
Fenton-Mediated Chlorophenol Degradation by Iron-Reducing Compounds Isolated from Endophytic Fungi in Atacama Puna Plateau Lecanicillium ATA01

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture, DNA Extraction, Amplification, Sequencing, and Phylogenetic Analysis
2.2. Morphological and Molecular Characterization of Fungal Isolate
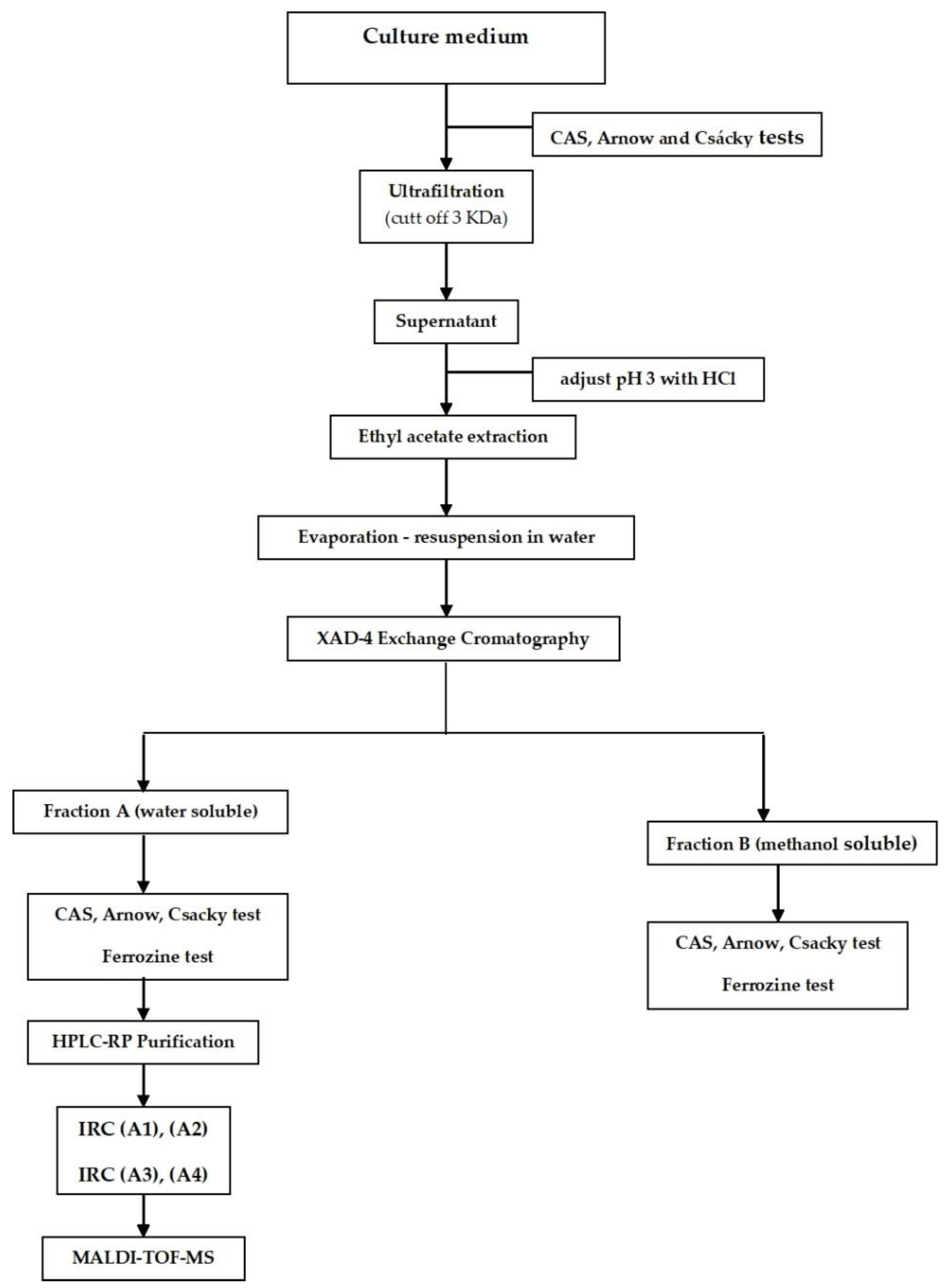
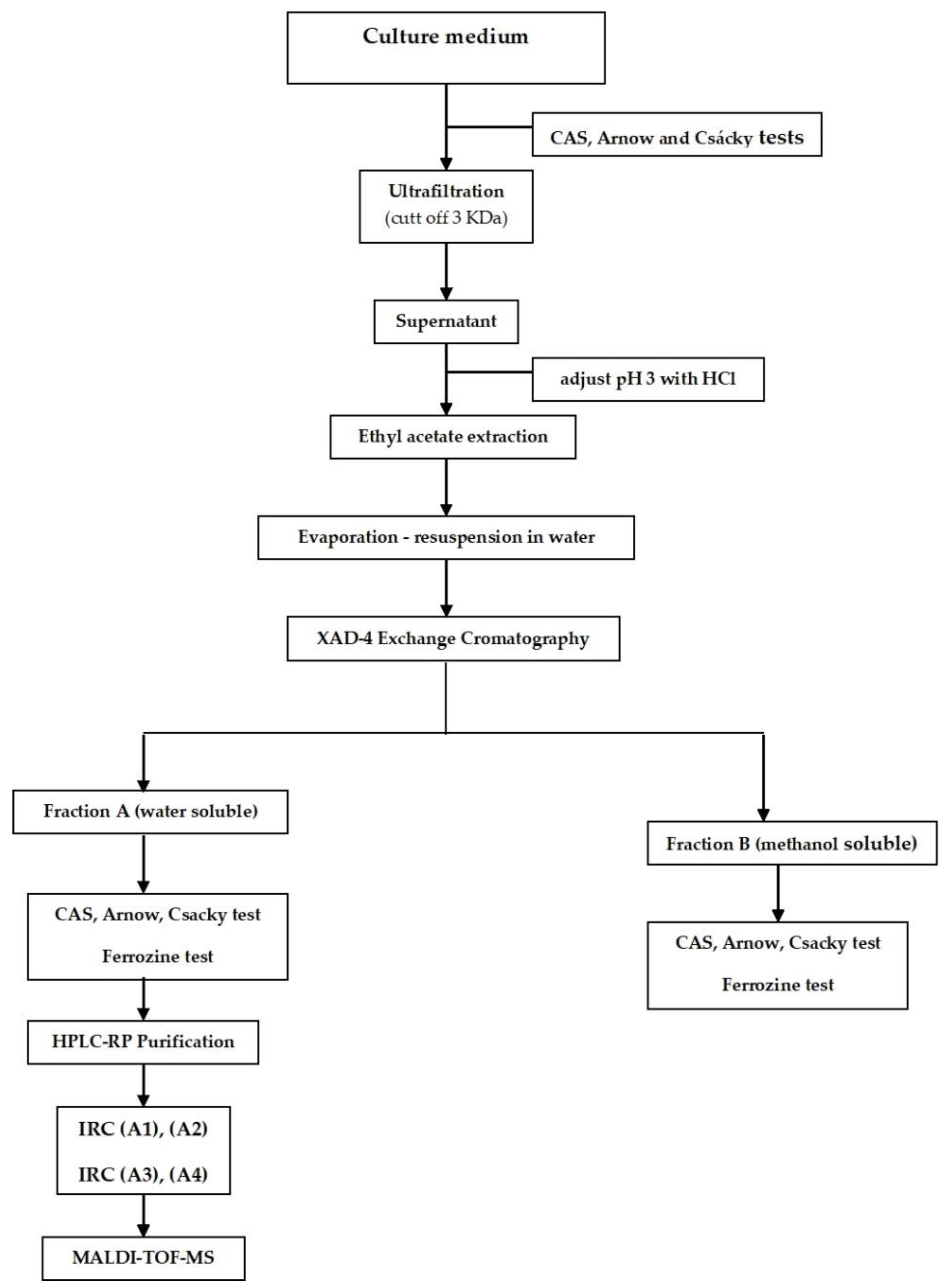
2.3. Production, Purification, and Characterization of IRCs
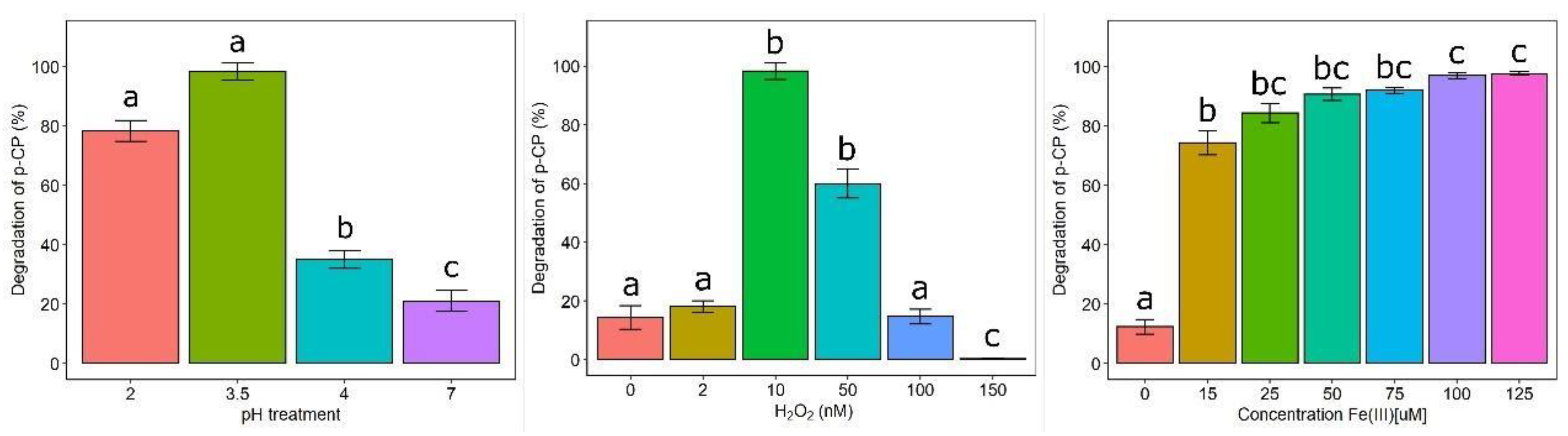
2.4. Optimization of Chlorophenol Degradation by IRC
2.5. DTPA-Fe+2 and EDTA-Fe+2 Iron Complexes
2.6. Degradation of Chlorophenols
2.7. Monitoring of Chlorophenol Degradation by High-Performance Liquid Chromatography (HPLC)
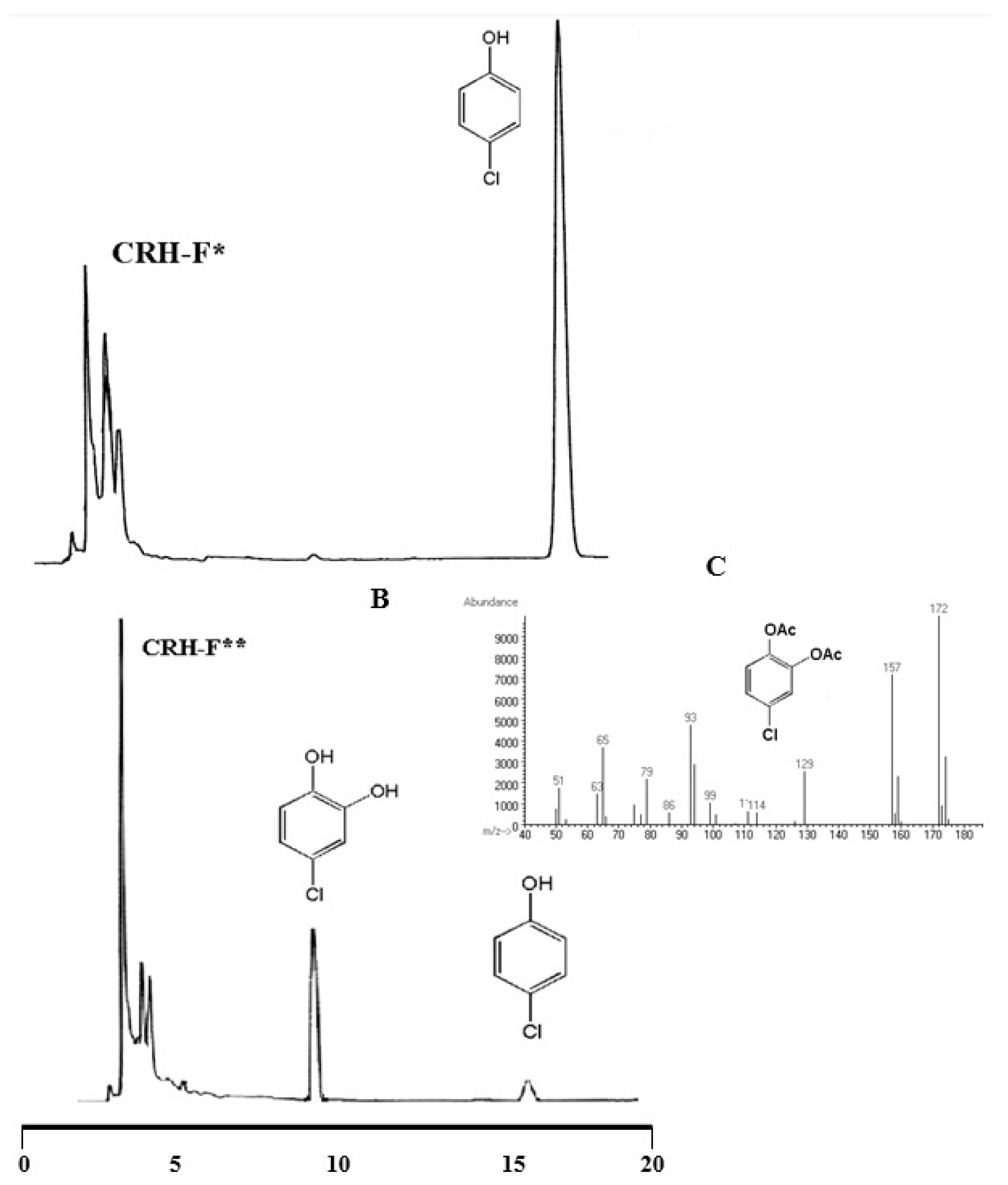
2.8. Identification of Degradation Products
2.9. Rate Constant and Yield of Chlorophenol Degradation by Fenton-like Reaction
2.10. Statistical Analyses
3. Results
3.1. Fungal Identification by Morphological and Molecular Characterization
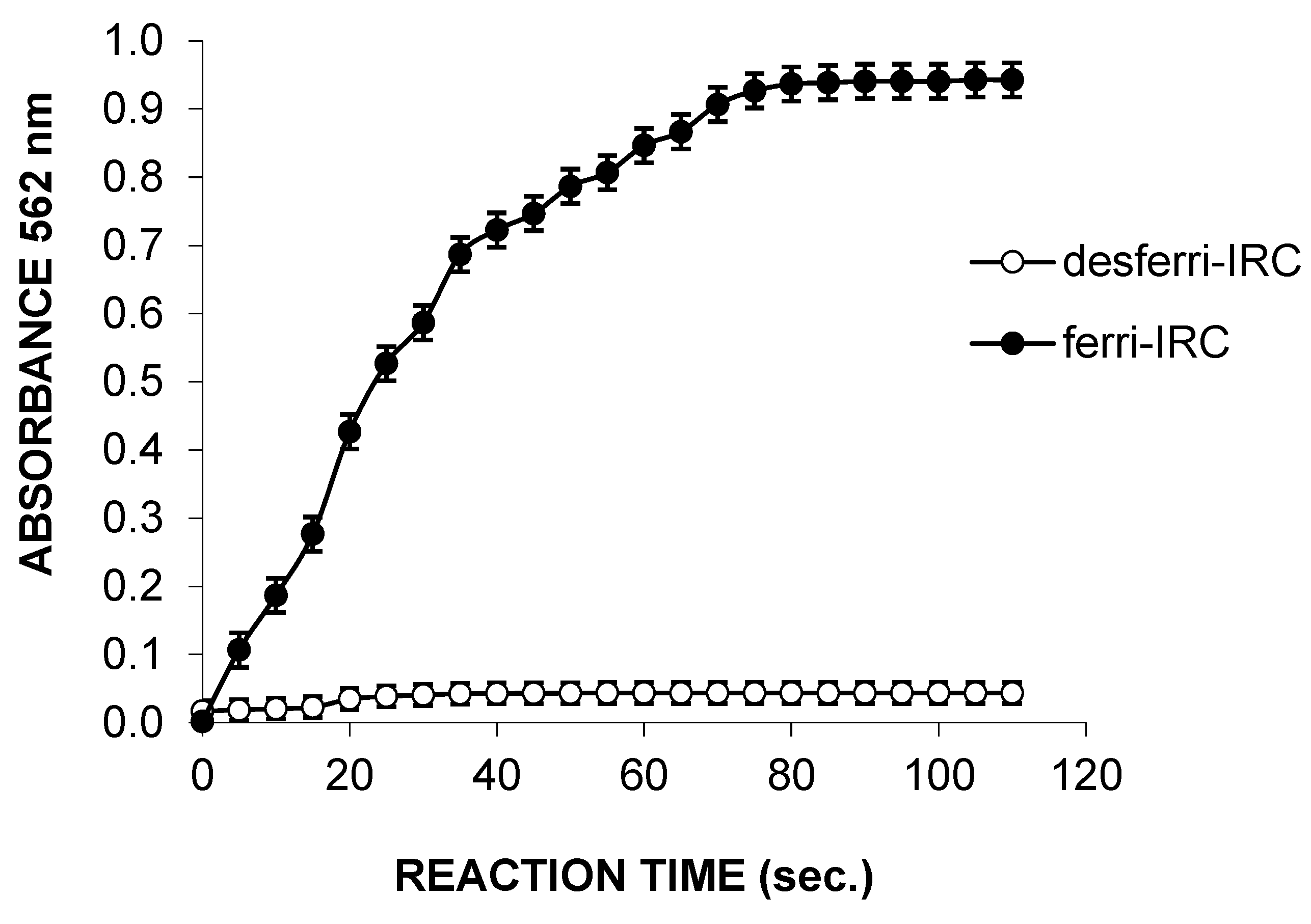
3.2. Production and Characterization of IRCs
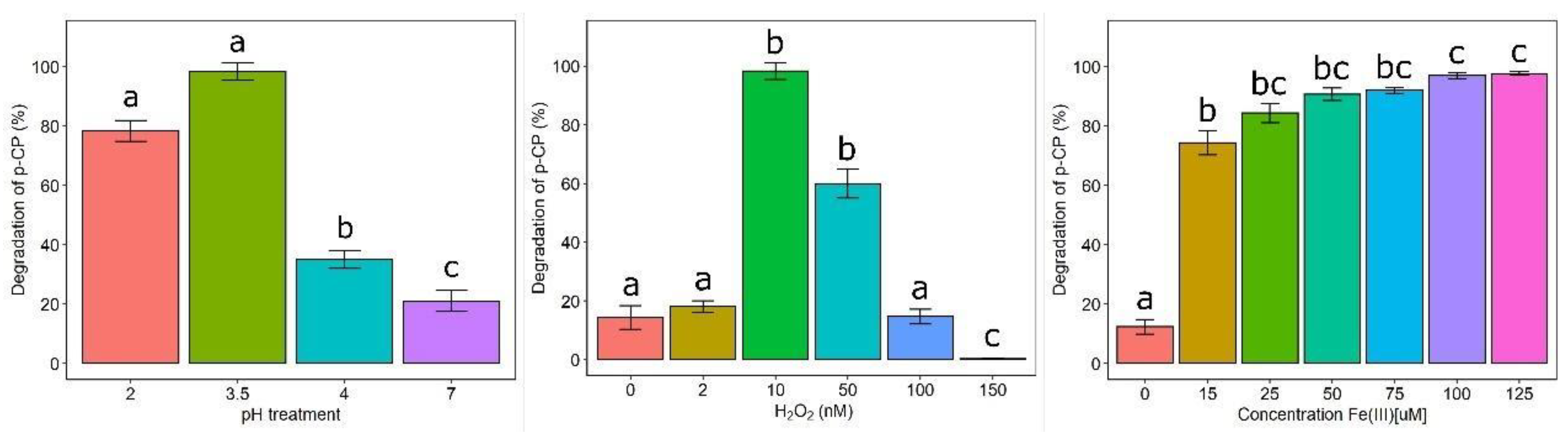
3.3. Optimization of Chlorophenol Degradation
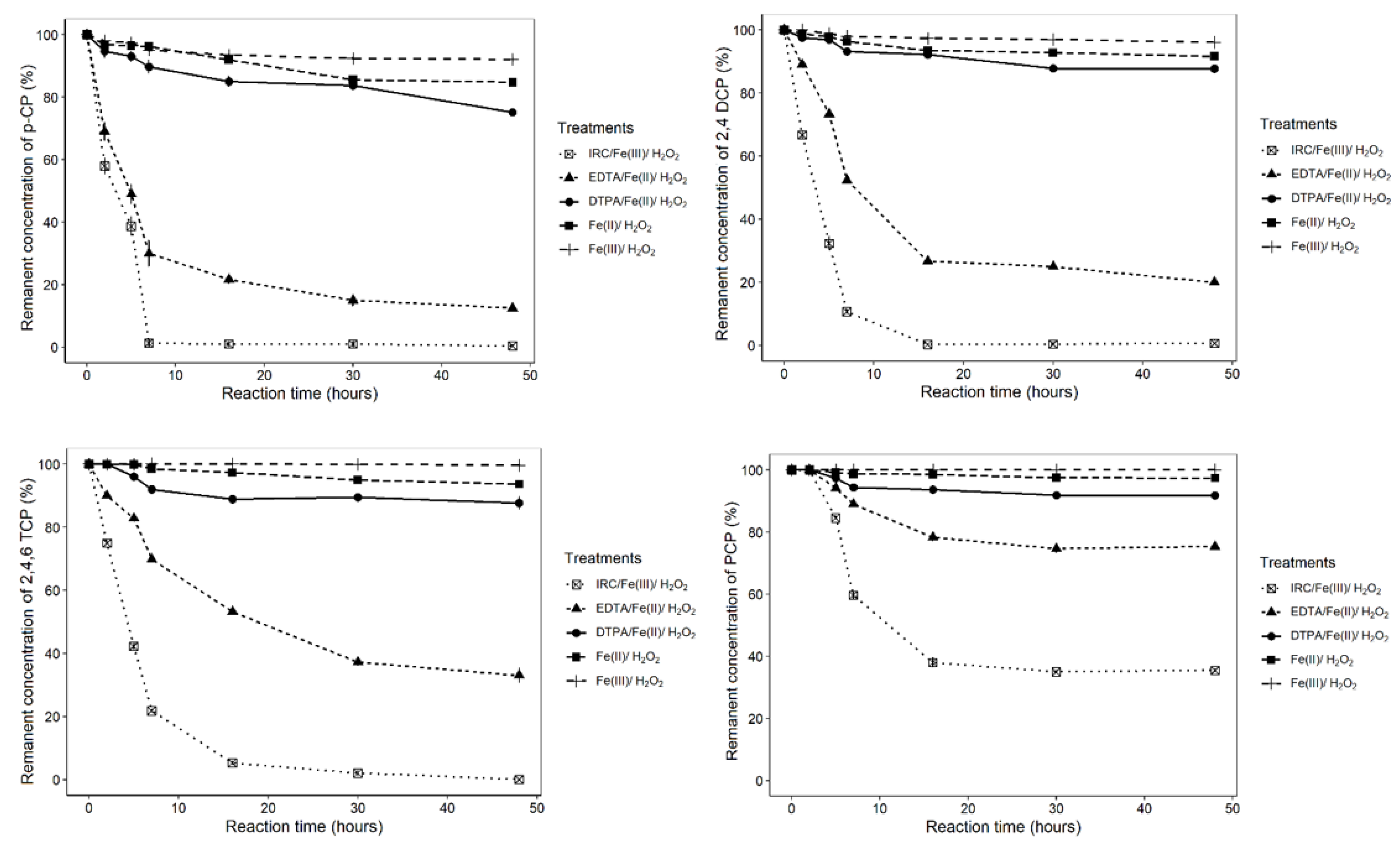
3.4. Degradation of Chlorinated Phenolic Compounds by Ferric-IRCs
3.5. Degradation of Chlorophenols by f-IRC and Synthetic Iron Chelates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arora, P.K.; Bae, H. Bacterial degradation of chlorophenols and their derivatives. Microb. Cell Factories 2014, 13, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igbinosa, E.; Odjadjare, E.; Chigor, V.; Igbinosa, I.H.; Emoghene, A.O.; Ekhaise, F.O.; Igiehon, N.O.; Idemudia, O.G. Toxicological Profile of Chlorophenols and Their Derivatives in the Environment: The Public Health Perspective. Sci. World J. 2013, 2013, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pera-Titus, M.; García-Molina, V.; Baños, M.A.; Giménez, J.; Esplugas, S. Degradation of chlorophenols by means of advanced oxidation processes: A general review. Appl. Catal. B Environ. 2004, 47, 219–256. [Google Scholar] [CrossRef]
- Czaplicka, M. Sources and transformations of chlorophenols in the natural environment. Sci. Total Environ. 2004, 322, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Oses, R.; Valenzuela, S.; Freer, J.; Sanfuentes, E.; Rodriguez, J. Fungal endophytes in xylem of healthy Chilean trees and their possible role in early wood decay. Fungal Divers 2008, 33, 77–86. Available online: https://www.fungaldiversity.org/fdp/sfdp/33-4.pdf (accessed on 11 February 2022).
- Oses, R.; Valenzuela, S.; Freer, J.; Baeza, J.; Rodríguez, J. Evaluation of fungal endophytes for lignocellulolytic enzyme production and wood biodegradation. Int. Biodeterior. Biodegrad. 2006, 57, 129–135. [Google Scholar] [CrossRef]
- Parray, J.A.; Shameem, N. Sustainable engineering technologies to promote activities of beneficial microbiome. In Sustainable Agriculture; Academic Pres: San Diego, CA, USA, 2020; pp. 231–275. [Google Scholar] [CrossRef]
- Arantes, V.; Jellison, J.; Goodell, B. Peculiarities of brown-rot fungi and biochemical Fenton reaction with regard to their potential as a model for bioprocessing biomass. Appl. Microbiol. Biotechnol. 2012, 94, 323–338. [Google Scholar] [CrossRef]
- Gόmez-Toribio, V.; García-Martín, A.B.; Martínez, M.J.; Martínez, A.T.; Guillén, F. Induction of Extracellular Hydroxyl Radical Production by White-Rot Fungi through Quinone Redox Cycling. Appl. Environ. Microbiol. 2009, 75, 3944–3953. [Google Scholar] [CrossRef] [Green Version]
- Hammel, K.E.; Kapich, A.N.; Jensen, K.A.; Ryan, Z.C. Reactive oxygen species as agents of wood decay by fungi. Enzym. Microb. Technol. 2002, 30, 445–453. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, M. A critical review of the application of chelating agents to enable Fenton and Fenton-like reactions at high pH values. J. Hazard. Mater. 2018, 362, 436–450. [Google Scholar] [CrossRef]
- Goodell, B.; Jellison, J.; Liu, J.; Daniel, G.; Paszczynski, A.; Fekete, F.; Krishnamurthy, S.; Jun, L.; Xu, G. Low molecular weight chelators and phenolic compounds isolated from wood decay fungi and their role in the fungal biodegradation of wood. J. Biotechnol. 1997, 53, 133–162. [Google Scholar] [CrossRef]
- Kerem, Z.; Jensen, K.A.; Hammel, K. Biodegradative mechanism of the brown rot basidiomyceteGloeophyllum trabeum: Evidence for an extracellular hydroquinone-driven fenton reaction. FEBS Lett. 1999, 446, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Mukhopadhyay, M.; Murthy, Z.V.P. Treatment of chlorophenols from wastewaters by advanced oxidation pro-cesses. Sep. Purif. Rev. 2013, 42, 263–295. [Google Scholar] [CrossRef]
- Rodríguez, J.; Parra, C.; Contreras, D.; Freer, J.; Baeza, J. Dihydroxybenzenes: Driven Fenton reactions. Water Sci. Technol. 2001, 44, 251–256. [Google Scholar] [CrossRef]
- Paszczynski, A.; Crawford, R.; Funk, D.; Goodell, B. De novo synthesis of 4, 5-Dimethoxycatechol and 2, 5-Dimethoxyhydroquinone by the brown-rot fungus Gloeophyllum trabeum. Appl. Environ. Microbiol. 1999, 65, 674–679. [Google Scholar] [CrossRef] [Green Version]
- Marco-Urrea, E.; García-Romera, I.; Aranda, E. Potential of non-ligninolytic fungi in bioremediation of chlorinated and polycyclic aromatic hydrocarbons. New Biotechnol. 2015, 32, 620–628. [Google Scholar] [CrossRef]
- Rocha-Pino, Z.; Marín-Cervantes, M.D.C.; Martínez-Archundia, M.; Soriano-Blancas, E.; Revah, S.; Shirai, K. Morphological changes, chitinolytic enzymes and hydrophobin-like proteins as responses of Lecanicillium lecanii during growth with hydrocarbon. Bioprocess Biosyst. Eng. 2012, 36, 531–539. [Google Scholar] [CrossRef]
- Pozdnyakova, N.N.; Varese, G.C.; Prigione, V.; Dubrovskaya, E.V.; Balandina, S.A.; Turkovskaya, O.V. Degradative properties of two newly isolated strains of the ascomycetes Fusarium oxysporum and Lecanicillium aphanocladii. Int. Microbiol. 2018, 22, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Fenice, M. The Psychrotolerant Antarctic Fungus Lecanicillium muscarium CCFEE 5003: A Powerful Producer of Cold-Tolerant Chitinolytic Enzymes. Molecules 2016, 21, 447. [Google Scholar] [CrossRef] [Green Version]
- Jaber, L.R.; Ownley, B. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Radwan, O.; Gunasekera, T.S.; Ruiz, O.N. Draft Genome Sequence of Lecanicillium sp. Isolate LEC01, a Fungus Capable of Hydrocarbon Degradation. Microbiol. Resour. Announc. 2019, 8, e01744-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Lefort, V.; Longueville, J.-E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Anisimova, M.; Gil, M.; Dufayard, J.-F.; Dessimoz, C.; Gascuel, O. Survey of Branch Support Methods Demonstrates Accuracy, Power, and Robustness of Fast Likelihood-based Approximation Schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Csáky, T.Z.; Hassel, O.; Rosenberg, T.; Loukamo, S.L.; Turunen, E.; Tuhkanen, A. On the Estimation of Bound Hydroxylamine in Biological Materials. Acta Chem. Scand. 1948, 2, 450–454. [Google Scholar] [CrossRef] [Green Version]
- Arnow, L.E. Colourimetric determination of the components of 3,4-dihydroxyphenylalanine-tyrosine mixtures. J. Biol. Chem 1937, 118, 531–537. Available online: https://www.jbc.org/article/S0021-9258(18)74509-2/pdf (accessed on 11 February 2022). [CrossRef]
- Voelker, B.M.; Sulzberger, B. Effects of Fulvic Acid on Fe(II) Oxidation by Hydrogen Peroxide. Environ. Sci. Technol. 1996, 30, 1106–1114. [Google Scholar] [CrossRef]
- Shao, Y.; Lin, A.H.-M. Improvement in the quantification of reducing sugars by miniaturizing the Somogyi-Nelson assay using a microtiter plate. Food Chem. 2018, 240, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cohen, G.; Sinet, P.M. The fenton reaction between ferrous-diethylenetriaminepentaacetic acid and hydrogen peroxide. FEBS Lett. 1982, 138, 258–260. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 11 February 2022).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Turkovskaya, O.; Dubrovskaya, E.; Grinev, V.; Balandina, S.; Pozdnyakova, N.; Institute of Biochemistry and Physiology of Plants and Microorganisms Ras. Degradative Activity and Production of the Extracellular Peroxidases by Micromycetes with Different Ecological Strategy. Sel’skokhozyaistvennaya Biol. 2019, 54, 65–75. [Google Scholar] [CrossRef]
- Ben Hammouda, S.; Adhoum, N.; Monser, L. Chemical oxidation of a malodorous compound, indole, using iron entrapped in calcium alginate beads. J. Hazard. Mater. 2016, 301, 350–361. [Google Scholar] [CrossRef]
- Ma, J.; Song, W.; Chen, C.; Ma, W.; Zhao, J.; Tang, Y. Fenton Degradation of Organic Compounds Promoted by Dyes under Visible Irradiation. Environ. Sci. Technol. 2005, 39, 5810–5815. [Google Scholar] [CrossRef]
- Barbeni, M.; Minero, C.; Pelizzetti, E. Chemical degradation of chlorophenols with Fenton’s reagent. Chemosphere 1987, 16, 2225–2237. [Google Scholar] [CrossRef]
- Gerginova, M.; Manasiev, J.; Yemendzhiev, H.; Terziyska, A.; Peneva, N.; Alexieva, Z. Biodegradation of Phenol by Antarctic Strains of Aspergillus fumigatus. Z. Für Nat. 2013, 68, 384. [Google Scholar] [CrossRef]
- Schlosser, D.; Fahr, K.; Karl, W.; Wetzstein, H.-G. Hydroxylated Metabolites of 2,4-Dichlorophenol Imply a Fenton-Type Reaction in Gloeophyllum striatum. Appl. Environ. Microbiol. 2000, 66, 3010–3015. [Google Scholar] [CrossRef] [Green Version]
- Copley, S.D. Evolution of a metabolic pathway for degradation of a toxic xenobiotic: The patchwork approach. Trends Biochem. Sci. 2000, 25, 261–265. [Google Scholar] [CrossRef]
- Methatham, T.; Lu, M.-C.; Ratanatamskul, C. Removal of 2,4-dichlorophenol as herbicide’s by-product by Fenton’s reagent combined with an electrochemical system. Desalination Water Treat. 2011, 32, 42–48. [Google Scholar] [CrossRef]
- Cho, N.-S.; Jarosz-Wilkolazka, A.; Leonowicz, A.; Oga, S. Removal of Chlorophenols by Fungal Laccase in the Presence of Aromatic Alcohols. J. Fac. Agric. Kyushu Univ. 2007, 52, 23–27. [Google Scholar] [CrossRef]
- Sutton, H.C.; Vile, G.F.; Winterbourn, C.C. Radical driven fenton reactions—Evidence from paraquat radical studies for production of tetravalent iron in the presence and absence of ethylenediaminetetraacetic acid. Arch. Biochem. Biophys. 1987, 256, 462–471. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p-chlorophenol | 2,4-chlorophenol | 2,4,6-chlorophenol | pentachlorophenol | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yield | Yield | Yield | Yield | |||||||||||||
| Experimental Treatments | Rate Constant # | Max.Deg. (%) | T | Max.Deg. 48 h (%) | Rate Constant # | Max.Deg. (%) | T | Max.Deg. 48 h (%) | Rate Constant # | Max.Deg. (%) | T | Max.Deg. 48 h (%) | Rate Constant # | Max.Deg. (%) | T | Max.Deg. 48 h (%) |
| IRC/Fe+3/H2O2 | 5.425 ± 0.082 a,(g) | 98.67 ± 1.247 | 7 | 99.6 ± 0.497 | 5.101 ± 0.052 b,c,d,(g) | 89.32 ± 0.472 | 7 | 99.32 ± 0.943 | 4.704 ± 0.009 e,(g) | 78.13 ± 0.262 | 7 | 99.93 ± 0.094 | 1.74 ± 0.013 f,(g) | 62.0 ± 1.633 | 16 | 64.53 ± 0.45 |
| EDTA/Fe+2/H2O2 | 4.996 ± 0.13 a,(h,i,j) | 70 ± 4.082 | 7 | 87.5 ± 0.408 | 3.582 ± 0.098 b,c,d,(h,l) | 73.32 ± 0.34 | 16 | 79.97 ± 0.126 | 3.459 ± 0.011 e,(i,m) | 30.16 ± 0.125 | 7 | 66.96 ± 2.176 | 1.191 ± 0.075 f,(j,l,m) | 21.66 ± 1.247 | 16 | 24.7 ± 0.356 |
| DTPA/Fe+2/H2O2 | 2.603 ± 0.479 a,(n,o,p) | 15 ± 1.633 | 16 | 24.87 ± 0.66 | 1.8 ± 0.17 b,c,d,(n,q,r) | 6.82 ± 0.236 | 7 | 12.32 ± 0.395 | 0.994 ± 0.103 e,(o,q) | 8.1 ± 0.294 | 7 | 12.33 ± 1.886 | 0.801 ± 0.104 f,(p,r) | 8.16 ± 0.236 | 30 | 8.26 ± 0.377 |
| Fe+2/H2O2 | 2.107 ± 0.152 a,(s) | 14.5 ± 0.408 | 30 | 15.25 ± 0.874 | 0.287 ± 0.239 c,(s) | 6.6 ± 0.082 | 16 | 8.42 ± 0.171 | 0.467 ± 0.013 e,(s) | 1.48 ± 0.062 | 7 | 6.36 ± 0.579 | 0.362 ± 0.078 f,(s) | 2.66 ± 0.492 | 30 | 2.66 ± 0.624 |
| Fe+3/H2O2 | 0.913 ± 0.043 a,(t,u,v) | 6.6 ± 0.432 | 16 | 8.0 ± 0.816 | 0.505 ± 0.099 d,(t) | 1.97 ± 0.126 | 7 | 3.97 ± 0.519 | 0.004 ± 0.003 e,(u) | 0.0 ± 0.005 | 7 | 0.35 ± 0.319 | 0.0 ± 0.0 f,(v) | 0.0 ± 0.0 | n.d. | 0.0 ± 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oses-Pedraza, R.; Bonnail, E.; Lavin, P.; Purcarea, C.; Alarcón, M.; Irribarren-Riquelme, D.; Rodríguez, J. Fenton-Mediated Chlorophenol Degradation by Iron-Reducing Compounds Isolated from Endophytic Fungi in Atacama Puna Plateau Lecanicillium ATA01. Fermentation 2022, 8, 147. https://doi.org/10.3390/fermentation8040147
Oses-Pedraza R, Bonnail E, Lavin P, Purcarea C, Alarcón M, Irribarren-Riquelme D, Rodríguez J. Fenton-Mediated Chlorophenol Degradation by Iron-Reducing Compounds Isolated from Endophytic Fungi in Atacama Puna Plateau Lecanicillium ATA01. Fermentation. 2022; 8(4):147. https://doi.org/10.3390/fermentation8040147
Chicago/Turabian StyleOses-Pedraza, Rómulo, Estefanía Bonnail, Paris Lavin, Cristina Purcarea, Manuel Alarcón, Danae Irribarren-Riquelme, and Jaime Rodríguez. 2022. "Fenton-Mediated Chlorophenol Degradation by Iron-Reducing Compounds Isolated from Endophytic Fungi in Atacama Puna Plateau Lecanicillium ATA01" Fermentation 8, no. 4: 147. https://doi.org/10.3390/fermentation8040147
APA StyleOses-Pedraza, R., Bonnail, E., Lavin, P., Purcarea, C., Alarcón, M., Irribarren-Riquelme, D., & Rodríguez, J. (2022). Fenton-Mediated Chlorophenol Degradation by Iron-Reducing Compounds Isolated from Endophytic Fungi in Atacama Puna Plateau Lecanicillium ATA01. Fermentation, 8(4), 147. https://doi.org/10.3390/fermentation8040147