
Assessing Hydrolyzed Gluten Content in Dietary Enzyme Supplements Following Fermentation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Samples for Multiplex and Immunoblotting
2.3. Hydrolysis of Gluten Using Pepsin and Trypsin Complex
2.4. Multiplex Competitive ELISA
2.5. Immunoblot Using Automated Capillary Electrophoretic-Based Immunoassays
2.6. Preparation of Gluten and Gliadin and PT-Digested Gluten for Cell Culture
2.7. Preparation of Supplemental Enzymes for Cell Culture
2.8. Cell Culture
2.9. Treatment Scheme
2.10. PBMCs Culture Conditions and Treatment
2.11. Cell Viability Assays
2.12. RNA Isolation and Quantitative Real-Time PCR Analysis
2.13. ELISA Cytokine Determination
2.14. Statistical Analysis
3. Results
3.1. Multiplex ELISA and Immunoblotting of Enzyme Fermentation Products
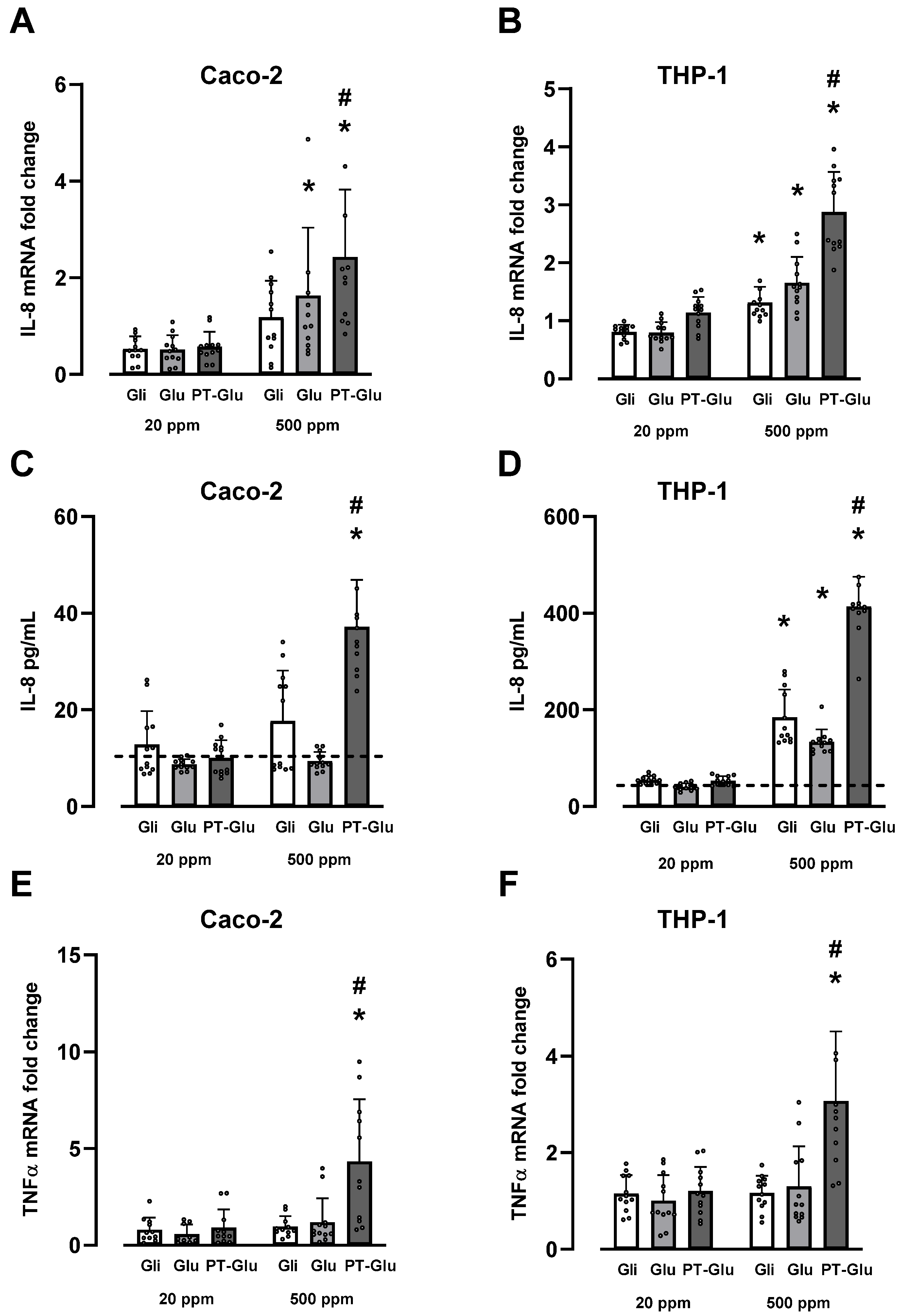
3.2. Effects of Gluten, Gliadin and Digested Gluten Standard on TNF-α and IL-8 Production in THP-1 and Caco-2 Cells
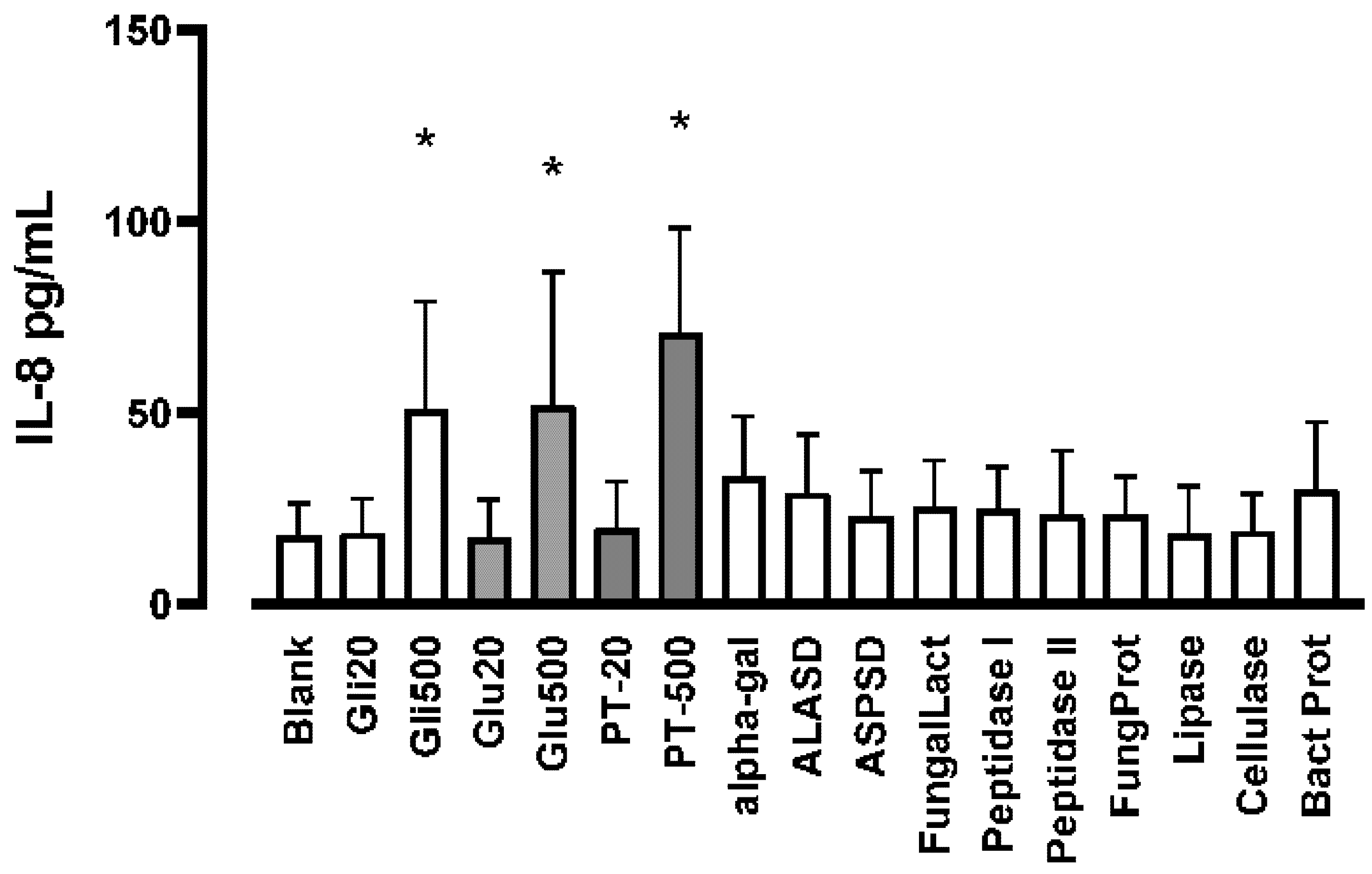
3.3. Effects of Gluten, Gliadin, Digested Gluten Standard and Supplemental Enzymes on IL-8 Production in THP-1 Cells
3.4. Effects of Gluten, Gliadin and Digested Gluten Standard on IL-8 Protein Production in Human Peripheral Blood Mononuclear Cells (PBMCs)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A
References
- Gao, H.; Jorgensen, R.; Raghunath, R.; Nagisetty, S.; Ng, P.K.W.; Gangur, V. Creating hypo-/nonallergenic wheat products using processing methods: Fact or fiction? Compr. Rev. Food Sci. Food Saf. 2021, 20, 6089–6115. [Google Scholar] [CrossRef] [PubMed]
- Vu, V.; Farkas, C.; Riyad, O.; Bujna, E.; Kilin, A.; Sipiczki, G.; Sharma, M.; Usmani, Z.; Gupta, V.K.; Nguyen, Q.D. Enhancement of the enzymatic hydrolysis efficiency of wheat bran using the Bacillus strains and their consortium. Bioresour. Technol. 2021, 343, 126092. [Google Scholar] [CrossRef] [PubMed]
- Dubos, R.J. The Adaptive Production of Enzymes by Bacteria. Bacteriol. Rev. 1940, 4, 1–16. [Google Scholar] [CrossRef]
- Cezairliyan, B.; Ausubel, F.M. Investment in secreted enzymes during nutrient-limited growth is utility dependent. Proc. Natl. Acad. Sci. USA 2017, 114, E7796–E7802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Stefano, M.; Miceli, E.; Gotti, S.; Missanelli, A.; Mazzocchi, S.; Corazza, G.R. The effect of oral alpha-galactosidase on intestinal gas production and gas-related symptoms. Dig. Dis. Sci. 2007, 52, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Oben, J.; Kothari, S.C.; Anderson, M.L. An open label study to determine the effects of an oral proteolytic enzyme system on whey protein concentrate metabolism in healthy males. J. Int. Soc. Sports Nutr. 2008, 5, 10. [Google Scholar] [CrossRef] [Green Version]
- Ianiro, G.; Pecere, S.; Giorgio, V.; Gasbarrini, A.; Cammarota, G. Digestive Enzyme Supplementation in Gastrointestinal Diseases. Curr. Drug Metab. 2016, 17, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Swami, O.C.; Shah, N.J. Functional dyspepsia and the role of digestive enzymes supplement in its therapy. Int. J. Basic Clin. Pharmacol. 2017, 6, 1035–1041. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Li, S.A.; Huang, C.H.; Su, H.H.; Chen, Y.H.; Chang, J.T.; Huang, S.S. Sirt1 Activation by Post-ischemic Treatment with Lumbrokinase Protects Against Myocardial Ischemia-Reperfusion Injury. Front. Pharmacol. 2018, 9, 636. [Google Scholar] [CrossRef] [Green Version]
- Altaf, F.; Wu, S.; Kasim, V. Role of Fibrinolytic Enzymes in Anti-Thrombosis Therapy. Front. Mol. Biosci. 2021, 8, 680397. [Google Scholar] [CrossRef]
- Townsend, J.R.; Morimune, J.E.; Jones, M.D.; Beuning, C.N.; Haase, A.A.; Boot, C.M.; Heffington, S.H.; Littlefield, L.A.; Henry, R.N.; Marshall, A.C.; et al. The Effect of ProHydrolase((R)) on the Amino Acid and Intramuscular Anabolic Signaling Response to Resistance Exercise in Trained Males. Sports 2020, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biller, J.A.; King, S.; Rosenthal, A.; Grand, R.J. Efficacy of lactase-treated milk for lactose-intolerant pediatric patients. J. Pediatr. 1987, 111, 91–94. [Google Scholar] [CrossRef]
- DiPalma, J.A.; Collins, M.S. Enzyme replacement for lactose malabsorption using a beta-D-galactosidase. J. Clin. Gastroenterol. 1989, 11, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Rosado, J.L.; Solomons, N.W.; Lisker, R.; Bourges, H. Enzyme replacement therapy for primary adult lactase deficiency. Effective reduction of lactose malabsorption and milk intolerance by direct addition of beta-galactosidase to milk at mealtime. Gastroenterology 1984, 87, 1072–1082. [Google Scholar] [CrossRef]
- Wei, G.; Helmerhorst, E.J.; Darwish, G.; Blumenkranz, G.; Schuppan, D. Gluten Degrading Enzymes for Treatment of Celiac Disease. Nutrients 2020, 12, 2095. [Google Scholar] [CrossRef] [PubMed]
- Deaton, J.; Cuentas, A.; Starnes, J. Tolerance and Efficacy of Glutalytic™: A Randomized, Double-Blind, Placebo Controlled Study. J. Nutr. Food Sci. 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Caio, G.; Volta, U.; Sapone, A.; Leffler, D.A.; De Giorgio, R.; Catassi, C.; Fasano, A. Celiac disease: A comprehensive current review. BMC Med. 2019, 17, 142. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Tapia, A.; Ludvigsson, J.F.; Brantner, T.L.; Murray, J.A.; Everhart, J.E. The prevalence of celiac disease in the United States. Am. J. Gastroenterol. 2012, 107, 1538–1544. [Google Scholar] [CrossRef]
- Aziz, I.; Hadjivassiliou, M.; Sanders, D.S. The spectrum of noncoeliac gluten sensitivity. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 516–526. [Google Scholar] [CrossRef]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef]
- Clemente, M.G.; De Virgiliis, S.; Kang, J.S.; Macatagney, R.; Musu, M.P.; Di Pierro, M.R.; Drago, S.; Congia, M.; Fasano, A. Early effects of gliadin on enterocyte intracellular signalling involved in intestinal barrier function. Gut 2003, 52, 218–223. [Google Scholar] [CrossRef]
- Abadie, V.; Jabri, B. IL-15: A central regulator of celiac disease immunopathology. Immunol. Rev. 2014, 260, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Leonard, M.M.; Sapone, A.; Catassi, C.; Fasano, A. Celiac Disease and Nonceliac Gluten Sensitivity: A Review. JAMA 2017, 318, 647–656. [Google Scholar] [CrossRef]
- Catassi, C.; Fabiani, E.; Iacono, G.; D’Agate, C.; Francavilla, R.; Biagi, F.; Volta, U.; Accomando, S.; Picarelli, A.; De Vitis, I.; et al. A prospective, double-blind, placebo-controlled trial to establish a safe gluten threshold for patients with celiac disease. Am. J. Clin. Nutr. 2007, 85, 160–166. [Google Scholar] [CrossRef]
- Halter, S.A.; Greene, H.L.; Helinek, G. Gluten-sensitive enteropathy: Sequence of villous regrowth as viewed by scanning electron microscopy. Hum. Pathol. 1982, 13, 811–818. [Google Scholar] [CrossRef]
- Halter, S.A.; Greene, H.L.; Helinek, G. Scanning electron microscopy of small intestinal repair following treatment for gluten sensitive enteropathy. Scan. Electron Microsc. 1980, 3, 155–161. [Google Scholar]
- Labeling, F.F. 21. CFR Part 101 [Docket No. FDA-2005-N-0404] RIN 0910-AG84. Federal Regist. 2013, 78, 47154–47179. [Google Scholar]
- Montserrat, V.; Bruins, M.J.; Edens, L.; Koning, F. Influence of dietary components on Aspergillus niger prolyl endoprotease mediated gluten degradation. Food Chem. 2015, 174, 440–445. [Google Scholar] [CrossRef]
- Scherf, K.A.; Wieser, H.; Koehler, P. Novel approaches for enzymatic gluten degradation to create high-quality gluten-free products. Food Res. Int. 2018, 110, 62–72. [Google Scholar] [CrossRef]
- Panda, R.; Garber, E.A.E. Western blot analysis of fermented-hydrolyzed foods utilizing gluten-specific antibodies employed in a novel multiplex competitive ELISA. Anal. Bioanal. Chem. 2019, 411, 5159–5174. [Google Scholar] [CrossRef]
- Rockendorf, N.; Meckelein, B.; Scherf, K.A.; Schalk, K.; Koehler, P.; Frey, A. Identification of novel antibody-reactive detection sites for comprehensive gluten monitoring. PLoS ONE 2017, 12, e0181566. [Google Scholar] [CrossRef] [Green Version]
- Panda, R.; Boyer, M.; Garber, E.A.E. A multiplex competitive ELISA for the detection and characterization of gluten in fermented-hydrolyzed foods. Anal. Bioanal. Chem. 2017, 409, 6959–6973. [Google Scholar] [CrossRef]
- Rivera Del Rio, A.; Keppler, J.K.; Boom, R.M.; Janssen, A.E.M. Protein acidification and hydrolysis by pepsin ensure efficient trypsin-catalyzed hydrolysis. Food Funct. 2021, 12, 4570–4581. [Google Scholar] [CrossRef]
- Nelson, G.M.; Guynn, J.M.; Chorley, B.N. Procedure and Key Optimization Strategies for an Automated Capillary Electrophoretic-based Immunoassay Method. J. Vis. Exp. 2017, 127, e55911. [Google Scholar] [CrossRef] [Green Version]
- Fu, B.X.; Sapirstein, H.D.; Bushuk, W. Salt-Induced Disaggregation/Solubilization of Gliadin and Glutenin Proteins in Water. J. Cereal Sci. 1996, 24, 241–246. [Google Scholar] [CrossRef]
- Tsuge, M.; Hiraga, N.; Zhang, Y.; Yamashita, M.; Sato, O.; Oka, N.; Shiraishi, K.; Izaki, Y.; Makokha, G.N.; Uchida, T.; et al. Endoplasmic reticulum-mediated induction of interleukin-8 occurs by hepatitis B virus infection and contributes to suppression of interferon responsiveness in human hepatocytes. Virology 2018, 525, 48–61. [Google Scholar] [CrossRef]
- Njock, M.S.; Cheng, H.S.; Dang, L.T.; Nazari-Jahantigh, M.; Lau, A.C.; Boudreau, E.; Roufaiel, M.; Cybulsky, M.I.; Schober, A.; Fish, J.E. Endothelial cells suppress monocyte activation through secretion of extracellular vesicles containing antiinflammatory microRNAs. Blood 2015, 125, 3202–3212. [Google Scholar] [CrossRef] [Green Version]
- Steinberg-Shemer, O.; Keel, S.; Dgany, O.; Walsh, T.; Noy-Lotan, S.; Krasnov, T.; Yacobovich, J.; Quarello, P.; Ramenghi, U.; King, M.C.; et al. Diamond Blackfan Anemia: A Nonclassical Patient with Diagnosis Assisted by Genomic Analysis. J. Pediatr. Hematol. Oncol. 2016, 38, e260–e262. [Google Scholar] [CrossRef] [Green Version]
- Chekhonin, V.P.; Lebedev, S.V.; Volkov, A.I.; Pavlov, K.A.; Ter-Arutyunyants, A.A.; Volgina, N.E.; Savchenko, E.A.; Grinenko, N.F.; Lazarenko, I.P. Activation of expression of brain-derived neurotrophic factor at the site of implantation of allogenic and xenogenic neural stem (progenitor) cells in rats with ischemic cortical stroke. Bull. Exp. Biol. Med. 2011, 150, 515–518. [Google Scholar] [CrossRef]
- Lacorn, M.; Lindeke, S.; Siebeneicher, S.; Weiss, T. Commercial ELISA Measurement of Allergens and Gluten: What We Can Learn from Case Studies. J. AOAC Int. 2018, 101, 102–107. [Google Scholar] [CrossRef]
- Panda, R.; Garber, E.A.E. Detection and Quantitation of Gluten in Fermented-Hydrolyzed Foods by Antibody-Based Methods: Challenges, Progress, and a Potential Path Forward. Front. Nutr. 2019, 6, 97. [Google Scholar] [CrossRef]
- Jelinkova, L.; Tuckova, L.; Cinova, J.; Flegelova, Z.; Tlaskalova-Hogenova, H. Gliadin stimulates human monocytes to production of IL-8 and TNF-alpha through a mechanism involving NF-kappaB. FEBS Lett. 2004, 571, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Lammers, K.M.; Khandelwal, S.; Chaudhry, F.; Kryszak, D.; Puppa, E.L.; Casolaro, V.; Fasano, A. Identification of a novel immunomodulatory gliadin peptide that causes interleukin-8 release in a chemokine receptor CXCR3-dependent manner only in patients with coeliac disease. Immunology 2011, 132, 432–440. [Google Scholar] [CrossRef]
- Kumar, S.; Prenner, S. Dyspepsia and Increased Levels of Liver Enzymes in a 24-Year-Old Man. Gastroenterology 2019, 157, 27–28. [Google Scholar] [CrossRef] [Green Version]
- De Palma, G.; Nadal, I.; Collado, M.C.; Sanz, Y. Effects of a gluten-free diet on gut microbiota and immune function in healthy adult human subjects. Br. J. Nutr. 2009, 102, 1154–1160. [Google Scholar] [CrossRef] [Green Version]
- Cinova, J.; Palova-Jelinkova, L.; Smythies, L.E.; Cerna, M.; Pecharova, B.; Dvorak, M.; Fruhauf, P.; Tlaskalova-Hogenova, H.; Smith, P.D.; Tuckova, L. Gliadin peptides activate blood monocytes from patients with celiac disease. J. Clin. Immunol. 2007, 27, 201–209. [Google Scholar] [CrossRef]
- Capozzi, A.; Vincentini, O.; Gizzi, P.; Porzia, A.; Longo, A.; Felli, C.; Mattei, V.; Mainiero, F.; Silano, M.; Sorice, M.; et al. Modulatory Effect of Gliadin Peptide 10-mer on Epithelial Intestinal CACO-2 Cell Inflammatory Response. PLoS ONE 2013, 8, e66561. [Google Scholar] [CrossRef] [Green Version]
- Palova-Jelinkova, L.; Rozkova, D.; Pecharova, B.; Bartova, J.; Sediva, A.; Tlaskalova-Hogenova, H.; Spisek, R.; Tuckova, L. Gliadin fragments induce phenotypic and functional maturation of human dendritic cells. J. Immunol. 2005, 175, 7038–7045. [Google Scholar] [CrossRef] [Green Version]
- Grover, J.; Chhuneja, P.; Midha, V.; Ghia, J.E.; Deka, D.; Mukhopadhyay, C.S.; Sood, N.; Mahajan, R.; Singh, A.; Verma, R.; et al. Variable Immunogenic Potential of Wheat: Prospective for Selection of Innocuous Varieties for Celiac Disease Patients via in vitro Approach. Front. Immunol. 2019, 10, 84. [Google Scholar] [CrossRef] [Green Version]
- Tuckova, L.; Novotna, J.; Novak, P.; Flegelova, Z.; Kveton, T.; Jelinkova, L.; Zidek, Z.; Man, P.; Tlaskalova-Hogenova, H. Activation of macrophages by gliadin fragments: Isolation and characterization of active peptide. J. Leukoc. Biol. 2002, 71, 625–631. [Google Scholar]
- Vincentini, O.; Maialetti, F.; Gonnelli, E.; Silano, M. Gliadin-dependent cytokine production in a bidimensional cellular model of celiac intestinal mucosa. Clin. Exp. Med. 2015, 15, 447–454. [Google Scholar] [CrossRef]
- Goel, G.; Tye-Din, J.A.; Qiao, S.W.; Russell, A.K.; Mayassi, T.; Ciszewski, C.; Sarna, V.K.; Wang, S.; Goldstein, K.E.; Dzuris, J.L.; et al. Cytokine release and gastrointestinal symptoms after gluten challenge in celiac disease. Sci. Adv. 2019, 5, eaaw7756. [Google Scholar] [CrossRef] [Green Version]
- Goel, G.; Daveson, A.J.M.; Hooi, C.E.; Tye-Din, J.A.; Wang, S.; Szymczak, E.; Williams, L.J.; Dzuris, J.L.; Neff, K.M.; Truitt, K.E.; et al. Serum cytokines elevated during gluten-mediated cytokine release in coeliac disease. Clin. Exp. Immunol. 2020, 199, 68–78. [Google Scholar] [CrossRef]
- Tye-Din, J.A.; Skodje, G.I.; Sarna, V.K.; Dzuris, J.L.; Russell, A.K.; Goel, G.; Wang, S.; Goldstein, K.E.; Williams, L.J.; Sollid, L.M.; et al. Cytokine release after gluten ingestion differentiates coeliac disease from self-reported gluten sensitivity. United Eur. Gastroenterol. J. 2020, 8, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Di Sabatino, A.; Giuffrida, P.; Fornasa, G.; Salvatore, C.; Vanoli, A.; Naviglio, S.; De Leo, L.; Pasini, A.; De Amici, M.; Alvisi, C.; et al. Innate and adaptive immunity in self-reported nonceliac gluten sensitivity versus celiac disease. Dig. Liver Dis. 2016, 48, 745–752. [Google Scholar] [CrossRef]
- Di Sabatino, A.; Corazza, G.R. True Nonceliac Gluten Sensitivity in Real Patients. Clin. Gastroenterol. Hepatol. 2016, 14, 168–169. [Google Scholar] [CrossRef]
- Brottveit, M.; Beitnes, A.C.; Tollefsen, S.; Bratlie, J.E.; Jahnsen, F.L.; Johansen, F.E.; Sollid, L.M.; Lundin, K.E. Mucosal cytokine response after short-term gluten challenge in celiac disease and non-celiac gluten sensitivity. Am. J. Gastroenterol. 2013, 108, 842–850. [Google Scholar] [CrossRef]
- Heydari, F.; Rostami-Nejad, M.; Moheb-Alian, A.; Mollahoseini, M.H.; Rostami, K.; Pourhoseingholi, M.A.; Aghamohammadi, E.; Zali, M.R. Serum cytokines profile in treated celiac disease compared with non-celiac gluten sensitivity and control: A marker for differentiation. J. Gastrointest. Liver Dis. 2018, 27, 241–247. [Google Scholar] [CrossRef]
- Manavalan, J.S.; Hernandez, L.; Shah, J.G.; Konikkara, J.; Naiyer, A.J.; Lee, A.R.; Ciaccio, E.; Minaya, M.T.; Green, P.H.; Bhagat, G. Serum cytokine elevations in celiac disease: Association with disease presentation. Hum. Immunol. 2010, 71, 50–57. [Google Scholar] [CrossRef]
- Chowers, Y.; Marsh, M.N.; De Grandpre, L.; Nyberg, A.; Theofilopoulos, A.N.; Kagnoff, M.F. Increased proinflammatory cytokine gene expression in the colonic mucosa of coeliac disease patients in the early period after gluten challenge. Clin. Exp. Immunol. 1997, 107, 141–147. [Google Scholar] [CrossRef]
- Street, M.E.; Volta, C.; Ziveri, M.A.; Zanacca, C.; Banchini, G.; Viani, I.; Rossi, M.; Virdis, R.; Bernasconi, S. Changes and relationships of IGFS and IGFBPS and cytokines in coeliac disease at diagnosis and on gluten-free diet. Clin. Endocrinol. 2008, 68, 22–28. [Google Scholar] [CrossRef]
- Lahat, N.; Shapiro, S.; Karban, A.; Gerstein, R.; Kinarty, A.; Lerner, A. Cytokine profile in coeliac disease. Scand J. Immunol. 1999, 49, 441–446. [Google Scholar] [CrossRef]
- Mohan Kumar, B.V.; Vijaykrishnaraj, M.; Kurrey, N.K.; Shinde, V.S.; Prabhasankar, P. Prolyl endopeptidase-degraded low immunoreactive wheat flour attenuates immune responses in Caco-2 intestinal cells and gluten-sensitized BALB/c mice. Food Chem. Toxicol. 2019, 129, 466–475. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Antibody | ELISA Kits | Manufacturer | Dilution | ||
|---|---|---|---|---|---|
| ELISA | Immunoblot | ||||
| Gtox-G12 | G12 | Gluten Tox ELISA Competitive G12 | Biomedal Diagnostics | to 30 ng/mL * | to 200 ng/mL |
| A-G12 | G12 | AgraQuant Gluten G12 | Romer Labs | 1 to 3 | 1 to 1 |
| R5Sand | R5 | RIDASCREEN Gliadin | R-BioPharm, AG | 1 to 35 | 1 to 5 |
| R5Comp | R5 | RIDASCREEN Gliadin Competitive | R-BioPharm, AG | 1 to 100 | 1 to 15 |
| V10-R5 | R5 | Veratox for Gliadin R5 (Cat # 8510) | Neogen Corp. | 1 to 15 | 1 to 10 |
| V80-GL | USDA | Veratox for Gliadin (Cat # 8480) | Neogen Corp. | 1 to 10 | 1 to 3 |
| MI-GL | MIoBS | Wheat/Gluten (Gliadin) MIoBS | Morinaga Institution of Biological Sciences Inc | 1 to 3 | 1 to 1 |
| AllSK | Skerritt | AllerTek Gluten | ELISA Technologies Inc. | 1 to 15 | 1 to 10 |
| 2D4 | 2D4 | Microbiologique Gluten Sandwich | Pi BioScientific Inc. | 1 to 10 | 1 to 1 |
| Target | Forward Primer | Reverse Primer | Reference |
|---|---|---|---|
| IL-8 | CAGTTTTGCCAAGGAGTGCT | CAACCCTCTGCACCCAGTTT | [36] |
| TNF-A | GCCAGAGGGCTGATTAGAGA | TCTTCTGCCTGCTGCACTT | [37] |
| RPL5 | GGTCTCTGTTCCGCAGGATG | CAGTTTTACCCTCTCGTCGTCT | [38] |
| GAPDH | CTTTGACGCTGGGGCTGGCATT | TTGTGCTCTTGCTGGGGCTGGT | [39] |
| Fermentation Product | Commercial ELISA Kit | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gtox-G12 | A-G12 | R5Sand | R5Comp | V10-R5 | V80-GL | MI-GL | AllSK | 2D4 | ||
| Antibody | ||||||||||
| G12 | G12 | R5 | R5 | R5 | USDA | MIoBS | Skerritt | 2D4 | ||
| PT-Glu | Pepsin-trypsin hydrolysis | 77.5 ± NA | 33.5 ± NA | 1.5 ± NA | 1.0 ± NA | 1.0 ± NA | 2.5 ± NA | 22.0 ± NA | 28.0 ± NA | 32 ± NA |
| Alpha-G | α-Galactosidase | ND | 0.0 ± NA | 0.36 ± 0.1 | 3.2 ± 3.0 | ND | ND | 5.3 ± 0.8 | ND | 1.63 ± 1.43 |
| ALA | Acid Lactase | ND | 0.5 ± NA | ND | 1.7 ± 1.7 | 2.0 ± 0.2 | ND | 2.2 ± 1.2 | ND | 0.1 ± NA |
| ASP | Acid Stable Protease | ND | 0.4 ± NA | 0.4 ± 0.1 | 2.9 ± 0.5 | 4.3 ± 0.2 | 2.4 ± NA | 1.6 ± 0.8 | ND | 3.8 ± 3.5 |
| FL | Fungal Lactase | ND | 0.4 ± NA | 0.0 ± 0.0 | 2.3 ± 0.1 | 4.9 ± 0.2 | ND | 3.4 ± 1.8 | 1.42 ± NA | 3.4 ± 3.3 |
| FLC | Fungal Lactase Conc. | ND | 0.8 ± NA | 0.4 ± 0.4 | 1.1 ± 0.6 | 8.0 ± 3.3 | 0.2 ± NA | 6.7 ± 0.8 | ND | 2.2 ± 2.1 |
| PI | Peptidase I | ND | 1.8 ± 0.1 | 0.8 ± 0.6 | 3.1 ± 0.5 | 0.4 ± 0.3 | 0.6 ± NA | 2.9 ± 2.4 | ND | 3.4 ± 2.9 |
| PIC | Peptidase I Conc. | ND | 1.4 ± 0.6 | 0.3 ± 0.1 | 1.4 ± 0.2 | 0.7 ± NA | 2.4 ± 1.3 | 1.4 ± 2.5 | ND | 2.4 ± 1.5 |
| PII | Peptidase II | ND | 0.8 ± NA | 0.2 ± 0.1 | 2.3 ± 0.1 | 6.5 ± 4.6 | ND | 2.8 ± 0.1 | ND | 4.5 ± 4.1 |
| PIIC | Peptidase II Conc. | ND | 0.8 ± 0.2 | 3.9 ± 3.1 | 4.0 ± 0.1 | 1.4 ± 0.3 | 0.2 ± NA | 3.7 ± 2.8 | ND | 3.5 ± 2.56 |
| Alpha-G-DE | α-Galactosidase II | ND | 0.8 ± NA | 0.5 ± NA | 2.3 ± 0.1 | 1.2 ± NA | 0.1 ± NA | 3.0 ± 1.4 | ND | 4.3 ± 3.4 |
| FPA | Fungal Protease A | ND | 0.8 ± 0.2 | 1.0 ± 0.7 | 6.8 ± 8.0 | 4.6 ± 4.8 | 6.7 ± 2.0 | 3.2 ± 1.0 | 1.0 ± NA | 4.3 ± 4.0 |
| BP | Bacterial Protease | 137 ± 4.0 | 85.5 ± 42.8 | 9.5 ± 4.1 | 3.1 ± 1.6 | 18.3 ± 1.0 | 14.9 ± 2.1 | 4.9 ± 2.1 | 1.8 ± 0.0 | 7.7 ± 2.9 |
| LipAN | Lipase AN | ND | 0.1 ± 0.1 | 4.1 ± 3.4 | 2.9 ± 0.2 | 3.2 ± 0.9 | 1.7 ± 1.7 | 3.0 ± 2.4 | ND | 4.8 ± 3.3 |
| CellAN | Cellulase AN | ND | 0.5 ± 0.4 | 0.4 ± 0.4 | 2.5 ± 0.3 | 1.2 ± 1.0 | 1.6 ± NA | 2.4 ± 2.1 | ND | 3.7 ± 3.7 |
| Fermentation Product | Commercial ELISA Kit | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gtox-G12 | A-G12 | R5Sand | R5Comp | V10-R5 | V80-GL | MI-GL | AllSK | 2D4 | ||
| Antibody | ||||||||||
| G12 | G12 | R5 | R5 | R5 | USDA | MIoBS | Skerritt | 2D4 | ||
| PT-Glu | Pepsin-trypsin hydrolysis | ND | ND | ND | ND | 36.5 | ND | ND | ND | 1.5 |
| Alpha-G | α-Galactosidase | ND | ND | ND | ND | ND | ND | ND | ND | 677.08 |
| ALA | Acid Lactase | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| ASP | Acid Stable Protease | ND | ND | ND | ND | 26.65 | ND | ND | ND | ND |
| FL | Fungal Lactase | ND | ND | ND | 95.86 | ND | ND | ND | ND | ND |
| FLC | Fungal Lactase Conc. | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| PI | Peptidase I | ND | ND | ND | ND | ND | ND | ND | 606.66 | ND |
| PIC | Peptidase I Conc. | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| PII | Peptidase II | ND | ND | ND | ND | ND | ND | ND | ND | 519.23 |
| PIIC | Peptidase II Conc. | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| Alpha-G-DE | α-Galactosidase II | ND | ND | 0.72 | ND | ND | ND | ND | ND | ND |
| FPA | Fungal Protease A | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| BP | Bacterial Protease | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| LipAN | Lipase AN | ND | ND | ND | ND | 29.53 | ND | ND | ND | ND |
| CellAN | Cellulase AN | ND | ND | ND | ND | ND | ND | ND | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khokhlova, E.; Kim, P.; Colom, J.; Bhat, S.; Curran, A.M.; Jouini, N.; Rea, K.; Phipps, C.; Deaton, J. Assessing Hydrolyzed Gluten Content in Dietary Enzyme Supplements Following Fermentation. Fermentation 2022, 8, 203. https://doi.org/10.3390/fermentation8050203
Khokhlova E, Kim P, Colom J, Bhat S, Curran AM, Jouini N, Rea K, Phipps C, Deaton J. Assessing Hydrolyzed Gluten Content in Dietary Enzyme Supplements Following Fermentation. Fermentation. 2022; 8(5):203. https://doi.org/10.3390/fermentation8050203
Chicago/Turabian StyleKhokhlova, Ekaterina, Pyeongsug Kim, Joan Colom, Shaila Bhat, Aoife M. Curran, Najla Jouini, Kieran Rea, Christopher Phipps, and John Deaton. 2022. "Assessing Hydrolyzed Gluten Content in Dietary Enzyme Supplements Following Fermentation" Fermentation 8, no. 5: 203. https://doi.org/10.3390/fermentation8050203
APA StyleKhokhlova, E., Kim, P., Colom, J., Bhat, S., Curran, A. M., Jouini, N., Rea, K., Phipps, C., & Deaton, J. (2022). Assessing Hydrolyzed Gluten Content in Dietary Enzyme Supplements Following Fermentation. Fermentation, 8(5), 203. https://doi.org/10.3390/fermentation8050203