachi, S. Capsular Polysaccharide of a Slime-Forming Lactococcus lactis ssp. Cremoris LAPT 3001 Isolated from Swedish Fermented Milk ‘Långfil.’. Int. J. Food Microbiol. 1991, 12, 167–171. [Google Scholar] [CrossRef]
- Neve, H.; Geis, A.; Teuber, M. Plasmid-Encoded Functions of Ropy Lactic Acid Streptococcal Strains from Scandinavian Fermented Milk. Biochimie 1988, 70, 437–442. [Google Scholar] [CrossRef]
- Ni, H.; Bao, Q.; Sun, T.; Chen, X.; Zhang, H. [Identification and biodiversity of yeasts isolated from Koumiss in Xinjiang of China]. Wei Sheng Wu Xue Bao 2007, 47, 578–582. [Google Scholar]
- Wang, H.; Hussain, T.; Yao, J.; Li, J.; Sabir, N.; Liao, Y.; Liang, Z.; Wang, Y.; Liu, Y.; Zhao, D.; et al. Koumiss Promotes Mycobacterium Bovis Infection by Disturbing Intestinal Flora and Inhibiting Endoplasmic Reticulum Stress. FASEB J. 2021, 35, e21777. [Google Scholar] [CrossRef]
- de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Maske, B.L.; De Dea Lindner, J.; Vale, A.S.; Favero, G.R.; Viesser, J.; de Carvalho, J.C.; Góes-Neto, A.; Soccol, C.R. An Updated Review on Bacterial Community Composition of Traditional Fermented Milk Products: What Next-Generation Sequencing Has Revealed so Far? Crit. Rev. Food Sci. Nutr. 2022, 62, 1870–1889. [Google Scholar] [CrossRef]
- Oki, K.; Dugersuren, J.; Demberel, S.; Watanabe, K. Pyrosequencing Analysis of the Microbial Diversity of Airag, Khoormog and Tarag, Traditional Fermented Dairy Products of Mongolia. Biosci. Microbiota Food Health 2014, 33, 53–64. [Google Scholar] [CrossRef]
- Abdelgadir, W.; Nielsen, D.S.; Hamad, S.; Jakobsen, M. A Traditional Sudanese Fermented Camel’s Milk Product, Gariss, as a Habitat of Streptococcus infantarius subsp. Infantarius. Int. J. Food Microbiol. 2008, 127, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.A.; Marette, A. Potential Health Benefits of Combining Yogurt and Fruits Based on Their Probiotic and Prebiotic Properties. Adv. Nutr. 2017, 8, 155S–164S. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, D.; Cichy, W.; Przysławski, J.; Drzymała-Czyż, S. The Role of Microbiota and Enteroendocrine Cells in Maintaining Homeostasis in the Human Digestive Tract. Adv. Med. Sci. 2021, 66, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Bibbò, S.; Ianiro, G.; Giorgio, V.; Scaldaferri, F.; Masucci, L.; Gasbarrini, A.; Cammarota, G. The Role of Diet on Gut Microbiota Composition. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4742–4749. [Google Scholar]
- Iebba, V.; Totino, V.; Gagliardi, A.; Santangelo, F.; Cacciotti, F.; Trancassini, M.; Mancini, C.; Cicerone, C.; Corazziari, E.; Pantanella, F.; et al. Eubiosis and Dysbiosis: The Two Sides of the Microbiota. New Microbiol. 2016, 39, 1–12. [Google Scholar]
- Gagliardi, A.; Totino, V.; Cacciotti, F.; Iebba, V.; Neroni, B.; Bonfiglio, G.; Trancassini, M.; Passariello, C.; Pantanella, F.; Schippa, S. Rebuilding the Gut Microbiota Ecosystem. Int. J. Environ. Res. Public Health 2018, 15, 1679. [Google Scholar] [CrossRef]
- Martinez-Guryn, K.; Leone, V.; Chang, E.B. Regional Diversity of the Gastrointestinal Microbiome. Cell Host Microbe 2019, 26, 314–324. [Google Scholar] [CrossRef]
- Senghor, B.; Sokhna, C.; Ruimy, R.; Lagier, J.-C. Gut Microbiota Diversity According to Dietary Habits and Geographical Provenance. Hum. Microbiome J. 2018, 7–8, 1–9. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.-X.; Chen, X.-Y.; Wang, J.-Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in Health and Diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef]
- Tasnim, N.; Abulizi, N.; Pither, J.; Hart, M.M.; Gibson, D.L. Linking the Gut Microbial Ecosystem with the Environment: Does Gut Health Depend on Where We Live? Front. Microbiol. 2017, 8, 1935. [Google Scholar] [CrossRef]
- Hakansson, A.; Molin, G. Gut Microbiota and Inflammation. Nutrients 2011, 3, 637–682. [Google Scholar] [CrossRef]
- Ianiro, G.; Bibbò, S.; Gasbarrini, A.; Cammarota, G. Therapeutic Modulation of Gut Microbiota: Current Clinical Applications and Future Perspectives. Curr. Drug Targets 2014, 15, 762–770. [Google Scholar] [CrossRef]
- Brenchley, J.M.; Douek, D.C. Microbial Translocation across the GI Tract. Annu Rev. Immunol. 2012, 30, 149–173. [Google Scholar] [CrossRef] [PubMed]
- Aslam, H.; Marx, W.; Rocks, T.; Loughman, A.; Chandrasekaran, V.; Ruusunen, A.; Dawson, S.L.; West, M.; Mullarkey, E.; Pasco, J.A.; et al. The Effects of Dairy and Dairy Derivatives on the Gut Microbiota: A Systematic Literature Review. Gut Microbes 2020, 12, 1799533. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.; Gasbarrini, A.; Mele, M. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the Human Gut Microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.-N.; Liu, X.-T.; Liang, Z.-H.; Wang, J.-H. Gut Microbiota in Obesity. WJG 2021, 27, 3837–3850. [Google Scholar] [CrossRef]
- Qiu, P.; Ishimoto, T.; Fu, L.; Zhang, J.; Zhang, Z.; Liu, Y. The Gut Microbiota in Inflammatory Bowel Disease. Front. Cell. Infect. Microbiol. 2022, 12, 733992. [Google Scholar] [CrossRef]
- Pascal, M.; Perez-Gordo, M.; Caballero, T.; Escribese, M.M.; Lopez Longo, M.N.; Luengo, O.; Manso, L.; Matheu, V.; Seoane, E.; Zamorano, M.; et al. Microbiome and Allergic Diseases. Front. Immunol. 2018, 9, 1584. [Google Scholar] [CrossRef]
- Vivarelli, S.; Salemi, R.; Candido, S.; Falzone, L.; Santagati, M.; Stefani, S.; Torino, F.; Banna, G.L.; Tonini, G.; Libra, M. Gut Microbiota and Cancer: From Pathogenesis to Therapy. Cancers 2019, 11, 38. [Google Scholar] [CrossRef]
- Tsikhan, N.; Belevtsev, M. Oral Tolerance Induction and Food Allergy Prevention: Oral Tolerance Induction and Food Allergy Prevention. JMS 2019, 88, 177–183. [Google Scholar] [CrossRef]
- Al-Domi, H. Paleolithic Hunter-Gatherers’ Dietary Patterns: Implications and Consequences. AJFAND 2015, 15, 9935–9948. [Google Scholar] [CrossRef]
- Alt, K.W.; Al-Ahmad, A.; Woelber, J.P. Nutrition and Health in Human Evolution–Past to Present. Nutrients 2022, 14, 3594. [Google Scholar] [CrossRef]
- Crittenden, A.N.; Schnorr, S.L. Current Views on Hunter-Gatherer Nutrition and the Evolution of the Human Diet. Am. J. Phys. Anthropol. 2017, 162, 84–109. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef] [PubMed]
- Romani, M.; Adouane, E.; Carrion, C.; Veckerlé, C.; Boeuf, D.; Fernandez, F.; Lefèvre, M.; Intertaglia, L.; Rodrigues, A.M.S.; Lebaron, P.; et al. Diversity and Activities of Pioneer Bacteria, Algae, and Fungi Colonizing Ceramic Roof Tiles during the First Year of Outdoor Exposure. Int. Biodeterior. Biodegrad. 2021, 162, 105230. [Google Scholar] [CrossRef]
- Armbruster, C.R.; Parsek, M.R. New Insight into the Early Stages of Biofilm Formation. Proc. Natl. Acad. Sci. USA 2018, 115, 4317–4319. [Google Scholar] [CrossRef] [PubMed]
- Cornejo Ulloa, P.; van der Veen, M.H.; Krom, B.P. Review: Modulation of the Oral Microbiome by the Host to Promote Ecological Balance. Odontology 2019, 107, 437–448. [Google Scholar] [CrossRef]
- Patangia, D.V.; Anthony Ryan, C.; Dempsey, E.; Paul Ross, R.; Stanton, C. Impact of Antibiotics on the Human Microbiome and Consequences for Host Health. MicrobiologyOpen 2022, 11, e1260. [Google Scholar] [CrossRef]
- Siddiqui, R.; Mungroo, M.R.; Alharbi, A.M.; Alfahemi, H.; Khan, N.A. The Use of Gut Microbial Modulation Strategies as Interventional Strategies for Ageing. Microorganisms 2022, 10, 1869. [Google Scholar] [CrossRef]
- Huurre, A.; Kalliomäki, M.; Rautava, S.; Rinne, M.; Salminen, S.; Isolauri, E. Mode of Delivery—Effects on Gut Microbiota and Humoral Immunity. Neonatology 2008, 93, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Chong, C.; Bloomfield, F.; O’Sullivan, J. Factors Affecting Gastrointestinal Microbiome Development in Neonates. Nutrients 2018, 10, 274. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, K.; Henriksen, L. Cesarean Section and Disease Associated with Immune Function. J. Allergy Clin. Immunol. 2016, 137, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Duan, Y.; Fouts, D.E.; Schnabl, B. Intestinal Virome and Therapeutic Potential of Bacteriophages in Liver Disease. J. Hepatol. 2021, 75, 1465–1475. [Google Scholar] [CrossRef]
- Li, Y.; Nishu; Yellezuome, D.; Chai, M.; Li, C.; Liu, R. Catalytic Pyrolysis of Biomass over Fe-Modified Hierarchical ZSM-5: Insights into Mono-Aromatics Selectivity and Pyrolysis Behavior Using Py-GC/MS and TG-FTIR. J. Energy Inst. 2021, 99, 218–228. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.A.; Ragab, S.H.; ElBaky, A.A.; Shoeib, A.R.S.; Alhosary, Y.; Fekry, D. Frequency of Firmicutes and Bacteroidetes in Gut Microbiota in Obese and Normal Weight Egyptian Children and Adults. Arch. Med. Sci. 2011, 3, 501–507. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel Disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Moraes, A.C.F.; de Almeida-Pittito, B.; Ferreira, S.R.G. The Gut Microbiome in Vegetarians. In Microbiome and Metabolome in Diagnosis, Therapy, and Other Strategic Applications; Elsevier: Amsterdam, The Netherlands, 2019; pp. 393–400. ISBN 978-0-12-815249-2. [Google Scholar]
- MetaHIT Consortium (Additional Members); Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; et al. Enterotypes of the Human Gut Microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Imhann, F.; Vich Vila, A.; Bonder, M.J.; Lopez Manosalva, A.G.; Koonen, D.P.Y.; Fu, J.; Wijmenga, C.; Zhernakova, A.; Weersma, R.K. The Influence of Proton Pump Inhibitors and Other Commonly Used Medication on the Gut Microbiota. Gut Microbes 2017, 8, 351–358. [Google Scholar] [CrossRef]
- Walsh, J.; Griffin, B.T.; Clarke, G.; Hyland, N.P. Drug-Gut Microbiota Interactions: Implications for Neuropharmacology: Drug-Gut Microbiota Interactions. Br. J. Pharmacol. 2018, 175, 4415–4429. [Google Scholar] [CrossRef]
- Weersma, R.K.; Zhernakova, A.; Fu, J. Interaction between Drugs and the Gut Microbiome. Gut 2020, 69, 1510–1519. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Singh, V.; Mani, I. Dysbiosis of Human Microbiome and Infectious Diseases. Prog. Mol. Biol. Transl. Sci. 2022, 192, 33–51. [Google Scholar] [CrossRef]
- Tiffany, C.R.; Bäumler, A.J. Dysbiosis: From Fiction to Function. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G602–G608. [Google Scholar] [CrossRef]
- Martin-Subero, M.; Anderson, G.; Kanchanatawan, B.; Berk, M.; Maes, M. Comorbidity between Depression and Inflammatory Bowel Disease Explained by Immune-Inflammatory, Oxidative, and Nitrosative Stress; Tryptophan Catabolite; and Gut-Brain Pathways. CNS Spectr. 2016, 21, 184–198. [Google Scholar] [CrossRef]
- Jalili-Firoozinezhad, S.; Gazzaniga, F.S.; Calamari, E.L.; Camacho, D.M.; Fadel, C.W.; Bein, A.; Swenor, B.; Nestor, B.; Cronce, M.J.; Tovaglieri, A.; et al. A Complex Human Gut Microbiome Cultured in an Anaerobic Intestine-on-a-Chip. Nat. Biomed. Eng. 2019, 3, 520–531. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Roos, S.; Eeckhaut, V.; MacKenzie, D.A.; Derde, M.; Verstraete, W.; Marzorati, M.; Possemiers, S.; Vanhoecke, B.; Van Immerseel, F.; et al. Incorporating a Mucosal Environment in a Dynamic Gut Model Results in a More Representative Colonization by Lactobacilli. Microb. Biotechnol. 2012, 5, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic Mechanisms of Action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Merenstein, D.; Gonzalez, J.; Young, A.G.; Roberts, R.F.; Sanders, M.E.; Petterson, S. Study to Investigate the Potential of Probiotics in Children Attending School. Eur. J. Clin. Nutr. 2011, 65, 447–453. [Google Scholar] [CrossRef]
- Laukens, D.; Brinkman, B.M.; Raes, J.; De Vos, M.; Vandenabeele, P. Heterogeneity of the Gut Microbiome in Mice: Guidelines for Optimizing Experimental Design. FEMS Microbiol. Rev. 2016, 40, 117–132. [Google Scholar] [CrossRef]
- Pace, F.; Pace, M.; Quartarone, G. Probiotics in Digestive Diseases: Focus on Lactobacillus GG. Minerva Gastroenterol. Dietol. 2015, 61, 273–292. [Google Scholar] [PubMed]
- Al-Rashidi, H.E. Gut Microbiota and Immunity Relevance in Eubiosis and Dysbiosis. Saudi J. Biol. Sci. 2022, 29, 1628–1643. [Google Scholar] [CrossRef]
- Bajinka, O.; Tan, Y.; Abdelhalim, K.A.; Özdemir, G.; Qiu, X. Extrinsic Factors Influencing Gut Microbes, the Immediate Consequences and Restoring Eubiosis. AMB Express 2020, 10, 130. [Google Scholar] [CrossRef] [PubMed]
- DeGruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current Understanding of Dysbiosis in Disease in Human and Animal Models. Inflamm. Bowel Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef]
- Wang, B.; Yao, M.; Lv, L.; Ling, Z.; Li, L. The Human Microbiota in Health and Disease. Engineering 2017, 3, 71–82. [Google Scholar] [CrossRef]
- Hasan, N.; Yang, H. Factors Affecting the Composition of the Gut Microbiota, and Its Modulation. PeerJ 2019, 7, e7502. [Google Scholar] [CrossRef]
- Hou, J.K.; Abraham, B.; El-Serag, H. Dietary Intake and Risk of Developing Inflammatory Bowel Disease: A Systematic Review of the Literature. Am. J. Gastroenterol. 2011, 106, 563–573. [Google Scholar] [CrossRef]
- Lo, C.-H.; Lochhead, P.; Khalili, H.; Song, M.; Tabung, F.K.; Burke, K.E.; Richter, J.M.; Giovannucci, E.L.; Chan, A.T.; Ananthakrishnan, A.N. Dietary Inflammatory Potential and Risk of Crohn’s Disease and Ulcerative Colitis. Gastroenterology 2020, 159, 873–883.e1. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N.; Khalili, H.; Song, M.; Higuchi, L.M.; Richter, J.M.; Nimptsch, K.; Wu, K.; Chan, A.T. High School Diet and Risk of Crohn’s Disease and Ulcerative Colitis. Inflamm. Bowel Dis. 2015, 21, 2311–2319. [Google Scholar] [CrossRef]
- Cozma-Petruţ, A.; Loghin, F.; Miere, D.; Dumitraşcu, D.L. Diet in Irritable Bowel Syndrome: What to Recommend, Not What to Forbid to Patients! WJG 2017, 23, 3771. [Google Scholar] [CrossRef]
- Pérez-Cobas, A.; Moya, A.; Gosalbes, M.; Latorre, A. Colonization Resistance of the Gut Microbiota against Clostridium Difficile. Antibiotics 2015, 4, 337–357. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human Gut Microbiota/Microbiome in Health and Diseases: A Review. Antonie Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, J.; Wang, L. Role and Mechanism of Gut Microbiota in Human Disease. Front. Cell Infect. Microbiol. 2021, 11, 625913. [Google Scholar] [CrossRef] [PubMed]
- Sorboni, S.G.; Moghaddam, H.S.; Jafarzadeh-Esfehani, R.; Soleimanpour, S. A Comprehensive Review on the Role of the Gut Microbiome in Human Neurological Disorders. Clin. Microbiol. Rev. 2022, 35, e0033820. [Google Scholar] [CrossRef]
- Curtis, M.A.; Diaz, P.I.; Van Dyke, T.E. The Role of the Microbiota in Periodontal Disease. Periodontol. 2000 2020, 83, 14–25. [Google Scholar] [CrossRef]
- Kitamoto, S.; Nagao-Kitamoto, H.; Hein, R.; Schmidt, T.M.; Kamada, N. The Bacterial Connection between the Oral Cavity and the Gut Diseases. J. Dent. Res. 2020, 99, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Kinashi, Y.; Hase, K. Partners in Leaky Gut Syndrome: Intestinal Dysbiosis and Autoimmunity. Front. Immunol. 2021, 12, 673708. [Google Scholar] [CrossRef]
- Amabebe, E.; Robert, F.O.; Agbalalah, T.; Orubu, E.S.F. Microbial Dysbiosis-Induced Obesity: Role of Gut Microbiota in Homoeostasis of Energy Metabolism. Br. J. Nutr. 2020, 123, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Bushyhead, D.; Quigley, E.M.M. Small Intestinal Bacterial Overgrowth-Pathophysiology and Its Implications for Definition and Management. Gastroenterology 2022, 163, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Laghi, L.; Mastromarino, P.; Elisei, W.; Capobianco, D.; Zhu, C.L.; Picchio, M.; Giorgetti, G.; Brandimarte, G.; Tursi, A. Impact of Treatments on Fecal Microbiota and Fecal Metabolome in Symptomatic Uncomplicated Diverticular Disease of the Colon: A Pilot Study. J. Biol. Regul. Homeost. Agents 2018, 32, 1421–1432. [Google Scholar]
- Santos-Marcos, J.A.; Perez-Jimenez, F.; Camargo, A. The Role of Diet and Intestinal Microbiota in the Development of Metabolic Syndrome. J. Nutr. Biochem. 2019, 70, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, M.C.; Andersen, J.F.; Gorelick, P.L.; Caspar, P.; Suerbaum, S.; Fox, J.G.; Cheever, A.W.; Jankovic, D.; Sher, A. Induction of Colitis by a CD4+ T Cell Clone Specific for a Bacterial Epitope. Proc. Natl. Acad. Sci. USA 2003, 100, 15830–15835. [Google Scholar] [CrossRef]
- Kullberg, M.C.; Jankovic, D.; Gorelick, P.L.; Caspar, P.; Letterio, J.J.; Cheever, A.W.; Sher, A. Bacteria-Triggered CD4(+) T Regulatory Cells Suppress Helicobacter Hepaticus-Induced Colitis. J. Exp. Med. 2002, 196, 505–515. [Google Scholar] [CrossRef]
- Andoh, A.; Imaeda, H.; Aomatsu, T.; Inatomi, O.; Bamba, S.; Sasaki, M.; Saito, Y.; Tsujikawa, T.; Fujiyama, Y. Comparison of the Fecal Microbiota Profiles between Ulcerative Colitis and Crohn’s Disease Using Terminal Restriction Fragment Length Polymorphism Analysis. J. Gastroenterol. 2011, 46, 479–486. [Google Scholar] [CrossRef]
- Sepehri, S.; Khafipour, E.; Bernstein, C.N.; Coombes, B.K.; Pilar, A.V.; Karmali, M.; Ziebell, K.; Krause, D.O. Characterization of Escherichia Coli Isolated from Gut Biopsies of Newly Diagnosed Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2011, 17, 1451–1463. [Google Scholar] [CrossRef]
- Matsuura, M. Structural Modifications of Bacterial Lipopolysaccharide That Facilitate Gram-Negative Bacteria Evasion of Host Innate Immunity. Front. Immunol. 2013, 4, 109. [Google Scholar] [CrossRef] [PubMed]
- Durack, J.; Lynch, S.V. The Gut Microbiome: Relationships with Disease and Opportunities for Therapy. J. Exp. Med. 2019, 216, 20–40. [Google Scholar] [CrossRef] [PubMed]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal Barrier and Gut Microbiota: Shaping Our Immune Responses throughout Life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Stecher, B. The Roles of Inflammation, Nutrient Availability and the Commensal Microbiota in Enteric Pathogen Infection. Microbiol. Spectr. 2015, 3, 1–17. [Google Scholar] [CrossRef]
- Guerin-Danan, C.; Chabanet, C.; Pedone, C.; Popot, F.; Vaissade, P.; Bouley, C.; Szylit, O.; Andrieux, C. Milk Fermented with Yogurt Cultures and Lactobacillus Casei Compared with Yogurt and Gelled Milk: Influence on Intestinal Microflora in Healthy Infants. Am. J. Clin. Nutr. 1998, 67, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Uyeno, Y.; Sekiguchi, Y.; Kamagata, Y. Impact of Consumption of Probiotic Lactobacilli-Containing Yogurt on Microbial Composition in Human Feces. Int. J. Food Microbiol. 2008, 122, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Odamaki, T.; Sugahara, H.; Yonezawa, S.; Yaeshima, T.; Iwatsuki, K.; Tanabe, S.; Tominaga, T.; Togashi, H.; Benno, Y.; Xiao, J. Effect of the Oral Intake of Yogurt Containing Bifidobacterium Longum BB536 on the Cell Numbers of Enterotoxigenic Bacteroides Fragilis in Microbiota. Anaerobe 2012, 18, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Merenstein, D.J.; Tan, T.P.; Molokin, A.; Smith, K.H.; Roberts, R.F.; Shara, N.M.; Mete, M.; Sanders, M.E.; Solano-Aguilar, G. Safety of Bifidobacterium Animalis Subsp. Lactis (B. Lactis) Strain BB-12-Supplemented Yogurt in Healthy Adults on Antibiotics: A Phase I Safety Study. Gut Microbes 2015, 6, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Volokh, O.; Klimenko, N.; Berezhnaya, Y.; Tyakht, A.; Nesterova, P.; Popenko, A.; Alexeev, D. Human Gut Microbiome Response Induced by Fermented Dairy Product Intake in Healthy Volunteers. Nutrients 2019, 11, 547. [Google Scholar] [CrossRef]
- El Manouni El Hassani, S.; de Boer, N.K.H.; Jansen, F.M.; Benninga, M.A.; Budding, A.E.; de Meij, T.G.J. Effect of Daily Intake of Lactobacillus Casei on Microbial Diversity and Dynamics in a Healthy Pediatric Population. Curr. Microbiol. 2019, 76, 1020–1027. [Google Scholar] [CrossRef]
- Amamoto, R.; Shimamoto, K.; Park, S.; Matsumoto, H.; Shimizu, K.; Katto, M.; Tsuji, H.; Matsubara, S.; Shephard, R.J.; Aoyagi, Y. Yearly Changes in the Composition of Gut Microbiota in the Elderly, and the Effect of Lactobacilli Intake on These Changes. Sci. Rep. 2021, 11, 12765. [Google Scholar] [CrossRef]
- Ba, Z.; Lee, Y.; Meng, H.; Kris-Etherton, P.M.; Rogers, C.J.; Lewis, Z.T.; Mills, D.A.; Furumoto, E.J.; Rolon, M.L.; Fleming, J.A.; et al. Matrix Effects on the Delivery Efficacy of Bifidobacterium Animalis Subsp. Lactis BB-12 on Fecal Microbiota, Gut Transit Time, and Short-Chain Fatty Acids in Healthy Young Adults. mSphere 2021, 6, e0008421. [Google Scholar] [CrossRef]
- Aumeistere, L.; Ķibilds, J.; Siksna, I.; Neimane, L.V.; Kampara, M.; Ļubina, O.; Ciproviča, I. The Gut Microbiome among Postmenopausal Latvian Women in Relation to Dietary Habits. Nutrients 2022, 14, 3568. [Google Scholar] [CrossRef]
- Dewit, O.; Boudraa, G.; Touhami, M.; Desjeux, J.F. Breath Hydrogen Test and Stools Characteristics after Ingestion of Milk and Yogurt in Malnourished Children with Chronic Diarrhoea and Lactase Deficiency. J. Trop. Pediatr. 1987, 33, 177–180. [Google Scholar] [CrossRef]
- Boudraa, G.; Touhami, M.; Pochart, P.; Soltana, R.; Mary, J.Y.; Desjeux, J.F. Effect of Feeding Yogurt versus Milk in Children with Persistent Diarrhea. J. Pediatr. Gastroenterol. Nutr. 1990, 11, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, S.; Singh, K.D.; Sazawal, S.; Saxena, S.K.; Bhan, M.K. Efficacy of Milk versus Yogurt Offered as Part of a Mixed Diet in Acute Noncholera Diarrhea among Malnourished Children. J. Pediatr. 1998, 132, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Pedone, C.A.; Bernabeu, A.O.; Postaire, E.R.; Bouley, C.F.; Reinert, P. The Effect of Supplementation with Milk Fermented by Lactobacillus Casei (Strain DN-114 001) on Acute Diarrhoea in Children Attending Day Care Centres. Int. J. Clin. Pract. 1999, 53, 179–184. [Google Scholar] [PubMed]
- Pashapour, N.; Iou, S.G. Evaluation of Yogurt Effect on Acute Diarrhea in 6-24-Month-Old Hospitalized Infants. Turk. J. Pediatr. 2006, 48, 115–118. [Google Scholar] [PubMed]
- Eren, M.; Dinleyici, E.C.; Vandenplas, Y. Clinical Efficacy Comparison of Saccharomyces Boulardii and Yogurt Fluid in Acute Non-Bloody Diarrhea in Children: A Randomized, Controlled, Open Label Study. Am. J. Trop. Med. Hyg. 2010, 82, 488–491. [Google Scholar] [CrossRef]
- Ishikawa, H.; Akedo, I.; Umesaki, Y.; Tanaka, R.; Imaoka, A.; Otani, T. Randomized Controlled Trial of the Effect of Bifidobacteria-Fermented Milk on Ulcerative Colitis. J. Am. Coll. Nutr. 2003, 22, 56–63. [Google Scholar] [CrossRef]
- Kato, K.; Mizuno, S.; Umesaki, Y.; Ishii, Y.; Sugitani, M.; Imaoka, A.; Otsuka, M.; Hasunuma, O.; Kurihara, R.; Iwasaki, A.; et al. Randomized Placebo-Controlled Trial Assessing the Effect of Bifidobacteria-Fermented Milk on Active Ulcerative Colitis. Aliment. Pharmacol. Ther. 2004, 20, 1133–1141. [Google Scholar] [CrossRef]
- Laake, K.O.; Bjørneklett, A.; Aamodt, G.; Aabakken, L.; Jacobsen, M.; Bakka, A.; Vatn, M.H. Outcome of Four Weeks’ Intervention with Probiotics on Symptoms and Endoscopic Appearance after Surgical Reconstruction with a J-Configurated Ileal-Pouch-Anal-Anastomosis in Ulcerative Colitis. Scand. J. Gastroenterol. 2005, 40, 43–51. [Google Scholar] [CrossRef]
- Matsuoka, K.; Uemura, Y.; Kanai, T.; Kunisaki, R.; Suzuki, Y.; Yokoyama, K.; Yoshimura, N.; Hibi, T. Efficacy of Bifidobacterium Breve Fermented Milk in Maintaining Remission of Ulcerative Colitis. Dig. Dis. Sci. 2018, 63, 1910–1919. [Google Scholar] [CrossRef]
- Hong, Y.-S.; Hong, K.S.; Park, M.-H.; Ahn, Y.-T.; Lee, J.-H.; Huh, C.-S.; Lee, J.; Kim, I.-K.; Hwang, G.-S.; Kim, J.S. Metabonomic Understanding of Probiotic Effects in Humans with Irritable Bowel Syndrome. J. Clin. Gastroenterol. 2011, 45, 415–425. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Nutrients | Cow Milk | Goat Milk | Sheep Milk | Buffalo Milk | Mare Milk | Camel (Dromedary) Milk | Donkey Milk | Yak Milk |
|---|---|---|---|---|---|---|---|---|
| Energy (kcal) | 59–66 | 57–69 | 93–108 | 71–118 | 42–50 | 44–79 | 32–51 | 87–91 |
| Energy (kJ) | 247–274 | 243–289 | 388–451 | 296–495 | 177–210 | 185–332 | 135–215 | 349–382 |
| Water (g) | 87.3–88.1 | 86.4–89.0 | 80.7–83.0 | 82.3–84.0 | 87.9–91.3 | 88.7–89.4 | 89.2–91.5 | 75.3–84.4 |
| Protein (g) | 3.2–3.4 | 2.9–3.8 | 5.4–6.0 | 2.7–4.6 | 1.4–3.2 | 2.4–4.2 | 1.4–1.8 | 4.2–5.9 |
| Fat (g) | 3.1–3.3 | 3.3–4.5 | 5.8–7.0 | 5.3–9.0 | 0.5–4.2 | 2.0–6.0 | 0.3–1.8 | 5.6–9.5 |
| Lactose (g) | 4.5–5.1 | 4.2–4.5 | 4.5–5.4 | 3.2–4.9 | 5.6–7.2 | 3.5–4.9 | 5.9–6.9 | 3.3–6.2 |
| Calcium (mg) | 91–120 | 100–134 | 170–207 | 147–220 | 76–124 | 105–120 | 68–115 | 119–134 |
| Iron (mg) | Tr.–0.2 | Tr.–0.6 | Tr.–0.1 | 0.2 * | Tr.–0.2 | 0.2–0.3 | 0.2–1.0 | |
| Magnesium (mg) | 10–11 | 13–14 | 18 * | 2–16 | 4–12 | 12–14 | 4 * | 8–12 |
| Phosphorus (mg) | 84–95 | 90–111 | 123–158 | 102–293 | 43–83 | 83–90 | 49–73 | 77–135 |
| Potassium (mg) | 132–155 | 170–228 | 120–187 | 112 * | 25–87 | 124–173 | 50 * | 83–107 |
| Sodium (mg) | 38–45 | 32–50 | 30–44 | 47 * | 13–20 | 59–73 | 22 * | 21–38 |
| Zinc (mg) | 0.3–0.4 | 0.1–0.5 | 0.5–0.7 | 0.5 * | 0.2–0.3 | 0.4–0.6 | 0.7–1.1 | |
| Copper (mg) | Tr. | Tr.–0.1 | 0.1–0.1 | Tr.–0.1 | 0.1–0.2 | 0.4 * | ||
| Selenium (μg) | 1.0–3.7 | 0.7–1.4 | 1.7 * | |||||
| Manganese (μg) | 4–10 | Tr.–18 | Tr.–18 | 60–180 | ||||
| Vitamin A (μg) | 30–46 | 30–74 | 64 * | 69 * | 14 * | |||
| Vitamin E (mg) | 0.1–0.1 | Tr.–0.1 | 0.1–0.1 | 0.2–2.0 | Tr | |||
| Thiamin (mg) | Tr. | Tr.–0.1 | 0.1–0.1 | 0.1 * | Tr. | 0.1 * | 0.1 * | |
| Riboflavin (mg) | 0.2–0.2 | Tr.–0.2 | 0.3–0.4 | 0.1 * | Tr. | 0.1 * | Tr. | 0.1 * |
| Niacin (mg) | 0.1–0.2 | 0.1–0.3 | 0.4–0.4 | 0.2 * | 0.1 * | 0.1 * | Tr. | |
| Pantothenic acid (mg) | 0.3–0.6 | 0.3–0.4 | 0.4–0.5 | 0.2 * | ||||
| Vitamin B6 (mg) | Tr.–0.1 | 0.1–0.1 | 0.1–0.1 | 0.3 * | Tr. | |||
| Folate (μg) | 5.0–8.0 | Tr.–1.0 | 5.0–7.0 | 0.6 * | ||||
| Biotin (μg) | 1.4–2.5 | 2.0–3.0 | 2.5–2.5 | 13.0 * | ||||
| Vitamin B12 (μg) | 0.3–0.9 | Tr.–0.1 | 0.6–0.7 | 0.4 * | ||||
| Vitamin C (mg) | Tr.–2.0 | 1.1–1.3 | 4.2–5.0 | 2.5 * | 1.7–8.1 | 2.5–18.4 | ||
| Vitamin D (μg) | 0.1–0.3 | 0.1–0.1 | 0.2–0.2 | 0.2 * |
| Fermented Milk Product | Type of Milk | Fermentation Culture | Basic Product Characteristic |
|---|---|---|---|
| Yogurt | All types, especially cow, goat, sheep, and buffalo milk | Streptococcus (Sc.) thermophilus and Lactobacillus (Lb.) delbrueckii sp. Bulgaricus | Tart flavor and texture related to the fermentation of sugars in milk and the production of lactic acid. |
| Kefir | Especially from cows, goats, or sheep milk | Lc. lactis subsp. lactis, Lc. lactis subsp. cremoris, citrate-positive Lc. lactis, Ln. mesenteroides subsp. cremoris, Ln. mesenteroides subsp. dextranicum, Sc. thermophilus, Lb. delbrueckii subsp. bulgaricus, Lb. acidophilus, Lb. helveticus, Lb. kefir, Lb. kefiranofaciens, Kluyveromyces marxianus, Saccharomyces spp. | From the North Caucasian regions and Turkey; contains the characteristic microflora of kefir grains; sour, bitter, and slightly carbonated taste similar to drinkable yogurt. The starter culture used affects the viscosity and chemical composition of kefir. |
| Kumis | Mare and donkey milk (Columbian kumis from cow milk) | Lb. acidophilus, Lb. delbrueckii subsp. bulgaricus, Saccharomyces lactis, Kluyveromyces marxianus Pichia membranaefaciens Saccharomyces cerevisiae | Traditionally produced by fermenting raw milk with yeast and lactic acid bacteria. |
| Långfil | Cow milk | Lc. lactis ssp. | Swedish ropy sour milk requires a low acidification temperature and long maturation; mildly acidic with a chewy and cohesive texture. |
| Viili | Cow and other milk | Lc. lactis, Geotrichum candidum | Finnish ropy milk product; semi-solid structure with a sharp taste and good diacetyl flavor. |
| Grassis | Camel milk | Lc. paracasei subsp., Lc. plantarum, Lc. lactis, Enterococcus spp., and Leuconostoc spp. | Consumed in various regions of the Sudan; obtained by semi-continuous or fed-batch fermentation process in large skin bags containing a large quantity of previously soured product. |
| Filmjölk | Cow milk | Lc. lactis, and Ln. mesenteroides subsp. Cremoris | Traditional fermented milk products from Sweden; a mild and slightly sour taste. |
| Buttermilk | All types of milk, especially cow milk | Lc. lactis subsp. lactis, Lc. lactis subsp. cremoris, Lc. lactis, and Ln. mesenteroides subsp. Cremoris | Obtained during the production of butter, containing water-soluble milk components and bioactive material derived from milk fat membrane globules. |
| Dadih | Buffalo milk | Lb. casei subsp. casei, Ln. paramesenteroides, Lb. plantarum, Lc. lactis subsp. lactis, Lc. lactis subsp. cremoris, citrate-positive Lc. lactis, Enterococcus faecium | Traditional fermented milk popular in West Sumatra (Indonesia); -produced by pouring fresh, raw, unheated milk into a capped bamboo tube and allowing it to ferment spontaneously at room temperature for a few days. |
| Dahi (Curud) | Cow milk, and sometimes buffalo, yak, or goat milk | Sc. thermophilus Lb. delbrueckii subsp. bulgaricus or Lc. lactis subsp. lactis, Lc. lactis subsp. cremoris, citrate-positive Lc. Lactis | Popular throughout the Indian subcontinent (around 90% of the total fermented milk products produced in India); obtained from pasteurized or boiled milk fermented with a culture. |
| Yakult | Cow milk | Lb. casei subsp. casei | Japanese sweetened fermented milk; consists of water, skimmed milk, glucose-fructose syrup, sucrose and bacterial strains. |
| Kurut | Yak milk and other animal milk | Lb. delbrueckii and Lb. helveticus | Traditional product in northwestern China; obtained by drying yogurt or ayran after filtration with the addition of salt. |
| Tarag | Goat and cow milk | Lb. helveticus and Lb. delbrueckii ssp. bulgaricus | Traditional product in Mongolia and China; produced from raw whole milk by backslopping method. |
| Leben | Cow, goat, sheep, and camel milk | Lc. lactis and Sc. thermophilus, Enterococcus faecium | Traditional fermented milk from the Middle East and North Africa; produced from raw milk |
| Khoormog | Camel milk | Lc. helveticus, Lc. kefiranofaciens and Lc. delbrueckii | Mongolian traditional food; a sour and alcoholic taste from raw milk. |
| Fermented Milk Products Used | Type of Bacteria | Dose | Time of Intervention | Study Population | Effect | References |
|---|---|---|---|---|---|---|
| Yogurt vs. Milk fermented with yogurt cultures and Lb. casei vs. Nonfermented gelled milk | Both fermented products contained at least 1 × 106 CFU/g Lb. bulgaricus, 1 × 109 CFU/g Sc. thermophilus, and 1 × 108 CFU/g Lb. casei | 125 g/d of one of the three products | 1 week baseline period, 1 month supplementation period, and 1 week follow-up Period | Infants: 39 healthy infants (randomly assigned to one of three groups) aged 10–18 months | In the yogurt group, the number of Enterococci in the feces increased, and the activity of β-glucuronidase significantly decreased. The percentage of branched-chain and long-chain fatty acids significantly decreased. | Guerin-Danan et al., 1998 [118] |
| Yogurt (three different yogurts) | Lactobacilli 6 × 107–2.4 × 108/g yogurt
| One serving per day depending on the study group: Yogurt 1–110 g/day, Yogurt 2–180 mL/day Yogurt 3–90 g/day | 20 days | Adults: 15 healthy adults (9 males and 6 females) were assigned to one of three groups; aged 24–46 years | The consumption of yogurts with probiotic strains was no more effective than yogurt which does not contain probiotic strains on the human fecal microbial composition. Bacteroides and Prevotella population levels and the Clostridium coccoides Eubacterium rectale group in fecal samples tended to change in response to ingestion, however, the change was not related to the yogurt type. The bacterial community in human feces may be altered by yogurt consumption but not related to probiotic lactic acid bacteria. | Uyeno et al., 2008 [119] |
| Strawberry yogurt with Bifidobacterium animalis subsp. lactis BB-12 vs. Yogurt without BB-12 (control group) | Bifidobacterium animalis subsp. lactis (1 × 1010 colony/100 mL) and Sc. thermophilus and Lb. delbrueckii subsp. Bulgaricus | Four fluid ounces (112 g) per day | 90 days | Children: 172 children from Washington (randomly assigned to one of two groups); aged 2–4 years | Yogurt was well tolerated in children but did not decrease absences due to illnesses in daycare. | Merenstein et al., 2011 [87] |
| Yogurt with Bifidobacterium longum BB536 vs. Ultra-high-temperature pasteurized milk | Bifidobacterium longum BB536 4.27 ± 1.25 × 108 CFU of living BB536 (more than 1.12 ± 0.62 × 108 CFU of BB536 at the end of the study) and 1 × 109 CFU of lactic acid bacteria | One portion per day
| 8 weeks | Adults: 32 healthy adults (11 male and 21 female) from Eastern Japan (randomly assigned to one of two groups); the mean age in the yogurt group was 41.1 ± 10.2 years, and in the milk group, 38.6 ± 7.5 years | The consumption of yogurt significantly decreases enterotoxigenic Bacteroides fragilis in the gut microbiota. | Odamaki et al., 2012 [120] |
| Yogurt with Bifidobacterium animalis subsp. lactis BB-12 vs. Yogurt without BB-12 (control group) | Bifidobacterium animalis subsp. lactis (1 × 1010 CFU/100 mL) | Four fluid ounces (112 g) per day | 10 days | Adults: 40 healthy adults (16 male and 24 female) randomly assigned to one of two groups; yogurts with BB-12 (n = 19) and control group (n = 21); mean age in the yogurt group of 33 years and the control group of 29 years | Bifidobacterium lactis fecal levels were modestly higher in the yogurt with BB-12 group. In a small subset of participants, consuming yogurt with BB-12 activated an array of immune genes associated with regulating and activating immune cells. | Merenstein et al., 2015 [121] |
| Yogurt with Bifidobacterium animalis subsp. lactis BB-12 | Bifidobacterium animalis subsp. lactis BB-12 | Twice a day, 125 mL of yogurt in the morning and evening | 30 days | Adults: 150 healthy volunteers from Russia (no exact information about the age of the patients) | Gut microbe content showed an increase in the presence of potentially beneficial bacteria, especially the genus Bifidobacterium, Adlercreutzia equolifaciens and Slackia isoflavoniconvertens. Increased ability to metabolize lactose and synthesize amino acids while reducing the potential for lipopolysaccharide synthesis. | Volokh et al., 2019 [122] |
| Fermented milk product vs. Control group (without any intervention) | Lactobacillus casei strain Shirota at the minimum concentration of 6.5 × 109 CFU | Commercially available fermented milk product (65 mL) taken during breakfast | 6 weeks | Children: 18 healthy children; study group (n = 6) and control group (n = 12); aged 12–18 years | Fermented milk product ingestion by healthy children does not result in a more diverse and stable gut microbiota composition. | El Manouni El Hassani et al., 2019 [123] |
| Fermented milk products | Lactocaseibacillus paracasei strain Shirota 0.9–40 billion CFU per bottle | Intake ≥ 3 days/week | 1 year | Adults: 218 Japanese participants; aged 66–91 years | Stabilisation of the gut microbiota in the elderly. | Amamoto et al., 2021 [124] |
| Strawberry yogurt (control group) vs. Strawberry yogurt with strain BB-12 added pre-fermentation vs. Strawberry yogurt with BB-12 added post-fermentation vs. Capsule containing BB-12 | Bifidobacterium animalis subsp. lactis BB-12 (log10 10 ± 0.5 × 109 or 3.16 × 109 and 3.16 × 1010 CFU of BB-12/ portion, in capsules log10 10 ± 0.5 CFU of BB-12/capsule | 240 g yogurt/day. | 4 treatments each lasting 4 weeks, and a 2 week wash-out compliance break between treatments | Adults: 36 healthy adults; 29 finished at least one treatment period (18 females and 11 males); mean age of 28.1 ± 0.6 years | Consumption of yogurt with BB-12 or capsule did not significantly alter the gut microbiota composition, gut transit times, and fecal excretion of short-chain fatty acids. A significant gender effect was observed when comparing the gut microbiota. Daily consumption of BB-12 in yogurt (with strain BB-12 added pre-fermentation and post-fermentation) resulted in a higher relative abundance of B. animalis. | Ba et al., 2021 [125] |
| Yogurt vs. Control group (without any intervention) | Lactic acid bacteria 1.4 × 109 CFU g−1 | 175 g of plain organic milk yogurt | 8 weeks | Adults: 52 postmenopausal women from Lativa; control (n = 26) and experimental group (n = 26); aged 44–69 years | No significant changes in the gut microbiome were related to the consumption of yogurt. Consumption of food products like grains, grain-based products, milk and milk products, and beverages (tea, coffee) is associated with differences in the composition of the gut microbiome. | Aumeistere et al., 2022 [126] |
| Fermented Milk Products Used | Type of Bacteria | Dose and Time of Intervention | Time of Intervention | Study Population | Effect | References |
|---|---|---|---|---|---|---|
| Diarrhea | ||||||
| Yogurt | Sc. thermophilus and Lb. bulgaricus | Individual dosage (depending on lactose) per kilogram of body weight | 4 days | Children: 9 Algerian boys with diarrhea of >1 month in duration, clinically mild malnutrition, villus atrophy, and lactose maldigestion; aged 7–29 months | Replacing milk (infant formula) with yogurt reduced lactose malabsorption and tended to improve lactose intolerance and diarrhea. | Dewit et al., 1987 [127] |
| Yogurt prepared from milk formulae vs. Milk formula | Sc. thermophilus and Lb. bulgaricus | Individual dosage per kilogram of body weight 150–180 kcal/kg/day for all foods (children aged 3–6 months received 4 servings of milk or yogurt, children aged 6–16 months received 3 servings, and children aged 12–36 months received 2 servings). | 5 days | Children: 52 children with persistent diarrhea (duration > 13 days but <29 days); randomly assigned to one of two groups; yogurt (n = 25) and milk (n = 27); age 3–36 months | Clinical failure was observed in 42% of children receiving milk and 14% receiving yogurt. Children consuming yogurt gained weight despite lower energy intake, had less liquid stools, and required less oral rehydration solution than children receiving milk. | Boudraa et al., 1990 [128] |
| Yogurt prepared from milk formulae vs. full-strength milk formulae | Sc. thermophilus and Lb. bulgaricus | 120 mL/kg body weight in seven divided feedings | 72 h | Children: 96 malnourished boys; randomly assigned to one of two groups; yogurt (n = 47) and milk (n = 49); age 4–47 months | The treatment failure rate was similar in both groups. Children who consumed milk had more weight gain at the end of the study and after recovery. Yogurt for malnourished children with acute diarrhea has no significant clinical benefit over milk. | Bhatnagar et al., 1998 [129] |
| Standard yogurt vs. Fermented milk with yogurt cultures and Lb. casei vs. Jellied milk (control group) | L casei 1 × 108 CFU/mL | One of three products 125 g or 250 g according to age | Three periods of 1 month, followed by 1 month without intervention | Children: 287 children with acute diarrhea over a 6-month observation period; mean age of 18.9 ± 6 months | The incidence of diarrhea was not different between the groups. The severity of diarrhea significantly decreased with the supplementation of L. casei fermented milk compared with the jellied milk. | Pedone et al., 1999 [130] |
| Pasteurized yogurt and routine hospital care vs. Routine hospital care (control group) | Lb. bulgaris 5 × 104 /mL and Sc. thermophilus 5 × 104/mL | 15 mL/kg/day | Until hospital discharge | Children: 80 children with moderate dehydration and acute non-bloody, non-mucoid diarrhea; randomly assigned to one of two groups; yogurt (n = 40) and control group (n = 40); aged 6–24 months | Children receiving yogurt observed a reduction in the frequency of diarrhea, fewer days in the hospital, and more weight gain compared to the control group. | Pashapour and Iou, 2006 [131] |
| Fluid yogurt prepared from commercial yogurt vs. Lyophilized Saccharomyces boulardii | 1 × 107 CFU/100 mL of Lb. bulgaricus and S. thermophilus (yogurt group) | Yogurt group: 15 mL twice a day for children < 2 years and 30 mL twice a day for children ≥ 2 years Lyophilized Saccharomyces boulardii group: 250 mg twice a day in children ≥ 2 years and 125 mg twice a day in children < 2 years of age | Until the resolution of the diarrhea | Children: 55 children with diarrhea; randomly assigned to one of two groups; yogurt (n = 27) and lyophilized Saccharomyces boulardii (n = 28); age 5 months–16 years | The effect of yogurt was comparable with that of lyophilized Saccharomyces boulardii in the treatment of acute diarrhea | Eren et al., 2010 [132] |
| Ulcerative Colitis (UC) | ||||||
| Bifidobacteria-fermented milk vs. control group | 1 × 1010 CFU of Bifidobacterium breve, and Bifidobacterium bifidum, and Lb. acidophillus YIT 0168 | 100 mL/day | 1 year | Adults: 21 patients with UC remission; randomly assigned to one of two groups; study group (n = 11), control group (n = 10); age 39–60 years | Significant reduction in exacerbation of symptoms after bifidobacteria fermented milk supplementation. Reduction in the percentage of Bacteroides vulgatus and luminal butyrate and good recovery of probiotic strains in the stools. Increases in protein and albumin levels. | Ishikawa et al., 2003 [133] |
| Bifidobacteria fermented milk vs. Fermented milk without live bifidobacteria (control group) | ≥1 × 1010 CFU of Bifidobacterium bifidum strain Yakult and Lb. acidophillus strain | 100 mL/day | 12 weeks | Adults: 20 patients with active UC, randomly assigned to one of two groups; study group (n = 10), control group (n = 10); mean age of 30.2 years for the study group and 33.7 years for the control group | Increase in probiotic strains and butyrate in the feces. Improved clinical activity index; endoscopic activity index and histological scores compared to the control group. | Kato et al., 2004 [134] |
| Fermented milk product (Cultura) | 1 × 108 CFU/mL milk Lb. acidophilus La-5 and B. lactis BB-12 | 500 mL | 4 weeks | Adults: three groups: UC group with ileal-pouch-anal-anastromosis (n = 51, mean of age 40 years), familial adenomatus polyposis with ileal-pouch-anal-anastromosis (n = 10, mean of age 35 years) and UC with ileorectal anastromosis (n = 6, mean of age 42 years) | Increased number of lactobacillus and bifidobacterium in the UC patients with ileal-pouch-anal-anastromosis and remained increased one week after intervention. No significant changes in blood tests (antinuclear antibody and antineutrophil autoantibodies), fecal fungi and fecal pH. | Laake et al., 2005 [135] |
| Fermented milk products with Bifdobacterium breve strain Yakult | 1 × 1010 CFU of Bifidobacterium breve Lb. acidophilus and 1 × 109 CFU of Lb. acidophilus | One pack (100 mL) of commercial B. breve strain Yakult fermented milk (Mil–Mil) | 48 weeks | Adults: 195 Japanese patients with quiescent UC; study group (n = 98) and placebo group (n = 97); aged 20–70 years | Bifidobacterium breve strain Yakult did not affect the time to relapse in UC patients compared with the placebo group. | Matsuoka et al., 2018 [136] |
| Irritable bowel syndrome (IBS) | ||||||
| Probiotic fermented yogurt drink vs. Placebo (the same product without lactic acid fermented bacteria) | 4 × 109 CFU of Lb. sp. HY7801, Lb. brevis HY7401, and Bifidobacterium longum HY8004 | One bottle (150 mL) of a probiotic yogurt drink, 3 times/day, within 10 min after breakfast, lunch, and dinner | 8 weeks | Adults: 74 IBS patients from the Republic of Corea; randomly assigned to one of two groups; study group (n = 37) and placebo group (n = 37); range age of 33 years | The amount of Lactobacilli species, which were included in the yogurt drink, significantly increased in the feces of IBS patients receiving treatment. Serum glucose and tyrosine levels in IBS patients were normalized to those of healthy individuals in the study group. | Hong et al., 2011 [137] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okoniewski, A.; Dobrzyńska, M.; Kusyk, P.; Dziedzic, K.; Przysławski, J.; Drzymała-Czyż, S. The Role of Fermented Dairy Products on Gut Microbiota Composition. Fermentation 2023, 9, 231. https://doi.org/10.3390/fermentation9030231
Okoniewski A, Dobrzyńska M, Kusyk P, Dziedzic K, Przysławski J, Drzymała-Czyż S. The Role of Fermented Dairy Products on Gut Microbiota Composition. Fermentation. 2023; 9(3):231. https://doi.org/10.3390/fermentation9030231
Chicago/Turabian StyleOkoniewski, Adam, Małgorzata Dobrzyńska, Paulina Kusyk, Krzysztof Dziedzic, Juliusz Przysławski, and Sławomira Drzymała-Czyż. 2023. "The Role of Fermented Dairy Products on Gut Microbiota Composition" Fermentation 9, no. 3: 231. https://doi.org/10.3390/fermentation9030231
APA StyleOkoniewski, A., Dobrzyńska, M., Kusyk, P., Dziedzic, K., Przysławski, J., & Drzymała-Czyż, S. (2023). The Role of Fermented Dairy Products on Gut Microbiota Composition. Fermentation, 9(3), 231. https://doi.org/10.3390/fermentation9030231