Abstract
Understanding the energy metabolism and its regulation is one of the clues to metabolic engineering of stress-resistant lignocellulose-converting microbial strains, also including the promising ethanologen Zymomonas mobilis. Z. mobilis is an obligately fermentative, facultatively anaerobic bacterium, carrying an active respiratory chain with low energy-coupling efficiency. Its respiration does not supply energy to aerobically growing cultures on sugary media, yet oxidative phosphorylation has been demonstrated in non-growing cells with ethanol. Here, we show, for the first time, that in respiring, non-growing Z. mobilis cells receiving regular small amounts of ethanol, oxidative phosphorylation significantly contributes to the maintenance of their viability. No improvement of viability is seen in the NADH dehydrogenase (ndh)-deficient respiratory mutant, which is unable to oxidize ethanol. The ethanol effect is also hampered by the protonophoric uncoupler CCCP, or the inhibitor of ATP synthase, DCCD. At higher concentrations (6% v/v), ethanol causes stress that slows down culture growth. By monitoring the activity of several respiratory gene promoters under ethanol stress with the green fluorescent protein reporter system, we demonstrate downregulation of these promoters, in particular the ndh promoter. We speculate that the decrease in respiratory chain activity in response to stress conditions mitigates the production of reactive oxygen species.
1. Introduction
Zymomonas mobilis is an ethanologenic, facultatively anaerobic alpha-proteobacterium. It possesses the most rapid homoethanol fermentation pathway among those of microorganisms [1], and apart from that, it can tolerate high sugar and ethanol concentrations [2,3]. These properties, in combination with a small genome size and relatively simple central metabolism, have put Z. mobilis in the focus of metabolic engineering work on the production of bioethanol and several other commodity chemicals from renewable substrates, primarily from lignocellulose hydrolysates [4,5,6,7,8]. Lignocellulose hydrolysates contain several types of inhibitory compounds; therefore, resistance to inhibitor stress, and more generally, the robustness of producer strains, is critical for lignocellulose bioconversion [5]. Generally speaking, robustness results from a complex network of physiological properties, yet in bacteria much of it is related to cellular energy management, and more specifically, to the function of respiratory chain [9,10,11] that has been shown to impact various types of stress resistance [12,13,14]. Understanding the respiratory energy metabolism and its regulation is one of the clues to metabolic engineering of stress-resistant lignocellulose-converting microbial strains.
While fermentative energy generation in Z. mobilis takes place in the Entner–Doudoroff pathway, and is fairly well understood, this is not the case with its respiratory energetics. Z. mobilis bears a constitutive respiratory chain (see Figure 1). Its major electron pathway involves the Type II NADH dehydrogenase, coenzyme Q10, and the cytochrome bd terminal oxidase. FAD-dependent D-lactate dehydrogenase and PQQ-dependent glucose dehydrogenase (Gdh) are minor dehydrogenase activities [15,16,17,18]. Cytochrome bd is the only terminal oxidase encoded in the genome [19,20] and detected spetroscopically [16,18]. Respiratory peroxidase, identified in the genome as a cytochrome c peroxidase (PerC), most likely functions as a quinol peroxidase [21]. The cytochrome bc1 branch of electron transport seems to be a ‘dead-end’, lacking a terminal cytochrome c oxidase, and hence, its function remains obscure.
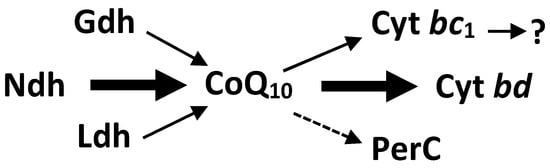
Figure 1.
Electron transport chain of Zymomonas mobilis. Ndh, type II NADH dehydrogenase; Ldh, lactate dehydrogenase; Gdh, glucose dehydrogenase; PerC, cytochrome c peroxidase; cyt bd, cytochrome bd terminal oxidase; Cyt bc1, cytochrome bc1 complex.
For some unknown reason, the energetic efficiency of Z. mobilis respiration in aerobically growing culture is much lower than that in other bacteria (e.g., L. lactis) that employ a similar electron transport route from the Type II NADH dehydrogenase to cytochrome bd terminal oxidase [9,22,23]. Although Oxidative phosphorylation capacity has not been demonstrated to have non-growing cells and membrane vesicles, it has been demonstrated in non-growing cells and membrane vesicles [24], aeration does not contribute to its biomass yield [25] (for a review, see [26]), making its aerobic energetics quite unique among those of the known biotechnological producer microorganisms. Furthermore, respiration hampers its aerobic growth by disturbing the fermentative redox balance. In the absence of a functional TCA cycle, the respiratory chain withdraws NADH mostly from the alcohol dehydrogenase reaction, thus causing the accumulation of the inhibitory metabolite acetaldehyde. Lowering Z. mobilis respiration decreases acetaldehyde accumulation, and improves aerobic growth and ethanol synthesis, as seen in the ndh knock-out mutant growing on a rich medium [17,27,28,29]. The complementation of this mutant strain with an Ndh-expressing plasmid vector restores its respiratory activity, and accordingly, its aerobic growth decreases to the level of that of the parent strain. These observations highlight a problem: for what purpose has Zymomonas mobilis preserved its respiratory chain, given that it is inhibitory for its own aerobic growth?
To understand the physiological rationale behind Z. mobilis respiration, it is helpful to compare its metabolism with that of other ethanologenic microorganisms, primarily with that of yeast. Some yeast species, including Saccharomyces cerevisiae, are Crabtree-positive, which means that in sugar-rich environments they perform fermentation along with respiration even under vigorous oxygenation [30]. Consuming sugar faster than other species, converting it into ethanol to inhibit the growth of competing bacteria, and then taking up the accumulated ethanol as the carbon and energy substrate after they have established dominance in their niche, is a growth strategy termed ‘make-accumulate-consume’ (MAC) [31,32]. Z. mobilis converts sugar into ethanol even faster than does the ethanologenic yeast [1], and is equally resistant to high ethanol concentrations. However, it is an obligately fermentative bacterium that can oxidize ethanol, but unlike yeast, cannot use it as a carbon source for growth [3]. Since respiration with ethanol in non-growing cells produces ATP via oxidative phosphorylation [24], we hypothesize that the Z. mobilis version of the MAC strategy might be to employ their accumulated ethanol merely as an oxidizable energy substrate, used to maintain the viability of non-growing cells. The MAC strategy can have also its disadvantages: although ethanol inhibits the growth of competing microorganisms and serves as an oxidizable energy source, at higher concentrations it causes stress to the producer organism itself [33,34]. How the respiratory chain functions under condition when Z. mobilis (if grown on a sugar rich media) accumulates high amounts of ethanol is thus another question related to its putative MAC strategy. In the present study, we demonstrate that ethanol can serve as a respiratory energy source for the maintenance of viability, yet under ethanol stress conditions, Z. mobilis partly turns down its respiratory chain activity.
2. Materials and Methods
2.1. Plasmid and Strain Construction
The plasmids and strains used in this study are listed in Table 1. For the monitoring of the promoter activity of Z. mobilis respiratory genes, reporter systems with engineered green fluorescent protein (EGFP) were constructed. The gene egfp was amplified via PCR, taking plasmid pQE-EGFP as the template. The PCR product was double-digested with HindIII and EcoRI restriction enzymes, and inserted into the MCS of pBBR1MCS-2 between the respective sites, yielding plasmid pBBR-EGFP. After that, chromosomal DNA sequences of the strain Zm6 (ATCC 29191), containing the promoter regions of cytochrome c peroxidase, cytochrome bc1 operon, cytochrome bd operon, and the type II NADH dehydrogenase, were amplified. Sequence data of the amplified DNA fragments and the corresponding primers are given in Table A1. These PCR products were double-digested and inserted into pBBR-EGFP directly upstream of egfp, between the EcoRI and BamHI sites of its MCS, yielding the four reporter plasmids listed in Table 1. Genomic DNA isolation from Z. mobilis and PCR assays, as well as DNA purification and cloning techniques were the same as those described before [17,35]. Escherichia coli strain DH5α was used for plasmid cloning. The obtained reporter plasmid constructs were verified via restriction analysis, and via PCR (not shown), using primer pairs consisting of an F primer for the respective promoter region and R primer for egfp (see Table A1). Finally, Z. mobilis Zm6 was transformed with pBBR-EGFP and with the four reporter plasmids via electroporation [36]. Successful transformations were verified via colony PCR.

Table 1.
Plasmids and strains used in this study.
2.2. Cultivation, Starvation and Assessment of Viability
Z. mobilis strains were maintained and cultivated at 30 °C, without aeration, in the medium containing (per 1 L of distilled water) 50 g of glucose, 5 g of yeast extract, 1 g of KH2PO4, 1 g of (NH4)2SO4, and 0.5 g of MgSO4 × 7H2O, at pH 5.8 [24]. Overnight cultivations of mutant/recombinant strains carrying antibiotic resistance markers were carried out with added antibiotics (chloramphenicol, in an amount of 120 μg mL−1, and/or kanamycin, in an amount of 310 μg mL−1, where required) [23,35]. For Z. mobilis electroporation, RM growth medium containing (per 1 L of distilled water) 20 g of glucose, 10 g of yeast extract, and 2 g of KH2PO4 was used. E. coli DH5α was grown on LB medium, containing (per liter) 10 g of tryptone, 5 g of yeast extract, and 10 g of NaCl. The medium for plasmid-bearing E. coli strains was supplemented with 60 μg mL−1 of kanamycin. For platings, all media were supplemented with 2 g L−1 of agar.
For starvation experiments, three strains (Table 1), (i) the wild type Zm6, (ii) the Ndh knock-out mutant strain Zm6-ndh, and (iii) the mutant strain complemented with a plasmid vector, carrying ndh under its native promoter, Zm6-ndh_pNdh, were cultivated overnight without shaking in a thermostat at 30 °C. The cells were harvested via centrifugation, washed and resuspended in 0.1 M potassium phosphate buffer (pH 7), supplemented with 5 mM MgSO4, to yield an OD550 around 10 ± 0.5 (equivalent to 2 mg dry wt mL−1). Briefly, amounts of 10 mL of the cell suspensions were transferred into 50 mL shaken flasks, and incubated on a shaker at 100 r.p.m., at 30 °C, for 6 h. To assess the cell viability, 50 µL samples were taken from the flasks at time zero and after the 6 h period. These samples were diluted with distilled water to an OD550 of 0.5, and for each experimental condition, four technical replicates of serial dilutions (from 100 to 10−4) were produced and spotted on agar plates in 5 µL aliquots. To determine the statistical significance of the difference between the outcomes of various experimental series, experiments were compared using the nonparametric Mann–Whitney U test (Wilcoxon rank sum test) based on the ranking of growth on the plates.
2.3. Cultivation of EGFP-Bearing Strains in the Microplate Reader and Monitoring of EGFP Fluorescence
The EGFP-bearing Z. mobilis strains were cultivated in a microplate (96-well plate) reader, Infinite M200 PRO Multimode Microplate Reader (Tecan, Maennedorf, Switzerland), at 30 °C, for 15 h. Each well contained 200 μL of culture. The plates were shaken at 200 r.p.m., with a 3.5 mm amplitude, in 490 s periods with interruptions for measurements. Optical density (at 600 nm) and fluorescence (excitation at 475 nm; emission at 510 nm) measurements were taken in 10.5 min intervals. Each experiment was carried out in at least 2 technical replicates.
2.4. FTIR Spectroscopy of Biomass
FTIR spectra of Z. mobilis biomass were recorded using Vertex 70 coupled with the microplate reader HTS-XT (Bruker, Germany). Spectra were recorded in the frequency range of 4000–600 cm−1, with a spectral resolution of 4 cm−1, and 64 scans were coadded. Only spectra with absorbance within the absorption limits between 0.30 and 1.00 (where the concentration of a component is proportional to the intensity of the absorption band) were used for analysis. Data were processed using OPUS 7.5 software (Bruker Optics GmbH, Ettlingen, Germany). The baseline of each spectrum was corrected using the rubber band method. The macromolecular composition of biomass was calculated as described previously [37].
2.5. Preparation of Cytoplasmic Membranes and Respiratory Chain NADH Oxidase Assay
For assaying respiratory NADH oxidase activity, cytoplasmic membrane vesicles were prepared from cells, grown aerobically in 250 mL shaken flasks (with a 100 mL culture volume) at 100 r.p.m. and 30 °C, and harvested after 8 h of cultivation, during their transition from the late exponential to early stationary growth phase. The harvesting of cells from a 100 mL culture volume via sedimentation, washing, and resuspension in 0.1 M potassium phosphate buffer with 5 mM MgSO4, at pH 7 (yielding a cell suspension with a 4 g dry wt L−1 biomass concentration), was followed by ultrasonic breakage and the removal of unbroken cells, and finally, the cell membrane fraction was pelleted via ultracentrifugation, basically following the procedures described previously [16,24]. Sedimentation was conducted in Thermo Scientific Sorvall WX+ Ultracentrifuge at 35,000 r.p.m. for 1 h, the pellet was washed with phosphate buffer, and the membranes were resuspended and centrifuged repeatedly for 1 h at the same speed. Finally, the pellet was resuspended in the same buffer to yield membrane vesicle suspension with the protein concentration in the range of 3–4 mg mL−1. Protein concentration was determined as previously described [16]. The NADH oxidase assay mixture contained 980 µL of 0.1 M phosphate buffer, at pH 7, 10 µL of 10 mM NADH, and 10 µL of the membrane vesicle suspension. NADH oxidase activity was measured by monitoring the decrease in NADH absorbance at 340 nm, taking 6.22 mM−1cm−1 as the millimolar extinction coefficient.
3. Results
3.1. The Impact of Respiratory Chain Activity on the Viability of Non-Growing Z. mobilis Cells
In the early stage of research on Z. mobilis (formerly, also Z. anaerobia; see [38]) physiology, Dawes and Large [39] showed that in a starving cell population, viability decreased exponentially as soon as starvation began, as might be expected since Zymomonas anaerobia contains no energy reserves to protect against starvation. The decrease in viability was accompanied by an exponential decrease in intracellular ATP which was most marked during the initial 6 h. Our FTIR spectroscopy data on the macromolecular constituents of the parent strain Zm6 before and after a 6 h starvation period (Table A2) supported the notion that Z. mobilis cells did not employ endogenous energy reserves; the spectra revealed fairly constant relative amounts of the bulk intracellular carbohydrate, nucleic acids, lipids, and protein. Accordingly, we concluded that viability should mostly depend on the external energy source. For Z. mobilis, when it runs out of carbohydrates in its growth medium, ethanol might be the energy source of choice. Here, we chose a 6 h starvation period to examine if ethanol oxidation could be used for energy production to maintain the viability of non-growing, respiring Z. mobilis cells.
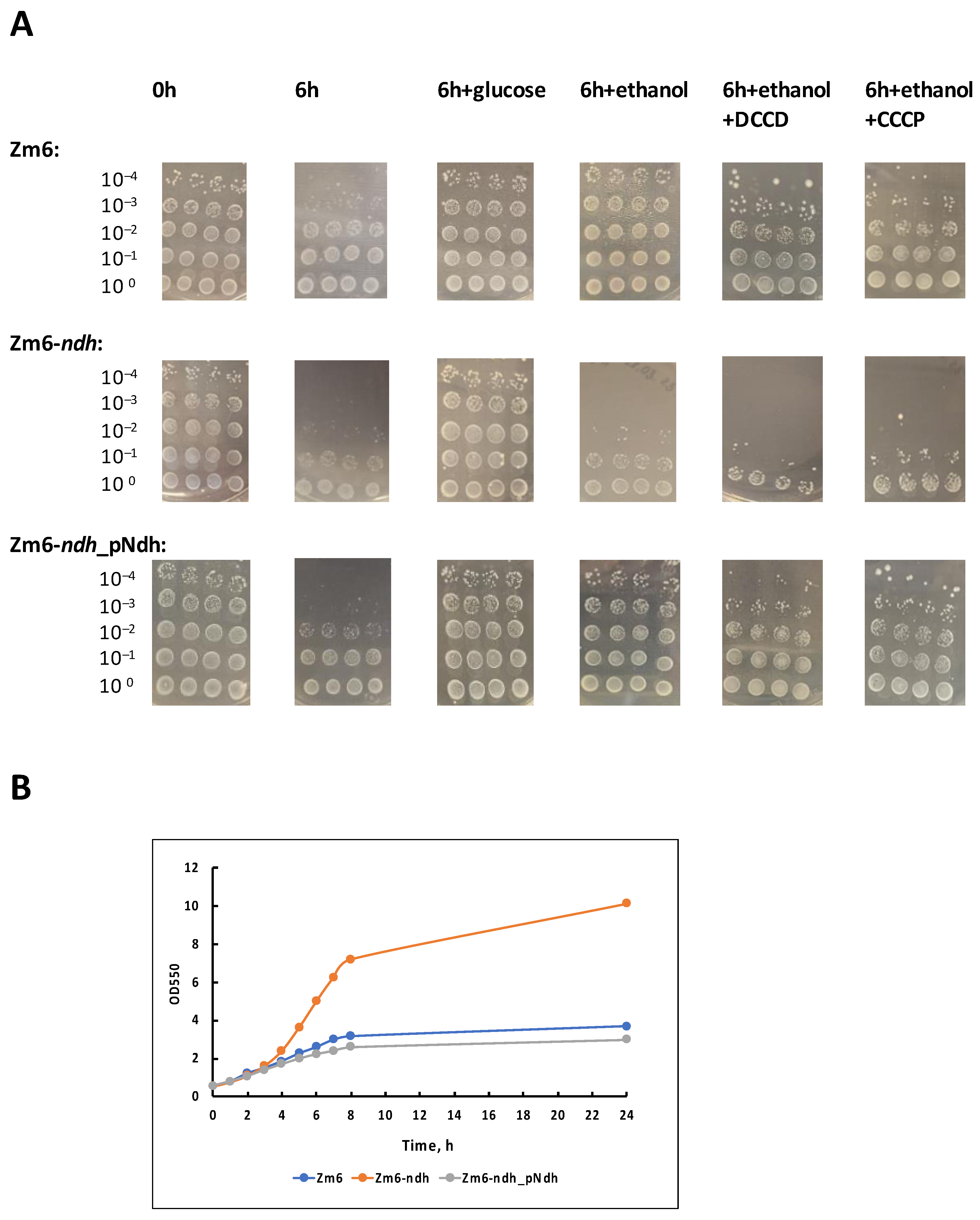
The results are presented in Figure 2A. Without the supply of an external energy source, there was a significant drop in viability after the 6 h starvation period for all strains. In the experimental series ‘6 h + glucose’, the cell suspensions were supplemented with small amounts of glucose once per hour, yielding a concentration of 1 g L−1 in the medium. Glucose was taken up within approx. 15 min (not shown), and hence, most of the time cells were starving. Nevertheless, for all strains, this amount of energy substrate proved sufficient to maintain their viability. In the experimental series ‘6 h + ethanol’, cells with the same regularity were supplied with ethanol in 5 g L−1 increments. Regular ethanol addition maintained viability in the respiring strains Zm6 and Zm6-ndh_pNdh to the same extent as did glucose supplements, but in Zm6-ndh, ethanol had no positive effect—a striking contrast to the aerobic growth observed on a rich medium (Figure 2B), where the mutant Zm6-ndh outperformed the other two strains by far.
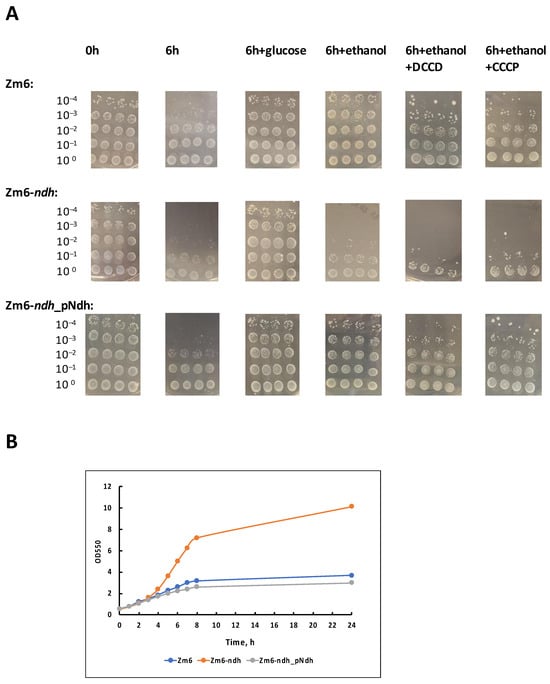
Figure 2.
Viability of aerobically incubated non-growing cells (A) and aerobic batch growth (B) of the strains Zm6, Zm6-ndh, and Zm6-ndh_pNdh. In (A), a representative set of spot tests is shown. The 6 h starvation experiment, and incubations with the addition of glucose or ethanol were conducted in four biological replicates (each with 4 technical repeats); incubations with added DCCD or CCCP were conducted in three biological replicates. According to the Mann–Whitney U test, statistically significant differences (p < 0.05) in viability were observed between the following samples: 0 h vs. 6 h for all strains; 6 h vs. 6 h + glucose for all strains; 6 h vs. 6 h + ethanol for Zm6 and Zm6-ndh_pNdh; 0 h (or 6 h + glucose) vs. 6 h + ethanol (also, 6 h + ethanol + DCCD, or +CCCP) for Zm6-ndh; 6 h + ethanol for Zm6-ndh vs. 6 h + ethanol for the other two strains; 6 h + ethanol vs. 6 h + ethanol + CCCP for Zm6. (B) A representative time course of the aerobic batch growth of the strains in the medium with glucose in 200 mL shaken flasks with 25 mL of culture, at 180 r.p.m. and 30 °C.
We have shown previously that non-growing respiring cells of Z. mobilis can perform oxidative phosphorylation using ethanol as a substrate [24]. The present finding strongly indicates that in Z. mobilis strains, which are able to oxidize ethanol, oxidative phosphorylation can serve as the energy source for the maintenance of their viability. Because of the lack of NADH dehydrogenase in its respiratory chain, the mutant Zm6-ndh cannot oxidize ethanol, and accordingly it cannot use ethanol for energy production via oxidative phosphorylation to maintain its viability. With glucose, on the other hand, the bulk of ATP comes from the fermentative catabolism that is roughly similar between all three strains.
To reinforce the proposed role of oxidative phosphorylation for the maintenance of viabilty, we performed an inhibitor analysis of the ethanol effect with the F1Fo-ATP synthase inhibitor N,N′-dicyclohexylcarbodiimide, DCCD (Figure 2A, series ‘6 h + ethanol + DCCD’), and the protonophoric uncoupler carbonyl cyanide m-chlorophenylhydrazone, CCCP (series ‘6 h + ethanol + CCCP’). DCCD at a 50 µM final concentration, or CCCP at a 10 µM final concentration, was added to the cell suspensions shortly before the start of incubations with the periodical ethanol supply. For the strains Zm6 and Zm6-ndh_pNdh, these inhibitors hampered the ethanol effect. A decrease in viability was seen with both inhibitors and in both strains. Yet, the most pronounced, statistically significant effect (p < 0.05) was obtained for Zm6 with CCCP.
3.2. Expression of the Respiratory Chain Complexes under Ethanol Stress
As we show here, at low ethanol concentrations, respiring Z. mobilis uses it as an energy source for the maintenance of viability. Yet, at higher concentrations, ethanol causes stress by inhibiting culture growth and metabolism. We assumed that monitoring the ethanol stress-induced changes of the expression pattern of Z. mobilis respiratory chain modules might help to draw conclusions about their involvement in ethanol stress protection. In order to see how the respiratory chain components responded to ethanol stress, we examined recombinant strains carrying the EGFP reporter plasmids with four respiratory gene promoters (see Table 1). Ethanol stress was induced via the addition of ethanol to the growth medium at a 6% (v/v) concentration, as described elsewhere [33,34], and cultivation was carried out in a microplate reader allowing the simultaneous recording of the culture OD and fluorescence. Ethanol in this concentration already significantly hampered culture growth (Figure 3, inset).
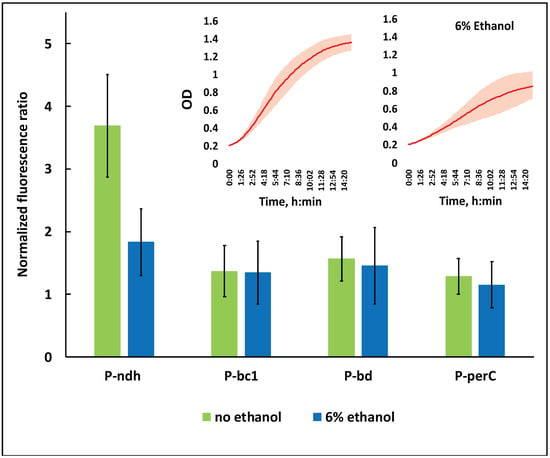
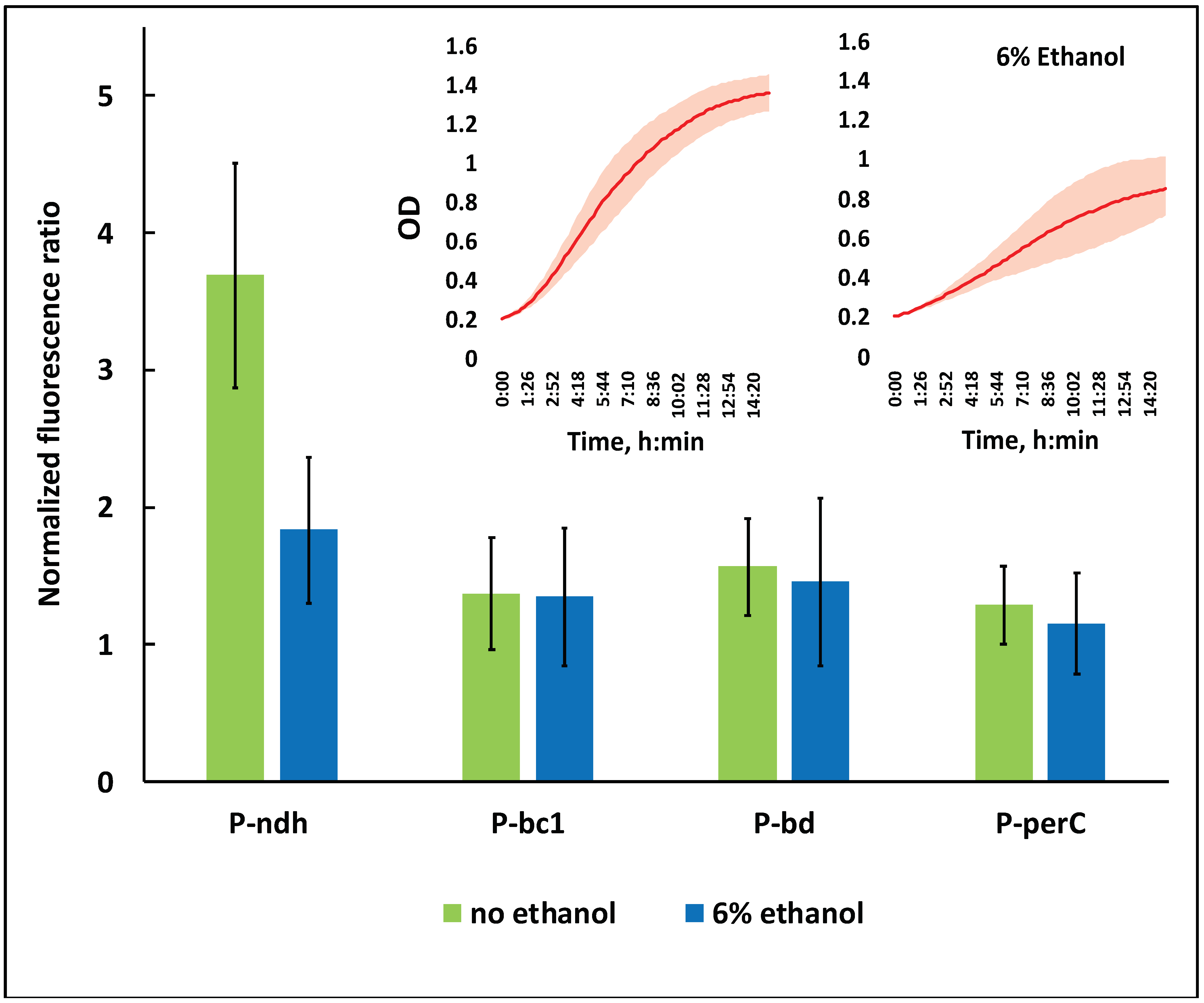
Figure 3.
The effect of ethanol stress on the relative activity of respiratory gene promoters of strain Zm6. P-ndh, type II NADH:ubiquinone oxidoreductase promoter; P-bc1, promoter of cytochrome bc1 operon; P-bd, promoter of cytochrome bd operon; P-perC, cytochrome c peroxidase promoter. Inset: the time course of batch growth of the strain Zm6_pBBR-EGFP in the microplate reader under non-stress (left side curve) and ethanol stress (right side curve) conditions. All presented values are the means of 8 biological replicates (non-stress condition) or 5 biological replicates (stress condition). For details on the calculation of the normalized fluorescence ratio, see the text.
EGFP fluorescence data in Figure 3 correspond to the 8th hour of cultivation in the microplate reader, when cultures were entering the early stationary growth phase. First, for each strain, the recorded fluorescence value was normalized per unit of its culture optical density. Since the normalized fluorescence of the strain Zm6_pBBR-EGFP was identical to that of Zm6 throughout the whole range of culture OD values (not shown), we concluded that EGFP did not become expressed from the plasmid pBBR-EGFP. The normalized fluorescence of Zm6_pBBR-EGFP was therefore taken as the baseline, implying that all of the fluorescence exceeding that level in the four other EGFP-bearing strains resulted from the activity of the respective respiratory gene promoters. Accordingly, the ‘normalized fluorescence ratio’ shown in Figure 3 denotes the ratio of the normalized fluorescence of any promoter-carrying strain to the normalized fluorescence of strain Zm6_pBBR-EGFP. A ratio of 1.0 on this scale would mean a complete lack of promoter activity.
As seen from Figure 3, under non-stressed conditions, all promoters were active. For all of them, the normalized fluorescence ratio was significantly above 1.0 (p < 0.05). The ndh promoter, however, stood out by being several times more active than were any of the three other promoters. Under ethanol stress, its activity was reduced by more than half, while only a slight (statistically not significant) decrease was seen for the rest of promoters. The perC promoter was the weakest of all. Its normalized fluorescence ratio was already the closest to 1.0 under non-stressed condition, but under ethanol stress it showed no statistically significant activity. For the ndh promoter, our data fell in line with previously published transcriptomic findings, demonstrating the downregulation of ndh under ethanol stress [33]. Yet, we did not see any significant effects with the cytochrome bd and cytochrome bc1 promoters, although the upregulation of bd and downregulation of bc1 transcripts were reported in the same study.
NADH oxidase activity in the membrane preparation of non-stressed Zm6_pBBR-EGFP was 0.64 (±0.08) U mg prot−1, while under ethanol stress it was down to only 0.28 (±0.05) U mg prot−1. This corresponded well to the observed decrease in promoter activity, especially with respect to the ndh promoter. Altogether, the downregulation of the respiratory gene promoters and the resulting decrease in NADH oxidase activity indicate that Z. mobilis does not cope with the ethanol stress via the activation of its respiratory chain or any of its branches, but rather reacts to that condition via a partial shutdown.
4. Discussion
In the present study, we show that the respiratory chain of Zymomonas mobilis has an important physiological function for keeping non-growing cells viable by driving oxidative phosphorylation with ethanol, the Zymomonas major catabolic product. Thus, Z. mobilis largely follows the MAC strategy of the Crabtree-positive yeast, implying that it consumes ethanol, ‘C’, specifically for the maintenance of viability, but not for growth. Sugar-depleted media, exposed to air, and containing some amount of ethanol, probably represent a common niche in the natural life cycle of this bacterium. Previously, we identified another advantage of respiration for the survival of Z. mobilis in the same type of niche. As shown by Rutkis et al. [23], after glucose is added to a starving Z. mobilis cell suspension, the respiring strain takes it up more rapidly than does the Ndh-deficient mutant. Entner–Doudoroff pathway simulations indicate that it is the oxidation of extra NAD(P)H by the respiratory chain that accelerates pathway flux, conferring a competitive advantage to the respiring strain, when a limited amount of glucose suddenly becomes available. Furthermore, Jones-Burrage et al. [40] have found that on minimal growth medium under oxic conditions, the respiring wild-type strain ZM4 grows better than does its Ndh-deficient mutant derivative—quite the opposite to what is seen on rich growth medium. In summary, survival from sugar deficiency and, perhaps, some other limitations in the growth medium is a distinct physiological domain where Z. mobilis makes use of its respiratory chain. The advantage provided by the respiratory chain for survival in sugar-depleted media might bear such high adaptive priority for this bacterium that it cancels out the disadvantage for its aerobic growth on rich medium.
We also wanted to see if the respiratory chain of Z. mobilis was involved in the protection of the cell under ethanol stress conditions. It has been demonstrated that various stressors in microorganisms induce a burst of reactive oxygen species (ROS), leading to secondary oxidative stress [41]. Elevated ROS production also takes place under ethanol stress, as reported, e.g., for E. coli and S. cerevisiae [42,43]. Accordingly, a respiratory chain that mitigates ethanol stress might be expected to carry this out primarily by coping with the associated ROS production. Some evidence has been accumulated on the role of Z. mobilis respiratory chain in oxidative stress defense. Martien et al. [44] reported the upregulation of several respiratory chain components after exposing a strictly anaerobic culture of Z. mobilis Zm4 to vigorous aeration. They suggested that cytochrome c peroxidase and cytochrome bd function to limit the toxic effects of oxygen. In support of that, the cytochrome c peroxidase-negative strains of Z. mobilis Zm6 and Z. mobilis TISTR 548 are shown to be hypersensitive to exogenously added hydrogen peroxide [21,45], pointing to protective activity of the respiratory peroxidase branch (Figure 1).
However, the relation of the respiratory chain to intracellular ROS levels is fairly complex. Along with activities such as those of cytochome c peroxidase that remove hydrogen peroxide, electron transport itself is a major source of ROS [46]. At a higher degree of reduction of its electron carriers, ROS production in the respiratory chain tends to increase [47]. We hypothesize that the downregulation of the respiratory promoters and the decrease in NADH oxidase activity that we see under ethanol stress are aimed primarily at minimizing respiratory ROS production. Since the ndh promoter is downregulated much more than the rest are, the ‘entrance gates’ of the electron transport chain might become the main bottleneck of electron transport flux, while the capacity of the terminal oxidase remains almost unaffected. This should result in a more oxidized steady state of respiratory electron carriers, and hence, in a lower rate of respiratory ROS generation. Such reasoning seems to be in line with the recent findings of Felczak and TerAvest [48]. They reported that, while the decrease in the respiration rate due to the deletion of ndh in the strain ZM4 stimulated aerobic growth, a similar decrease in respiration attained in result of the deletion of cytochrome bd had an opposite effect. One might speculate that in the ndh mutant, the steady state degree of reduction in electron carriers (and accordingly, ROS production) is much lower than that in the terminal oxidase-deficient strain. This might have contributed to the observed difference in their aerobic growth patterns. This hypothesis, of course, needs experimental verification via the monitoring of the redox state of electron carriers (primarily, that of ubiquinone) and measurement of the rate of respiratory ROS production.
5. Conclusions
In the present study, we show that non-growing Zymomonas mobilis employs its respiratory chain for oxidative phosphorylation to survive in sugar-depleted media that contain its catabolic product ethanol. At the same time, its respiratory chain seems unlikely to provide any specific protection against stress caused by elevated ethanol concentrations. Altogether, the physiology and regulation of Zymomonas mobilis respiration under stress conditions requires further research, as it might reveal novel, non-conventional functions of bacterial respiration. A better understanding of the alternative roles of the respiratory chain is important both for the rational metabolic engineering of this bacterium, as well as for microbial biotechnology in general.
Author Contributions
Conceptualization, U.K. and I.S.; methodology, I.S., R.R. and M.G.; software, R.R. and K.S.; validation, M.B., E.R.P. and U.K.; formal analysis, U.K.; investigation, I.S., M.B., E.R.P., K.S. and Z.L.; resources, R.R.; data curation, I.S., M.G. and U.K.; writing—original draft preparation, M.B., E.R.P. and U.K.; writing—review and editing, U.K. and M.G.; visualization, M.B. and E.R.P.; supervision, U.K. and I.S.; project administration, R.R.; funding acquisition, R.R. and U.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Latvian State Education Development Agency ERDF, project no. 1.1.1.1/19/A/097.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
The amplified promoter regions of the Z. mobilis Zm6 respiratory chain components, the egfp sequence, and the respective primer sequences used in the study. Sequences of the structural gene ORFs are shown in blue. The primer binding regions are given in bold. Sequences, edited to introduce restriction sites, are underlined.
Table A1.
The amplified promoter regions of the Z. mobilis Zm6 respiratory chain components, the egfp sequence, and the respective primer sequences used in the study. Sequences of the structural gene ORFs are shown in blue. The primer binding regions are given in bold. Sequences, edited to introduce restriction sites, are underlined.
| Gene/Operon Names, and Sequence i.d. | Sequences of the Amplified Chromosomal Regions, Bearing the Promoters of the Respective Genes/Operons (5’ → 3’) | Forward and Reverse Primer Sequences (5’ → 3’) |
|---|---|---|
| Cytochrome c peroxidase perC ZZ6_0192 | →ZZ6_0191 CCGATGAATGGGTAAAATAAGACCTTTTCCCTTTCTAAAATTTCAGAGGTGGCTTTATTC CTGTTAAAGCCACCTTTTATTTTATCTTTTTCTTTATTCTTTCTCAGCTTTTCTGATTAA AAAATTTTGTAAATATAGCCAATATTTCCAATGAAATATCAGGTTAAAATATCTGTAAAG AACGAGAATTTTCTTATCACTTTAATGATAACTATTGGCATCATCAAAATCATCAAGAAT TAATCAAAATTGAAATGACAGTTATGCCTTTTACTCTAAAAGGGATGTGCTTTCTGTCTT GGAATAAGGCCATATTTTTGTTAATTCACAATAAATAAAAAGGTAAATCATCTTCAATAG GGAATCGGATACTATAAATTTATAGCCGACCATCCGATCATCTCTGGCGCTGGATTAAGA TAACGAGGGGCAAGCACAGTTTTATGAATATCAAGGCTTTATT →ZZ6_0192 | F primer BamHI CCGATGAATGGGATCCATAAGACCTTTTCC |
| R primer EcoRI AGCCTTGATATTCATGAATTCGTGCTTGCC | ||
| Cytochrome bc1 (ubiquinol:cytc oxidoreductase) operon: FeS cluster (Rieske) subunit ZZ6_0349; Cyt b subunit ZZ6_0348; Cyt c1 subunit ZZ6_0347 | → ZZ6_0350 CTCGCCGCAACCGTCTTGTTCTGGCTATGAGTCCTGAGCCTAGATTTCATGATAGTCAAT ATATTAAAAAATTATATATTGTAGTAATCTTTCTTAAAAAATCAGAATAATTGATTTAAA TATCATTAATTATGGCATTTTAATTATCATTTTTAATATTTTCATGGACAATGAAGAAAA CTTATTATGTTTTTATTGTAGTAAGCGTTGCAAAAAATATTGTAAGTAATTTACAGTAGA GTAGGTTCGTCAAGGGATATGAAGAATAGCGAAGCGAA → ZZ6_0349 | F primer BamHI TTGTTCTGGCTATGGATCCTGAGCCTAGAT |
| R primer EcoRI TTCGCTATTCTTCATGAATTCTGACGAACC | ||
| Cytochrome bd operon: cydA ZZ6_1532; cydB ZZ6_1531 | ZZ6_1533 ← TCAAGGGTGGAAATGGTCATGTCGCTTCAACCTTTGATTCTATGAGCGAGGAAAGACTTA GGAGATATATCTAACAAAATTCGACCCGATCTGACAGCTATTATTAAAATGCAGAATAAG GTCGCGGGCTTTTTATCAGCTAACTCTAACAAAAAACAGAATCATTCGAAAAAGAATAAT AGAATACTCTATATTTAAATTATAAATTGATCCTTTTTCATATGGCTTTTTAATCACAAC CGCCCTAAAATCGGAAAATATCTGTTCCCTTGGGTGGTCTGTTATCAGAGATAGAAAATA ATTCGATTTCAAAAATCGTTATTTTCAATATTGAGACCTAAGCCCCTTTTCCCTTGTTCC TATCGTGACTTTCTTTTGAAATCGCCCCTATTCAACGAATTTAGGGGCAAGTTTTAAAAT AAAGCCCTTAATCGCTTCATCACTATTTTATTTAAAAAATGTTATTGATCTATAAAAAGA TACTATAACTACAGCAATATACTCAAAACAATCTGACATGGTCATATTAAATTTTAAAAT TTATCAAAAAAATAATTAAAATTAAAGAAATATAAAACGATAATTAAACCCAATTGAAGA TAAATAGACTGTAGATAGAGATTATTACCCCCGAGGATCAAGAAATTCTCTATTGACACA ACACCCGATGCTATCAATACACCGATAATATATAGGGGGCATTTTACAGATTCAGGAAGC TGATCCCTGAAGAAAAGCCCGTAGACCCTAATAATTCCTAATATTATCGACGGAGCCATT TCTATGGTACCAGATGCGACCGC → ZZ6_1532 | F primer BamHI GGTGGAAATGGATCCGTCGCTTCAACCTTT |
| R primer EcoRI CGCATCTGGTACCATGAATTCGGCTCCGTC | ||
| Type II NADH: ubiquionone oxidoreductase ndh ZZ6_0213 | ZZ6_0214 ← TGATCGGTTTGTTTTTCCATAACCCATATTGACGGGAAAAGAAGCCAAAGGTCAATCAGA GAAAGCGCGCTTTAAACAAAGCCCATGAGGATGATCAAAGACAGAGACCGGATAAGGGCA AAGATAGTTTCACAGTAAGCAATAAAAACTTTCACTCTGACTTATAATTTATGTGAAACA GATTTGAATGTTCTTCGTTAGTTACAGCAAAAGGTAATTGAGGCGGCTCGCAAACCACCA CGCCAAAGGGGGCTGGGTCGCTTGGAAGAGAATAGAGGTTTCAAATGTCGAAGAATGGTA → ZZ6_0213 | F primer BamHI GGTTTGTGGATCCATAACCCATATTGACGG |
| R primer EcoRI GACATGAATTCCCTCTATTCTCTTCCAAGC | ||
| EGFP gene in plasmid pQE-EGFP egfp | → egfp ATACGGATCCACCGGTCGCCACCATGGTGAGCAAGGGCGAGGAGCTGTTCACCGGGGTGGTGCCCATCCTGGTCGAGCTGGACGGCGACGTAAACGGCCACAAGTTCAGCGTGTCCGGCGAGGGCGAGGGCGATGCCACCTACGGCAAGCTGACCCTGAAGTTCATCTGCACCACCGGCAAGCTGCCCGTGCCCTGGCCCACCCTCGTGACCACCCTGACCTACGGCGTGCAGTGCTTCAGCCGCTACCCCGACCACATGAAGCAGCACGACTTCTTCAAGTCCGCCATGCCCGAAGGCTACGTCCAGGAGCGCACCATCTTCTTCAAGGACGACGGCAACTACAAGACCCGCGCCGAGGTGAAGTTCGAGGGCGACACCCTGGTGAACCGCATCGAGCTGAAGGGCATCGACTTCAAGGAGGACGGCAACATCCTGGGGCACAAGCTGGAGTACAACTACAACAGCCACAACGTCTATATCATGGCCGACAAGCAGAAGAACGGCATCAAGGTGAACTTCAAGATCCGCCACAACATCGAGGACGGCAGCGTGCAGCTCGCCGACCACTACCAGCAGAACACCCCCATCGGCGACGGCCCCGTGCTGCTGCCCGACAACCACTACCTGAGCACCCAGTCCGCCCTGAGCAAAGACCCCAACGAGAAGCGCGATCACATGGTCCTGCTGGAGTTCGTGACCGCCGCCGGGATCACTCTCGGCATGGACGAGCTGTACAAGTAAGGAGGTTCCCGCGGA | F primer EcoRI ATACGGATCCACCGGTCGAATTCATGGTGA |
| R primer HindIII TCCGCGGGAAAGCTTTTACTTGTACAGCTC |
Appendix B
Table A2.
The FTIR spectroscopy data on the relative macromolecular composition (% of dry weight) of the wild type strain Zm6 biomass before and after the 6h aerobic incubation period. Mean values of 4 biological replicates are presented.
Table A2.
The FTIR spectroscopy data on the relative macromolecular composition (% of dry weight) of the wild type strain Zm6 biomass before and after the 6h aerobic incubation period. Mean values of 4 biological replicates are presented.
| Incubation without Substrate Addition | Carbohydrates | Nucleic Acids | Proteins | Lipids | ||||
|---|---|---|---|---|---|---|---|---|
| 0 h | 6 h | 0 h | 6 h | 0 h | 6 h | 0 h | 6 h | |
| Mean value | 12.65 | 11.97 | 17.57 | 18.45 | 57.74 | 57.71 | 2.05 | 1.87 |
| ±SEM | ±0.47 | ±0.39 | ±0.35 | ±0.61 | ±0.61 | ±0.27 | ±0.24 | ±0.17 |
| p value | 0.33 | 0.28 | 0.97 | 0.55 | ||||
| Incubation with 1 g L−1 glucose additions, once per hour | ||||||||
| Mean value | 12.70 | 12.21 | 15.93 | 15.48 | 59.13 | 60.06 | 2.24 | 2.25 |
| ±SEM | ±0.87 | ±0.39 | ±0.19 | ±0.15 | ±0.68 | ±0.49 | ±0.36 | ±0.10 |
| p value | 0.63 | 0.11 | 0.31 | 0.98 | ||||
References
- Rogers, P.L.; Lee, K.J.; Skotnicki, M.L.; Tribe, D.E. Ethanol production by Zymomonas mobilis. In Microbial Reactions; Advances in Biochemical Engineering; Springer: Berlin/Heidelberg, Germany, 1982; Volume 23, pp. 37–84. [Google Scholar]
- Sprenger, G. Carbohydrate metabolism in Zymomonas mobilis: A catabolic highway with some scenic routes. FEMS Microbiol. Lett. 1996, 145, 301–307. [Google Scholar] [CrossRef]
- Swings, J.; De Ley, J. The biology of Zymomonas. Bacteriol. Rev. 1977, 41, 1–46. [Google Scholar] [CrossRef]
- Zhang, M.; Eddy, C.; Deanda, K.; Finkelstein, M.; Picataggio, S. Metabolic engineering of a pentose metabolism pathway in ethanologenic Zymomonas mobilis. Science 1995, 267, 240–243. [Google Scholar]
- Mohagheghi, A.; Linger, J.G.; Yang, S.H.; Smith, H.; Dowe, N.; Zhang, M.; Pienkos, P.T. Improving a recombinant Zymomonas mobilis strain 8b through continuous adaptation on dilute acid pretreated corn stover hydrolysate. Biotechnol. Biofuels 2015, 8, 55. [Google Scholar]
- Yang, S.; Mohagheghi, A.; Franden, M.A.; Chou, Y.C.; Chen, X.; Dowe, N.; Himmel, M.E.; Zhang, M. Metabolic engineering of Zymomonas mobilis for 2,3-butanediol production from lignocellulosic biomass sugars. Biotechnol. Biofuels 2016, 9, 189. [Google Scholar]
- Qiu, M.; Shen, W.; Yan, X.; He, Q.; Cai, D.; Chen, S.; Wei, H.; Knoshaug, E.P.; Zhang, M.; Himmel, M.E.; et al. Metabolic engineering of Zymomonas mobilis for anaerobic isobutanol production. Biotechnol. Biofuels 2020, 13, 15. [Google Scholar]
- Kalnenieks, U.; Pappas, K.M.; Bettenbrock, K. Zymomonas mobilis metabolism: Novel tools and targets for its rational engineering. Adv. Micr. Physiol. 2020, 77, 37–88. [Google Scholar]
- Brooijmans, R.J.W.; Poolman, B.; Schuurman-Wolters, G.K.; De Vos, W.M.; Hugenholtz, J. Generation of a membrane potential by Lactococcus lactis through aerobic electron transport. J. Bacteriol. 2007, 189, 5203–5209. [Google Scholar] [CrossRef]
- Kalnenieks, U.; Balodite, E.; Rutkis, R. Metabolic engineering of bacterial respiration: High vs. low P/O and the case of Zymomonas mobilis. Front. Bioeng. Biotechnol. 2019, 7, 327. [Google Scholar]
- Zelle, E.N.; Oldiges, M.; Koch-Koerfges, A.; Bott, M.; Nöh, K.; Wiechert, W. An energetic profile of Corynebacterium glutamicum underpinned by measured biomass yield on ATP. Metabol. Eng. 2021, 65, 66–78. [Google Scholar] [CrossRef]
- Zotta, T.; Guidone, A.; Ianniello, R.G.; Parente, E.; Ricciardi, A. Temperature and respiration affect the growth and stress resistance of Lactobacillus plantarum C17. J. Appl. Microbiol. 2013, 115, 848–858. [Google Scholar] [CrossRef]
- Hayashi, T.; Kato, T.; Watakabe, S.; Song, W.; Aikawa, S.; Furukawa, K. The respiratory chain provides salt stress tolerance by maintaining a low NADH/NAD+ ratio in Zymomonas mobilis. Microbiology 2015, 161, 2384–2394. [Google Scholar]
- Murata, M.; Ishii, A.; Fujimoto, H.; Nishimura, K.; Kosaka, T.; Mori, H.; Yamada, M. Update of thermotolerant genes essential for survival at a critical high temperature in Escherichia coli. PLoS ONE 2018, 13, e0189487. [Google Scholar] [CrossRef]
- Strohdeicher, M.; Neuß, B.; Bringer-Meyer, S.; Sahm, H. Electron transport chain of Zymomonas mobilis. Interaction with the membrane-bound glucose dehydrogenase and identification of ubiquinone 10. Arch. Microbiol. 1990, 154, 536–543. [Google Scholar]
- Kalnenieks, U.; Galinina, N.; Bringer-Meyer, S.; Poole, R.K. Membrane d-lactate oxidase in Zymomonas mobilis: Evidence for a branched respiratory chain. FEMS Microbiol. Lett. 1998, 168, 91–97. [Google Scholar]
- Kalnenieks, U.; Galinina, N.; Strazdina, I.; Kravale, Z.; Pickford, J.L.; Rutkis, R.; Poole, R.K. NADH dehydrogenase deficiency results in low respiration rate and improved aerobic growth of Zymomonas mobilis. Microbiology 2008, 154, 989–994. [Google Scholar] [CrossRef]
- Sootsuwan, K.; Lertwattanasakul, N.; Thanonkeo, P.; Matsushita, K.; Yamada, M. Analysis of the respiratory chain in ethanologenic Zymomonas mobilis with a cyanide-resistant bd-type ubiquinol oxidase as the only terminal oxidase and its possible physiological roles. J. Mol. Microbiol. Biotechnol. 2008, 14, 163–175. [Google Scholar] [CrossRef]
- Seo, J.-S.; Chong, H.; Park, H.S.; Yoon, K.-O.; Jung, C.; Hong, J.H.; Kim, H.; Kim, J.-H.; Kil, J.-I.; Park, C.J.; et al. The genome sequence of the ethanologenic bacterium Zymomonas mobilis ZM4. Nat. Biotechnol. 2005, 23, 63–68. [Google Scholar] [CrossRef]
- Yang, S.; Pappas, K.M.; Hauser, L.J.; Land, M.L.; Chen, G.L.; Hurst, G.B.; Pan, C.; Kouvelis, V.N.; A Typas, M.; A Pelletier, D.; et al. Improved genome annotation for Zymomonas mobilis. Nat. Biotechnol. 2009, 27, 893–894. [Google Scholar] [CrossRef]
- Balodite, E.; Strazdina, I.; Galinina, N.; McLean, S.; Rutkis, R.; Poole, R.K.; Kalnenieks, U. Structure of the Zymomonas mobilis respiratory chain: Oxygen affinity of electron transport and the role of cytochrome c peroxidase. Microbiology 2014, 160, 2045–2052. [Google Scholar] [CrossRef]
- Rutkis, R.; Galinina, N.; Strazdina, I.; Kalnenieks, U. The inefficient aerobic energetics of Zymomonas mobilis: Identifying the bottleneck. J. Basic Microbiol. 2014, 54, 1090–1097. [Google Scholar] [CrossRef]
- Rutkis, R.; Strazdina, I.; Balodite, E.; Lasa, Z.; Galinina, N.; Kalnenieks, U. The low energy-coupling respiration in Zymomonas mobilis accelerates flux in the Entner–Doudoroff pathway. PLoS ONE 2016, 11, e0153866. [Google Scholar]
- Kalnenieks, U.; de Graaf, A.A.; Bringer-Meyer, S.; Sahm, H. Oxidative phosphorylation in Zymomonas mobilis. Arch. Microbiol. 1993, 160, 74–79. [Google Scholar] [CrossRef]
- Belaich, J.P.; Senez, J.C. Influence of aeration and pantothenate on growth yields of Zymomonas mobilis. J. Bacteriol. 1965, 89, 1195–1200. [Google Scholar]
- Kalnenieks, U. Physiology of Zymomonas mobilis: Some unanswered questions. Adv. Microb. Physiol. 2006, 51, 73–117. [Google Scholar] [CrossRef]
- Hayashi, T.; Furuta, Y.; Furukawa, K. Respiration-deficient mutants of Zymomonas mobilis show improved growth and ethanol fermentation under aerobic and high temperature conditions. J. Biosci. Bioeng. 2011, 111, 414–419. [Google Scholar]
- Hayashi, T.; Kato, T.; Furukawa, K. Respiratory chain analysis of Zymomonas mobilis mutants producing high levels of ethanol. Appl. Environ. Microbiol. 2012, 78, 5622–5629. [Google Scholar] [CrossRef]
- Kalnenieks, U.; Balodite, E.; Strähler, S.; Strazdina, I.; Rex, J.; Pentjuss, A.; Fuchino, K.; Bruheim, P.; Rutkis, R.; Pappas, K.M.; et al. Improvement of acetaldehyde production in Zymomonas mobilis by engineering of its aerobic metabolism. Front. Microbiol. 2019, 10, 2533. [Google Scholar] [CrossRef]
- De Deken, R.H. The Crabtree effect: A regulatory system in yeast. J. Gen. Microbiol. 1966, 44, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Dashko, S.; Zhou, N.; Compagno, C.; Piskur, J. Why, when, and how did yeast evolve alcoholic fermentation? FEMS Yeast Res. 2014, 14, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Swamy, K.B.S.; Leu, J.-Y.; McDonald, M.J.; Galafassi, S.; Compagno, C.; Piškur, J. Coevolution with bacteria drives the evolution of aerobic fermentation in Lachancea kluyveri. PLoS ONE 2017, 12, e0173318. [Google Scholar] [CrossRef]
- He, M.X.; Wu, B.; Shui, Z.X.; Hu, Q.C.; Wang, W.G.; Tan, F.R.; Tang, X.-Y.; Zhu, Q.-L.; Pan, K.; Li, Q.; et al. Transcriptome profiling of Zymomonas mobilis under ethanol stress. Biotechnol. Biofuels 2012, 5, 75. [Google Scholar] [CrossRef]
- Yang, S.; Pan, C.; Tschaplinski, T.J.; Hurst, G.B.; Engle, N.L.; Zhou, W.; Dam, P.; Xu, Y.; Rodriguez, M.; Dice, L.; et al. Systems Biology Analysis of Zymomonas mobilis ZM4 Ethanol Stress Responses. PLoS ONE 2013, 7, e68886. [Google Scholar] [CrossRef]
- Strazdina, I.; Balodite, E.; Lasa, Z.; Rutkis, R.; Galinina, N.; Kalnenieks, U. Aerobic catabolism and respiratory lactate bypass in Ndh-negative Zymomonas mobilis. Metabol. Eng. Commun. 2018, 7, e00081. [Google Scholar]
- Zou, S.; Zhang, K.; You, L.; Zhao, X.; Jing, X.; Zhang, M. Enhanced electrotransformation of the ethanologen Zymomonas mobilis ZM4 with plasmids. Eng. Life Sci. 2012, 12, 152–161. [Google Scholar]
- Grube, M.; Bekers, M.; Upite, D.; Kaminska, E. IR-spectroscopic studies of Zymomonas mobilis and levan precipitate. Vibr. Spectrosc. 2002, 28, 277–285. [Google Scholar] [CrossRef]
- DeLey, J.; Swings, J. Phenotypic description, numerical analysis, and proposal for an improved taxonomy and nomeclature of the genus Zymomonas Kluyver and van Niel 1936. Int. J. Syst. Bacteriol. 1976, 26, 146–157. [Google Scholar]
- Dawes, E.A.; Large, P.J. Effect of starvation on the viability and cellular constituents of Zymomonas anaerobia and Zymomonas mobilis. J. Gen. Microbiol. 1970, 60, 31–62. [Google Scholar]
- Jones-Burrage, S.E.; Kremer, T.A.; McKinlay, J.B. Cell aggregation and aerobic respiration are important for Zymomonas mobilis ZM4 survival in an aerobic minimal medium. Appl. Environ. Microbiol. 2019, 85, e00193-19. [Google Scholar] [CrossRef]
- Hong, Y.; Zeng, J.; Wang, X.; Drlica, K.; Zhao, X. Post-stress bacterial cell death mediated by reactive oxygen species. Proc. Natl. Acad. Sci. USA 2019, 116, 10064–10071. [Google Scholar] [CrossRef]
- Cao, H.; Wei, D.; Yang, Y.; Shang, Y.; Li, G.; Zhou, Y.; Ma, Q.; Xu, Y. Systems-level understanding of ethanol-induced stresses and adaptation in E. coli. Sci. Rep. 2017, 7, 44150. [Google Scholar] [CrossRef]
- Perez-Gallardo, R.V.; Briones, L.S.; Diaz-Perez, A.L.; Gutierrez, S.; Rodriguez-Zavala, J.S.; Campos-Garcia, J. Reactive oxygen species production induced by ethanol in Saccharomyces cerevisiae increases because of a dysfunctional mitochondrial iron–sulfur cluster assembly system. FEMS Yeast Res. 2013, 13, 804–819. [Google Scholar] [CrossRef]
- Martien, J.I.; Hebert, A.S.; Stevenson, D.M.; Regner, M.R.; Khana, D.B.; Coon, J.J.; Amador-Noguez, D. Systems-level analysis of oxygen exposure in Zymomonas mobilis: Implications for isoprenoid production. mSystems 2019, 4, e00284-18. [Google Scholar] [CrossRef]
- Charoensuk, K.; Irie, A.; Lertwattanasakul, N.; Sootsuwan, K.; Thanonkeo, P.; Yamada, M. Physiological importance of cytochrome c peroxidase in ethanologenic thermotolerant Zymomonas mobilis. J. Mol. Microbiol. Biotechnol. 2011, 20, 70–82. [Google Scholar] [CrossRef]
- Messner, K.R.; Imlay, J.A. The Identification of Primary Sites of Superoxide and Hydrogen Peroxide Formation in the Aerobic Respiratory Chain and Sulfite Reductase Complex of Escherichia coli. J. Biol. Chem. 1999, 274, 10119–10128. [Google Scholar]
- Jarmuszkiewicz, W.; Dominiak, K.; Budzinska, A.; Wojcicki, K.; Galganski, L. Mitochondrial Coenzyme Q Redox Homeostasis and Reactive Oxygen Species Production. Front. Biosci. 2023, 28, 61. [Google Scholar] [CrossRef]
- Felczak, M.M.; TerAvest, M.A. Respiration is essential for aerobic growth of Zymomonas mobilis ZM4. bioRxiv 2023. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).