
Bacterial Interactions with Nanoplastics and the Environmental Effects They Cause

Abstract
:1. Introduction
2. Effects of Nanoplastics on Bacteria
2.1. Inhibitory Growth
2.2. Biofilm Formation Impact
2.3. Genetic Linkage
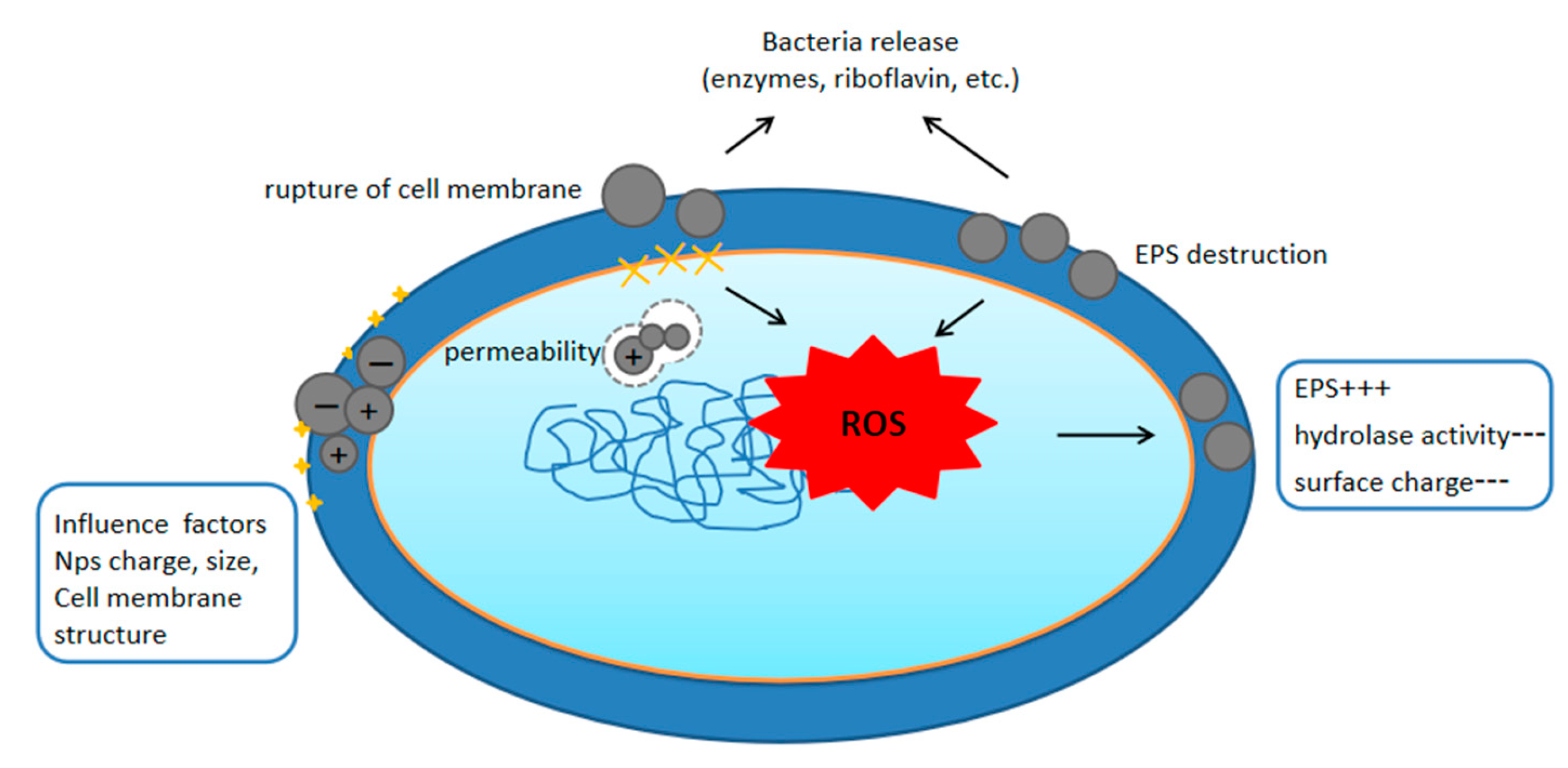
3. Mechanisms by Which Nanoplastics Affect Bacteria
3.1. Nanoplastics and Cell Membrane Interactions

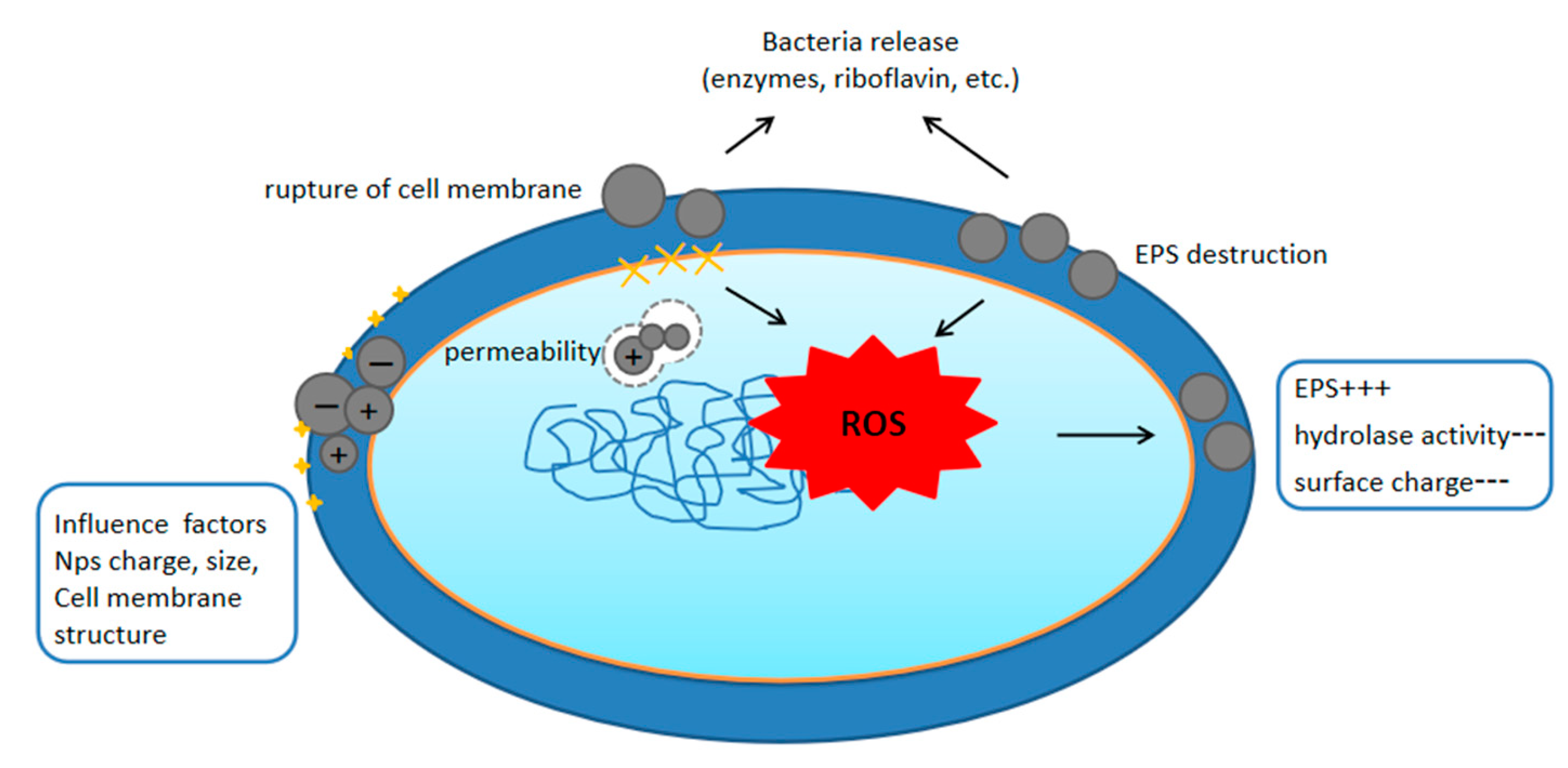{kind=link}
| Types | Size | Concentrations | Time | Mechanisms of Interaction | Ref. |
|---|---|---|---|---|---|
| Staphylococcus aureus, Bacillus subtilis, Escherichia coli | PCLA: 50 nm HDPE: 615 nm PESa: 396 nm PP: 531nm | 10–100 mg/L | 24 h | Decreased lipid peroxidase activity and increased antioxidant ratio lead to the occurrence of oxidative stress | [27] |
| Pseudomonas aeruginosa PAO1 | PS: 120 nm | 0.1, 1, 10, 20, 50 mg/L | 72 h | ROS generation, oxidative stress induction, membrane interaction | [28] |
| Methanosarcina acetivorans | PS-NH2: 30 nm | 5, 8 mg/L | 4 days | ROS generation, oxidative stress induction, membrane interaction | [29] |
| Methanosarcina acetivorans | PS-NH2: 30 nm | 20, 50 mg/L | 4 days | ROS generation, oxidative stress induction, membrane interaction | [29] |
| Alkalophilic halomonas | PS: 50 nm PS-NH2: 55 nm | 80 mg/L | 0.5–2 h | ROS generation, oxidative stress induction, membrane interaction | [11] |
| Lactobacillus plantarum ZP-6 Lactobacillus brevis ZB-1 Lactobacillus fermentum ZF-3 Lactobacillus gasseri ZG-4 | PP: 100 nm PE: 100 nm PVC: 100 nm | 30 mg/L | 2 h, 4 h, 6 h, 8 h | Membrane interactions were observed | [30] |
| Sylvatica | PS: 159nm | 150 mg/L | 4 h–5 day | Membrane interactions were observed | [21] |
| Escherichia coli | PS: 200nm | 4 mg/L | 24 h | Membrane interactions were observed | [23] |
3.2. Nanoplastics Induce Oxidative Stress in Bacteria
4. Factors Affecting the Interaction of Nanoplastics with Bacteria
4.1. Properties of Plastic Pellets
4.2. Properties of the Medium
5. Ecological Impacts of Nanoplastics and Bacteria Working Together
5.1. Nanoplastics Affect the Environment by Influencing Bacteria
5.2. Nanoplastics and Bacteria form Ecological Corona That Affects the Environment
| Types | Size | Concentrations | Time | Ecocorona Composition | Aims | Ref. |
|---|---|---|---|---|---|---|
| Halomonas genus | 50–500 nm | 5 mg/L | 24 h | Glycoprotein polymers | The generation of NPs-EPS ecological corona affects the sinking rate. The EPS composition may affect the buoyant density of NPs’ agglomerates. | [36] |
| Chlorella vulgaris | 200 nm | 1 mg/L | 72 h | 20 kDa to >100 kDa in size | EPS further enhanced the positive effect of PS MPs on bacterial growth. | [42] |
| Marine diatoms | 60 nm | 1–50 mg/L | 3 h | Carbohydrates and proteins | EPS increased the colloidal stability of PS M/NPs in NaCl or CaCl2. | [44] |
| Escherichia coli | 20–50 nm | 200 mg/L | 40 min | Proteins dominate over polysaccharides in the corona | EPS corona formation readily adsorbs metal ions, forming NPs-EPS-metal complexes and promoting sedimentation. | [61] |
| Escherichia coli | 500nm | 10 mg/L | 40 min | Approximately 70% of the adsorbed HA macromolecules were over 30 kDa | EPS corona enhances the transport of nanoplastics. | [62] |
| Bacillus subtilis | <1000 µm | 10 mg/L | 40 min | Glycoprotein polymers | The NPs-EPS corona increased the toxicity of NPs to the environment by accumulating highly toxic heavy metals. | [63] |
6. Conclusions
- (1)
- In order to mimic and illustrate the toxicity of nanoplastics in true natural habitats, it is advised that future research concentrate more on the toxicity of nanoplastics at realistic concentrations in natural environments;
- (2)
- Nanoplastics are materials with extremely small particles that are chemically similar to organic matter. Future research should be done to fill the gap in this field. There is only one experimental way to study the interactions between nanoplastics and bacteria and it is difficult to directly detect the distribution and abundance of nanoplastics in the environment due to shortcomings in detection methods and tools;
- (3)
- To gather accurate information on the toxicity of NPs to microorganisms, it is important to examine the kinetics and absorption pathways of various MP types (polymer type, size, and shape) when interacting with various bacterial species.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, Y.; Zhou, D.; Li, Z.; Zhu, C. Advancement and Challenges of Microplastic Pollution in the Aquatic Environment: A Review. Water Air Soil Pollut. 2018, 229, 140. [Google Scholar] [CrossRef]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic waste inputs from land into the ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Koelmans, A.A.; Besseling, E.; Foekema, E.; Kooi, M.; Mintenig, S.; Ossendorp, B.C.; Redondo-Hasselerharm, P.E.; Verschoor, A.; van Wezel, A.P.; Scheffer, M. Risks of Plastic Debris: Unravelling Fact, Opinion, Perception, and Belief. Environ. Sci. Technol. 2017, 51, 11513–11519. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wei, W.; Liu, X.; Ni, B. Emerging electrochemical techniques for identifying and removing micro/nanoplastics in urban waters. Water Res. 2022, 221, 118846. [Google Scholar] [CrossRef] [PubMed]
- Materić, D.; Holzinger, R.; Niemann, H. Nanoplastics and ultrafine microplastic in the Dutch Wadden Sea—The hidden plastics debris? Sci. Total Environ. 2022, 846, 157371. [Google Scholar] [CrossRef] [PubMed]
- Hüffer, T.; Weniger, A.; Hofmann, T. Sorption of organic compounds by aged polystyrene microplastic particles. Environ. Pollut. 2018, 236, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Gao, P.; Li, H.; Huang, J.; Zhang, Y.; Ding, H.; Zhang, W. Mechanism of the inhibition and detoxification effects of the interaction between nanoplastics and microalgae Chlorella pyrenoidosa. Sci. Total Environ. 2021, 783, 146919. [Google Scholar] [CrossRef]
- Qiu, Y.; Zhou, S.; Zhang, C.; Zhou, Y.; Qin, W. Soil microplastic characteristics and the effects on soil properties and biota: A systematic review and meta-analysis. Environ. Pollut. 2022, 313, 120183. [Google Scholar] [CrossRef]
- Ruvini, L.; Dabare, P.; Bachhuka, A.; Palms, D.; Parkinson-Lawrence, E.; Hayball, J.D.; Mierczynska, A.; Vasilev, K. Surface chemistry mediated albumin adsorption, conformational changes and influence on innate immune responses. Appl. Surf. Sci. 2022, 596, 153518. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, Y.J.; Lee, S.; Lee, E. Interactions between bacteria and nano (micro)-sized polystyrene particles by bacterial responses and microscopy. Chemosphere 2022, 306, 135584. [Google Scholar] [CrossRef]
- Sun, X.; Chen, B.; Li, Q.; Liu, N.; Xia, B.; Zhu, L.; Qu, K. Toxicities of polystyrene nano- and microplastics toward marine bacterium Halomonas alkaliphila. Sci. Total Environ. 2018, 642, 1378–1385. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wu, R.; Yang, X.; Fang, S.; Xiang, Z.; Yang, S.; Ouyang, G. Water-dispersible nano-pollutions reshape microbial metabolism in type-specific manners: A metabolic and bacteriological investigation in Escherichia coli. Front. Environ. Sci. Eng. 2022, 16, 116. [Google Scholar] [CrossRef]
- Fu, S.; Ding, J.; Zhang, Y.; Li, Y.; Zhu, R.; Yuan, X.; Zou, H. Exposure to polystyrene nanoplastic leads to inhibition of anaerobic digestion system. Sci. Total Environ. 2018, 625, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Okshevsky, M.; Gautier, E.; Farner, J.M.; Schreiber, L.; Tufenkji, N. Biofilm formation by marine bacteria is impacted by concentration and surface functionalization of polystyrene nanoparticles in a species-specific manner. Environ. Microbiol. Rep. 2020, 12, 203–213. [Google Scholar] [CrossRef]
- Arias-Andres, M.; Klümper, U.; Rojas-Jimenez, K.; Grossart, H. Microplastic pollution increases gene exchange in aquatic ecosystems. Environ. Pollut. 2018, 237, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xu, R.; Wu, H.; Xu, K.; Zhang, W.; Wang, Z.; Gong, W. Nanoplastics promote the dissemination of antibiotic resistance through conjugative gene transfer: Implications from oxidative stress and gene expression. Environ. Sci. Nano 2023, 10, 1329–1340. [Google Scholar] [CrossRef]
- Ning, Q.; Wang, D.; An, J.; Ding, Q.; Huang, Z.; Zou, Y.; Wu, F.; You, J. Combined effects of nanosized polystyrene and erythromycin on bacterial growth and resistance mutations in Escherichia coli. J. Hazard. Mater. 2022, 422, 126858. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ma, D.; Feng, K.; Lou, Y.; Zhou, H.; Liu, B.; Xie, G.; Ren, N.; Xing, D. Polystyrene nanoplastics shape microbiome and functional metabolism in anaerobic digestion. Water Res. 2022, 219, 118606. [Google Scholar] [CrossRef]
- Septiadi, D.; Crippa, F.; Moore, T.L.; Rothen-Rutishauser, B.; Petri-Fink, A. Nanoparticle–Cell Interaction: A Cell Mechanics Perspective. Adv. Mater. 2018, 30, 1704463. [Google Scholar] [CrossRef]
- Liang, Z.; Das, A.; Hu, Z. Bacterial response to a shock load of nanosilver in an activated sludge treatment system. Water Res. 2010, 44, 5432–5438. [Google Scholar] [CrossRef]
- Fringer, V.S.; Fawcett, L.P.; Mitrano, D.M.; Maurer-Jones, M.A. Impacts of Nanoplastics on the Viability and Riboflavin Secretion in the Model Bacteria Shewanella oneidensis. Front. Environ. Sci. 2020, 8, 97. [Google Scholar] [CrossRef]
- Rossi, G.; Barnoud, J.; Monticelli, L. Polystyrene Nanoparticles Perturb Lipid Membranes. J. Phys. Chem. Lett. 2014, 5, 241–246. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wu, D.; Rong, H.; Li, M.; Tong, M.; Kim, H. Influence of Nano- and Microplastic Particles on the Transport and Deposition Behaviors of Bacteria in Quartz Sand. Environ. Sci. Technol. 2018, 52, 11555–11563. [Google Scholar] [CrossRef] [PubMed]
- Zha, Y.; Li, Z.; Zhong, Z.; Ruan, Y.; Sun, L.; Zuo, F.; Li, L.; Hou, S. Size-dependent enhancement on conjugative transfer of antibiotic resistance genes by micro/nanoplastics. J. Hazard. Mater. 2022, 431, 128561. [Google Scholar] [CrossRef] [PubMed]
- Perini, D.A.; Parra-Ortiz, E.; Varó, I.; Queralt-Martín, M.; Malmsten, M.; Alcaraz, A. Surface-Functionalized Polystyrene Nanoparticles Alter the Transmembrane Potential via Ion-Selective Pores Maintaining Global Bilayer Integrity. Langmuir 2022, 38, 14837–14849. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Ye, R.; Huang, J.; Wang, B.; Xie, Z.; Ou, X.; Yu, N.; Huang, C.; Hua, Y.; Zhou, R.; et al. Distinct lipid membrane interaction and uptake of differentially charged nanoplastics in bacteria. J. Nanobiotechnol. 2022, 20, 191. [Google Scholar] [CrossRef] [PubMed]
- Saygin, H.; Baysal, A. Similarities and Discrepancies Between Bio-Based and Conventional Submicron-Sized Plastics: In Relation to Clinically Important Bacteria. Bull. Environ. Contam. Toxicol. 2020, 105, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Kang, Y.; Huang, F.; Su, Y.; Zhou, X.; Wang, A.; Gao, S. Distinct responses of Pseudomonas aeruginosa PAO1 exposed to different levels of polystyrene nanoplastics. Sci. Total Environ. 2022, 852, 158214. [Google Scholar] [CrossRef]
- Liu, X.; Ma, J.; Duan, J.; Sun, X.; Feng, L.; Li, X.; Han, Y.; Zhang, K.; Zhang, M.; Wang, Y.; et al. The surface groups of polystyrene nanoparticles control their interaction with the methanogenic archaeon Methanosarcina acetivorans. Water Res. 2022, 223, 118993. [Google Scholar] [CrossRef]
- Zhao, L.; Dou, Q.; Chen, S.; Wang, Y.; Yang, Q.; Chen, W.; Zhang, H.; Du, Y.; Xie, M. Adsorption abilities and mechanisms of Lactobacillus on various nanoplastics. Chemosphere 2023, 320, 138038. [Google Scholar] [CrossRef]
- Zheng, T.; Yuan, D.; Liu, C. Molecular toxicity of nanoplastics involving in oxidative stress and desoxyribonucleic acid damage. J. Mol. Recognit. 2019, 32, e2804. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yuan, D.; Shan, M.; Yang, Z.; Liu, C. Single and combined effects of amino polystyrene and perfluorooctane sulfonate on hydrogen-producing thermophilic bacteria and the interaction mechanisms. Sci. Total Environ. 2020, 703, 135015. [Google Scholar] [CrossRef]
- Chen, Z.; Shi, X.; Zhang, J.; Wu, L.; Wei, W.; Ni, B. Nanoplastics are significantly different from microplastics in urban waters. Water Res. X 2023, 19, 100169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W. Nanoparticle Aggregation: Principles and Modeling; Springer: Dordrecht, The Netherlands, 2014; pp. 19–43. ISBN 978-94-017-8739-0. [Google Scholar]
- He, Y.T.; Wan, J.; Tokunaga, T. Kinetic stability of hematite nanoparticles: The effect of particle sizes. J. Nanopart. Res. 2008, 10, 321–332. [Google Scholar] [CrossRef]
- Summers, S.; Henry, T.; Gutierrez, T. Agglomeration of nano- and microplastic particles in seawater by autochthonous and de novo-produced sources of exopolymeric substances. Mar. Pollut. Bull. 2018, 130, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Yang, X.; Chen, F.; Zhao, F.; Zhao, Y.; Ruan, L.; Wang, Y.; Yang, Y. Fate and transport of nanoplastics in complex natural aquifer media: Effect of particle size and surface functionalization. Sci. Total Environ. 2019, 669, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Zhou, D.; Yang, S.; Shi, Z.; Pan, H.; Jin, Q.; Ding, Z. Neurotoxicity of polystyrene nanoplastics with different particle sizes at environment-related concentrations on early zebrafish embryos. Sci. Total Environ. 2023, 872, 162096. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Ma, X.; Guo, B.; Zhang, K. Environmental factors-mediated behavior of microplastics and nanoplastics in water: A review. Chemosphere 2021, 271, 129597. [Google Scholar] [CrossRef]
- Fan, X.; Gan, R.; Liu, J.; Xie, Y.; Xu, D.; Xiang, Y.; Su, J.; Teng, Z.; Hou, J. Adsorption and desorption behaviors of antibiotics by tire wear particles and polyethylene microplastics with or without aging processes. Sci. Total Environ. 2021, 771, 145451. [Google Scholar] [CrossRef]
- Oliveira, M.M.; Proenca, A.M.; Moreira-Silva, E.; de Castro, A.M.; Dos Santos, F.M.; Marconatto, L.; Medina-Silva, R. Biofilms of Pseudomonas and Lysinibacillus Marine Strains on High-Density Polyethylene. Microb. Ecol. 2021, 81, 833–846. [Google Scholar] [CrossRef]
- Natarajan, L.; Omer, S.; Jetly, N.; Jenifer, M.A.; Chandrasekaran, N.; Suraishkumar, G.K.; Mukherjee, A. Eco-corona formation lessens the toxic effects of polystyrene nanoplastics towards marine microalgae Chlorella sp. Environ. Res. 2020, 188, 109842. [Google Scholar] [CrossRef] [PubMed]
- Canesi, L.; Balbi, T.; Fabbri, R.; Salis, A.; Damonte, G.; Volland, M.; Blasco, J. Biomolecular coronas in invertebrate species: Implications in the environmental impact of nanoparticles. Nanoimpact 2017, 8, 89–98. [Google Scholar] [CrossRef]
- Grassi, G.; Gabellieri, E.; Cioni, P.; Paccagnini, E.; Faleri, C.; Lupetti, P.; Corsi, I.; Morelli, E. Interplay between extracellular polymeric substances (EPS) from a marine diatom and model nanoplastic through eco-corona formation. Sci. Total Environ. 2020, 725, 138457. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, C.; Tallec, K.; Le Goïc, N.; Lambert, C.; Soudant, P.; Huvet, A.; Suquet, M.; Berchel, M.; Paul-Pont, I. Cellular responses of Pacific oyster (Crassostrea gigas) gametes exposed in vitro to polystyrene nanoparticles. Chemosphere 2018, 208, 764–772. [Google Scholar] [CrossRef]
- Guo, J.; Huang, X.; Xiang, L.; Wang, Y.; Li, Y.; Li, H.; Cai, Q.; Mo, C.; Wong, M. Source, migration and toxicology of microplastics in soil. Environ. Int. 2020, 137, 105263. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Jiang, R.; Lin, W.; Ouyang, G. Effect of salinity and humic acid on the aggregation and toxicity of polystyrene nanoplastics with different functional groups and charges. Environ. Pollut. 2019, 245, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Bolan, N.; Tsang, D.C.W.; Sarkar, B.; Bradney, L.; Li, Y. A review of microplastics aggregation in aquatic environment: Influence factors, analytical methods, and environmental implications. J. Hazard. Mater. 2021, 402, 123496. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Hu, L.; Shi, H.; Ye, J.; Zhang, Y.; Kim, H. Effects of inorganic ions and natural organic matter on the aggregation of nanoplastics. Chemosphere 2018, 197, 142–151. [Google Scholar] [CrossRef]
- Shams, M.; Alam, I.; Chowdhury, I. Aggregation and stability of nanoscale plastics in aquatic environment. Water Res. 2020, 171, 115401. [Google Scholar] [CrossRef]
- Li, Z.; Yi, X.; Zhou, H.; Chi, T.; Li, W.; Yang, K. Combined effect of polystyrene microplastics and dibutyl phthalate on the microalgae Chlorella pyrenoidosa. Environ. Pollut. 2020, 257, 113604. [Google Scholar] [CrossRef]
- Romero-Cano, M.S.; Martín-Rodríguez, A.; de Las Nieves, F.J. Electrosteric Stabilization of Polymer Colloids with Different Functionality. Langmuir 2001, 17, 3505–3511. [Google Scholar] [CrossRef]
- Mccormick, A.; Hoellein, T.J.; Mason, S.A.; Schluep, J.; Kelly, J.J. Microplastic is an Abundant and Distinct Microbial Habitat in an Urban River. Environ. Sci. Technol. 2014, 48, 11863–11871. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.K.; Hong, S.H.; Jang, M.; Han, G.M.; Rani, M.; Lee, J.; Shim, W.J. A comparison of microscopic and spectroscopic identification methods for analysis of microplastics in environmental samples. Mar. Pollut. Bull. 2015, 93, 202–209. [Google Scholar] [CrossRef]
- Kesy, K.; Oberbeckmann, S.; Müller, F.; Labrenz, M. Polystyrene influences bacterial assemblages in Arenicola marina-populated aquatic environments in vitro. Environ. Pollut. 2016, 219, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Zheng, Y. Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef] [PubMed]
- Awet, T.T.; Kohl, Y.; Meier, F.; Straskraba, S.; Grün, A.L.; Ruf, T.; Jost, C.; Drexel, R.; Tunc, E.; Emmerling, C. Effects of polystyrene nanoparticles on the microbiota and functional diversity of enzymes in soil. Environ. Sci. Eur. 2018, 30, 11. [Google Scholar] [CrossRef]
- Huang, D.; Xu, Y.; Lei, F.; Yu, X.; Ouyang, Z.; Chen, Y.; Jia, H.; Guo, X. Degradation of polyethylene plastic in soil and effects on microbial community composition. J. Hazard. Mater. 2021, 416, 126173. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Peng, H.; Xiao, L. The influence of microplastics for the transportation of E. coli using column model. Sci. Total Environ. 2021, 786, 147487. [Google Scholar] [CrossRef]
- Ni, B.; Fang, F.; Xie, W.; Sun, M.; Sheng, G.; Li, W.; Yu, H. Characterization of extracellular polymeric substances produced by mixed microorganisms in activated sludge with gel-permeating chromatography, excitation–emission matrix fluorescence spectroscopy measurement and kinetic modeling. Water Res. 2009, 43, 1350–1358. [Google Scholar] [CrossRef]
- Zhang, P.; Xu, X.; Zhang, X.; Zou, K.; Liu, B.; Qing, T.; Feng, B. Nanoparticles-EPS corona increases the accumulation of heavy metals and biotoxicity of nanoparticles. J. Hazard. Mater. 2021, 409, 124526. [Google Scholar] [CrossRef]
- Zhu, M.; Zhang, Z.; Zhang, T.; Hofmann, T.; Chen, W. Eco-Corona Dictates Mobility of Nanoplastics in Saturated Porous Media: The Critical Role of Preferential Binding of Macromolecules. Environ. Sci. Technol. 2023, 57, 331–339. [Google Scholar] [CrossRef]
- Xiong, S.; Cao, X.; Eggleston, I.; Chi, Y.; Li, A.; Liu, X.; Zhao, J.; Xing, B. Role of extracellular polymeric substances in the aggregation and biological response of micro(nano)plastics with different functional groups and sizes. J. Hazard. Mater. 2023, 446, 130713. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Li, X.; Li, J.; Dai, W.; Luan, Y. Bacterial Interactions with Nanoplastics and the Environmental Effects They Cause. Fermentation 2023, 9, 939. https://doi.org/10.3390/fermentation9110939
Wang R, Li X, Li J, Dai W, Luan Y. Bacterial Interactions with Nanoplastics and the Environmental Effects They Cause. Fermentation. 2023; 9(11):939. https://doi.org/10.3390/fermentation9110939
Chicago/Turabian StyleWang, Rongyu, Xiaodong Li, Jing Li, Wei Dai, and Yaning Luan. 2023. "Bacterial Interactions with Nanoplastics and the Environmental Effects They Cause" Fermentation 9, no. 11: 939. https://doi.org/10.3390/fermentation9110939
APA StyleWang, R., Li, X., Li, J., Dai, W., & Luan, Y. (2023). Bacterial Interactions with Nanoplastics and the Environmental Effects They Cause. Fermentation, 9(11), 939. https://doi.org/10.3390/fermentation9110939