
Novel Developments on Stimuli-Responsive Probiotic Encapsulates: From Smart Hydrogels to Nanostructured Platforms

 ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
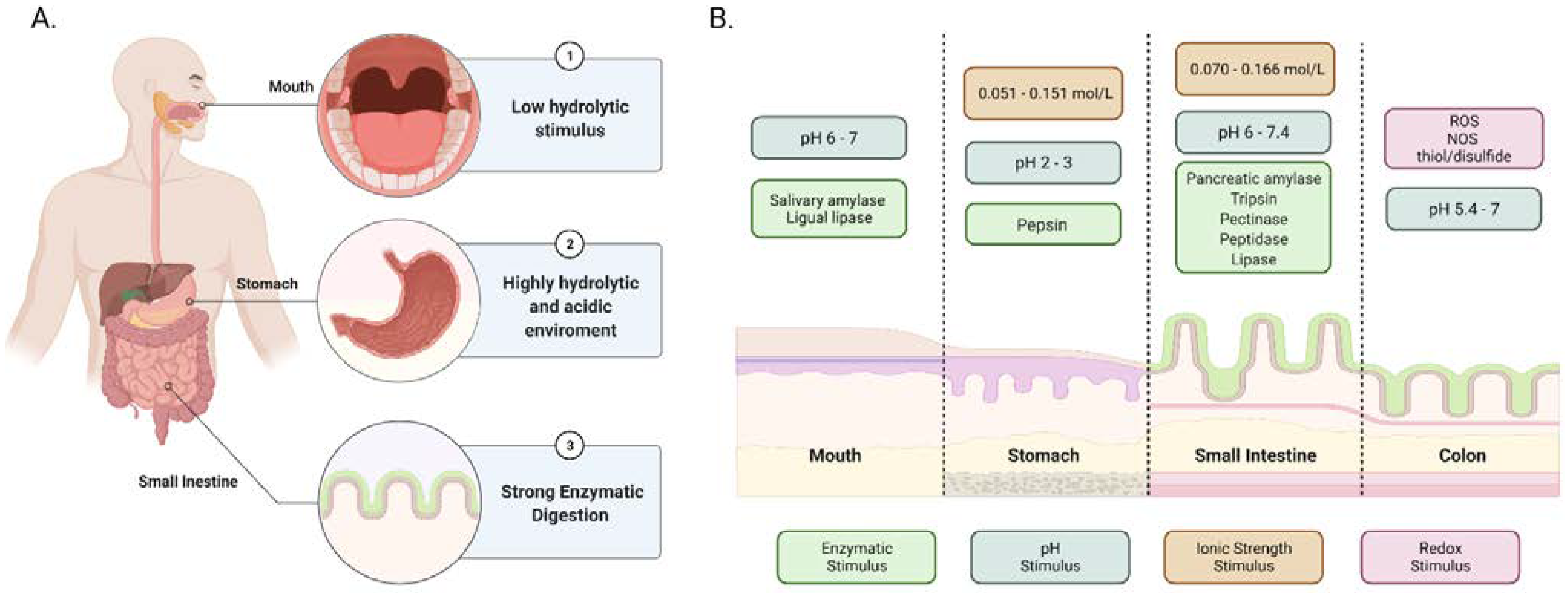
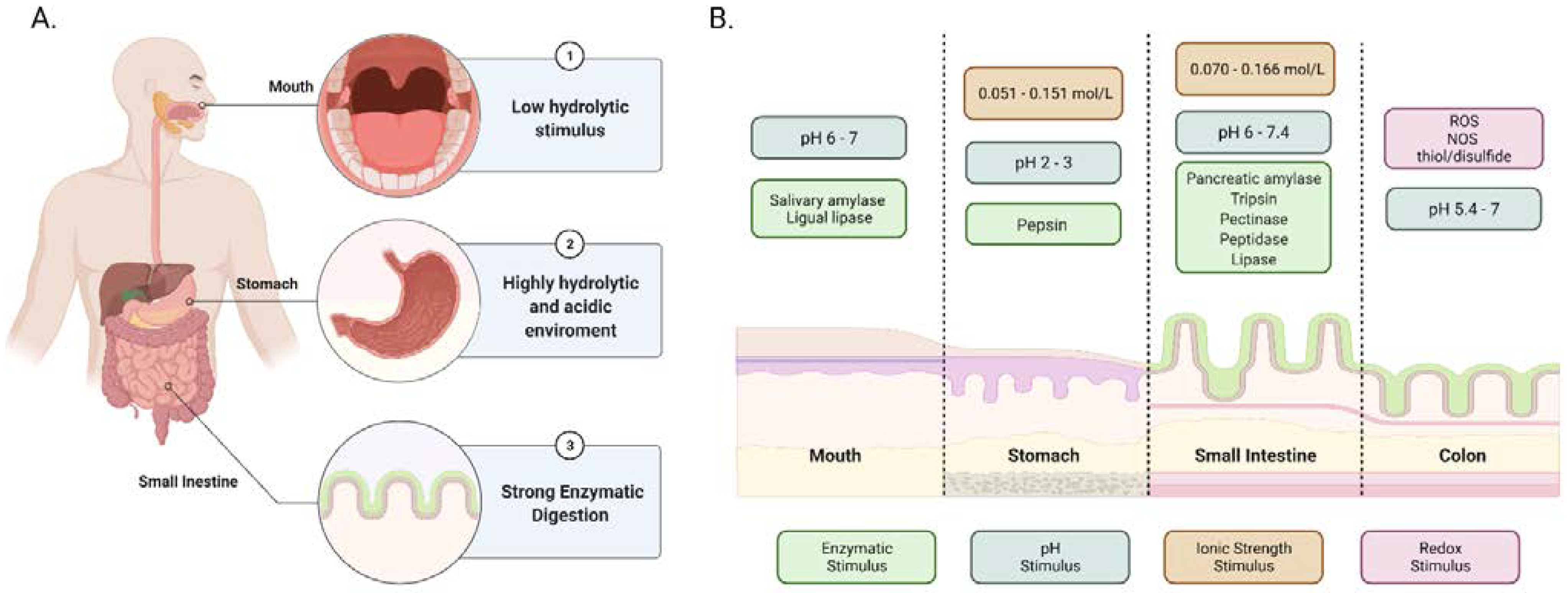
2. Gastrointestinal Tract (GIT) Stimuli as Tool for Design
2.1. Enzymatic Degradation and pH Changes
2.2. Ionic Strength and Redox Potential
3. Polymeric Platforms and Delivery Systems of Probiotics
3.1. Polymeric Materials in Probiotic Encapsulation
3.2. Hydrogels
3.3. Microencapsulates
3.4. Nanostructured Platforms
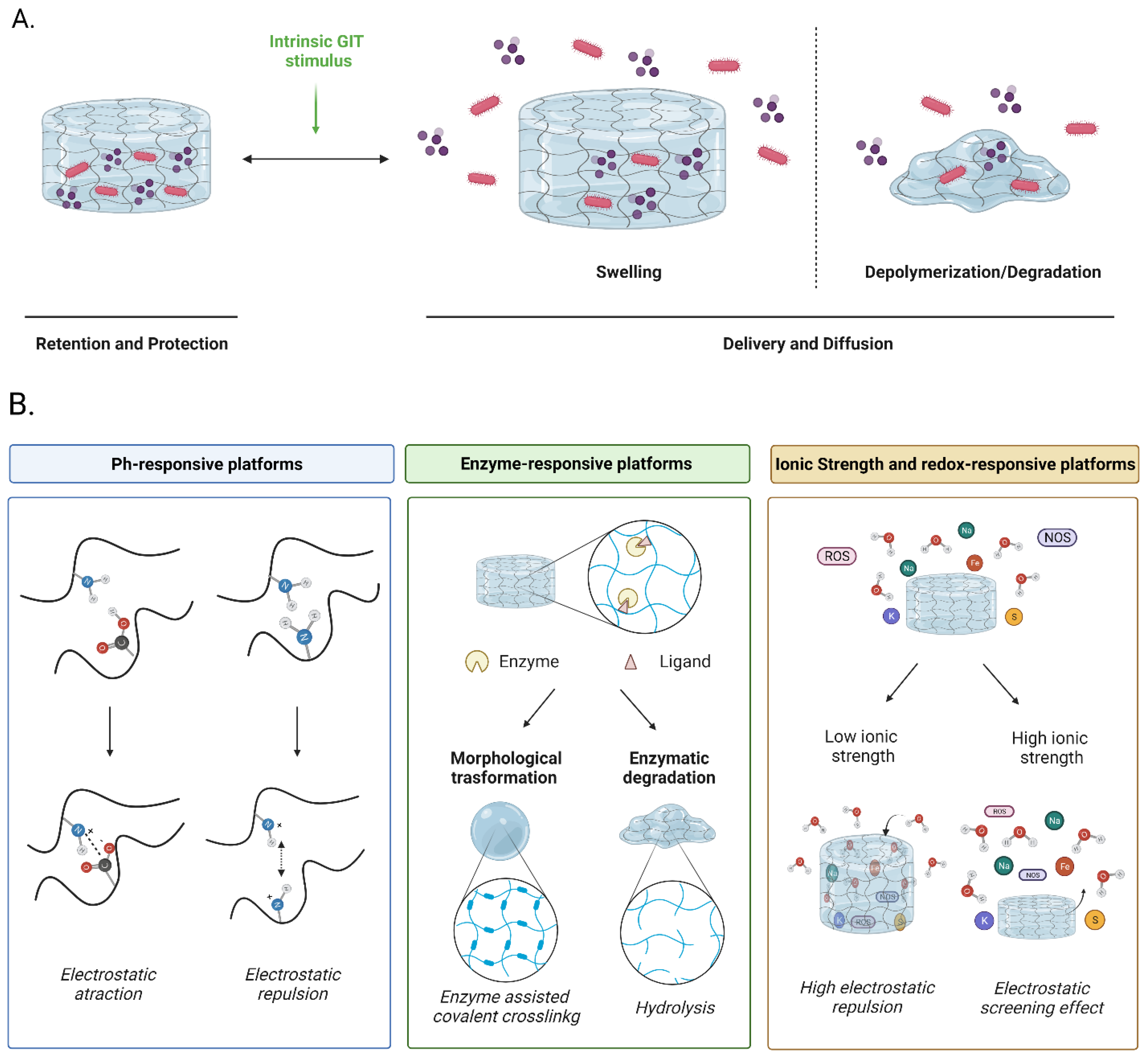
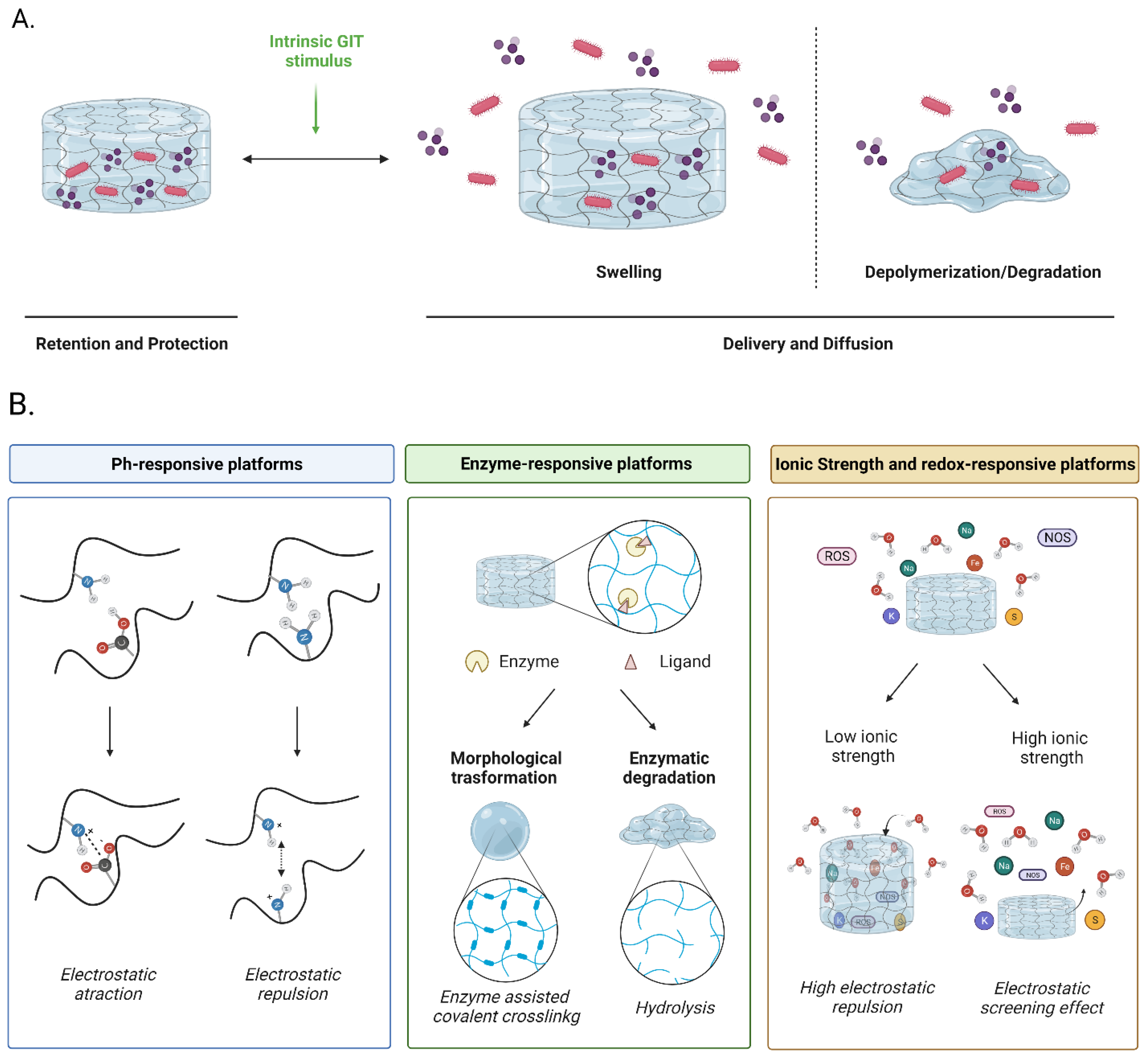
4. Improving Functionalities of Polymeric Matrices for Probiotic Encapsulation
4.1. pH-Responsive Matrices
4.2. Enzyme-Responsive Matrices
4.3. Ionic Strength and Redox-Responsive Matrices
5. Physiological Barriers to Overcome
5.1. Composition of the Gut Microbiome
5.2. Colonization Resistance
5.3. Mucosal Adhesion in Probiotics Colonization
5.4. Prebiotics Co-Immobilization as an Alternative to Improve Probiotics Colonization
6. Future Directions for Probiotic Encapsulation and Clinical Practice
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ranjha, M.M.A.N.; Shafique, B.; Batool, M.; Kowalczewski, P.; Shehzad, Q.; Usman, M.; Manzoor, M.F.; Zahra, S.M.; Yaqub, S.; Aadil, R.M. Nutritional and Health Potential of Probiotics: A Review. Appl. Sci. 2021, 11, 11204. [Google Scholar] [CrossRef]
- Quigley, E.M. Prebiotics and Probiotics in Digestive Health. Clin. Gastroenterol. Hepatol. 2018, 17, 333–344. [Google Scholar] [CrossRef]
- La Fata, G.; Weber, P.; Mohajeri, M.H. Probiotics and the Gut Immune System: Indirect Regulation. Probiot. Antimicrob. Proteins 2018, 10, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Oberoi, K.; Tolun, A.; Altintas, Z.; Sharma, S. Effect of Alginate-Microencapsulated Hydrogels on the Survival of Lactobacillus rhamnosus under Simulated Gastrointestinal Conditions. Foods 2021, 10, 1999. [Google Scholar] [CrossRef]
- Mattila-Sandholm, T.; Myllärinen, P.; Crittenden, R.; Mogensen, G.; Fondén, R.; Saarela, M. Technological challenges for future probiotic foods. Int. Dairy J. 2002, 12, 173–182. [Google Scholar] [CrossRef]
- Butel, M.-J. Probiotics, gut microbiota and health. Med. Mal. Infect. 2014, 44, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Vitetta, L.; Briskey, D.; Alford, H.; Hall, S.; Coulson, S. Probiotics, prebiotics and the gastrointestinal tract in health and disease. Inflammopharmacology 2014, 22, 135–154. [Google Scholar] [CrossRef]
- Chugh, B.; Kamal-Eldin, A. Bioactive compounds produced by probiotics in food products. Curr. Opin. Food Sci. 2020, 32, 76–82. [Google Scholar] [CrossRef]
- Tan-Lim, C.S.C.; Esteban-Ipac, N.A.R. Probiotics as treatment for food allergies among pediatric patients: A meta-analysis. World Allergy Organ. J. 2018, 11, 25. [Google Scholar] [CrossRef]
- Maragkoudakis, P.A.; Zoumpopoulou, G.; Miaris, C.; Kalantzopoulos, G.; Pot, B.; Tsakalidou, E. Probiotic potential of Lactobacillus strains isolated from dairy products. Int. Dairy J. 2006, 16, 189–199. [Google Scholar] [CrossRef]
- Rolim, F.R.; Neto, O.C.F.; Oliveira, M.E.G.; Oliveira, C.J.; Queiroga, R.C. Cheeses as food matrixes for probiotics: In vitro and in vivo tests. Trends Food Sci. Technol. 2020, 100, 138–154. [Google Scholar] [CrossRef]
- Soares, M.B.; Martinez, R.C.; Pereira, E.P.; Balthazar, C.F.; Cruz, A.G.; Ranadheera, C.S.; Sant’Ana, A.S. The resistance of Bacillus, Bifidobacterium, and Lactobacillus strains with claimed probiotic properties in different food matrices exposed to simulated gastrointestinal tract conditions. Food Res. Int. 2019, 125, 108542. [Google Scholar] [CrossRef] [PubMed]
- McClements, D.J. Designing biopolymer microgels to encapsulate, protect and deliver bioactive components: Physicochemical aspects. Adv. Colloid Interface Sci. 2016, 240, 31–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas-Jara, M.J.; Ilabaca, A.; Vega, M.; García, A. Biofilm Forming Lactobacillus: New Challenges for the Development of Probiotics. Microorganisms 2016, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Trush, E.; Poluektova, E.A.; Beniashvilli, A.G.; Shifrin, O.S.; Poluektov, Y.M.; Ivashkin, V.T. The Evolution of Human Probiotics: Challenges and Prospects. Probiot. Antimicrob. Proteins 2020, 12, 1291–1299. [Google Scholar] [CrossRef] [PubMed]
- Huq, T.; Khan, A.; Khan, R.A.; Riedl, B.; Lacroix, M. Encapsulation of Probiotic Bacteria in Biopolymeric System. Crit. Rev. Food Sci. Nutr. 2013, 53, 909–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieterich, W.; Schink, M.; Zopf, Y. Microbiota in the Gastrointestinal Tract. Med. Sci. 2018, 6, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanaro, S.; Chierici, R.; Guerrini, P.; Vigi, V. Intestinal microflora in early infancy: Composition and development. Acta Paediatr. 2007, 92, 48–55. [Google Scholar] [CrossRef]
- Tannock, G.W. Molecular assessment of intestinal microflora. Am. J. Clin. Nutr. 2001, 73, 410s–414s. [Google Scholar] [CrossRef] [Green Version]
- Baral, K.C.; Bajracharya, R.; Lee, S.H.; Han, H.-K. Advancements in the Pharmaceutical Applications of Probiotics: Dosage Forms and Formulation Technology. Int. J. Nanomed. 2021, 16, 7535–7556. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, X.; Xue, C.; Wei, Z. Latest developments in food-grade delivery systems for probiotics: A systematic review. Crit. Rev. Food Sci. Nutr. 2021, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Asgari, S.; Pourjavadi, A.; Licht, T.R.; Boisen, A.; Ajalloueian, F. Polymeric carriers for enhanced delivery of probiotics. Adv. Drug Deliv. Rev. 2020, 161–162, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Yoha, K.S.; Nida, S.; Dutta, S.; Moses, J.A.; Anandharamakrishnan, C. Targeted Delivery of Probiotics: Perspectives on Research and Commercialization. Probiot. Antimicrob. Proteins 2021, 14, 15–48. [Google Scholar] [CrossRef] [PubMed]
- El-Husseiny, H.M.; Mady, E.A.; Hamabe, L.; Abugomaa, A.; Shimada, K.; Yoshida, T.; Tanaka, T.; Yokoi, A.; Elbadawy, M.; Tanaka, R. Smart/stimuli-responsive hydrogels: Cutting-edge platforms for tissue engineering and other biomedical applications. Mater. Today Bio 2021, 13, 100186. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Xie, J.; Du, H.; McClements, D.J.; Xiao, H.; Li, L. Progress in microencapsulation of probiotics: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 857–874. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Lu, Y.; Xie, J.; Fei, Y.; Zheng, G.; Wang, Z.; Liu, J.; Lv, L.; Ling, Z.; Berglund, B.; et al. Probiotic Gastrointestinal Transit and Colonization After Oral Administration: A Long Journey. Front. Cell. Infect. Microbiol. 2021, 11, 609722. [Google Scholar] [CrossRef]
- Yao, M.; Li, B.; Ye, H.; Huang, W.; Luo, Q.; Xiao, H.; McClements, D.J.; Li, L. Enhanced viability of probiotics (Pediococcus pentosaceus Li05) by encapsulation in microgels doped with inorganic nanoparticles. Food Hydrocoll. 2018, 83, 246–252. [Google Scholar] [CrossRef]
- Patricia, J.J.; Dhamoon, A.S. Physiology, Digestion, StatPearls. September 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK544242/ (accessed on 24 January 2022).
- Gachons, C.P.D.; Breslin, P.A.S. Salivary Amylase: Digestion and Metabolic Syndrome. Curr. Diabetes Rep. 2016, 16, 102. [Google Scholar] [CrossRef]
- Patel, G.; Misra, A. Oral Delivery of Proteins and Peptides: Concepts and Applications. In Challenges in Delivery of Therapeutic Genomics and Proteomics; Elsevier: Amsterdam, The Netherlands, 2011; pp. 481–529. [Google Scholar] [CrossRef]
- Bertoni, S.; Machness, A.; Tiboni, M.; Bártolo, R.; Santos, H.A. Reactive oxygen species responsive nanoplatforms as smart drug delivery systems for gastrointestinal tract targeting. Biopolymers 2019, 111, e23336. [Google Scholar] [CrossRef]
- Shubakov, A.; Mikhailova, E.; Prosheva, V.; Belyy, V. Swelling and Degradation of Calcium-Pectic Gel Particles Made of Pectins of Silene vulgaris and Lemna minor Callus Cultures at Different Concentrations of Pectinase in an Artificial Colon Environment. Int. J. Biomed. 2018, 8, 60–64. [Google Scholar] [CrossRef]
- Zhu, W.; Han, C.; Dong, Y.; Jian, B. Enzyme-responsive mechanism based on multi-walled carbon nanotubes and pectin complex tablets for oral colon-specific drug delivery system. J. Radioanal. Nucl. Chem. Artic. 2019, 320, 503–512. [Google Scholar] [CrossRef]
- Kim, Y.S.; Erickson, R.H. Role of Peptidases of the Human Small Intestine in Protein Digestion. Gastroenterology 1985, 88, 1071–1073. [Google Scholar] [CrossRef]
- Pirahanchi, Y.; Sharma, S. Biochemistry, Lipase, StatPearls, July 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK537346/ (accessed on 24 January 2022).
- Circu, M.L.; Aw, T.Y. Redox biology of the intestine. Free Radic. Res. 2011, 45, 1245–1266. [Google Scholar] [CrossRef] [Green Version]
- Arnaut, L. Elementary reactions in solution. In Chemical Kinetics; Elsevier: Amsterdam, The Netherlands, 2021; pp. 263–293. [Google Scholar] [CrossRef]
- Qi, X.; Simsek, S.; Chen, B.; Rao, J. Alginate-based double-network hydrogel improves the viability of encapsulated probiotics during simulated sequential gastrointestinal digestion: Effect of biopolymer type and concentrations. Int. J. Biol. Macromol. 2020, 165, 1675–1685. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Han, Z.; Chen, C.; Li, Z.; Yu, S.; Qu, Y.; Zeng, R. Novel probiotic-bound oxidized Bletilla striata polysaccharide-chitosan composite hydrogel. Mater. Sci. Eng. C 2020, 117, 111265. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Cai, Z.; Wang, F.; Hong, L.; Deng, L.; Zhong, J.; Wang, Z.; Cui, W. Colon-Targeted Adhesive Hydrogel Microsphere for Regulation of Gut Immunity and Flora. Adv. Sci. 2021, 8, 2101619. [Google Scholar] [CrossRef]
- Bearat, H.; Vernon, B. Environmentally responsive injectable materials. In Injectable Biomaterials; Elsevier: Amsterdam, The Netherlands, 2011; pp. 263–297. [Google Scholar] [CrossRef]
- Alkekhia, D.; Hammond, P.T.; Shukla, A. Layer-by-Layer Biomaterials for Drug Delivery. Annu. Rev. Biomed. Eng. 2020, 22, 1–24. [Google Scholar] [CrossRef]
- Pei, J.; Xiong, L.; Bao, P.; Chu, M.; Yan, P.; Guo, X. Secondary Structural Transformation of Bovine Lactoferricin Affects Its Antibacterial Activity. Probiot. Antimicrob. Proteins 2020, 13, 873–884. [Google Scholar] [CrossRef]
- Rattanaburi, P.; Charoenrat, N.; Pongtharangkul, T.; Suphantharika, M.; Wongkongkatep, J. Hydroxypropyl methylcellulose enhances the stability of o/w Pickering emulsions stabilized with chitosan and the whole cells of Lactococcus lactis IO-1. Food Res. Int. 2018, 116, 559–565. [Google Scholar] [CrossRef]
- Taniguchi, M.; Nambu, M.; Katakura, Y.; Yamasaki-Yashiki, S. Adhesion mechanisms of Bifidobacterium animalis subsp. lactis JCM 10602 to dietary fiber. Biosci. Microbiota Food Health 2021, 40, 59–64. [Google Scholar] [CrossRef]
- Lindahl, A.; Ungell, A.-L.; Knutson, L.; Lennernäs, H. Characterization of Fluids from the Stomach and Proximal Jejunum in Men and Women. Pharm. Res. 1997, 14, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Million, M.; Raoult, D. Linking gut redox to human microbiome. Hum. Microbiome J. 2018, 10, 27–32. [Google Scholar] [CrossRef]
- Vincent, S.G.T.; Jennerjahn, T.; Ramasamy, K. Environmental variables and factors regulating microbial structure and functions. In Microbial Communities in Coastal Sediments; Elsevier: Amsterdam, The Netherlands, 2021; pp. 79–117. [Google Scholar] [CrossRef]
- Kwiecień, I.; Kwiecień, M. Application of Polysaccharide-Based Hydrogels as Probiotic Delivery Systems. Gels 2018, 4, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abad, I.; Conesa, C.; Sánchez, L. Development of Encapsulation Strategies and Composite Edible Films to Maintain Lactoferrin Bioactivity: A Review. Materials 2021, 14, 7358. [Google Scholar] [CrossRef]
- Gheorghita, R.; Anchidin-Norocel, L.; Filip, R.; Dimian, M.; Covasa, M. Applications of Biopolymers for Drugs and Probiotics Delivery. Polymers 2021, 13, 2729. [Google Scholar] [CrossRef] [PubMed]
- Saboktakin, M.R.; Tabatabaei, R.M. Supramolecular hydrogels as drug delivery systems. Int. J. Biol. Macromol. 2015, 75, 426–436. [Google Scholar] [CrossRef]
- Municoy, S.; Álvarez Echazú, M.I.; Antezana, P.E.; Galdopórpora, J.M.; Olivetti, C.; Mebert, A.M.; Foglia, M.L.; Tuttolomondo, M.V.; Alvarez, G.S.; Hardy, J.G.; et al. Stimuli-Responsive Materials for Tissue Engineering and Drug Delivery. Int. J. Mol. Sci. 2020, 21, 4724. [Google Scholar] [CrossRef]
- Pinteala, M.; Abadie, M.J.M.; Rusu, R.D. Smart Supra-and Macro-Molecular Tools for Biomedical Applications. Materials 2020, 13, 3343. [Google Scholar] [CrossRef]
- Safi, M.A.E. Materials and Techniques for Microencapsulation of Probiotics. Biosci. Biotechnol. Res. Commun. 2021, 14, 922–928. [Google Scholar] [CrossRef]
- da Silva, B.V.; Barreira, J.C.; Oliveira, M.B.P. Natural phytochemicals and probiotics as bioactive ingredients for functional foods: Extraction, biochemistry and protected-delivery technologies. Trends Food Sci. Technol. 2016, 50, 144–158. [Google Scholar] [CrossRef] [Green Version]
- Survival of Lactobacillus Acidophilus as Probiotic Bacteria using Chitosan Nanoparticles. Int. J. Eng. 2017, 30, 456–463. [CrossRef]
- Allan-Wojtas, P.; Hansen, L.T.; Paulson, A. Microstructural studies of probiotic bacteria-loaded alginate microcapsules using standard electron microscopy techniques and anhydrous fixation. LWT 2008, 41, 101–108. [Google Scholar] [CrossRef]
- Shori, A.B. Microencapsulation Improved Probiotics Survival During Gastric Transit. HAYATI J. Biosci. 2017, 24, 1–5. [Google Scholar] [CrossRef]
- Hansen, L.T.; Allan-Wojtas, P.; Jin, Y.-L.; Paulson, A. Survival of Ca-alginate microencapsulated Bifidobacterium spp. in milk and simulated gastrointestinal conditions. Food Microbiol. 2002, 19, 35–45. [Google Scholar] [CrossRef]
- Etchepare, M.D.A.; Barin, J.S.; Cichoski, A.J.; Jacob-Lopes, E.; Wagner, R.; Fries, L.L.M.; De Menezes, C.R. Microencapsulation of probiotics using sodium alginate. Ciência Rural 2015, 45, 1319–1326. [Google Scholar] [CrossRef] [Green Version]
- Mathews, S. Microencapsulation of Probiotics by Calcium Alginate and Gelatin and Evaluation of its Survival in Simulated Human Gastro-Intestinal Condition. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2080–2087. [Google Scholar] [CrossRef] [Green Version]
- Khosravi Zanjani, M.A.; Tarzi, B.G.; Sharifan, A.; Mohammadi, N. Microencapsulation of probiotics by calcium alginate-gelatinized starch with chitosan coating and evaluation of survival in simulated human gastro-intestinal condition. Iran. J. Pharm. Res. 2014, 13, 843–852. [Google Scholar] [CrossRef]
- Krasaekoopt, W.; Bhandari, B.; Deeth, H. The influence of coating materials on some properties of alginate beads and survivability of microencapsulated probiotic bacteria. Int. Dairy J. 2004, 14, 737–743. [Google Scholar] [CrossRef]
- Shu, X.; Zhu, K. The release behavior of brilliant blue from calcium–alginate gel beads coated by chitosan: The preparation method effect. Eur. J. Pharm. Biopharm. 2002, 53, 193–201. [Google Scholar] [CrossRef]
- Lee, J.S.; Cha, D.S.; Park, H.J. Survival of Freeze-Dried Lactobacillus bulgaricus KFRI 673 in Chitosan-Coated Calcium Alginate Microparticles. J. Agric. Food Chem. 2004, 52, 7300–7305. [Google Scholar] [CrossRef]
- Koo, S.M.; Cho, Y.H.; Huh, C.S.; Baek, Y.J.; Park, J.Y. Improvement of the Stability of Lactobacillus casei YIT 9018 by Microencapsulation Using Alginate and Chitosan. J. Microbiol. Biotechnol. 2001, 11, 376–383. [Google Scholar]
- Călinoiu, L.-F.; Ştefănescu, B.E.; Pop, I.D.; Muntean, L.; Vodnar, D.C. Chitosan Coating Applications in Probiotic Microencapsulation. Coatings 2019, 9, 194. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.A.S.; Machado, M.T.C. Coated alginate–chitosan particles to improve the stability of probiotic yeast. Int. J. Food Sci. Technol. 2020, 56, 2122–2131. [Google Scholar] [CrossRef]
- Albadran, H.A.; Monteagudo-Mera, A.; Khutoryanskiy, V.V.; Charalampopoulos, D. Development of chitosan-coated agar-gelatin particles for probiotic delivery and targeted release in the gastrointestinal tract. Appl. Microbiol. Biotechnol. 2020, 104, 5749–5757. [Google Scholar] [CrossRef]
- Annan, N.; Borza, A.; Hansen, L.T. Encapsulation in alginate-coated gelatin microspheres improves survival of the probiotic Bifidobacterium adolescentis 15703T during exposure to simulated gastro-intestinal conditions. Food Res. Int. 2008, 41, 184–193. [Google Scholar] [CrossRef]
- Liu, J.; Liu, F.; Ren, T.; Wang, J.; Yang, M.; Yao, Y.; Chen, H. Fabrication of fish gelatin/sodium alginate double network gels for encapsulation of probiotics. J. Sci. Food Agric. 2021, 101, 4398–4408. [Google Scholar] [CrossRef]
- Ramos, P.E.; Cerqueira, M.A.; Teixeira, J.A.; Vicente, A.A. Physiological protection of probiotic microcapsules by coatings. Crit. Rev. Food Sci. Nutr. 2018, 58, 1864–1877. [Google Scholar] [CrossRef]
- Hlaing, S.P.; Kim, J.; Lee, J.; Kwak, D.; Kim, H.; Yoo, J.-W. Enhanced Viability of Probiotics Against Gastric Acid by One-Step Coating Process with Poly-L-Lysine: In Vitro and In Vivo Evaluation. Pharmaceutics 2020, 12, 662. [Google Scholar] [CrossRef]
- Misra, S.; Pandey, P.; Mishra, H.N. Novel approaches for co-encapsulation of probiotic bacteria with bioactive compounds, their health benefits and functional food product development: A review. Trends Food Sci. Technol. 2021, 109, 340–351. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Z.; Bai, L.; Deng, J.; Zhou, Q. Biomaterial-based encapsulated probiotics for biomedical applications: Current status and future perspectives. Mater. Des. 2021, 210, 110018. [Google Scholar] [CrossRef]
- Xiao, Y.; Lu, C.; Liu, Y.; Kong, L.; Bai, H.; Mu, H.; Li, Z.; Geng, H.; Duan, J. Encapsulation of Lactobacillus rhamnosus in Hyaluronic Acid-Based Hydrogel for Pathogen-Targeted Delivery to Ameliorate Enteritis. ACS Appl. Mater. Interfaces 2020, 12, 36967–36977. [Google Scholar] [CrossRef] [PubMed]
- Mosqueda, I.S.; Bousquets, A.L.; Montiel-Sosa, J.F.; Corona, L.; Álvarez, Z.G.; Gochi, L.C. Encapsulation of Lactobacillus plantarum ATCC 8014 and Pediococcus acidilactici ATCC 8042 in a freeze-dried alginate-gum arabic system and its in vitro testing under gastrointestinal conditions. J. Microencapsul. 2019, 36, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, Y.; Polk, D.B.; Tomasula, P.M.; Yan, F.; Liu, L. Preserving viability of Lactobacillus rhamnosus GG in vitro and in vivo by a new encapsulation system. J. Control. Release 2016, 230, 79–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.; Cai, Y.; Zhi, Z.; Guo, Q.; Mao, L.; Gao, Y.; Yuan, F.; Van der Meeren, P. Assembly of propylene glycol alginate/β-lactoglobulin composite hydrogels induced by ethanol for co-delivery of probiotics and curcumin. Carbohydr. Polym. 2020, 254, 117446. [Google Scholar] [CrossRef]
- Dafe, A.; Etemadi, H.; Dilmaghani, A.; Mahdavinia, G.R. Investigation of pectin/starch hydrogel as a carrier for oral delivery of probiotic bacteria. Int. J. Biol. Macromol. 2017, 97, 536–543. [Google Scholar] [CrossRef]
- Sharpe, L.A.; Daily, A.M.; Horava, S.D.; A Peppas, N. Therapeutic applications of hydrogels in oral drug delivery. Expert Opin. Drug Deliv. 2014, 11, 901–915. [Google Scholar] [CrossRef] [Green Version]
- Patarroyo, J.L.; Florez-Rojas, J.S.; Pradilla, D.; Valderrama-Rincón, J.D.; Cruz, J.C.; Reyes, L.H. Formulation and Characterization of Gelatin-Based Hydrogels for the Encapsulation of Kluyveromyces lactis—Applications in Packed-Bed Reactors and Probiotics Delivery in Humans. Polymers 2020, 12, 1287. [Google Scholar] [CrossRef]
- Ghibaudo, F.; Gerbino, E.; Orto, V.C.D.; Gomez-Zavaglia, A. Pectin-iron capsules: Novel system to stabilise and deliver lactic acid bacteria. J. Funct. Foods 2017, 39, 299–305. [Google Scholar] [CrossRef]
- Falco, C.Y.; Falkman, P.; Risbo, J.; Cárdenas, M.; Medronho, B. Chitosan-dextran sulfate hydrogels as a potential carrier for probiotics. Carbohydr. Polym. 2017, 172, 175–183. [Google Scholar] [CrossRef]
- Patarroyo, J.; Fonseca, E.; Cifuentes, J.; Salcedo, F.; Cruz, J.; Reyes, L. Gelatin-Graphene Oxide Nanocomposite Hydrogels for Kluyveromyces lactis Encapsulation: Potential Applications in Probiotics and Bioreactor Packings. Biomolecules 2021, 11, 922. [Google Scholar] [CrossRef]
- Badeau, B.A.; Deforest, C.A. Programming Stimuli-Responsive Behavior into Biomaterials. Annu. Rev. Biomed. Eng. 2019, 21, 241–265. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Samanta, P.; Dhara, D. Temperature, pH and redox responsive cellulose based hydrogels for protein delivery. Int. J. Biol. Macromol. 2016, 87, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Noirbent, G.; Dumur, F. Photoinitiators of polymerization with reduced environmental impact: Nature as an unlimited and renewable source of dyes. Eur. Polym. J. 2020, 142, 110109. [Google Scholar] [CrossRef]
- Ahmed, E.M. Hydrogel: Preparation, characterization, and applications: A review. J. Adv. Res. 2015, 6, 105–121. [Google Scholar] [CrossRef] [Green Version]
- Samal, S.K.; Dash, M.; Dubruel, P.; Van Vlierberghe, S. Smart polymer hydrogels: Properties, synthesis and applications. In Smart Polymers and their Applications, 1st ed.; Aguilar, M.R., Román, J.S., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2014; pp. 237–270. ISBN 978-085-709-702-6. [Google Scholar]
- Liu, L.; Yao, W.; Rao, Y.; Lu, X.; Gao, J. pH-Responsive carriers for oral drug delivery: Challenges and opportunities of current platforms. Drug Deliv. 2017, 24, 569–581. [Google Scholar] [CrossRef] [Green Version]
- Peppas, N.A.; Huang, Y. Nanoscale technology of mucoadhesive interactions. Adv. Drug Deliv. Rev. 2004, 56, 1675–1687. [Google Scholar] [CrossRef]
- Tang, Y.; Heaysman, C.L.; Willis, S.; Lewis, A.L. Physical hydrogels with self-assembled nanostructures as drug delivery systems. Expert Opin. Drug Deliv. 2011, 8, 1141–1159. [Google Scholar] [CrossRef]
- Sun, Q.; Wicker, L. Hydrogel Encapsulation of Lactobacillus casei by Block Charge Modified Pectin and Improved Gastric and Storage Stability. Foods 2021, 10, 1337. [Google Scholar] [CrossRef]
- Yeung, T.W.; Üçok, E.F.; Tiani, K.A.; McClements, D.J.; Sela, D.A. Microencapsulation in Alginate and Chitosan Microgels to Enhance Viability of Bifidobacterium longum for Oral Delivery. Front. Microbiol. 2016, 7, 494. [Google Scholar] [CrossRef] [Green Version]
- Sohail, A.; Turner, M.; Coombes, A.G.; E Bostrom, T.; Bhandari, B. Survivability of probiotics encapsulated in alginate gel microbeads using a novel impinging aerosols method. Int. J. Food Microbiol. 2011, 145, 162–168. [Google Scholar] [CrossRef]
- Yeung, T.W.; Arroyo-Maya, I.J.; McClements, D.J.; Sela, D.A. Microencapsulation of probiotics in hydrogel particles: Enhancing Lactococcus lactis subsp. cremoris LM0230 viability using calcium alginate beads. Food Funct. 2015, 7, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Feng, C.; Li, J.; Mu, Y.; Liu, Y.; Kong, M.; Cheng, X.; Chen, X. Construction of multilayer alginate hydrogel beads for oral delivery of probiotics cells. Int. J. Biol. Macromol. 2017, 105, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Liu, C.; Zhang, H.; Hossen, A.; Sameen, D.E.; Dai, J.; Qin, W.; Liu, Y.; Li, S. In vitro digestion of sodium alginate/pectin co-encapsulated Lactobacillus bulgaricus and its application in yogurt bilayer beads. Int. J. Biol. Macromol. 2021, 193, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Kahieshesfandiari, M.; Nami, Y.; Lornezhad, G.; Kiani, A.; Javanmard, A.; Jaymand, M.; Haghshenas, B. Herbal hydrogel-based encapsulated Enterococcus faecium ABRIINW.N7 improves the resistance of red hybrid tilapia against Streptococcus iniae. J. Appl. Microbiol. 2021, 131, 2516–2527. [Google Scholar] [CrossRef]
- Ming, Z.; Han, L.; Bao, M.; Zhu, H.; Qiang, S.; Xue, S.; Liu, W. Living Bacterial Hydrogels for Accelerated Infected Wound Healing. Adv. Sci. 2021, 8, 2102545. [Google Scholar] [CrossRef]
- Riaz, T.; Iqbal, M.W.; Saeed, M.; Yasmin, I.; Hassanin, H.A.M.; Mahmood, S.; Rehman, A. In vitro survival of Bifidobacterium bifidum microencapsulated in zein-coated alginate hydrogel microbeads. J. Microencapsul. 2019, 36, 192–203. [Google Scholar] [CrossRef]
- Črnivec, I.G.O.; Neresyan, T.; Gatina, Y.; Bučar, V.K.; Skrt, M.; Dogša, I.; Matijašić, B.B.; Kulikova, I.; Lodygin, A.; Ulrih, N.P. Polysaccharide Hydrogels for the Protection of Dairy-Related Microorganisms in Adverse Environmental Conditions. Molecules 2021, 26, 7484. [Google Scholar] [CrossRef]
- Mirmazloum, I.; Ladányi, M.; Omran, M.; Papp, V.; Ronkainen, V.-P.; Pónya, Z.; Papp, I.; Némedi, E.; Kiss, A. Co-encapsulation of probiotic Lactobacillus acidophilus and Reishi medicinal mushroom (Ganoderma lingzhi) extract in moist calcium alginate beads. Int. J. Biol. Macromol. 2021, 192, 461–470. [Google Scholar] [CrossRef]
- Afzaal, M.; Khan, A.U.; Saeed, F.; Arshad, M.S.; Khan, M.A.; Saeed, M.; Maan, A.A.; Khan, M.K.; Ismail, Z.; Ahmed, A.; et al. Survival and stability of free and encapsulated probiotic bacteria under simulated gastrointestinal conditions and in ice cream. Food Sci. Nutr. 2020, 8, 1649–1656. [Google Scholar] [CrossRef] [Green Version]
- Tarifa, M.C.; Piqueras, C.M.; Genovese, D.B.; Brugnoni, L.I. Microencapsulation of Lactobacillus casei and Lactobacillus rhamnosus in pectin and pectin-inulin microgel particles: Effect on bacterial survival under storage conditions. Int. J. Biol. Macromol. 2021, 179, 457–465. [Google Scholar] [CrossRef]
- Praepanitchai, O.-A.; Noomhorm, A.; Anal, A.K. Survival and Behavior of Encapsulated Probiotics (Lactobacillus plantarum) in Calcium-Alginate-Soy Protein Isolate-Based Hydrogel Beads in Different Processing Conditions (pH and Temperature) and in Pasteurized Mango Juice. BioMed Res. Int. 2019, 2019, 9768152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, M.T.; Tzortzis, G.; Charalampopoulos, D.; Khutoryanskiy, V.V. Microencapsulation of a synbiotic into PLGA/alginate multiparticulate gels. Int. J. Pharm. 2014, 466, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultana, K.; Godward, G.; Reynolds, N.; Arumugaswamy, R.; Peiris, P.; Kailasapathy, K. Encapsulation of probiotic bacteria with alginate–starch and evaluation of survival in simulated gastrointestinal conditions and in yoghurt. Int. J. Food Microbiol. 2000, 62, 47–55. [Google Scholar] [CrossRef]
- Holkem, A.T.; Favaro-Trindade, C.S. Potential of solid lipid microparticles covered by the protein-polysaccharide complex for protection of probiotics and proanthocyanidin-rich cinnamon extract. Food Res. Int. 2020, 136, 109520. [Google Scholar] [CrossRef] [PubMed]
- González-Ferrero, C.; Irache, J.; González-Navarro, C. Soybean protein-based microparticles for oral delivery of probiotics with improved stability during storage and gut resistance. Food Chem. 2018, 239, 879–888. [Google Scholar] [CrossRef]
- Harshitha, K.; Kulkarni, P.; Vaghela, R.; Varma, V.K.; Deshpande, D.; Hani, U. Probiotic and Prebiotic-probiotic PEC Microparticles for Sustaining and Enhancing Intestinal Probiotic Growth. Curr. Drug Deliv. 2015, 12, 299–307. [Google Scholar] [CrossRef]
- Haffner, F.B.; Girardon, M.; Fontanay, S.; Canilho, N.; Duval, R.E.; Mierzwa, M.; Etienne, M.; Diab, R.; Pasc, A. Core–shell alginate@silica microparticles encapsulating probiotics. J. Mater. Chem. B 2016, 4, 7929–7935. [Google Scholar] [CrossRef]
- Di Natale, C.; Lagreca, E.; Panzetta, V.; Gallo, M.; Passannanti, F.; Vitale, M.; Fusco, S.; Vecchione, R.; Nigro, R.; Netti, P. Morphological and Rheological Guided Design for the Microencapsulation Process of Lactobacillus paracasei CBA L74 in Calcium Alginate Microspheres. Front. Bioeng. Biotechnol. 2021, 9, 660691. [Google Scholar] [CrossRef]
- Anselmo, A.C.; McHugh, K.J.; Webster, J.; Langer, R.; Jaklenec, A. Layer-by-Layer Encapsulation of Probiotics for Delivery to the Microbiome. Adv. Mater. 2016, 28, 9486–9490. [Google Scholar] [CrossRef] [Green Version]
- Fakhrullin, R.F.; Lvov, Y.M. “Face-Lifting” and “Make-Up” for Microorganisms: Layer-by-Layer Polyelectrolyte Nanocoating. ACS Nano 2012, 6, 4557–4564. [Google Scholar] [CrossRef]
- Tripathi, P.; Beaussart, A.; Alsteens, D.; Dupres, V.; Claes, I.; von Ossowski, I.E.; De Vos, W.M.; Palva, A.; Lebeer, S.; Vanderleyden, J.; et al. Adhesion and Nanomechanics of Pili from the Probiotic Lactobacillus rhamnosus GG. ACS Nano 2013, 7, 3685–3697. [Google Scholar] [CrossRef] [PubMed]
- Pothakamury, U.R.; Barbosa-Cánovas, G.V. Fundamental aspects of controlled release in foods. Trends Food Sci. Technol. 1995, 6, 397–406. [Google Scholar] [CrossRef]
- Chen, M.-J.; Chen, K.-N. Applications of Probiotic Encapsulation in Dairy Products. In Encapsulation and Controlled Release Technologies in Food Systems; Blackwell Publishing: Ames, IA, USA, 2007; pp. 83–112. [Google Scholar] [CrossRef]
- Kailasapathy, K. Encapsulation technologies for functional foods and nutraceutical product development. CAB Rev. Perspect. Agric. Veter. Sci. Nutr. Nat. Resour. 2009, 4. [Google Scholar] [CrossRef]
- de Vos, P.; Faas, M.M.; Spasojevic, M.; Sikkema, J. Encapsulation for preservation of functionality and targeted delivery of bioactive food components. Int. Dairy J. 2010, 20, 292–302. [Google Scholar] [CrossRef]
- Picot, A.; Lacroix, C. Encapsulation of bifidobacteria in whey protein-based microcapsules and survival in simulated gastrointestinal conditions and in yoghurt. Int. Dairy J. 2004, 14, 505–515. [Google Scholar] [CrossRef]
- Heidebach, T.; Först, P.; Kulozik, U. Microencapsulation of probiotic cells by means of rennet-gelation of milk proteins. Food Hydrocoll. 2009, 23, 1670–1677. [Google Scholar] [CrossRef]
- Heidebach, T.; Först, P.; Kulozik, U. Transglutaminase-induced caseinate gelation for the microencapsulation of probiotic cells. Int. Dairy J. 2009, 19, 77–84. [Google Scholar] [CrossRef]
- Li, X.Y.; Chen, X.G.; Cha, D.S.; Park, H.J.; Liu, C. Microencapsulation of a probiotic bacteria with alginate–gelatin and its properties. J. Microencapsul. 2008, 26, 315–324. [Google Scholar] [CrossRef]
- Semyonov, D.; Ramon, O.; Kaplun, Z.; Levin-Brener, L.; Gurevich, N.; Shimoni, E. Microencapsulation of Lactobacillus paracasei by spray freeze drying. Food Res. Int. 2010, 43, 193–202. [Google Scholar] [CrossRef]
- Wang, Z.; Finlay, W.; Peppler, M.; Sweeney, L. Powder formation by atmospheric spray-freeze-drying. Powder Technol. 2006, 170, 45–52. [Google Scholar] [CrossRef]
- Zuidam, N.J.; Shimoni, E. Overview of Microencapsulates for Use in Food Products or Processes and Methods to Make Them. In Encapsulation Technologies for Active Food Ingredients and Food Processing; Zuidam, N.J., Nedovic, V., Eds.; Springer: New York, NY, USA, 2010; Volume 1, pp. 3–29. [Google Scholar] [CrossRef]
- Champagne, C.; Fustier, P. Microencapsulation for delivery of probiotics and other ingredients in functional dairy products. In Functional Dairy Products; Elsevier: Amsterdam, The Netherlands, 2007; pp. 404–426. [Google Scholar] [CrossRef]
- Liu, H.; Cui, S.W.; Chen, M.; Li, Y.; Liang, R.; Xu, F.; Zhong, F. Protective approaches and mechanisms of microencapsulation to the survival of probiotic bacteria during processing, storage and gastrointestinal digestion: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 2863–2878. [Google Scholar] [CrossRef] [PubMed]
- Zaghari, L.; Basiri, A.; Rahimi, S. Preparation and characterization of double-coated probiotic bacteria via a fluid-bed process: A case study on Lactobacillus reuteri. Int. J. Food Eng. 2020, 16. [Google Scholar] [CrossRef]
- Krasaekoopt, W.; Bhandari, B.; Deeth, H. Evaluation of encapsulation techniques of probiotics for yoghurt. Int. Dairy J. 2003, 13, 3–13. [Google Scholar] [CrossRef]
- Lee, Y.; Ji, Y.R.; Lee, S.; Choi, M.-J.; Cho, Y. Microencapsulation of Probiotic Lactobacillus acidophilus KBL409 by Extrusion Technology to Enhance Survival under Simulated Intestinal and Freeze-Drying Conditions. J. Microbiol. Biotechnol. 2019, 29, 721–730. [Google Scholar] [CrossRef] [Green Version]
- Kanmani, P.; Kumar, R.S.; Yuvaraj, N.; Paari, K.; Pattukumar, V.; Arul, V. Effect of cryopreservation and microencapsulation of lactic acid bacterium Enterococcus faecium MC13 for long-term storage. Biochem. Eng. J. 2011, 58-59, 140–147. [Google Scholar] [CrossRef]
- Cook, M.T.; Tzortzis, G.; Charalampopoulos, D.; Khutoryanskiy, V.V. Production and Evaluation of Dry Alginate-Chitosan Microcapsules as an Enteric Delivery Vehicle for Probiotic Bacteria. Biomacromolecules 2011, 12, 2834–2840. [Google Scholar] [CrossRef]
- Chávarri, M.; Marañón, I.; Ares, R.; Ibáñez, F.C.; Marzo, F.; Villarán, M.D.C. Microencapsulation of a probiotic and prebiotic in alginate-chitosan capsules improves survival in simulated gastro-intestinal conditions. Int. J. Food Microbiol. 2010, 142, 185–189. [Google Scholar] [CrossRef]
- Li, X.Y.; Chen, X.G.; Sun, Z.W.; Park, H.J.; Cha, D.-S. Preparation of alginate/chitosan/carboxymethyl chitosan complex microcapsules and application in Lactobacillus casei ATCC 393. Carbohydr. Polym. 2011, 83, 1479–1485. [Google Scholar] [CrossRef]
- Zaeim, D.; Sarabi-Jamab, M.; Ghorani, B.; Kadkhodaee, R.; Tromp, R. Electrospray-assisted drying of live probiotics in acacia gum microparticles matrix. Carbohydr. Polym. 2018, 183, 183–191. [Google Scholar] [CrossRef]
- Ivanovska, T.P.; Mladenovska, K.; Zhivikj, Z.; Pavlova, M.J.; Gjurovski, I.; Ristoski, T.; Petrushevska-Tozi, L. Synbiotic loaded chitosan-Ca-alginate microparticles reduces inflammation in the TNBS model of rat colitis. Int. J. Pharm. 2017, 527, 126–134. [Google Scholar] [CrossRef]
- Hadzieva, J.; Mladenovska, K.; Crcarevska, M.S.; Dodov, M.G.; Dimchevska, S.; Geškovski, N.; Grozdanov, A.; Popovski, E.; Petruševski, G.; Chachorovska, M.; et al. Lactobacillus casei loaded Soy Protein-Alginate Microparticles prepared by Spray-Drying. Food Technol. Biotechnol. 2017, 55, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Moghanjougi, Z.M.; Bari, M.R.; Khaledabad, M.A.; Amiri, S.; Almasi, H. Microencapsulation of Lactobacillus acidophilus LA-5 and Bifidobacterium animalis BB-12 in pectin and sodium alginate: A comparative study on viability, stability, and structure. Food Sci. Nutr. 2021, 9, 5103–5111. [Google Scholar] [CrossRef] [PubMed]
- Coghetto, C.C.; Brinques, G.B.; Siqueira, N.; Pletsch, J.; Soares, R.M.; Ayub, M.A.Z. Electrospraying microencapsulation of Lactobacillus plantarum enhances cell viability under refrigeration storage and simulated gastric and intestinal fluids. J. Funct. Foods 2016, 24, 316–326. [Google Scholar] [CrossRef]
- Reddy, S.; He, L.; Ramakrishana, S.; Luo, H. Graphene nanomaterials for regulating stem cell fate in neurogenesis and their biocompatibility. Curr. Opin. Biomed. Eng. 2019, 10, 69–78. [Google Scholar] [CrossRef]
- Shi, H.; Liu, W.; Xie, Y.; Yang, M.; Liu, C.; Zhang, F.; Wang, S.; Liang, L.; Pi, K. Synthesis of carboxymethyl chitosan-functionalized graphene nanomaterial for anticorrosive reinforcement of waterborne epoxy coating. Carbohydr. Polym. 2020, 252, 117249. [Google Scholar] [CrossRef]
- Gholamali, I.; Yadollahi, M. Bio-nanocomposite Polymer Hydrogels Containing Nanoparticles for Drug Delivery: A Review. Regen. Eng. Transl. Med. 2021, 7, 129–146. [Google Scholar] [CrossRef]
- Lattuada, E.; Leo, M.; Caprara, D.; Salvatori, L.; Stoppacciaro, A.; Sciortino, F.; Filetici, P. DNA-GEL, Novel Nanomaterial for Biomedical Applications and Delivery of Bioactive Molecules. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef]
- Rabiee, N.; Bagherzadeh, M.; Ghadiri, A.M.; Fatahi, Y.; Baheiraei, N.; Safarkhani, M.; Aldhaher, A.; Dinarvand, R. Bio-multifunctional noncovalent porphyrin functionalized carbon-based nanocomposite. Sci. Rep. 2021, 11, 6604. [Google Scholar] [CrossRef]
- Thangrongthong, S.; Puttarat, N.; Ladda, B.; Itthisoponkul, T.; Pinket, W.; Kasemwong, K.; Taweechotipatr, M. Microencapsulation of probiotic Lactobacillus brevis ST-69 producing GABA using alginate supplemented with nanocrystalline starch. Food Sci. Biotechnol. 2020, 29, 1475–1482. [Google Scholar] [CrossRef]
- Yuan, L.; Wei, H.; Yang, X.-Y.; Geng, W.; Peterson, B.W.; van der Mei, H.C.; Busscher, H.J. Escherichia coli Colonization of Intestinal Epithelial Layers In Vitro in the Presence of Encapsulated Bifidobacterium breve for Its Protection against Gastrointestinal Fluids and Antibiotics. ACS Appl. Mater. Interfaces 2021, 13, 15973–15982. [Google Scholar] [CrossRef]
- Duarte, I.F.B.; Mergulhão, N.L.O.N.; Silva, V.D.C.; de Bulhões, L.C.G.; Júnior, I.D.B.; Silva, A.C. Natural Probiotics and Nanomaterials: A New Functional Food. In Prebiotics and Probiotics—From Food to Health; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Yu, H.; Park, J.-Y.; Kwon, C.W.; Hong, S.-C.; Park, K.-M.; Chang, P.-S. An Overview of Nanotechnology in Food Science: Preparative Methods, Practical Applications, and Safety. J. Chem. 2018, 2018, 5427978. [Google Scholar] [CrossRef]
- Razavi, S.; Janfaza, S.; Tasnim, N.; Gibson, D.L.; Hoorfar, M. Nanomaterial-based encapsulation for controlled gastrointestinal delivery of viable probiotic bacteria. Nanoscale Adv. 2021, 3, 2699–2709. [Google Scholar] [CrossRef]
- Mettu, S.; Hathi, Z.; Athukoralalage, S.; Priya, A.; Lam, T.N.; Ong, K.L.; Choudhury, N.R.; Dutta, N.K.; Curvello, R.; Garnier, G.; et al. Perspective on Constructing Cellulose-Hydrogel-Based Gut-Like Bioreactors for Growth and Delivery of Multiple-Strain Probiotic Bacteria. J. Agric. Food Chem. 2021, 69, 4946–4959. [Google Scholar] [CrossRef] [PubMed]
- Hasan, N.; Rahman, L.; Kim, S.-H.; Cao, J.; Arjuna, A.; Lallo, S.; Jhun, B.H.; Yoo, J.-W. Recent advances of nanocellulose in drug delivery systems. J. Pharm. Investig. 2020, 50, 553–572. [Google Scholar] [CrossRef]
- Park, M.; Lee, D.; Hyun, J. Nanocellulose-alginate hydrogel for cell encapsulation. Carbohydr. Polym. 2015, 116, 223–228. [Google Scholar] [CrossRef]
- Huq, T.; Fraschini, C.; Khan, A.; Riedl, B.; Bouchard, J.; Lacroix, M. Alginate based nanocomposite for microencapsulation of probiotic: Effect of cellulose nanocrystal (CNC) and lecithin. Carbohydr. Polym. 2017, 168, 61–69. [Google Scholar] [CrossRef]
- Liu, Y.; Lyu, Y.; Hu, Y.; An, J.; Chen, R.; Chen, M.; Du, J.; Hou, C. Novel Graphene Oxide Nanohybrid Doped Methacrylic Acid Hydrogels for Enhanced Swelling Capability and Cationic Adsorbability. Polymers 2021, 13, 1112. [Google Scholar] [CrossRef]
- Phan, L.; Vo, T.; Hoang, T.; Cho, S. Graphene Integrated Hydrogels Based Biomaterials in Photothermal Biomedicine. Nanomaterials 2021, 11, 906. [Google Scholar] [CrossRef]
- Wang, X.; Guo, W.; Li, L.; Yu, F.; Li, J.; Liu, L.; Fang, B.; Xia, L. Photothermally triggered biomimetic drug delivery of Teriparatide via reduced graphene oxide loaded chitosan hydrogel for osteoporotic bone regeneration. Chem. Eng. J. 2020, 413, 127413. [Google Scholar] [CrossRef]
- Lu, Y.-J.; Lan, Y.-H.; Chuang, C.-C.; Lu, W.-T.; Chan, L.-Y.; Hsu, P.-W.; Chen, J.-P. Injectable Thermo-Sensitive Chitosan Hydrogel Containing CPT-11-Loaded EGFR-Targeted Graphene Oxide and SLP2 shRNA for Localized Drug/Gene Delivery in Glioblastoma Therapy. Int. J. Mol. Sci. 2020, 21, 7111. [Google Scholar] [CrossRef]
- Kim, J.; Hlaing, S.P.; Lee, J.; Saparbayeva, A.; Kim, S.; Hwang, D.S.; Lee, E.H.; Yoon, I.-S.; Yun, H.; Kim, M.-S.; et al. Exfoliated bentonite/alginate nanocomposite hydrogel enhances intestinal delivery of probiotics by resistance to gastric pH and on-demand disintegration. Carbohydr. Polym. 2021, 272, 118462. [Google Scholar] [CrossRef]
- Batista, D.P.C.; de Oliveira, I.N.; Ribeiro, A.R.B.; Fonseca, E.J.S.; Santos-Magalhães, N.S.; de Sena-Filho, J.G.; Teodoro, A.V.; Grillo, L.A.M.; de Almeida, R.S.; Dornelas, C.B. Encapsulation and release of Beauveria bassiana from alginate–bentonite nanocomposite. RSC Adv. 2017, 7, 26468–26477. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, D.M.; Nunes, Y.L.; Figueirêdo, M.C.B.; de Azeredo, H.M.C.; Aouada, F.A.; Feitosa, J.P.A.; Rosa, M.F.; Dufresne, A. Nanocellulose nanocomposite hydrogels: Technological and environmental issues. Green Chem. 2018, 20, 2428–2448. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Chen, F. pH-Responsive Drug-Delivery Systems. Chem. Asian J. 2014, 10, 284–305. [Google Scholar] [CrossRef]
- Lavanya, K.; Chandran, S.V.; Balagangadharan, K.; Selvamurugan, N. Temperature- and pH-responsive chitosan-based injectable hydrogels for bone tissue engineering. Mater. Sci. Eng. C 2020, 111, 110862. [Google Scholar] [CrossRef]
- Andrade, F.; Roca-Melendres, M.M.; Durán-Lara, E.F.; Rafael, D.; Schwartz, S., Jr. Stimuli-Responsive Hydrogels for Cancer Treatment: The Role of pH, Light, Ionic Strength and Magnetic Field. Cancers 2021, 13, 1164. [Google Scholar] [CrossRef]
- Bazban-Shotorbani, S.; Hasani-Sadrabadi, M.M.; Karkhaneh, A.; Serpooshan, V.; Jacob, K.I.; Moshaverinia, A.; Mahmoudi, M. Revisiting structure-property relationship of pH-responsive polymers for drug delivery applications. J. Control. Release 2017, 253, 46–63. [Google Scholar] [CrossRef]
- Mei, L.; He, F.; Zhou, R.-Q.; Wu, C.-D.; Liang, R.; Xie, R.; Ju, X.-J.; Wang, W.; Chu, L.-Y. Novel Intestinal-Targeted Ca-Alginate-Based Carrier for pH-Responsive Protection and Release of Lactic Acid Bacteria. ACS Appl. Mater. Interfaces 2014, 6, 5962–5970. [Google Scholar] [CrossRef]
- Singh, I.; Singh, I.; Kumar, P.; Kumar, P.; Pillay, V.; Pillay, V.; Singh, I.; Singh, I.; Kumar, P.; Kumar, P.; et al. Site-specific delivery of polymeric encapsulated microorganisms: A patent evaluation of US20170165201A1. Expert Opin. Ther. Patents 2018, 28, 703–708. [Google Scholar] [CrossRef]
- Gately, N.M.; Kennedy, J.E. The Development of a Melt-Extruded Shellac Carrier for the Targeted Delivery of Probiotics to the Colon. Pharmaceutics 2017, 9, 38. [Google Scholar] [CrossRef]
- Stratford, M.; Steels, H.; Novodvorska, M.; Archer, D.B.; Avery, S.V. Extreme Osmotolerance and Halotolerance in Food-Relevant Yeasts and the Role of Glycerol-Dependent Cell Individuality. Front. Microbiol. 2019, 9, 3238. [Google Scholar] [CrossRef]
- Shen, T.-Y.; Qin, H.-L.; Gao, Z.-G.; Fan, X.-B.; Hang, X.-M.; Jiang, Y.-Q. Influences of enteral nutrition combined with probiotics on gut microflora and barrier function of rats with abdominal infection. World J. Gastroenterol. 2006, 12, 4352–4358. [Google Scholar] [CrossRef]
- Ma, J.; Wang, W.; Sun, C.; Gu, L.; Liu, Z.; Yu, W.; Chen, L.; Jiang, Z.; Hou, J. Effects of environmental stresses on the physiological characteristics, adhesion ability and pathogen adhesion inhibition of Lactobacillus plantarum KLDS 1.0328. Process Biochem. 2020, 92, 426–436. [Google Scholar] [CrossRef]
- Singh, P.; Medronho, B.; Alves, L.; Da Silva, G.J.; Miguel, M.; Lindman, B. Development of carboxymethyl cellulose-chitosan hybrid micro- and macroparticles for encapsulation of probiotic bacteria. Carbohydr. Polym. 2017, 175, 87–95. [Google Scholar] [CrossRef]
- Enck, K.; Banks, S.; Yadav, H.; Welker, M.E.; Opara, E.C. Development of a Novel Oral Delivery Vehicle for Probiotics. Curr. Pharm. Des. 2020, 26, 3134–3140. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, C.; Zhou, W.; Luan, Q.; Li, W.; Deng, Q.; Dong, X.; Tang, H.; Huang, F. A pH-Responsive Gel Macrosphere Based on Sodium Alginate and Cellulose Nanofiber for Potential Intestinal Delivery of Probiotics. ACS Sustain. Chem. Eng. 2018, 6, 13924–13931. [Google Scholar] [CrossRef]
- Wang, M.; Zang, Y.; Hong, K.; Zhao, X.; Yu, C.; Liu, D.; An, Z.; Wang, L.; Yue, W.; Nie, G. Preparation of pH-sensitive carboxymethyl cellulose/chitosan/alginate hydrogel beads with reticulated shell structure to deliver Bacillus subtilis natto. Int. J. Biol. Macromol. 2021, 192, 684–691. [Google Scholar] [CrossRef]
- Zheng, H.; Gao, M.; Ren, Y.; Lou, R.; Xie, H.; Yu, W.; Liu, X.; Ma, X. An improved pH-responsive carrier based on EDTA-Ca-alginate for oral delivery of Lactobacillus rhamnosus ATCC 53103. Carbohydr. Polym. 2016, 155, 329–335. [Google Scholar] [CrossRef]
- Ben Thomas, M.; Vaidyanathan, M.; Radhakrishnan, K.; Raichur, A.M. Enhanced viability of probiotic Saccharomyces boulardii encapsulated by layer-by-layer approach in pH responsive chitosan–dextran sulfate polyelectrolytes. J. Food Eng. 2014, 136, 1–8. [Google Scholar] [CrossRef]
- Chandrawati, R. Enzyme-responsive polymer hydrogels for therapeutic delivery. Exp. Biol. Med. 2016, 241, 972–979. [Google Scholar] [CrossRef]
- Wang, Y.; Heng, C.; Zhou, X.; Cao, G.; Jiang, L.; Wang, J.; Li, K.; Wang, D.; Zhan, X. Supplemental Bacillus subtilis DSM 29784 and enzymes, alone or in combination, as alternatives for antibiotics to improve growth performance, digestive enzyme activity, anti-oxidative status, immune response and the intestinal barrier of broiler chickens. Br. J. Nutr. 2020, 125, 494–507. [Google Scholar] [CrossRef]
- Liu, L.; Fishman, M.L.; Kost, J.; Hicks, K.B. Pectin-based systems for colon-specific drug delivery via oral route. Biomaterials 2003, 24, 3333–3343. [Google Scholar] [CrossRef]
- Lai, H.; Lin, K.; Zhang, W.; Zhang, Z.; Jie, L.; Wu, Y.; He, Q. Development of pH- and enzyme-controlled, colon-targeted, pulsed delivery system of a poorly water-soluble drug: Preparation and in vitro evaluation. Drug Dev. Ind. Pharm. 2010, 36, 81–92. [Google Scholar] [CrossRef]
- Pinto, J.F. Site-specific drug delivery systems within the gastro-intestinal tract: From the mouth to the colon. Int. J. Pharm. 2010, 395, 44–52. [Google Scholar] [CrossRef]
- Shrivastava, S.; Shrivastava, A.; Sinha, S. Dextran carrier macromolecules for colon-specific delivery of 5-aminosalicylic acid. Indian J. Pharm. Sci. 2013, 75, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Chen, L.; McClements, D.J.; Qiu, C.; Li, C.; Zhang, Z.; Miao, M.; Tian, Y.; Zhu, K.; Jin, Z. Stimulus-responsive hydrogels in food science: A review. Food Hydrocoll. 2021, 124, 107218. [Google Scholar] [CrossRef]
- Li, H.; Yew, Y.K. Simulation of soft smart hydrogels responsive to pH stimulus: Ionic strength effect and case studies. Mater. Sci. Eng. C 2009, 29, 2261–2269. [Google Scholar] [CrossRef]
- Rasool, N.; Yasin, T.; Heng, J.Y.; Akhter, Z. Synthesis and characterization of novel pH-, ionic strength and temperature- sensitive hydrogel for insulin delivery. Polymer 2010, 51, 1687–1693. [Google Scholar] [CrossRef]
- Neish, A.S. Redox signaling mediated by the gut microbiota. Free Radic. Res. 2013, 47, 950–957. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wang, Z.; Zhang, G.; Ren, J.; Yu, L.; Liu, X.; Yang, Y.; Ravindran, A.; Wong, C.; Chen, R. A pH/redox-dual responsive, nanoemulsion-embedded hydrogel for efficient oral delivery and controlled intestinal release of magnesium ions. J. Mater. Chem. B 2021, 9, 1888–1895. [Google Scholar] [CrossRef]
- Liu, H.; Rong, L.; Wang, B.; Xie, R.; Sui, X.; Xu, H.; Zhang, L.; Zhong, Y.; Mao, Z. Facile fabrication of redox/pH dual stimuli responsive cellulose hydrogel. Carbohydr. Polym. 2017, 176, 299–306. [Google Scholar] [CrossRef]
- Cresci, G.A.; Izzo, K. Gut Microbiome. In Adult Short Bowel Syndrome; Academic Press: Cambridge, MA, USA, 2019; pp. 45–54. ISBN 9780128143308. [Google Scholar]
- Durack, J.; Lynch, S.V. The gut microbiome: Relationships with disease and opportunities for therapy. J. Exp. Med. 2019, 216, 20–40. [Google Scholar] [CrossRef] [Green Version]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Ruan, W.; Engevik, M.; Spinler, J.K.; Versalovic, J. Healthy Human Gastrointestinal Microbiome: Composition and Function After a Decade of Exploration. Am. J. Dig. Dis. 2020, 65, 695–705. [Google Scholar] [CrossRef] [Green Version]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.R.; Yu, W.-H.; Lakshmanan, A.; Wade, W.G. The Human Oral Microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [Green Version]
- Quigley, E.M.M. Gut microbiome as a clinical tool in gastrointestinal disease management: Are we there yet? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 315–320. [Google Scholar] [CrossRef]
- Okubo, C.K. Biochemical studies of CELO virus: An oncogenic avian adenovirus. Experientia 1983, 39, 303–304. [Google Scholar] [CrossRef]
- Vasapolli, R.; Schütte, K.; Schulz, C.; Vital, M.; Schomburg, D.; Pieper, D.H.; Vilchez-Vargas, R.; Malfertheiner, P. Analysis of Transcriptionally Active Bacteria Throughout the Gastrointestinal Tract of Healthy Individuals. Gastroenterology 2019, 157, 1081–1092. [Google Scholar] [CrossRef] [Green Version]
- Hillman, E.T.; Lu, H.; Yao, T.; Nakatsu, C.H. Microbial Ecology along the Gastrointestinal Tract. Microbes Environ. 2017, 32, 300–313. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.; Chen, D.; Zhang, J.-N.; Lv, X.; Wang, K.; Duan, L.-P.; Nie, Y.; Wu, X.-L. Bacterial Community Mapping of the Mouse Gastrointestinal Tract. PLoS ONE 2013, 8, e74957. [Google Scholar] [CrossRef] [Green Version]
- Ducarmon, Q.R.; Zwittink, R.D.; Hornung, B.V.H.; Van Schaik, W.; Young, V.B.; Kuijper, E.J. Gut Microbiota and Colonization Resistance against Bacterial Enteric Infection. Microbiol. Mol. Biol. Rev. 2019, 83, e00007-19. [Google Scholar] [CrossRef] [PubMed]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Ross, R.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Genet. 2012, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, J.; Sineva, E.; Knight, J.; Levy, R.M.; Ebright, R.H. Antibacterial Peptide Microcin J25 Inhibits Transcription by Binding within and Obstructing the RNA Polymerase Secondary Channel. Mol. Cell 2004, 14, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Destoumieux-Garzón, D.; Péduzzi, J.; Thomas, X.; Djediat, C.; Rebuffat, S. Parasitism of Iron-siderophore Receptors of Escherichia Coli by the Siderophore-peptide Microcin E492m and its Unmodified Counterpart. BioMetals 2006, 19, 181–191. [Google Scholar] [CrossRef]
- Parks, W.M.; Bottrill, A.R.; Pierrat, O.A.; Durrant, M.C.; Maxwell, A. The action of the bacterial toxin, microcin B17, on DNA gyrase. Biochimie 2007, 89, 500–507. [Google Scholar] [CrossRef]
- Thanissery, R.; Winston, J.A.; Theriot, C.M. Inhibition of spore germination, growth, and toxin activity of clinically relevant C. difficile strains by gut microbiota derived secondary bile acids. Anaerobe 2017, 45, 86–100. [Google Scholar] [CrossRef]
- Watanabe, M.; Fukiya, S.; Yokota, A. Comprehensive evaluation of the bactericidal activities of free bile acids in the large intestine of humans and rodents. J. Lipid Res. 2017, 58, 1143–1152. [Google Scholar] [CrossRef] [Green Version]
- Kurdi, P.; Kawanishi, K.; Mizutani, K.; Yokota, A. Mechanism of Growth Inhibition by Free Bile Acids in Lactobacilli and Bifidobacteria. J. Bacteriol. 2006, 188, 1979–1986. [Google Scholar] [CrossRef] [Green Version]
- Sannasiddappa, T.H.; Lund, P.A.; Clarke, S.R.; Sannasiddappa, T. In Vitro Antibacterial Activity of Unconjugated and Conjugated Bile Salts on Staphylococcus aureus. Front. Microbiol. 2017, 8, 1581. [Google Scholar] [CrossRef] [Green Version]
- Chua, J.C.L.; Hale, J.D.F.; Silcock, P.; Bremer, P.J. Bacterial survival and adhesion for formulating new oral probiotic foods. Crit. Rev. Food Sci. Nutr. 2019, 60, 2926–2937. [Google Scholar] [CrossRef] [PubMed]
- Van Tassell, M.L.; Miller, M.J. Lactobacillus Adhesion to Mucus. Nutrients 2011, 3, 613–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, M.B.; Diaz, J.P.; Muñoz-Quezada, S.; Llorente, C.G.; Gil, A. Probiotic Mechanisms of Action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Mercier-Bonin, M.; Chapot-Chartier, M.-P. Surface Proteins of Lactococcus lactis: Bacterial Resources for Muco-adhesion in the Gastrointestinal Tract. Front. Microbiol. 2017, 8, 2247. [Google Scholar] [CrossRef] [PubMed]
- Juge, N. Microbial adhesins to gastrointestinal mucus. Trends Microbiol. 2012, 20, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Schwarz-Linek, U.; Höök, M.; Potts, J.R. Fibronectin-binding proteins of Gram-positive cocci. Microbes Infect. 2006, 8, 2291–2298. [Google Scholar] [CrossRef] [PubMed]
- Saára, M.; Sleytr, U.B. S-Layer Proteins. J. Bacteriol. 2000, 182, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Han, J.; Gong, G.; Koffas, M.A.G.; Zha, J. Wall teichoic acids: Physiology and applications. FEMS Microbiol. Rev. 2020, 45, fuaa064. [Google Scholar] [CrossRef]
- Zhou, Y.; Cui, Y.; Qu, X. Exopolysaccharides of lactic acid bacteria: Structure, bioactivity and associations: A review. Carbohydr. Polym. 2018, 207, 317–332. [Google Scholar] [CrossRef]
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and aggregation ability of probiotic strainLactobacillus acidophilusM92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Cao, Y.; Ferguson, L.R.; Shu, Q.; Garg, S. Evaluation of mucoadhesive coatings of chitosan and thiolated chitosan for the colonic delivery of microencapsulated probiotic bacteria. J. Microencapsul. 2012, 30, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-C.; Mi, F.-L.; Liao, Z.-X.; Hsiao, C.-W.; Sonaje, K.; Chung, M.-F.; Hsu, L.-W.; Sung, H.-W. Recent advances in chitosan-based nanoparticles for oral delivery of macromolecules. Adv. Drug Deliv. Rev. 2013, 65, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.M.; Teague, S.J. Hydrogen Bonding, Hydrophobic Interactions, and Failure of the Rigid Receptor Hypothesis. Angew. Chem. Int. Ed. 1999, 38, 736–749. [Google Scholar] [CrossRef]
- Shi, L.; Caldwell, K.D. Mucin Adsorption to Hydrophobic Surfaces. J. Colloid Interface Sci. 2000, 224, 372–381. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Walter, J.; Maldonado-Gómez, M.X.; Martínez, I. To engraft or not to engraft: An ecological framework for gut microbiome modulation with live microbes. Curr. Opin. Biotechnol. 2017, 49, 129–139. [Google Scholar] [CrossRef]
- Ma, C.; Wasti, S.; Huang, S.; Zhang, Z.; Mishra, R.; Jiang, S.; You, Z.; Wu, Y.; Chang, H.; Wang, Y.; et al. The gut microbiome stability is altered by probiotic ingestion and improved by the continuous supplementation of galactooligosaccharide. Gut Microbes 2020, 12, 1785252. [Google Scholar] [CrossRef]
- Mao, B.; Gu, J.; Li, D.; Cui, S.; Zhao, J.; Zhang, H.; Chen, W. Effects of Different Doses of Fructooligosaccharides (FOS) on the Composition of Mice Fecal Microbiota, Especially the Bifidobacterium Composition. Nutrients 2018, 10, 1105. [Google Scholar] [CrossRef] [Green Version]
- Hansen, C.H.F.; Larsen, C.S.; Petersson, H.O.; Zachariassen, L.F.; Vegge, A.; Lauridsen, C.; Kot, W.; Krych, A.; Nielsen, D.S.; Hansen, A.K. Targeting gut microbiota and barrier function with prebiotics to alleviate autoimmune manifestations in NOD mice. Diabetologia 2019, 62, 1689–1700. [Google Scholar] [CrossRef] [Green Version]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Wang, J.; Sailer, M.; Theis, S.; Verbeke, K.; Raes, J. Prebiotic inulin-type fructans induce specific changes in the human gut microbiota. Gut 2017, 66, 1968–1974. [Google Scholar] [CrossRef]
- Islam, S.U. Clinical Uses of Probiotics. Medicine 2016, 95, e2658. [Google Scholar] [CrossRef] [PubMed]
- Doron, S.; Snydman, D. Risk and Safety of Probiotics. Clin. Infect. Dis. 2015, 60, S129–S134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA. Generally Recognized as Safe (GRAS). Available online: https://www.fda.gov/food/food-ingredients-packaging/generally-recognized-safe-gras (accessed on 25 January 2022).
- Dronkers, T.M.; Ouwehand, A.C.; Rijkers, G.T. Global analysis of clinical trials with probiotics. Heliyon 2020, 6, e04467. [Google Scholar] [CrossRef]
- Degnan, F.H. Clinical studies involving probiotics: When FDA’s investigational new drug rubric applies-and when it may not. Gut Microbes 2012, 3, 485–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
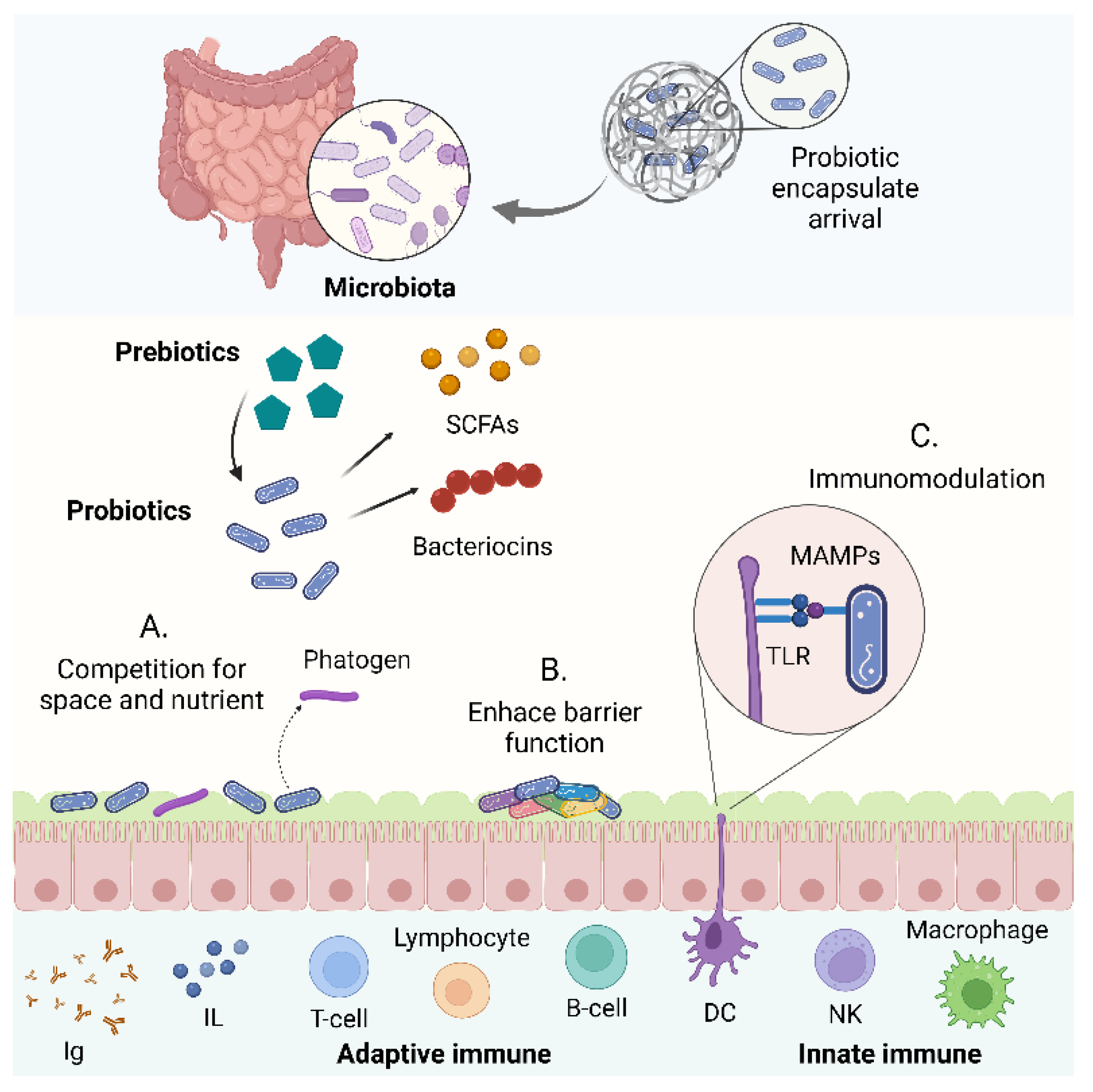{kind=link}
| GI Tract Part | Enzyme | Function and Characteristics | Ref. |
|---|---|---|---|
| Mouth | Salivary amylase | It digests starch into maltose and maltotriose by working at an optimum pH of 6.7 to 7. This cleavage decreases the glucose-polymer chain length and the viscosity of starch. | [28,29] |
| Lingual lipase | It catalyzes the hydrolysis of fatty acids. It hydrolyzes the ester bonds in the triglycerides to form diacylglycerols and monoacylglycerols. | [28,30] | |
| Stomach | Pepsin | It breaks down the internal peptide bonds of proteins at an optimal pH from 2 to 3. | [28] |
| Small intestine | Pancreatic amylase | It digests starch in the duodenal portion into maltose and maltotriose. | [29] |
| Trypsin | It is an endopeptidase that hydrolyzes the internal peptide bonds of proteins. It converts chymotrypsinogen, procarboxypeptidase, and proelastase to their active forms. | [28,31] | |
| Pectinase | It causes the degradation of pectin chains, a polysaccharide found in the cell wall of plants. | [32,33] | |
| Peptidase | It plays a role in protein digestion before absorption. | [34] | |
| Lipase | It is produced in the pancreas and oversees fat digestion. It breaks down triglycerides into free fatty acids and glycerol. | [35] |
| Material | Processing Method | Crosslinking Agent | Delivery System | Encapsulated Probiotics Strain | Ref. |
|---|---|---|---|---|---|
| Thiolated hyaluronic acid | Self-crosslinking | - | Macrogels | L. rhamnosus | [77] |
| Pectin methylesterase | Ionotropic gelation | CaCl2 | Macrogels | L. casei | [95] |
| Type A Gelatin | Cold gelation | Glutaraldehyde | Macrogels | K. lactis | [83] |
| Chitosan-coated alginate | Dual aerosols Ionotropic gelation | CaCl2 | Core/shell beads | L. rhamnosus L. acidophilus B. longum | [96,97] |
| Calcium-alginate | Ionotropic gelation | CaCl2 | Microgels | L. lactis | [98] |
| Alginate | Ionic crosslinking | Calcium ion (Ca2+) | Multilayer beads | B. breve | [99] |
| Pectin/Glucose | Ionotropic gelation | CaCl2 | Freeze-dried gels | L. rhamnosus | [79] |
| Alginate/low methoxyl pectin Alginate/κ-carrageenan | Ionotropic gelation | CaCl2 | Double layer beads | L. rhamnosus | [38] |
| Pectin/Iron | Ionotropic gelation | FeSO4 | Freeze-dried beads | L. plantarum | [84] |
| Gelatin/Sodium Alginate | Ionotropic gelation | CaCl2 | Gels Microbeads | B. longum L. bulgaricus | [72,100] |
| Alginate/Basil seed mucilage | Ionotropic gelation | CaCl2 | Hydrogel beads | E. faecium | [101] |
| methacrylate-modified gelatin methacrylate-modified hyaluronic acid | Light-irradiation crosslinking | LAP | Microbeads immobilized in a hydrogel matrix | L.reuteri | [102] |
| Zein-coated alginate | Ionotropic gelation | CaCl2 | Core-shell beds | B. bifidum | [103] |
| Alginate/xanthan gum Alginate/gum acacia Alginate/sodium caseinate Alginate/chitosan Alginate/starch Alginate/carrageenan | Ionic crosslinking | CaCl2 | Core-shell beds | L. rhamnosus | [4] |
| Calcium Alginate | Vibrating nozzle encapsulator Ionotropic gelation | CaCl2 | Single-layer beads | L. plantarum L.rhamnosus L. lactis L. acidophilus L. casei | [104,105,106] |
| Pectin/Inulin Pectin | Water/Oil blending Ionotropic gelation | CaCl2 | Single-layer beads | L. casei L. rhamnosus | [107] |
| Calcium-alginate-soy protein isolate | Ionotropic gelation | CaCl2 | Single-layer beads | L. plantarum | [108] |
| PLGA/alginate | Solvent Evaporation | - | Microbeads immobilized in a hydrogel matrix | B. breve | [109] |
| Chitosan/Dextran Sulfate (DXS) | Gelation | Genipin | Bulky Hydrogel Single-layer beads | L. rhamnosus | [85] |
| Delivery System | Material | Processing Method | Encapsulated Probiotics Strain | Ref. |
|---|---|---|---|---|
| Microcapsules | Alginate/chitosan | Freeze drying External gelation | E. faecium B. breve | [135,136] |
| Microspheres | Alginate/chitosan | Extrusion | L. gasseri B. bifidum | [137] |
| Microcapsules | Alginate/chitosan/carboxymethyl cellulose | Extrusion | L. casei | [138] |
| Microparticles | Acacia gum | Electrospray | L. plantarum | [139] |
| Microparticles | Chitosan/Calcium/alginate | Spray drying | L. casei | [140] |
| Microparticles | Soy protein isolate/Alginate | Spray drying | L. casei | [141] |
| Microcapsules | Pectin/Sodium alginate | Emulsion | L. acidophilus B. animalis | [142] |
| Microcapsules | Sodium alginate Sodium alginate-citric pectin | Electrospray | L. plantarum | [143] |
| Platform Material | Nanomaterial | Encapsulated Probiotics Strain | Attractive Properties | Ref. |
|---|---|---|---|---|
| Propylene glycol alginate | β-lactoglobulin (β-lg) Nanoparticles | L. rhamnosus | High trapping efficiency for bioactive molecules. | [80] |
| Alginate/lecithin | Cellulose Nanocrystals (CNC) | L. rhamnosus | Improved compression strength, decreased swelling in the gastric fluid, and increased cell viability than unmodified alginate encapsulates. | [157] |
| Alginate | Bentonite | B. bassiana L. rhamnosus | Reduced permeability, release kinetics, and pore size. Delayed gastric fluid penetration. | [162,163] |
| Gelatin | Graphene Oxide (GO) | K. lactis | Higher mechanical stability and integrity. Tunable degradation rates by varying GO composition. | [86] |
| Material | Processing Method | Configuration | Encapsulated Probiotics | Functionality | Ref. |
|---|---|---|---|---|---|
| Carboxymethyl Cellulose/Chitosan | Drop-wise addition Nozzle-spray | Macroparticles Microparticles | L. rhamnosus | Reduced swelling at pH 2.4 and remarkable swelling at pH 7.4. | [175] |
| Oxidized Alginate | Ionotropic gelation | Single-layer beads | L. casei | Highly resistant to the acidic environment by polymeric rearrangement and sensitive degradation in neutral-basic pH. | [176] |
| Alginate (SA) and TEMPO-oxidized cellulose nanofiber | Ionotropic gelation | Macrospheres | L. plantarum | Controlled swelling in the intestinal fluid and core compact in the gastric fluid. | [177] |
| Carboxymethyl cellulose/chitosan/alginate | Ionotropic gelation | Multilayer beads | B. subtilis | pH-dependent degradability rates. | [178] |
| EDTA-Ca- Alginate | Emulsification | Microspheres | L. rhamnosus | pH-driven swelling and disassembly mediated by calcium (Ca2+) release. | [179] |
| Chitosan and dextran sulfate | Layer-by-layer technique | Microcapsules | S. boulardii | pH-dependent electrostatic interactions between raw materials enable an almost impermeable protective matrix at low pH and the appearance of pores at neutral pH. | [180] |
| Ca-alginate/protamine | Co-extrusion | Microcapsule | L. casei | Blockage of diffusion channels at acidic pH and easy degradation at a neutral pH environment. | [169] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Brand, A.J.; Quezada, V.; Gonzalez-Melo, C.; Bolaños-Barbosa, A.D.; Cruz, J.C.; Reyes, L.H. Novel Developments on Stimuli-Responsive Probiotic Encapsulates: From Smart Hydrogels to Nanostructured Platforms. Fermentation 2022, 8, 117. https://doi.org/10.3390/fermentation8030117
Garcia-Brand AJ, Quezada V, Gonzalez-Melo C, Bolaños-Barbosa AD, Cruz JC, Reyes LH. Novel Developments on Stimuli-Responsive Probiotic Encapsulates: From Smart Hydrogels to Nanostructured Platforms. Fermentation. 2022; 8(3):117. https://doi.org/10.3390/fermentation8030117
Chicago/Turabian StyleGarcia-Brand, Andres J., Valentina Quezada, Cristina Gonzalez-Melo, Angie D. Bolaños-Barbosa, Juan C. Cruz, and Luis H. Reyes. 2022. "Novel Developments on Stimuli-Responsive Probiotic Encapsulates: From Smart Hydrogels to Nanostructured Platforms" Fermentation 8, no. 3: 117. https://doi.org/10.3390/fermentation8030117
APA StyleGarcia-Brand, A. J., Quezada, V., Gonzalez-Melo, C., Bolaños-Barbosa, A. D., Cruz, J. C., & Reyes, L. H. (2022). Novel Developments on Stimuli-Responsive Probiotic Encapsulates: From Smart Hydrogels to Nanostructured Platforms. Fermentation, 8(3), 117. https://doi.org/10.3390/fermentation8030117