Abstract
Reactive compounds with one or more sulfane sulfur atoms can be an important source of reductive off-odors in wine. These substances contain labile sulfur, which can participate in microbiological (enzymatic) and chemical transformations (including in the post-bottling period), releasing malodorous hydrogen sulfide (H2S) and its derivatives (MeSH, EtSH, etc.). The following sulfane sulfur compounds were considered in this review as important precursors in the wine chemistry of reductive aromas: elemental sulfur (S8), persulfides (R-S-S-H), polysulfanes (R-Sn-R(′)), polythionates (−O3S-Sn-SO3−), thiosulfate (S2O32−) and derivatives of (poly)sulfane monosulfonic acids (R-Sn-SO3H). This review discusses the formation of these compounds, their reactivity and chemical transformations in wine, including reactions of nucleophilic substitution. In particular, the reactions of thiolysis, thiosulfatolysis and sulfitolysis of sulfane sulfur compounds are described, which lead in the end to reductive aroma compounds. In this way, the review attempts to shed light on some of the mysteries in the field of sulfur chemistry in wine and the reappearance of reductive off-odors after bottling.
1. Introduction
Hydrogen sulfide (H2S) and its derivatives (MeSH, EtSH, etc.) are among the main molecules responsible for reductive defects in wine, providing distinctive odor of “rotten eggs”, “cooked cabbage”, “onion”, etc. [1]. The sensory threshold levels in wine for the listed sulfur compounds are often presented in the range of 0.3–3 µg/L [2,3]). However, other studies demonstrated that reductive off-odors only start to be perceived at much higher concentrations [4,5,6,7]. Thus, in the case of H2S, the sensory thresholds can reach 80–150 µg/L. These values can depend on different factors, including wine matrix, individuality and training of the panelists, etc. Despite the variation of sensory thresholds, the content of sulfur compounds responsible for reductive aromas can be well above these values in some wines [1,5,6]. Therefore, the analysis of H2S, MeSH, EtSH and other sulfur-containing compounds is important for wine quality control and research. The efficient analytical approaches based on gas chromatography with sulfur chemiluminescence detection (GC-SCD) or pulse flame photometric detection (GC-PFPD) were proposed about two decades ago [3,8,9,10,11]. Comprehensive reviews of the analysis of various sulfur compounds in wine are given by Mestres et al. [1] and Fracassetti and Vigentini [6].
Faulty wines with reductive off-flavors remain a problem for the wine industry, resulting in large economic losses each year [12]. Possible solutions to this problem have long been one of the main topics in winemaking research. The compounds with reductive aromas can appear in wine during fermentation as by-products released by yeasts or after wine bottling due to chemical transformations of various precursors. They can also reappear in wines that have been specifically treated before bottling (e.g., with Cu salts) to remove H2S and its derivatives. This fact raises questions about the nature of sulfur compounds in wine and their transformations, which still remain unclear. This review is a continuation of our research to comprehend the phenomena of sulfur compounds in wine [13] based on recent studies.
When it comes to living organisms, the release of H2S is closely related to its endogenous role as a small signaling molecule in different kinds of living matter. The first manifestations of this phenomenon began in the early stages of life on Earth with sulfur bacteria, followed by a variety of eubacteria, fungi and yeasts (including those used in winemaking), up to the high forms of life, like plants and mammals [14,15,16]. From the winemaking point of view, yeasts (e.g., Saccharomyces cerevisiae) with their metabolic pathways have a considerable impact on the character of wine. They release not only ethanol and CO2, but other substances in smaller quantities, including various sulfur compounds [17,18,19]. Since the latter can strongly influence wine aroma, both positively (e.g., 3-mercaptohexanol (3-MH), 4-methyl-4-mercaptopentan-2-one (4-MMP)) and negatively (e.g., H2S, MeSH, EtSH), the sulfur metabolism of yeasts is one of the main factors affecting the quality of wine [20]. Due to the reactivity of sulfhydryl (HS-containing) compounds with wine components, the reductive aroma substances can be trapped by converting into products (precursors). Later, during the wine aging, these precursors can decompose, releasing the initial malodorous compounds. The formation of such precursors can be additionally induced by wine treatments (e.g., aeration, copper fining), the purpose of which is to eliminate reductive aromas in wine before bottling. Therefore, these treatments often turn out to be temporary measures when curing defected wines, and reductive aromas can reappear, for example, already after seven weeks of anoxic wine storage [21].
There are various candidates for precursors of reductive sulfur substances in wine, which were discussed in detail by Kreitman and colleagues in a review some years ago [22]: quinone- and carbonyl-sulfhydryl adducts, metal-sulfhydryl complexes, S-alkyl thioacetates, precursors with S-S groups (sulfane sulfur compounds). In recent years, significant progress has been made in identifying sulfane sulfur substances in wine and investigating their role as latent precursors of reductive off-odors. Therefore, the objective of this review was to analyze and summarize the latest data on sulfane sulfur precursors in wine. Here we provide an overview on the recent developments concerning the reappearance of sulfur compounds responsible for reductive defects in wine after bottling, including:
- A description of sulfane sulfur precursors and a discussion of their importance;
- Possible reactions of chemicals and enzymatic formation of sulfane sulfur precursors, including the role of the sulfur metabolism of Saccharomyces cerevisiae yeasts;
- Chemical reactions and their mechanisms which lead to the formation of reductive off-odors in wine from sulfane sulfur precursors after bottling.
2. Definition of Sulfane Sulfur Compounds
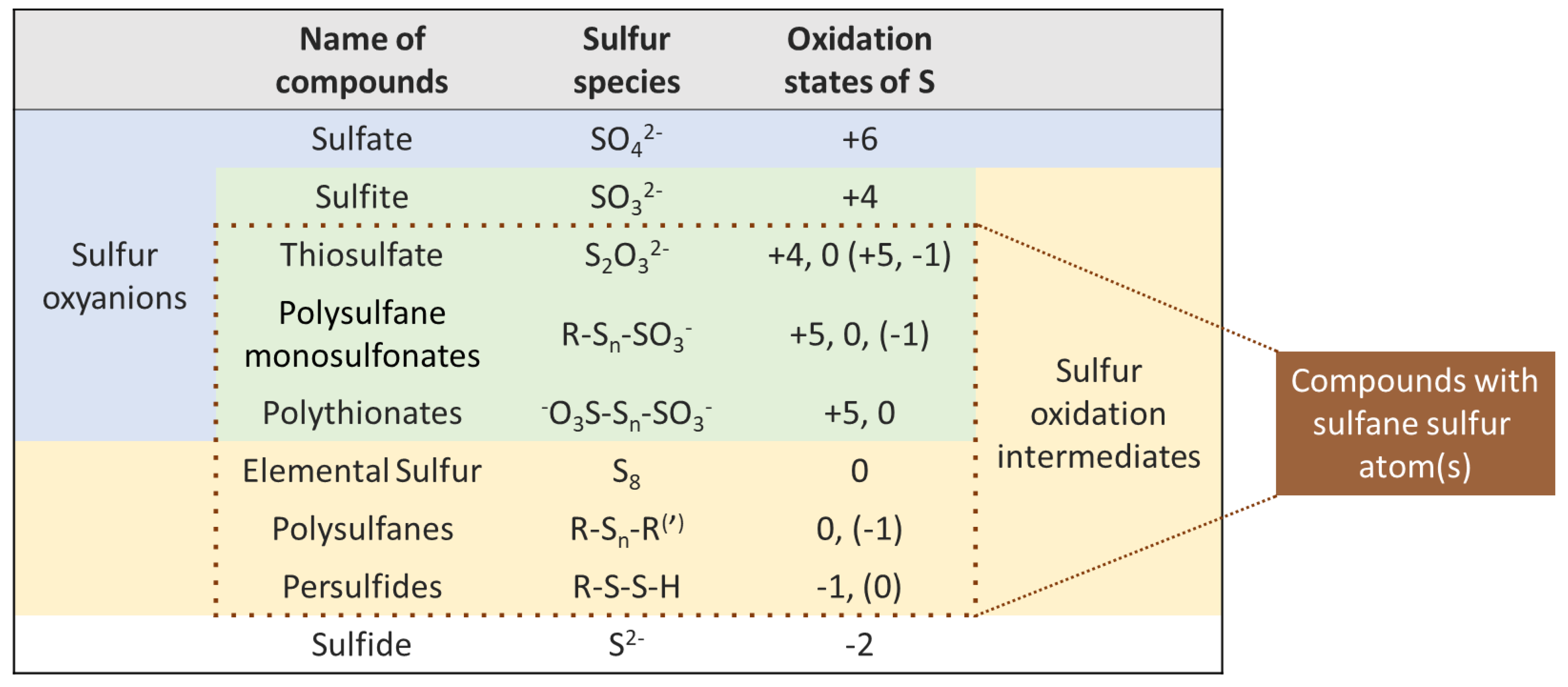
The notion of “sulfane sulfur compounds” varies somewhat among different authors. On one side, the term “sulfane sulfur” describes sulfur atoms that possess only covalent bonds with other sulfur atoms, i.e., the “-S-S-S-” fragment [23]. For example, elemental sulfur comprises exclusively sulfane sulfur atoms, which tend to self-associate and usually form an allotrope of S8 cycles. The broader concept of “sulfane sulfur” refers to a divalent sulfur atom bonded to at least one other sulfur atom, i.e., the “-S-S-” fragment [24,25,26]. In this review, we follow the second notion, since it allows us to discuss different types of substances with similar chemical properties together. Therefore, the following compounds can be considered as ones with sulfane sulfur atoms: persulfides (R-S-S-H), polysulfides, which are also called polysulfanes (R-Sn-R(′), n ≥ 2), polythionates (−O3S-Sn-SO3−, n ≥ 1), thiosulfate (S2O32−) and derivatives of (poly)sulfane monosulfonic acids (R-Sn-SO3H, n ≥ 1). Sulfane sulfur compounds have to be distinguished from sulfur oxyacids and their derivatives (e.g., sulfates and sulfites), which do not have the corresponding “-S-S-” fragment (Figure 1). The latter compounds possess higher oxidation states of sulfur and also play an important role in winemaking.
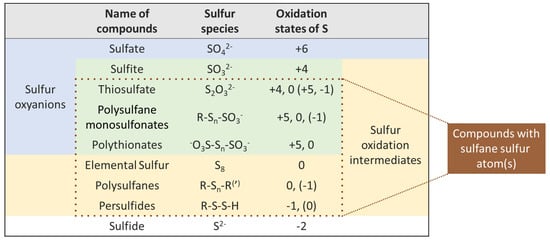
Figure 1.
Selected sulfur compounds and their oxidation states (values in parentheses vary due to resonance structures of sulfur species or R substituent).
Describing sulfane sulfur atoms more specifically, they do not exist in a free form but have the unique ability to be reversibly attached to other sulfur atoms [26]. The concept of sulfane sulfur was useful to consider the particularities of polysulfanes and persulfides, which were introduced into biological studies in the 1950s [16]. The term “sulfane sulfur” can be replaced by “bound sulfur” (which includes persulfides) [16]. Nowadays, both definitions are found in the scientific literature.
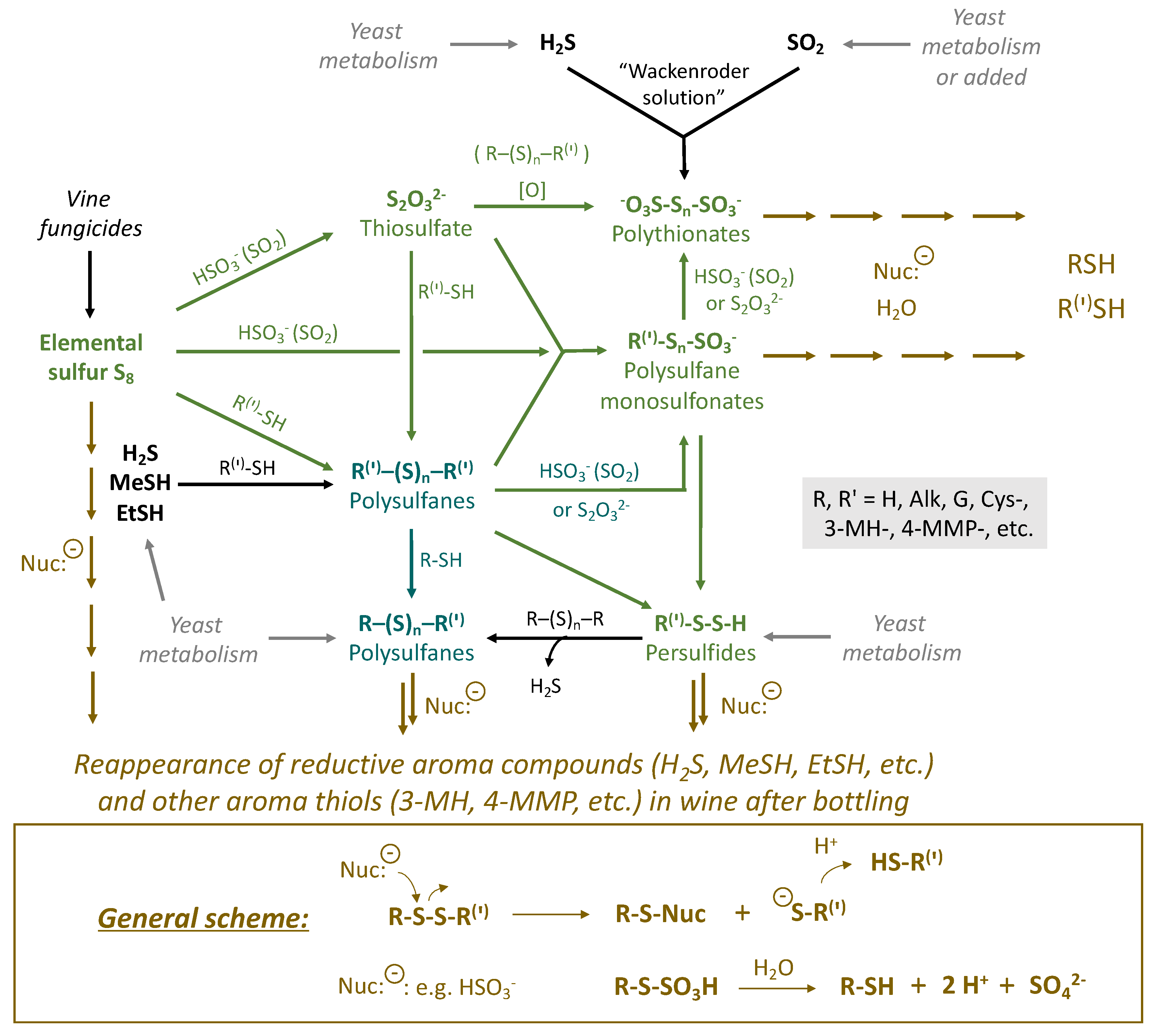
Sulfur compounds well-known in wine, whether from natural sources or artificially added, are present in all common oxidation states: hydrogen sulfide “−2”, elemental sulfur “0”, sulfur dioxide “+4”, sulfate “+6”, etc. A number of more complex sulfur compounds, including those with sulfane sulfurs and multiple oxidation states of S, can also occur in wine conditions, at least as short-lived species (Figure 1). Many of these compounds are suspected of being responsible for the reappearance of reductive faults after bottling and have been identified in wine. In general, sulfane sulfur compounds are considered as latent precursors of reductive off-odors due to their high reactivity and low oxidation states of sulfur atoms, which are similar to H2S and its derivatives (MeSH, EtSH, etc.). Various sulfane sulfur compounds are interrelated and can be mutually interconverted in wine (Figure 2). Many of the indicated reactions can be reversible under wine conditions. The presented intermediates are susceptible to further nucleophilic attacks, making this reaction scheme rather complicated.
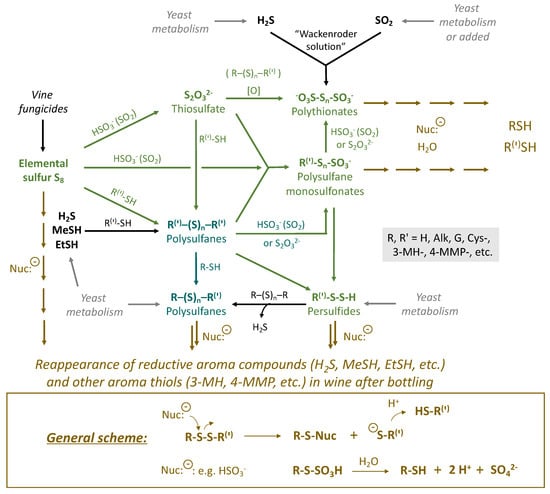
Figure 2.
General scheme of the interrelation of sulfane sulfur compounds in wine.
Figure 2 demonstrates the general scheme of chemical transformations, which will be presented in more detail in the subsequent figures separately with the corresponding colors green and turquoise (Section 3.1 and Section 3.4). The brown color corresponds to the transformations, which lead to the appearance of reductive aroma compounds in wine in the post-bottling period. Various nucleophiles in wine (e.g., HSO3−) can attack precursors by sulfane sulfur atoms and break the molecules. This results in the formation of smaller molecules (R(‘)-SH) [27], including H2S and thiols (Figure 2). In turn, polythionates and polysulfane monosulfonates may hydrolyze with the formation of various sulfhydryls (R-SH) as well [28]. Similar processes can be relevant not only for the reappearance of reductive off-odor compounds but also for other odorous thiols, e.g., 3-MH and 4-MMP, which are responsible for varietal wine aromas. The latter, however, are not the main subject of the actual review but can assist other investigations related to the development and stability of the aroma bouquet of wines rich in varietal thiols. At the same time, excessive release of 3-MH and 4-MMP can turn their positive fruity sensory perception into unpleasant odors in wine, which can be described, for example, as a “cat urine” smell.
3. Sulfane Sulfur Compounds in Wine and Their Transformations
Sulfane sulfur compounds have attracted the attention of scientists in various fields. For example, persulfides and polysulfanes were studied in mammalian cells and tissues as potentially important signaling/effector species [29]. Polysulfanes were detected in the fungus Aspergillus nidulans [30], and the question of their presence in Saccharomyces cerevisiae has also been raised. Currently we deal with a number of sulfane sulfur compounds, which have been identified in wine or hypothesized to be present as intermediates (Table 1).

Table 1.
Compounds with sulfane sulfur in wine.
One of the potential sources of sulfane sulfur compounds in wine is yeast metabolism. Yeasts usually assimilate hydrogensulfite HSO3− and sulfate SO42−, integrating them into sulfur metabolism and releasing reductive aroma compounds in wine, such as H2S, MeSH, EtSH, etc. [5,52]. These substances could subsequently react with various sulfur compounds present in wine, such as R(I)-SH or SO2, yielding sulfane sulfur compounds (Figure 2). The latter can serve later, after bottling, as precursors of reductive aroma substances.
Another source of sulfane sulfur substances in must and wine can be related to the chemical reactions of elemental sulfur (traces of vineyard fungicides in must/wine) and various sulfur nucleophiles, e.g., hydrogensulfite HSO3−, which is added in different steps of winemaking (Figure 2).
Sulfur nucleophiles can react with various sulfur compounds in wine and differ in their content and reactivity. The latter is related to nucleophilicity (in the case of sulfane sulfur compounds, it is thiophilicity, rather), which primarily is a function of basicity. The values of acid dissociation constants (pKa) of selected sulfur compounds are given in Table 2. For example, it is noteworthy that glutathione persulfide GSSH and similar compounds with longer sulfur chains are more acidic than glutathione (GSH), which means a higher concentration of corresponding anions (GSS−).
Table 2.
pKa of selected sulfur species in wine (based on [53,54]).
Steric effects of bigger substituents (R) in compounds with HS-groups also have an impact and decrease their reactivity. Thus, in reactions with 4-methyl-quinone, the nucleophilicity of the R-SH compounds was as follows [55]:
4-MMP < 3-MH < SO2 ≈ GSH ≈ H2S
In recent years, sulfane sulfur compounds and their reactions in wine have been intensively studied and many scientific works have been published. In order to keep our discussion comprehensive and up to date, this review is mainly based on the scientific articles presented over the past five years. Different types of sulfane sulfur compounds are discussed separately in the following subsections. These substances are usually present in a free form in wine. Later, in Section 5, we will also briefly discuss sulfane sulfur compounds bound to proteins.
3.1. Elemental Sulfur (S8)
Of all sulfane sulfur compounds, elemental sulfur can be considered the ideal representative of this family. Cyclooctasulfur S8 is the most common sulfur allotrope at ambient temperatures [56]. Each sulfur atom in the ring is bonded to two other sulfur atoms and they are all in zero oxidation state. Despite the fact that sulfur has been known from ancient times, the study of peculiarities of its chemical behavior is still ongoing [57]. Elemental sulfur is considered rather chemically inert, but under wine conditions it can react with various components of wine. For example, sulfur can be attacked by nucleophiles, such as hydrogensulfite (HSO3−, sulfitolysis), thiols (thiolysis) and other HS-containing compounds (e.g., glutathione), due to the electrophilic nature of S-S bonds of elemental sulfur (Figure 2 and Figure 3) [22]. These reactions open the door for various types of sulfane sulfur compounds and, finally, for reductive aroma compounds before and after bottling via numerous side reactions [17,32,33,39,41,58,59]. For example, it has been demonstrated that residues of elemental sulfur can induce production of H2S not only during fermentation but also after bottle storage [32]. According to this study, elemental sulfur content above 1 mg/L might cause the later formation of H2S in concentrations detrimental to wine quality.
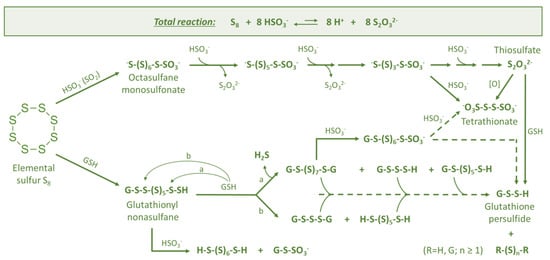
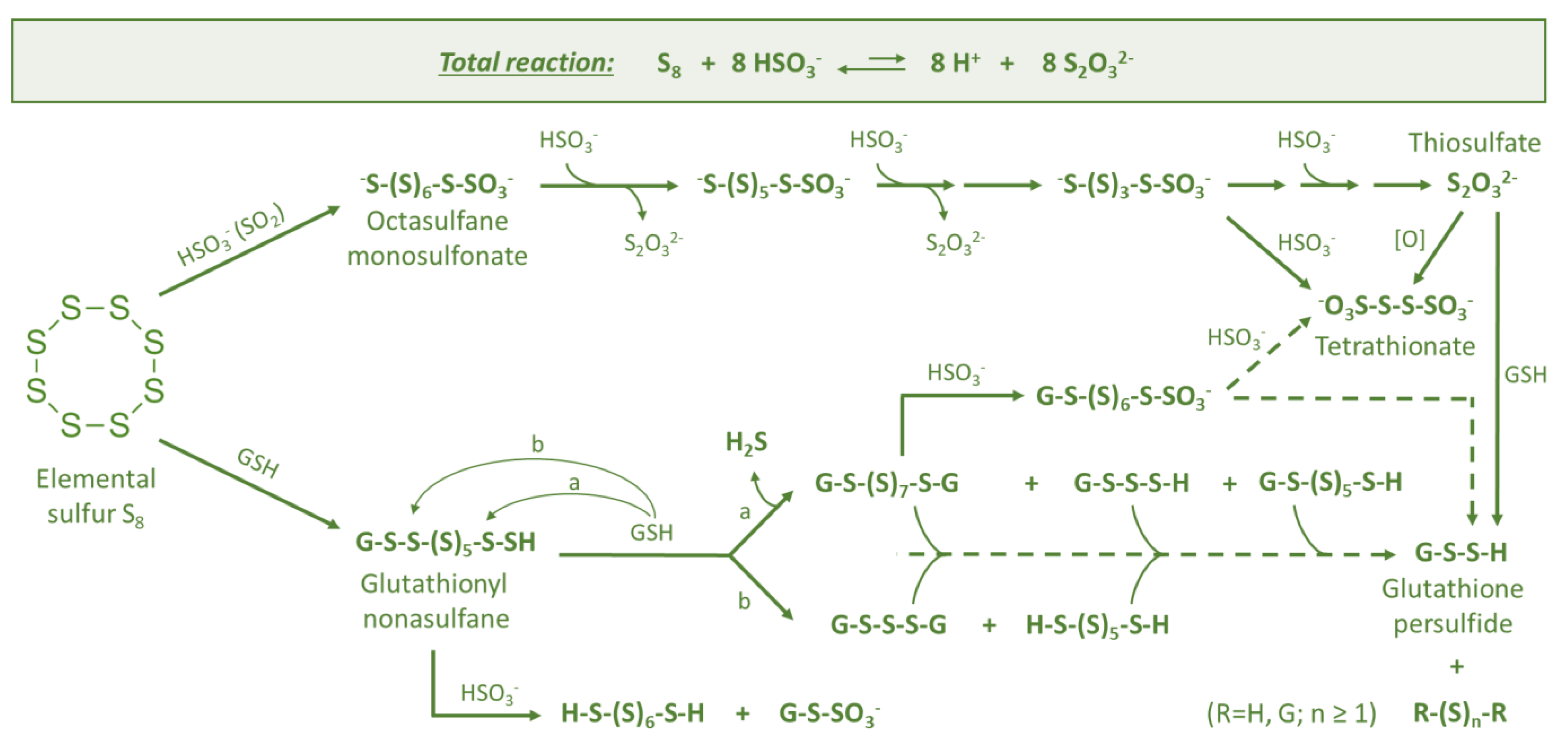
Figure 3.
Sulfane sulfur compounds formed due to sulfitolysis (reaction with hydrogensulfite HSO3−) and thiolysis (reaction with glutathione GSH) of elemental sulfur at wine pH: detailed chemical transformations for Figure 2, the part colored green (based on [32,33]).
Elemental sulfur is usually applied in vineyards to control powdery mildew. There are no comprehensive data about the typical content of elemental sulfur in grapes during the harvest, but it can reach 3 μg/g berry weight (equal to 3.4 mg/L of must) [60] and more up to 100 mg/L in a worst-case scenario [32,61]. It is worth noting that the addition of elemental sulfur prior to fermentation can increase the formation of not only H2S but also varietal thiols such as 3-MH [31].
The nature of the initial reaction between elemental sulfur (S8) and glutathione (GSH) is being discussed among scientists. While this reaction is chemically justified, some authors have questioned if it can occur under must/wine conditions due to the relatively low content of GSH, typically up to 100 mg/L [62]. Therefore, it was suggested that this initial reaction, which leads to the accumulation of sulfane sulfur precursors of reductive compounds, is rather enzymatic. In order to understand this biochemical process, we can go back to the earliest forms of life on Earth to find parallels with sulfur metabolism in Saccharomyces cerevisiae [63,64]. The early work of Rohwerder and Sand [41] on the sulfur-oxidizing organism Acidobacillus thiooxidans demonstrated that “biological” oxidation of elemental sulfur starts with the nucleophilic attack of glutathione on one of the sulfur atoms of cyclooctasulfur, leading to glutathionyl nonasulfane (GS-(S)7-SH) (Figure 3). This unstable primary product may further be stepwise degraded to persulfide (GSSH). The latter serves as a substrate for the glutathione-dependent sulfur dioxygenase (EC 1.13.11.18) enzyme, yielding sulfite as a product. Later, in Section 4, it is shown that Saccharomyces cerevisiae has the same enzyme at its disposal.
The degradation of glutathionyl nonasulfane (GS-(S)7-SH) in wine can occur as a result of reactions with other GSH molecules (originated from grapes and yeasts) producing various mono- and disubstituted polysulfanes and H2S (Figure 3). Glutathionyl nonasulfane (GS-(S)7-SH) and other intermediates can react also with hydrogensulfite (HSO3−) to form (poly)sulfane monosulfonates (R-Sn-SO3−) and polythionates (-O3S-Sn-SO3-).
Hydrogensulfite (HSO3−) is an important nucleophile in wine, derived from sulfur dioxide (SO2) [50,65]. It is added to wine at various winemaking stages at allowed concentrations of typically up to 150–400 mg/L. Moreover, even if no SO2 is added to must or wine, certain amounts of it are usually produced by yeasts [66]. It is well known that elemental sulfur reacts in an alkaline medium with nucleophilic reagents, like Na2SO3, with cleavage of the S8 ring followed by degradation of the sulfur chain. A similar reaction also takes place in acidic media, when hydrogensulfite is acting as a nucleophile [33]. The cleavage of the S8 ring in the first step results in the formation of octasulfane monosulfonic acid (Figure 3). Since the open chain sulfane intermediates are more reactive in nucleophilic substitutions, they are finally degraded to thiosulfate as presented in the total reaction (Figure 3). Similar reactions are expected to occur in real wines as well, since tetrathionate (a potential intermediate) was recently detected in experiments in which wine contained elemental sulfur. The formed tetrathionate can serve later as a precursor of reductive aromas. The same can occur also with sulfitolysis intermediates with different sulfur chain lengths, which can further react as nucleophiles or serve as substrates, yielding numerous products (Figure 2).
3.2. Thiosulfate (S2O32−)
Thiosulfuric acid H2S2O3 is not stable in aqueous solutions and, depending on the conditions, decomposes, producing elemental sulfur, SO2, H2S, H2SO4 and even polysulfanes (R-Sn-R(′)) and polythionates (−O3S-Sn-SO3−). Therefore, thiosulfuric acid can be obtained only by anhydrous methods [67]. In turn, thiosulfates as salts are stable, but only in neutral or alkaline solutions. Under acidic conditions, thiosulfate anion (S2O32−) disproportionates, yielding sulfur and hydrogensulfite (Figure 3, total reaction). To our knowledge, thiosulfate has not been detected in wine, but it may exist at least for a short period, for example, as a by-product of the reaction of elemental sulfur with hydrogensulfite (HSO3−) (Figure 2 and Figure 3).
Despite its simple structure, thiosulfate can serve as a precursor for many sulfane sulfur compounds and, therefore, is considered as one of the key components of sulfane sulfur chemistry. A complete description of thiosulfate chemistry is beyond the scope of this paper. An excellent review of thiosulfate’s chemical properties has been published by Miethe [68]. In wine conditions, thiosulfate is expected to be easily oxidized to tetrathionate chemically in the presence of Cu(II) or Fe(III) (similar to the oxidation in gold leaching solutions [69]) (Figure 3) or enzymatically by thiosulfate oxidase (EC 1.8.5.2) (more details in Section 4).
Considering microbiological aspects, it was shown that thiosulfate can be assimilated by Saccharomyces cerevisiae and used as a source of sulfur. In fact, it can serve as the only sulfur source in yeast metabolism, substituting sulfate in yeast nutrition [34,70]. Thiosulfate can also be energetically more preferable for yeasts than sulfate, since one of its sulfur atoms is in an oxidation state close to the target metabolic products, such as cysteine, methionine, H2S, etc. The enzyme rhodanese (EC 2.8.1.1) catalyzes the first step of thiosulfate metabolism and, together with its subclass “relative” thiosulfate sulfur transferase (TST), plays an important role in persulfidation reactions. The latter are the basis for the biological production of polysulfanes in wine (Section 4). It was also shown that thiosulfate can improve the growth of yeasts and their ethanol-production rate [70]. At the same time, the other study demonstrated that thiosulfate revealed stronger inhibitory properties than sulfite to Saccharomyces cerevisiae [35]. Most of the described experiments were performed at pH 6, while at pH < 4 thiosulfate decomposed to elemental sulfur and hydrogensulfite (HSO3−) (sulfurous acid).
3.3. Persulfides (R-S-S-H)
In terms of chemical structure, persulfides are monosubstituted derivatives of hydrogen disulfide (H2S2). Persulfides are quite reactive and participate in different chemical and biochemical transformations (including the formation of various polysulfanes (R-Sn-R(′))), which makes them interesting objects of study [24,27,29,53,71,72]. Thus, persulfides play an important role in biological systems, being intermediates in the synthesis of sulfur-containing biomolecules and are considered to mediate the signaling effects of H2S. Persulfides are often formed from thiols via different pathways, including the participation of H2S. These processes are described in detail in the recent review of Benchoam and colleagues [73]. Persulfides of cysteine and glutathione are being extensively studied in different organisms, mainly in mammal cells [20]. For example, it was demonstrated that persulfides of cysteine residues in proteins are important intermediates in the biological chemistry of H2S and its role in signaling processes, including in human beings [74]. Glutathione persulfide (GSSH) is produced by yeasts from thiosulfate and glutathione (GSH) by means of mitochondrial rhodanese Rdl1 (Figure 3). Then, GSSH spontaneously reacts with another GSH molecule, yielding glutathione disulfide (GSSG) and H2S [35,36]. In general, biochemical transformations of glutathione play an outstanding role in life processes. It is present not only in animal and fungi cells, but also in grapes.
Compared to thiols, persulfides are more nucleophilic and acidic. For example, the acidity of GSSH is much higher (pKa = 5.45) than that of glutathione (pKa = 8.94). As a result, glutathione persulfide reacts much faster in nucleophilic reactions with sulfane sulfur compounds [53]. In contrast to thiols, persulfides can also exhibit electrophilic properties, which makes them labile in aqueous systems and, hence, difficult to analyze [53]. Under wine conditions, persulfides are expected to react with many wine components, including various polysulfanes. Reductive aroma compounds, such as H2S, can be released directly from these reactions (Figure 2).
3.4. Polysulfanes (R-Sn-R(′))
Polysulfanes are intermediate sulfur species that are important for many environmentally and biologically relevant processes [63,64]. Polysulfanes are not exclusive to grapes and wine but are ubiquitous in many other organisms: plants [75,76], mammalian/human cells [77], yeasts [78], invertebrates [14], fungi [79], prokaryotes [80], etc. Laboratory experiments with microbial cultures and enzymes indicate both indirect and direct roles of microorganisms in the production and assimilation of polysulfanes. It is challenging to quantify polysulfanes in natural systems due to their reactivity, sensitivity to sample handling and the analytical complexity associated with their measurements.
The variability of polysulfanes makes them one of the most important precursors of reductive aroma compounds in wine. The simplest representatives are inorganic polysulfanes (H-(S)n-H), starting from hydrogen disulfide H2S2 (disulfane), which was detected in wine along with trisulfane (H2S3), and which were associated with flintstone aroma [38] (Table 1). Monosubstituted polysulfanes (R-Sn-H) can be formed in wine as intermediates, but they are difficult to identify due to their high reactivity. Recently, Pilkington and colleagues developed synthetic methods to produce authentic samples of cysteinyl (Cys-Sn-H) and glutathionyl hydropolysulfanes (G-Sn-H) (n = 2–4), as well as disubstituted polysulfanes (Cys-Sn-Cys) and (G-Sn-G, n = 2–5), and the approaches of their analysis [48]. Monosubstituted polysulfanes (R-Sn-H) can further react with various sulfhydryl compounds R(’)-SH resulting in disubstituted polysulfanes (R-Sn-R(′)).
Disubstituted polysulfanes can exist in symmetric (R-Sn-R) and asymmetric (R-(S)n-R’) forms (Table 1). Dialkyl polysulfanes (Me-S3-Me, Et-S3-Et, Me-S3-Et) and their formation in alcoholic drinks have been known for decades [46]. However, representatives with glutathionyl and cysteinyl groups have been found in wine only in recent years. For example, first diglutathionyl polysulfanes, GSSG and GSSSG, were detected in wine containing sulfur residues by Jastrzemsky and colleagues and these results were published in 2017 [32]. The mixtures of bigger diglutathionyl polysulfanes (G-(S)n-G, n = 2–7) and dicysteinyl polysulfanes (Cys-(S)n-Cys, n = 2–6) were observed in model wines due to the reactions of hydrogen sulfide with glutathione and cysteine catalyzed by Cu(II) [28,44]. In the presence of MeSH, mixed polysulfanes, Cys-Sn-Me (n = 2–6) and G-Sn-Me (n = 2–8), were also formed [44]. Recently, in 2020, a series of symmetric and asymmetric polysulfanes with cysteine and glutathione substituents were reported in real wines: G-Sn-G, Cys-Sn-Cys, G-Sn-Cys (n = 2–5) [45,49]. A number of symmetric and asymmetric polysulfanes with aroma thiols 3-MH and 4-MMP were also detected in model and real wines: 3MH-Sn-3MH, 4MMP-Sn-4MMP, 3MH-Sn-4MMP, Cys-Sn-3MH, Cys-Sn-4MMP, G-Sn-3MH, G-Sn-4MMP (n = 2–5) [43,81]. The formation of such disulfides (n = 2) was catalyzed by Cu(II) ions. Corresponding polysulfanes with n = 3–4 were obtained when elemental sulfur was added. The composition and ratio of polysulfanes differed in model and real wines, which should be related to the various stabilities and reactivities of these compounds [81]. In addition, the reactions of polysulfane formation can be affected by the matrix of real wines.
The mechanisms of polysulfane formation are currently being discussed. As described, they can be obtained chemically in wines and model aqueous solutions due to the reaction of elemental sulfur or H2S with thiols [28,44,46,81] (Figure 2). In both cases, Cu(II) ions catalyze these reactions, which are usually present in must and wine, and which can be added before bottling for copper fining. The enzymatic genesis of polysulfanes in wine is also plausible, which has been argued by several authors [20,22]. Then, Pilkington and colleagues observed the presence of cysteinyl and glutathionyl polysulfanes in Saccharomyces cerevisiae for the first time [48]. Therefore, the increase of polysulfane concentrations in wine can be expected due to yeast autolysis, as was shown for dimethyl trisulfide (DMTS) (Me-S3-Me) [47]. Studies of polysulfane formation in yeast cells are currently in progress.
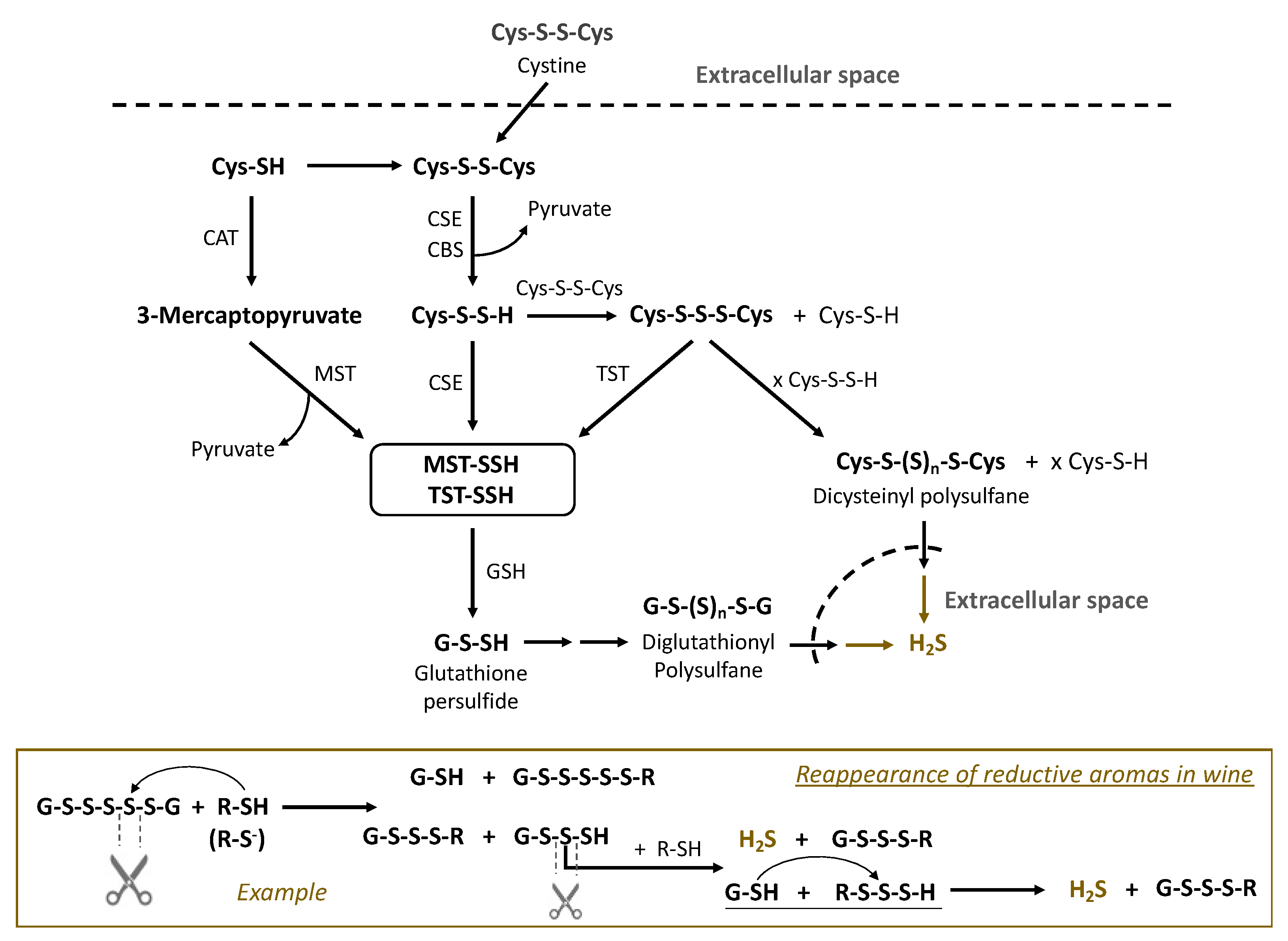
Sulfur metabolism in Saccharomyces cerevisiae is integrated in many processes, including transformations of sulfur amino acids and their derivatives [18]. Experiments of Huang explored whether sulfane sulfur compounds are involved in cysteine catabolism in Saccharomyces cerevisiae. The presented results suggested the contribution of cysteine persulfide (Cys-S-SH) to the formation of polysulfanes and their presence in wine [37]. The formation of polysulfanes and their role as latent precursors of reductive off-odors became part of the “polysulfide theory”. We assume that biological and chemical synthesis and conversions of polysulfanes occur in parallel during the winemaking process. As an example, the transformations of diglutathionyl, dicysteinyl and other polysulfanes is outlined in Figure 4. Cystine, a dimer of cysteine with a disulfide bond, is assimilated from the extracellular space, i.e., must/wine. The enzyme cystathionine-γ-lyase (CSE), which typically cleaves cystathionine into cysteine and pyruvate, catalyzes the formation of cysteine persulfide (CysSSH) from cystine and uses pyridoxal phosphate as a co-factor [82]. Transsulfidation with the formation of protein persulfides of 3-mercaptopyruvate sulfurtransferase (MST-SSH) or rhodanese (TST-SSH) is followed by further transsulfidation and leads to glutathione persulfide (GSSH). Both persulfides CysSSH and GSSH can react with corresponding disulfides to form trisulfides GSSSG and CySSSCys, which in subsequent reactions with persulfides result in elongation of the polysulfane chain (n > 4). The formed polysulfanes may undergo further nucleophlic substitutions, reacting with sulfur nucleophiles in wine, and finally release reductive aroma compounds, e.g., H2S (Figure 4).
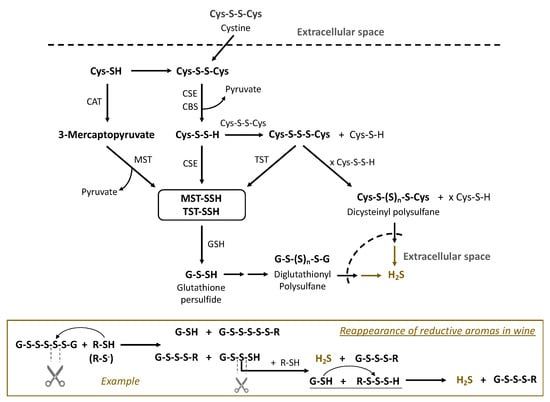
Figure 4.
Formation of polysulfanes in Saccharomyces cerevisiae and the enzymes involved in sulfur metabolism (based on [24,29]). CAT—cysteine aminotransferase; CSE—cystathionine-γ-lyase; MST—3-mercaptopyruvate sulfurtransferase; TST—thiosulfate sulfurtransferase (rhodanese); also involved is CBS: cystathionine-ß-synthase. MST-SSH and TST-SSH stand for the persulfides of the corresponding enzymes in Table 3. The lower block is an example of polysulfane reactions in wine releasing reductive aromas, e.g., H2S.
The reactivity of polysulfanes depends on their structure, in particular on the length of sulfur chains. Polysulfanes with three sulfur atoms are among the most stable [28,32], e.g., 84% of Cys-S3-Cys content in a model wine remained after six months. In contrast, the molecules with four and more sulfur atoms decomposed faster in model wine, losing about three quarters of the initial amount over the same half-year period [28]. Decomposition of polysulfanes can result in formation of starting compounds, in particular H2S. An example of such polysulfane tranformations in wine is presented in Figure 4 (lower block). HS-containing nucleophiles attack sulfane sulfur in the β-position of diglutathionyl pentasulfide (GSSSSSG). There are two possibilities of subsequent cleavage of S-S bonds in polysulfane, yielding two pairs of compounds. The mechanisms of these thiol–polysulfane exchange reactions in wine require more investigation. Some authors suggest that the SN2 mechanism can be realized in such reactions, starting from the formation of thiolate (R-S−), which attacks and cleaves polysulfanes. The priority for the leaving groups will depend on their pKa: a modest preference for the product with a lower pKa. In the example in Figure 4, the leaving group GSSS− will have more priority than GS− (pKa GSSS− < pKa GS−). Both of the products in acidic wine conditions will mostly exist as GSH and GSSSH. The latter can undergo subsequent thiol–polysulfane exchange reactions, releasing H2S.
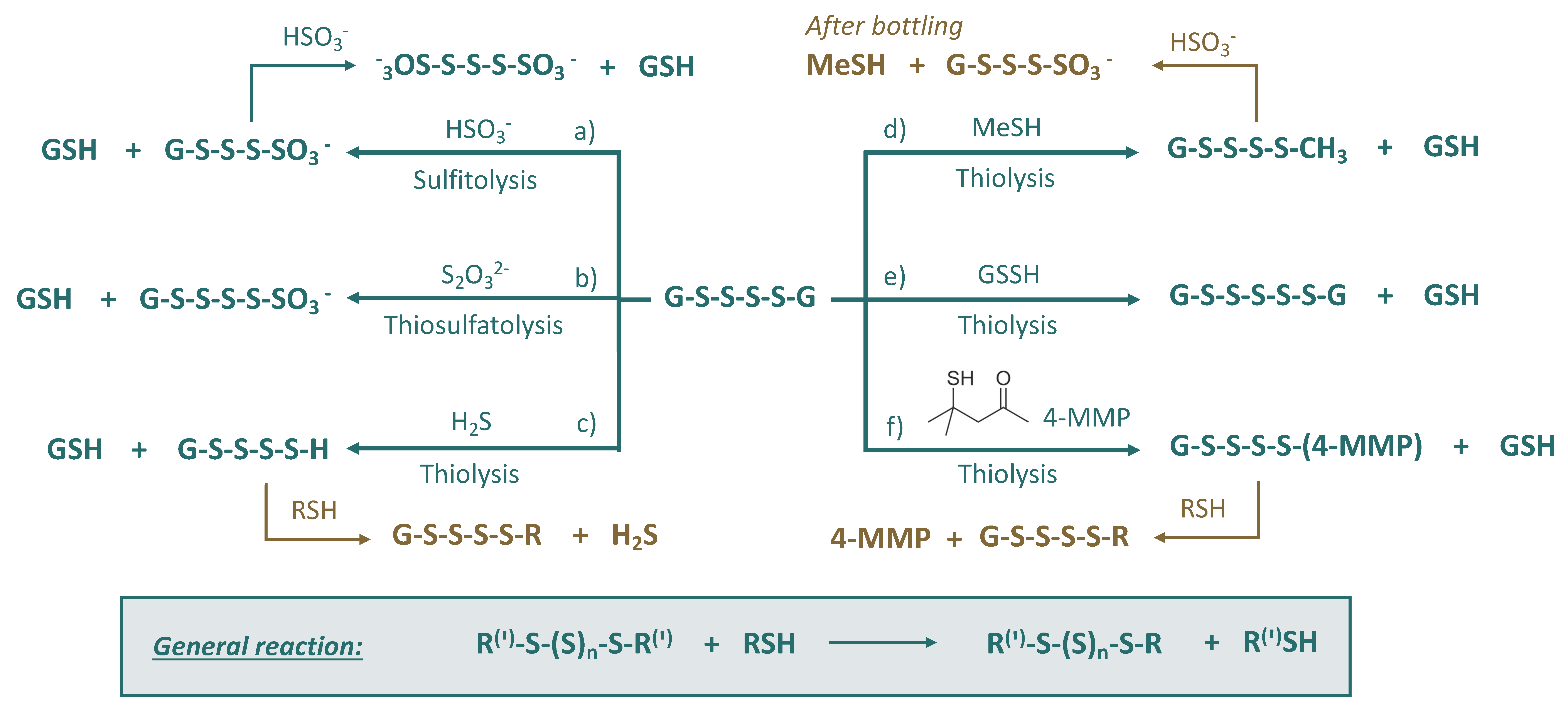
Besides R-SH nucleophiles, polysulfanes can be attacked by hydrogensulfite HSO3− (sulfitolysis) or thiosulfate S2O32− (thiosulfatolysis). It was demonstrated that the presence of sulfur dioxide (hydrogensulfite) was associated with the quick degradation of polysulfanes and consequent liberation of H2S [28]. Possible reactions of polysulfanes (on the example of diglutathionyl tetrasulfide (GSSSSG)) with various wine nucleophiles are presented in Figure 5. The role of these reactions can be summarized as follows:
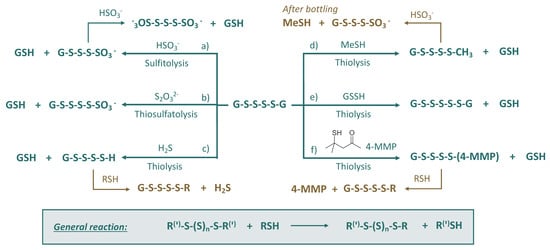
Figure 5.
Suggested reactions of diglutathionyl tetrasulfide with S-nucleophiles in wine: examples of chemical transformations for Figure 2, the part colored turquoise.
- (a)
- Sulfitolysis: formation of polysulfane monosulfonates and polythionates, which will be discussed in the next subsections;
- (b)
- Thiosulfatolysis: results in polysulfane monosulfonates with sulfur chain elongation;
- (c)
- Thiolysis by H2S: trapping of malodorous H2S, which can reappear later due to the subsequent reactions, e.g., with HS-containing nucleophiles;
- (d)
- Thiolysis by MeSH: trapping of malodorous MeSH, which can be released later due to the consequent reactions, e.g., sulfitolysis;
- (e)
- Thiolysis by glutathione persulfide: formation of polysulfanes with longer sulfur chains;
- (f)
- Thiolysis with 4-MMP: trapping of varieatal thiol 4-MMP, which can reappear later due to the subsequent reactions, e.g., with HS-containing nucleophiles.
3.5. Polythionates (−O3S-Sn-SO3−)
Polythionates, also known as polysulfane disulfonates, are sulfur-based oxyanions in which the sulfane sulfur chain is terminated at both ends with SO3− groups. These compounds are important intermediate species in the redox transformations of sulfur substances in various media and in the metabolism of sulfur-oxidizing and sulfur-reducing microorganisms [83,84,85]. Polythionates have long been known in the form of salts with the length of sulfur chains up to several dozens. However, the most common representatives have from one to four sulfane sulfur atoms. These salts are metastable in aqueous acidic solutions. The corresponding free acids H2SxO6 are unstable in the pure form [86].
In the past, polythionates were neglected due to their instability and unclear formation in complex mixtures. An example of such a mixture is “Wackenroder solution”, which is obtained by passing gaseous H2S through an aqueous solution of SO2. As a result of complex and not fully understood chemical reactions, the final solution contains various products, such as colloidal sulfur, polythionates (in particular with n = 1–3) and thiosulfate [13]. The presence of H2S and SO2 in wine (similar to “Wackenroder solution”, but at lower concentrations) makes the formation of polythionates under wine conditions highly probable (Figure 2).
The other potential source of polythionates in wine are thiosulfatolysis and sulfitolysis of polysulfanes (Figure 2, Figure 3 and Figure 5). The mechanism of the latter reaction may also involve thiosulfate S2O32−, which can be also formed due to the sulfitolysis of elemental sulfur [32]. In turn, the oxidation of thiosulfate can lead to the formation of tetrathionate (-O3S-S-S-SO3-) (Equation (1)) [87], which was recently detected in wine [32]. Jastrzembski and colleagues investigated the role of elemental sulfur in wine and suggested this pathway of tetrathionate formation [32]. In general, tetrathionate is one of the most stable polythionates at wine pH. Therefore, its detection in wine was not unexpected but became a very important part of the general theory of sulfane sulfur compounds in wine.
4 S2O32− + O2 + 4H+ = 2 S4O62− + H2O
Despite the fact that polythionates are metastable under acidic conditions, they are difficult to detect in wine for the following reasons. First, polythionates can easily react with nucleophiles and reducing agents, especially at higher pH values of wine. Second, oxidizers can convert polythionates to sulfate SO42−. The role of an oxidizing agent can be played, for example, by hydrogen peroxide H2O2, which is formed in wine due to the reactions of dissolved oxygen O2 with phenolic compounds, catalyzed by metal ions Fe(III) and Cu(II).
3.6. Polysulfane Monosulfonates (R-Sn-SO3−)
Polysulfane monosulfonates, in contrast to polythionates, have only one SO3− group at the end of the sulfane sulfur chain. The other side of the molecule is terminated with hydrogen or a different organic substituent (R). Polysulfane monosulfonates are unstable, easily subjected to nucleophilic reactions, e.g., sulfitolysis or thiosulfatolysis (Figure 2 and Figure 3), and play an outstanding role as intermediates in sulfane sulfur chemistry.
Among (poly)sulfane monosulfonates that have been already detected in wine are S-sulfonated derivatives of glutathione, cysteine and pantetheine: G-S-SO3−, Cys-S-SO3−, Pan-S-SO3− [28,32,39,50]. They are suggested to be obtained due to the sulfitolysis of corresponding sulfhydryl compounds, such as amino acids and peptides. In the case of glutathionyl sulfane monosulfonate (G-S-SO3−), the sulfitolysis of glutathione disulfide (G-S-S-G) is assumed [50]. Polysulfane monosulfonates with longer sulfane sulfur chains were also determined under wine conditions, e.g., Cys-S2-SO3−, Cys-S3-SO3−, G-S2-SO3−, G-S3-SO3− [28,32,39]. Other reactions leading to polysulfane monosulfonates are sulfitolysis of elemental sulfur (Figure 3) or various polysulfanes (Figure 2 and Figure 5).
Polysulfane monosulfonates may be subjected to further sulfitolysis or thiosulfatolysis, yielding polythionates. Then, both families of compounds can react with various nucleophiles in wine, producing directly reductive aromas, H2S (Figure 2).
4. Biosynthesis of Sulfane Sulfur Compounds by Microorganisms during Fermentation and the Enzymes Involved
In the previous sections we discussed chemical pathways of sulfane sulfur compound formation in wine and mentioned that many of these substances can also be formed enzymatically by microorganisms. Thus, polysulfanes may be originated from enzymatically produced persulfides and disulfides [53,71]. Both persulfides and polysulfanes are formed in noticeable amounts during sulfur-containing amino acid metabolism and possess important chemical and biological properties. Some of these sulfane sulfur compounds have also been suggested to exist endogenously in mammalian systems. Reports also indicated that a persulfide moiety (RSSH) may be formed on the cysteine (Cys) residues of specific proteins, mostly in the presence of sulfur-transferring enzymes, i.e., sulfurtransferases (similar to what is shown in Figure 4) [29]. These enzymes also play an important role in the sulfur metabolism of Saccharomyces yeasts, after the sulfate reduction pathway (SRP), followed by biosynthesis of cysteine and methionine, resulting in similar reactions yielding polysulfanes.
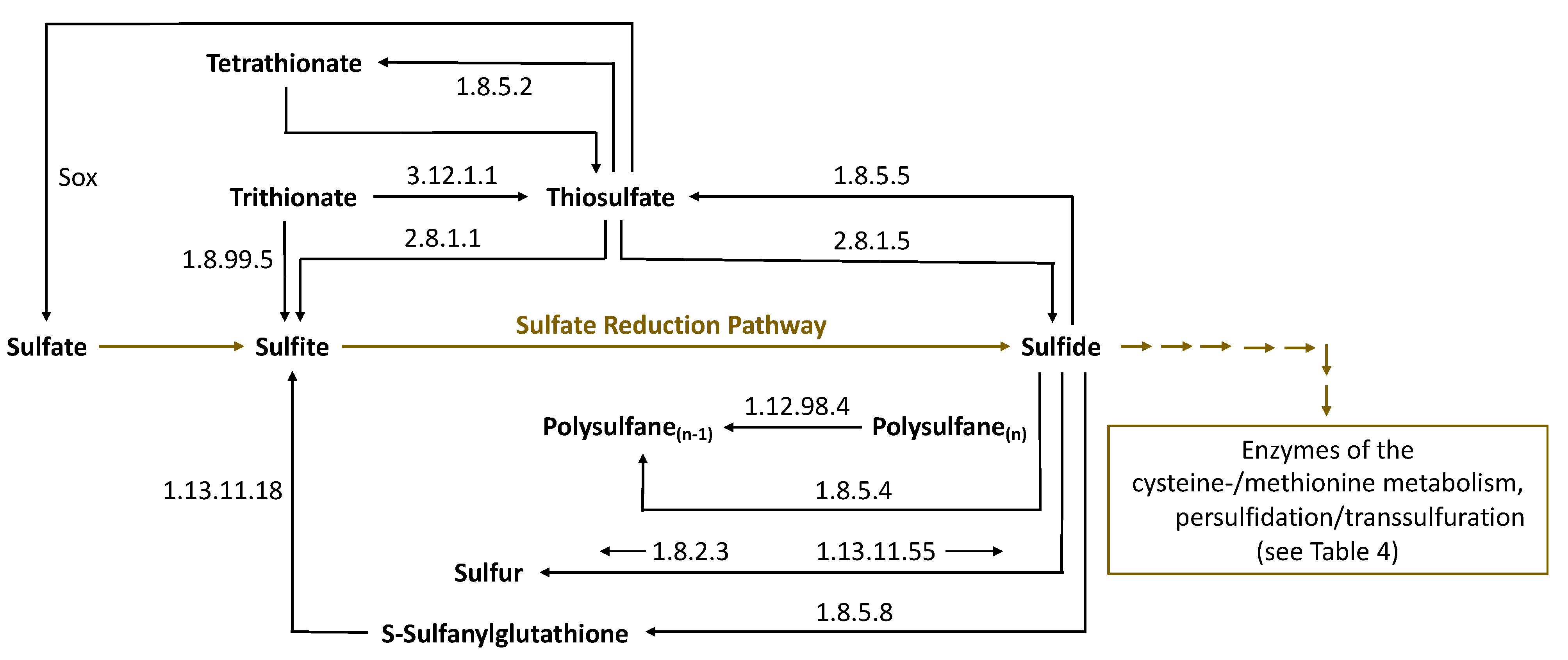
The detection of polysulfanes in Saccharomyces cerevisiae [48] gives rise to the suspicion that enzymes of sulfur metabolism could also act via persulfidation in their formation, as indicated in Figure 4. Indeed, the enzymes 3-mercaptopyruvate sulfur transferase (MST) and thiosulfate sulfurtransferase (TST, rhodanese), among others (Table 3), may perform this function. The enzymes involved in the formation of polysulfanes belong to the most abundant biocatalysts responsible for the metabolism of sulfur compounds in Saccharomyces cerevisiae. A closer look at the sulfur metabolism of the BRENDA (BRaunschweig ENzyme DAtabase) [88] and KEGG (Kyoto Encyclopedia of Genes and Genomes) [89] databanks helps to clarify actual questions related to sulfane sulfur substances. Thus, many sulfane sulfur compounds can be found there, together with the corresponding enzymes related to their formation and further metabolism. In particular, Figure 6 provides a detailed section from the sulfur metabolism of Saccharomyces cerevisiae in the KEGG databank concerning sulfane sulfur compounds. The corresponding enzymes are listed below in Table 3 and Table 4.
Table 3.
Enzymes involved in cysteine/methionine metabolism, persulfidation/transsulfuration.
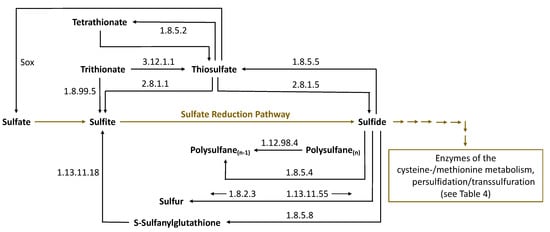
Figure 6.
Sulfane sulfur compounds in the sulfur metabolism of Saccharomyces cerevisiae (including the enzymes involved), based on BRENDA [88] and KEGG [89] (Sox—sulfur oxidation enzyme system).
Table 4.
Enzymes for sulfur metabolism involved in the transformation of sulfane sulfur compounds (selected from KEGG [89] (sulfur metabolism of Saccharomyces cerevisiae)).
The second major player in the winemaking process, lactic acid bacteria, operates with similar enzymatic equipment to Saccharomyces cerevisiae in sulfur metabolic processes [17,97,98,99]. Therefore, it may be expected that these bacteria have a similar role in the formation of sulfane sulfur compounds and that, consequently, there will be an intensification of research activities in this area in the future. In addition, the impact on the formation of volatile sulfur compounds of non-conventional yeasts, which are becoming more of a focus in the winemaking process, should be also considered and studied in the future [100,101]. The more we advance in these studies, the clearer it becomes that the sulfur metabolism in microorganisms involved in wine fermentation leads to a myriad of reactive sulfane sulfur products and intermediates, which can be released into wine and serve as latent precursors of reductive compounds in the post-bottling period.
5. Proteins with Various Sulfane Sulfur Groups
Proteins can be found in wine from different sources: grapes, yeast and lactic acid bacteria metabolism, wine aging on lees, the protein fining process, etc. [102,103]. In the later steps of winemaking the protein content in wine can decrease, especially if the wine is treated with bentonite. Due to the varying content of proteins in wines, usually from 15 to 230 mg/L [102], it is difficult to speak about their impact on the post-bottling development of reductive defects. Nevertheless, these proteins can be degraded and release an additional amount of H2S and its derivatives.
Protein persulfides in general play an important role in sulfane sulfur chemistry. Some renowned papers on this topic were published more than 30 years ago, indicating their general importance in biological systems [26,104]. The method utilized by Mustafa et al. detected protein persulfides in the range of 10–25% of all cysteine residues in liver cells [105]. Later, the levels of persulfides in cells and tissues of 1–12 μg/mg were confirmed [106] by a novel detection method.
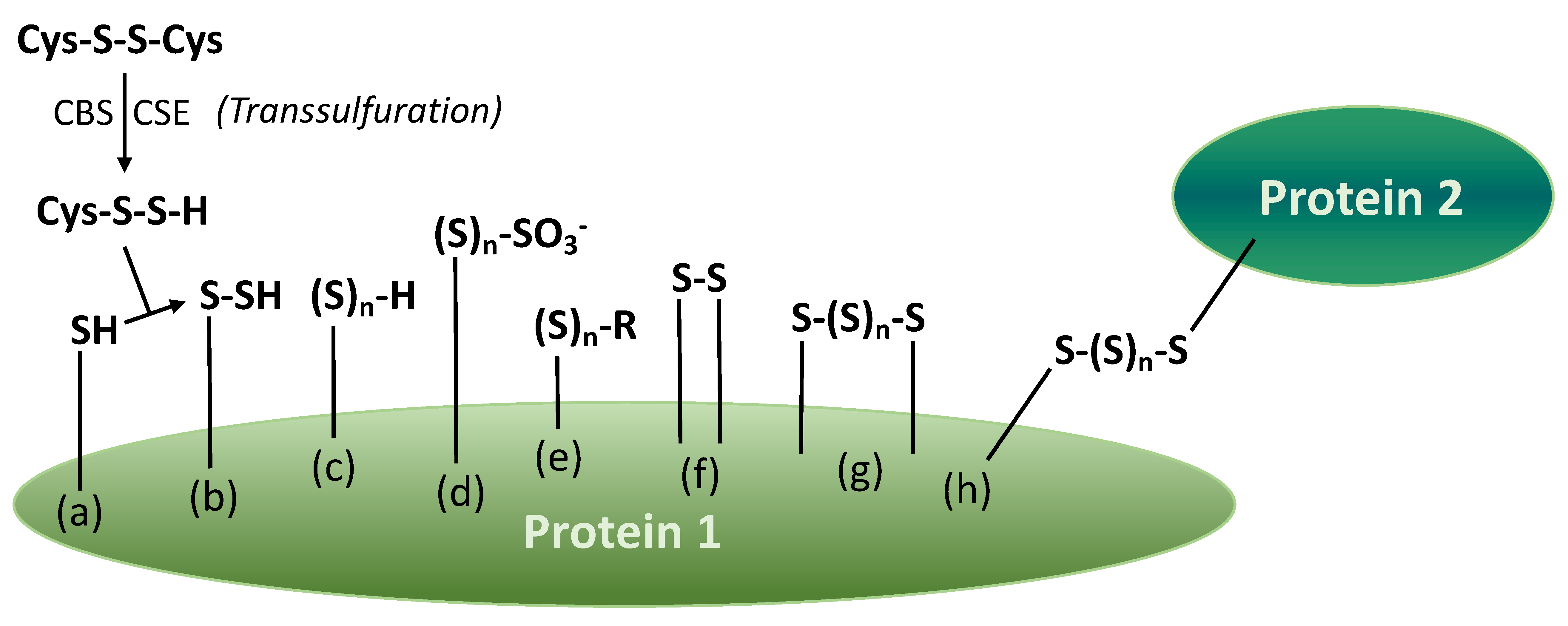
Persulfides, polysulfane monosulfonates and various polysulfane fragments can be present in the structure of proteins in wine. Certain fragments are originated from enzymatic reactions, e.g., a simplified pathway of the persulfidation process is shown in Figure 7a,b [27]. The others are formed due to a number of chemical transformations in must and wine. For example, Cu(II) ions may catalyze the formation of disulfide and polysulfane fragments (Figure 7f–h). In general, many reactions with sulfur nucleophiles (Figure 1 and Figure 5) may take place with protein persulfides and polysulfanes, including those generating varietal thiols and reductive compounds, H2S, in the post-bottling period.
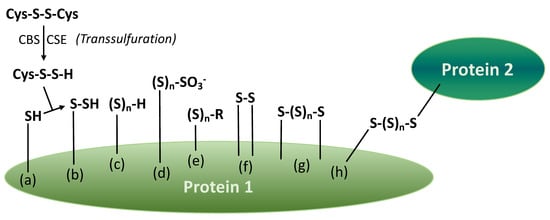
Figure 7.
Products of protein (enzyme) transsulfuration and consecutive reactions (based on [27]). Legends: (a) cysteine in protein 1 (transsulfuration by cysteine persulfide); (b) protein persulfide; (c) protein monosubstituted polysulfane; (d) protein polysulfane monosulfonic acid; (e) protein polysulfane (R = Me, Et, G, Cys-, 3-MH-, 4-MMP-, other “wine thiols”); (f) internal disulfide bridge; (g) internal polysulfane bridge; (h) polysulfane bridge between two proteins.
6. Final Remarks
As outlined in this review, the reappearance of reductive off-odors traces back to an extremely complex interplay between different factors in wine and during fermentation: the choice of yeast starter cultures and nutrient supports, the pH value of wine, the presence and nature of precursors, nucleophiles, temperature, redox potential, etc. It has been shown that well-known approaches for removing or preventing reductive off-odors (caused by H2S or other low volatile sulfur compounds, e.g., MeSH, EtSH, MeSSMe, etc.) often have only a temporary effect. Moreover, they can cause even more problems later due to the reappearance of reductive faults after bottling due to the formation and accumulation of corresponding precursors. Among these known methods are wine treatment with copper or silver salts, macro-oxygenation, addition of ascorbic acid, lees from a donor wine, diammonium phosphate (DAP) as a yeast nutrient, etc. [45,107]. The complexity of the reactions described in this review demonstrates why it is difficult to recommend remediation strategies to remove or avoid reductive off-odors in wine. However, this complexity should not preclude the study of sulfane sulfur compounds as precursors, as this can help to deal with the problem of reductive aromas in wine.
The determination of possible latent sources of H2S and other causes of reductive aromas would help to eliminate an increasing problem for the wine industry [108]. However, so far, many attempts to achieve a total balance of all sulfur compounds in wine have failed [109]. Thus, in the cited publication, the sum of the identified sulfur species in wine accounted only for 65–77% of the total sulfur concentration, which indicates the existence of many unknown compounds. Moreover, comparing the composition of sulfite-treated wine with an untreated one, a dataset of more than 11,000 new sulfur-containing compounds was found [110]. This goes along with our knowledge about numerous reaction pathways of wine nucleophiles with all the sulfane sulfur compounds dealt with in this review.
7. Conclusions
In this review, it was shown how sulfane sulfur compounds can act as latent precursors of reductive off-odors in wine, such as H2S and its derivatives, MeSH, EtSH, etc. It was clarified that the formation of sulfane sulfur precursors results from a mixture of chemical and enzymatic reactions. The key role of microorganisms, in particular yeasts (Saccharomyces cerevisiae), was discussed and corresponding metabolic pathways, including persulfides as intermediates, were presented. Elemental sulfur from fungicide residues and H2S, MeSH, etc., generated by yeasts can be among the main components that are the basis for the formation of latent precursors of reductive aroma compounds during the winemaking process. It can be considered that generated H2S, MeSH and other reductive aroma compounds become trapped in the structure of these precursors. Polysulfanes (R-Sn-R(′)) were identified as one of the most variable and important sulfane sulfur precursors. Based on the available literature and general chemistry concepts, we summarized possible transformation pathways of polysulfanes in wine, in the post-bottling period in particular. The presented figures show how these precursors can be attacked by various nucleophiles, specifically sulfhydryl (HS-containing) compounds and hydrogensulfite (HSO3−), which is usually available in wine in relatively high amounts. The products of these reactions can be other sulfane sulfur compounds and reductive aroma substances (H2S, MeSH, etc.). Finally, it was demonstrated that sulfane sulfur precursors can be in a free form or bound to proteins.
The more light is shed on the processes responsible for the reappearance of reductive aromas in wine, the more wine appears as a complex system and more new questions are raised. Sulfur compounds remain some of the most crucial and intriguing components for life on Earth, as well as for understanding wine.
Author Contributions
Conceptualization, investigation, N.M.; resources, funding acquisition, D.R.; project administration, visualization, A.T.; writing—original draft, review and editing, N.M., D.R. and A.T. All authors have read and agreed to the published version of the manuscript.
Funding
We acknowledge support by the Open Access Publishing Fund of Geisenheim University.
Institutional Review Board Statement
Not applicable.
Acknowledgments
The authors thank Tatyana Felyust for preparation of the graphical abstract.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mestres, M.; Busto, O.; Guasch, J. Analysis of organic sulfur compounds in wine aroma. J. Chromatogr. A 2000, 881, 569–581. [Google Scholar] [CrossRef]
- Siebert, T.; Bramley, B.; Solomon, M. Technical notes. Hydrogen sulfide: Aroma detection threshold study in white and red wine. Tech. Rev. 2009, 183, 14–16. [Google Scholar]
- Siebert, T.E.; Solomon, M.R.; Pollnitz, A.P.; Jeffery, D.W. Selective Determination of Volatile Sulfur Compounds in Wine by Gas Chromatography with Sulfur Chemiluminescence Detection. J. Agric. Food Chem. 2010, 58, 9454–9462. [Google Scholar] [CrossRef] [PubMed]
- Goniak, O.J.; Noble, A.C. Sensory Study of Selected Volatile Sulfur Compounds in White Wine. Am. J. Enol. Vitic. 1987, 38, 223–227. [Google Scholar]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Fracassetti, D.; Vigentini, I. Occurrence and Analysis of Sulfur Compounds in Wine. In Grapes and Wines; Jordão, A.M., Cosme, F., Eds.; IntechOpen: Rijeka, Croatia, 2018. [Google Scholar]
- Tarasov, A.; Garzelli, F.; Schuessler, C.; Fritsch, S.; Loisel, C.; Pons, A.; Patz, C.-D.; Rauhut, D.; Jung, R. Wine Storage at Cellar vs. Room Conditions: Changes in the Aroma Composition of Riesling Wine. Molecules 2021, 26, 6256. [Google Scholar] [CrossRef]
- Rauhut, D.; Kürbel, H.; MacNamara, K.; Grossmann, M. Headspace GC-SCD monitoring of low volatile sulfur compounds during fermentation and in wine. Analusis 1998, 26, 142–144. [Google Scholar] [CrossRef] [Green Version]
- Rauhut, D. Impact of volatile sulfur compounds on wine quality. In Sulfur Transport and Assimilation in Plants; Davidian, J.-C., Grill, D., De Kok, L.J., Stulen, I., Hawkesford, M.J., Schnug, E., Rennenberg, H., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 121–131. [Google Scholar]
- Rauhut, D.; Kürbel, H.; Fischer, S.; Beisert, B. Simultaneous analysis of N- and S-compounds as screening for the identification of off-flavours in wine. In Recent Highlights in Flavor Chemistry and Biology; Hofmann, T., Meyerhof, W., Schieberle, P., Eds.; Deutsche Forschungsanstalt für Lebensmittelchemie: Garching, Germany, 2007; pp. 314–317. [Google Scholar]
- Rauhut, D.; Beisert, B.; Berres, M.; Gawron-Scibek, M.; Kürbel, H. Pulse flame photometric detection: An innovative technique to analyse volatile sulfur compounds in wine and other beverages. In State of the Art in flavour Chemistry and Biology; Hofmann, T., Rothe, M., Schieberle, P., Eds.; Deutsche Forschungsanstalt für Lebensmittelchemie: Garching, Germany, 2005; pp. 363–368. [Google Scholar]
- Goode, J.; Harrop, S. Wine faults and their prevalence: Data from the world’s largest blind tasting. In Proceedings of the 20th Entretiens Scientifiques Lallemand, Horsens, Denmark, 15 May 2008; pp. 7–9. [Google Scholar]
- Müller, N.; Rauhut, D. Recent Developments on the Origin and Nature of Reductive Sulfurous Off-Odours in Wine. Fermentation 2018, 4, 62. [Google Scholar] [CrossRef] [Green Version]
- Hildebrandt, T.M.; Grieshaber, M.K. Three enzymatic activities catalyze the oxidation of sulfide to thiosulfate in mammalian and invertebrate mitochondria. FEBS J. 2008, 275, 3352–3361. [Google Scholar] [CrossRef]
- Jiang, G.; Li, M.; Wen, Y.; Zeng, W.; Zhao, Q.; Chen, C.; Yuan, H.; Liu, C.; Liu, C. Visualization of Sulfane Sulfur in Plants with a Near-Infrared Fluorescent Probe. ACS Sens. 2019, 4, 434–440. [Google Scholar] [CrossRef]
- Koike, S.; Ogasawara, Y. Sulfur Atom in its Bound State Is a Unique Element Involved in Physiological Functions in Mammals. Molecules 2016, 21, 1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauhut, D. Usage and Formation of Sulphur Compounds. In Biology of Microorganisms on Grapes, in Must and in Wine; König, H., Unden, G., Fröhlich, J., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 255–291. ISBN 978-3-319-60021-5. [Google Scholar]
- Thomas, D.; Surdin-Kerjan, Y. Metabolism of sulfur amino acids in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 1997, 61, 503–532. [Google Scholar] [CrossRef] [PubMed]
- Allison, R.B.; Sacks, G.L. Brine-Releasable Hydrogen Sulfide in Wine: Mechanism of Release from Copper Complexes and Effects of Glutathione. J. Agric. Food Chem. 2021, 69, 13164–13172. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-W.; Walker, M.E.; Fedrizzi, B.; Gardner, R.C.; Jiranek, V. Hydrogen sulfide and its roles in Saccharomyces cerevisiae in a winemaking context. FEMS Yeast Res. 2017, 17, fox058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vela, E.; Hernández-Orte, P.; Franco-Luesma, E.; Ferreira, V. The effects of copper fining on the wine content in sulfur off-odors and on their evolution during accelerated anoxic storage. Food Chem. 2017, 231, 212–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreitman, G.Y.; Elias, R.J.; Jeffery, D.W.; Sacks, G.L. Loss and formation of malodorous volatile sulfhydryl compounds during wine storage. Crit. Rev. Food Sci. Nutr. 2019, 59, 1728–1752. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.L. Sulfane sulfur. In Methods in Enzymology; Sulfur and Sulfur Amino Acids; Academic Press: Cambridge, MA, USA, 1987; Volume 143, pp. 25–29. [Google Scholar]
- Iciek, M.; Bilska-Wilkosz, A.; Górny, M. Sulfane sulfur—New findings on an old topic. Acta Biochim. Pol. 2019, 66, 533–544. [Google Scholar] [CrossRef]
- Westley, J.; Adler, H.; Westley, L.; Nishida, C. The sulfurtransferases. Fundam. Appl. Toxicol. 1983, 3, 377–382. [Google Scholar] [CrossRef]
- Toohey, J.I. Sulfur signaling: Is the agent sulfide or sulfane? Anal. Biochem. 2011, 413, 1–7. [Google Scholar] [CrossRef]
- Fukuto, J.M.; Ignarro, L.J.; Nagy, P.; Wink, D.A.; Kevil, C.G.; Feelisch, M.; Cortese-Krott, M.M.; Bianco, C.L.; Kumagai, Y.; Hobbs, A.J.; et al. Biological hydropersulfides and related polysulfides—A new concept and perspective in redox biology. FEBS Lett. 2018, 592, 2140–2152. [Google Scholar] [CrossRef] [Green Version]
- Bekker, M.Z.; Kreitman, G.Y.; Jeffery, D.W.; Danilewicz, J.C. Liberation of Hydrogen Sulfide from Dicysteinyl Polysulfanes in Model Wine. J. Agric. Food Chem. 2018, 66, 13483–13491. [Google Scholar] [CrossRef] [PubMed]
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive cysteine persulfides and S-polythiolation regulate oxidative stress and redox signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7606–7611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wróbel, M.; Lewandowska, I.; Bronowicka-Adamska, P.; Paszewski, A. The level of sulfane sulfur in the fungus Aspergillus nidulans wild type and mutant strains. Amino Acids 2008, 37, 565. [Google Scholar] [CrossRef] [PubMed]
- Araujo, L.D.; Vannevel, S.; Buica, A.; Callerot, S.; Fedrizzi, B.; Kilmartin, P.A.; du Toit, W.J. Indications of the prominent role of elemental sulfur in the formation of the varietal thiol 3-mercaptohexanol in Sauvignon blanc wine. Food Res. Int. 2017, 98, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Jastrzembski, J.A.; Allison, R.B.; Friedberg, E.; Sacks, G.L. Role of Elemental Sulfur in Forming Latent Precursors of H2S in Wine. J. Agric. Food Chem. 2017, 65, 10542–10549. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Heinrich, H. Reaktion von schwefliger Säure mit elementarem Schwefel. Über Säuren des Schwefels, XI. Angew. Chem. 1958, 70, 572. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, X.; Li, H.; Liu, H.; Xia, Y.; Xun, L. The Complete Pathway for Thiosulfate Utilization in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2018, 84, e01241-18. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Xia, Y.; Liu, H.; Liu, H.; Xun, L. The Mechanisms of Thiosulfate Toxicity against Saccharomyces cerevisiae. Antioxidants 2021, 10, 646. [Google Scholar] [CrossRef]
- Melideo, S.L.; Jackson, M.R.; Jorns, M.S. Biosynthesis of a Central Intermediate in Hydrogen Sulfide Metabolism by a Novel Human Sulfurtransferase and Its Yeast Ortholog. Biochemistry 2014, 53, 4739–4753. [Google Scholar] [CrossRef]
- Huang, C.-W. Identification of Yeast Genes Affecting Production of Hydrogen Sulfide and Volatile Thiols from Cysteine Treatment during Fermentation. Ph.D. Thesis, University of Adelaide, Adelaide, Australia, 2017. [Google Scholar]
- Starkenmann, C.; Chappuis, C.J.-F.; Niclass, Y.; Deneulin, P. Identification of Hydrogen Disulfanes and Hydrogen Trisulfanes in H2S Bottle, in Flint, and in Dry Mineral White Wine. J. Agric. Food Chem. 2016, 64, 9033–9040. [Google Scholar] [CrossRef]
- Jastrzembski, J.A. Mass Spectrometry for the High-Throughput Quantification and Mechanistic Investigation of Odor-Active Volatiles in Grapes and Wine. Ph.D. Thesis, Cornell University, Ithaca, NY, USA, 2017. [Google Scholar] [CrossRef]
- Kumar, M.R.; Farmer, P.J. Characterization of Polysulfides, Polysulfanes, and Other Unique Species in the Reaction between GSNO and H2S. Molecules 2019, 24, 3090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohwerder, T.; Sand, W. The sulfane sulfur of persulfides is the actual substrate of the sulfur-oxidizing enzymes from Acidithiobacillus and Acidiphilium spp. Microbiol. Read. Engl. 2003, 149, 1699–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobet, R.A.; Noble, A.C.; Boulton, R.B. Kinetics of the ethanethiol and diethyl disulfide interconversion in wine-like solutions. J. Agric. Food Chem. 1990, 38, 449–452. [Google Scholar] [CrossRef]
- Kreitman, G.Y.; Danilewicz, J.C.; Jeffery, D.W.; Elias, R.J. Reaction Mechanisms of Metals with Hydrogen Sulfide and Thiols in Model Wine. Part 2: Iron- and Copper-Catalyzed Oxidation. J. Agric. Food Chem. 2016, 64, 4105–4113. [Google Scholar] [CrossRef]
- Kreitman, G.Y.; Danilewicz, J.C.; Jeffery, D.W.; Elias, R.J. Copper(II)-Mediated Hydrogen Sulfide and Thiol Oxidation to Disulfides and Organic Polysulfanes and Their Reductive Cleavage in Wine: Mechanistic Elucidation and Potential Applications. J. Agric. Food Chem. 2017, 65, 2564–2571. [Google Scholar] [CrossRef]
- Nardin, T.; Larcher, R.; van Leeuwen, K.A.; Pilkington, L.I.; Roman, T.; Malacarne, M.; Fedrizzi, B. Effect of antioxidant supplementation on the polysulfides of white wines. LWT 2020, 134, 110132. [Google Scholar] [CrossRef]
- Nedjma, M.; Hoffmann, N. Hydrogen Sulfide Reactivity with Thiols in the Presence of Copper(II) in Hydroalcoholic Solutions or Cognac Brandies: Formation of Symmetrical and Unsymmetrical Dialkyl Trisulfides. J. Agric. Food Chem. 1996, 44, 3935–3938. [Google Scholar] [CrossRef]
- Nishibori, N.; Kuroda, A.; Yamada, O.; Goto-Yamamoto, N. Factors Affecting Dimethyl Trisulfide Formation in Wine. Food Sci. Technol. Res. 2017, 23, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Pilkington, L.I.; Deed, R.C.; Parish-Virtue, K.; Huang, C.-W.; Walker, M.E.; Jiranek, V.; Barker, D.; Fedrizzi, B. Iterative synthetic strategies and gene deletant experiments enable the first identification of polysulfides in Saccharomyces cerevisiae. Chem. Commun. 2019, 55, 8868–8871. [Google Scholar] [CrossRef]
- van Leeuwen, K.A.; Nardin, T.; Barker, D.; Fedrizzi, B.; Nicolini, G.; Larcher, R. A novel LC-HRMS method reveals cysteinyl and glutathionyl polysulfides in wine. Talanta 2020, 218, 121105. [Google Scholar] [CrossRef]
- Arapitsas, P.; Ugliano, M.; Perenzoni, D.; Angeli, A.; Pangrazzi, P.; Mattivi, F. Wine metabolomics reveals new sulfonated products in bottled white wines, promoted by small amounts of oxygen. J. Chromatogr. A 2016, 1429, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Dekker, S.; Fedrizzi, B.; van Leeuwen, K.A.; Nardin, T.; Dell’Anna, C.; Larcher, R. Time course accumulation of polysulfides in Chardonnay and model juice fermentations. Food Chem. 2022, 371, 131341. [Google Scholar] [CrossRef] [PubMed]
- Hallinan, C.P.; Saul, D.J.; Jiranek, V. Differential utilisation of sulfur compounds for H2S liberation by nitrogen-starved wine yeasts. Aust. J. Grape Wine Res. 1999, 5, 82–90. [Google Scholar] [CrossRef]
- Benchoam, D.; Semelak, J.A.; Cuevasanta, E.; Mastrogiovanni, M.; Grassano, J.S.; Ferrer-Sueta, G.; Zeida, A.; Trujillo, M.; Möller, M.N.; Estrin, D.A.; et al. Acidity and nucleophilic reactivity of glutathione persulfide. J. Biol. Chem. 2020, 295, 15466–15481. [Google Scholar] [CrossRef]
- Dupraz, S.; Ménez, B.; Guyot, F. Fast Determination of the Main Reduced Sulfur Species in Aquatic Systems by a Direct and Second-Derivative Spectrophotometric Method. J. Chem. 2019, 2019, e1039487. [Google Scholar] [CrossRef] [Green Version]
- Nikolantonaki, M.; Waterhouse, A.L. A Method To Quantify Quinone Reaction Rates with Wine Relevant Nucleophiles: A Key to the Understanding of Oxidative Loss of Varietal Thiols. J. Agric. Food Chem. 2012, 60, 8484–8491. [Google Scholar] [CrossRef]
- Steudel, R.; Eckert, B. Solid Sulfur AllotropesSulfur Allotropes. In Elemental Sulfur and Sulfur-Rich Compounds I; Steudel, R., Ed.; Topics in Current Chemistry; Springer: Berlin/Heidelberg, Germany, 2003; pp. 1–80. ISBN 978-3-540-44855-6. [Google Scholar]
- Kafantaris, C. On the Reactivity of Nanoparticulate Elemental Sulfur: Experimentation and Field Observations. Ph.D. Thesis, Indiana Univerity, Bloomington, IN, USA, 2017. [Google Scholar]
- Rauhut, D.; Kürbel, H. The production from H2S from elemental sulfur residues during fermentation and its influence on the formation of sulfur metabolites causing off-flavours in wine. Vitic. Enol. Sci 1994, 49, 27–36. [Google Scholar]
- Schütz, M.; Kunkee, R.E. Formation of Hydrogen Sulfide from Elemental Sulfur during Fermentation by Wine Yeast. Am. J. Enol. Vitic. 1977, 28, 137–144. [Google Scholar]
- Thomas, C.S.; Gubler, W.D.; Silacci, M.W.; Miller, R. Changes in Elemental Sulfur Residues on Pinot noir and Cabernet Sauvignon Grape Berries during the Growing Season. Am. J. Enol. Vitic. 1993, 44, 205. [Google Scholar]
- Kwasniewski, M.T.; Sacks, G.L.; Wilcox, W.F. Persistence of Elemental Sulfur Spray Residue on Grapes during Ripening and Vinification. Am. J. Enol. Vitic. 2014, 65, 453. [Google Scholar] [CrossRef] [Green Version]
- Kritzinger, E.C.; Bauer, F.F.; du Toit, W.J. Role of Glutathione in Winemaking: A Review. J. Agric. Food Chem. 2013, 61, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Dahl, C.; Friedrich, C.; Kletzin, A. Sulfur Oxidation in Prokaryotes. In eLS; American Cancer Society: Atlanta, GA, USA, 2008; ISBN 978-0-470-01590-2. [Google Scholar]
- Dahl, C.; Hell, R.; Leustek, T.; Knaff, D. Introduction to Sulfur Metabolism in Phototrophic Organisms. In Sulfur Metabolism in Phototrophic Organisms; Hell, R., Dahl, C., Knaff, D., Leustek, T., Eds.; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2008; pp. 1–14. ISBN 978-1-4020-6863-8. [Google Scholar]
- Arapitsas, P.; Guella, G.; Mattivi, F. The impact of SO2 on wine flavanols and indoles in relation to wine style and age. Sci. Rep. 2018, 8, 858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandri, M.P. Nutritional and Genetic Aspects of Sulfite Excretion during Fermentation by Wine Strains of Saccharomyces cerevisiae. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2018. [Google Scholar]
- Schmidt, M. Über Säuren des Schwefels. I. zur Kenntnis der wasserfreien Thioschwefelsäure. Z. Für Anorg. Allg. Chem. 1957, 289, 141–157. [Google Scholar] [CrossRef]
- Miethe, G. Untersuchungen zum Zerfall und zur Analytik der Zersetzungsprodukte von Natriumthiosulfat-Injektionslösungen. Ph.D. Dissertation, Humbolt-Universität Berlin, Berlin, Germany, 2003. [Google Scholar]
- Breuer, P.L.; Jeffrey, M.I. The reduction of copper(II) and the oxidation of thiosulfate and oxysulfur anions in gold leaching solutions. Hydrometallurgy 2003, 70, 163–173. [Google Scholar] [CrossRef]
- Funahashi, E.; Saiki, K.; Honda, K.; Sugiura, Y.; Kawano, Y.; Ohtsu, I.; Watanabe, D.; Wakabayashi, Y.; Abe, T.; Nakanishi, T.; et al. Finding of thiosulfate pathway for synthesis of organic sulfur compounds in Saccharomyces cerevisiae and improvement of ethanol production. J. Biosci. Bioeng. 2015, 120, 666–669. [Google Scholar] [CrossRef] [PubMed]
- Benchoam, D.; Cuevasanta, E.; Möller, M.N.; Alvarez, B. Hydrogen Sulfide and Persulfides Oxidation by Biologically Relevant Oxidizing Species. Antioxidants 2019, 8, 48. [Google Scholar] [CrossRef] [Green Version]
- Cuevasanta, E.; Lange, M.; Bonanata, J.; Coitiño, E.L.; Ferrer-Sueta, G.; Filipovic, M.R.; Alvarez, B. Reaction of Hydrogen Sulfide with Disulfide and Sulfenic Acid to Form the Strongly Nucleophilic Persulfide. J. Biol. Chem. 2015, 290, 26866–26880. [Google Scholar] [CrossRef] [Green Version]
- Benchoam, D.; Cuevasanta, E.; Möller, M.N.; Alvarez, B. Persulfides, at the crossroads between hydrogen sulfide and thiols. Essays Biochem. 2020, 64, 155–168. [Google Scholar] [CrossRef]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical Biology of H2S Signaling through Persulfidation. Chem. Rev. 2018, 118, 1253–1337. [Google Scholar] [CrossRef]
- Aroca, A.; Gotor, C.; Romero, L.C. Hydrogen Sulfide Signaling in Plants: Emerging Roles of Protein Persulfidation. Front. Plant Sci. 2018, 9, 1369. [Google Scholar] [CrossRef] [Green Version]
- Vojtovič, D.; Luhová, L.; Petřivalský, M. Something smells bad to plant pathogens: Production of hydrogen sulfide in plants and its role in plant defence responses. J. Adv. Res. 2021, 27, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Corvino, A.; Frecentese, F.; Magli, E.; Perissutti, E.; Santagada, V.; Scognamiglio, A.; Caliendo, G.; Fiorino, F.; Severino, B. Trends in H2S-Donors Chemistry and Their Effects in Cardiovascular Diseases. Antioxidants 2021, 10, 429. [Google Scholar] [CrossRef] [PubMed]
- Ran, M.; Wang, T.; Shao, M.; Chen, Z.; Liu, H.; Xia, Y.; Xun, L. Sensitive Method for Reliable Quantification of Sulfane Sulfur in Biological Samples. Anal. Chem. 2019, 91, 11981–11986. [Google Scholar] [CrossRef] [PubMed]
- Pluth, M.D.; Bailey, T.S.; Hammers, M.D.; Hartle, M.D.; Henthorn, H.A.; Steiger, A.K. Natural Products Containing Hydrogen Sulfide Releasing Moieties. Synlett 2015, 26, 2633–2643. [Google Scholar] [CrossRef]
- Dahl, C. Sulfur Metabolism in Phototrophic Bacteria. In Modern Topics in the Phototrophic Prokaryotes: Metabolism, Bioenergetics, and Omics; Hallenbeck, P.C., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 27–66. ISBN 978-3-319-51365-2. [Google Scholar]
- Dekker, S.; Nardin, T.; Mattana, M.; Fochi, I.; Larcher, R. Identification and characterisation of thiolated polysulfides in must and wine using online SPE UHPLC-HRMS. Anal. Bioanal. Chem. 2020, 412, 5229–5245. [Google Scholar] [CrossRef]
- Müller, N. Iminiumsalz-Strukturen bei der durch Pyridoxalphosphat (Vitamin B6) katalysierten Bildung von Aromastoffen und Fehlaromen im Wein. Z. Nat. B 2018, 73, 521–533. [Google Scholar] [CrossRef]
- Kelly, D.P. Thermodynamic aspects of energy conservation by chemolithotrophic sulfur bacteria in relation to the sulfur oxidation pathways. Arch. Microbiol. 1999, 171, 219–229. [Google Scholar] [CrossRef]
- Steudel, R.; Holdt, G.; Göbel, T.; Hazeu, W. Chromatographic Separation of Higher Polythionates SnO (n = 3…22) and Their Detection in Cultures of Thiobacillus ferroxidans; Molecular Composition of Bacterial Sulfur Secretions. Angew. Chem. Int. Ed. Engl. 1987, 26, 151–153. [Google Scholar] [CrossRef]
- Xu, Y.; Schoonen, M.A.A.; Nordstrom, D.K.; Cunningham, K.M.; Ball, J.W. Sulfur geochemistry of hydrothermal waters in Yellowstone National Park, Wyoming, USA. II. Formation and decomposition of thiosulfate and polythionate in Cinder Pool. J. Volcanol. Geotherm. Res. 2000, 97, 407–423. [Google Scholar] [CrossRef]
- Lens, P.; Pol, L.H. Environmental Technologies to Treat Sulfur Pollution—Principles and Applications; IWA Publishing: London, UK, 2005. [Google Scholar] [CrossRef]
- Rickard, D. Sulfidic Sediments and Sedimentary Rocks, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 65, ISBN 978-0-444-52989-3. [Google Scholar]
- BRENDA BRaunschweig ENzyme DAtabase. Available online: https://www.brenda-enzymes.org (accessed on 10 November 2021).
- KEGG Kyoto Encyclopedia of Genes and Genomes. Available online: https://www.genome.jp/kegg/ (accessed on 10 November 2021).
- Pedre, B.; Dick, T.P. 3-Mercaptopyruvate sulfurtransferase: An enzyme at the crossroads of sulfane sulfur trafficking. Biol. Chem. 2021, 402, 223–237. [Google Scholar] [CrossRef]
- Sbodio, J.I.; Snyder, S.H.; Paul, B.D. Regulators of the transsulfuration pathway. Br. J. Pharmacol. 2019, 176, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Kreinbring, C.A.; Hill, M.; Liu, C.; Petsko, G.A.; McCune, C.D.; Berkowitz, D.B.; Liu, D.; Ringe, D. Crystal Structures of Cystathionine β-Synthase from Saccharomyces cerevisiae: One Enzymatic Step at a Time. Biochemistry 2018, 57, 3134–3145. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, N.; Wróbel, M. H2S, Polysulfides, and Enzymes: Physiological and Pathological Aspects. Biomolecules 2020, 10, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selles, B.; Moseler, A.; Rouhier, N.; Couturier, J. Rhodanese domain-containing sulfurtransferases: Multifaceted proteins involved in sulfur trafficking in plants. J. Exp. Bot. 2019, 70, 4139–4154. [Google Scholar] [CrossRef] [PubMed]
- Andreeßen, C.; Gerlt, V.; Steinbüchel, A. Conversion of cysteine to 3-mercaptopyruvic acid by bacterial aminotransferases. Enzyme Microb. Technol. 2017, 99, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Xin, Y.; Xuan, G.; Zhao, R.; Liu, H.; Xia, Y.; Xun, L. Escherichia coli Uses Separate Enzymes to Produce H2S and Reactive Sulfane Sulfur From L-cysteine. Front. Microbiol. 2019, 10, 298. [Google Scholar] [CrossRef] [Green Version]
- Knoll, C.; du Toit, M.; Schnell, S.; Rauhut, D.; Irmler, S. Cloning and characterisation of a cystathionine β/γ-lyase from two Oenococcus oeni oenological strains. Appl. Microbiol. Biotechnol. 2011, 89, 1051–1060. [Google Scholar] [CrossRef]
- Matoba, Y.; Noda, M.; Yoshida, T.; Oda, K.; Ezumi, Y.; Yasutake, C.; Izuhara-Kihara, H.; Danshiitsoodol, N.; Kumagai, T.; Sugiyama, M. Catalytic specificity of the Lactobacillus plantarum cystathionine γ-lyase presumed by the crystallographic analysis. Sci. Rep. 2020, 10, 14886. [Google Scholar] [CrossRef]
- Pripis-Nicolau, L.; de Revel, G.; Bertrand, A.; Lonvaud-Funel, A. Methionine catabolism and production of volatile sulphur compounds by Oenococcus oeni. J. Appl. Microbiol. 2004, 96, 1176–1184. [Google Scholar] [CrossRef]
- Rollero, S.; Bloem, A.; Brand, J.; Ortiz-Julien, A.; Camarasa, C.; Divol, B. Nitrogen metabolism in three non-conventional wine yeast species: A tool to modulate wine aroma profiles. Food Microbiol. 2021, 94, 103650. [Google Scholar] [CrossRef]
- Benito, S.; Ruiz, J.; Belda, I.; Kiene, F.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Santos, A.; Rauhut, D. Application of Non-Saccharomyces Yeasts in Wine Production. In Non-Conventional Yeasts: From Basic Research to Application; Sibirny, A., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 75–89. ISBN 978-3-030-21110-3. [Google Scholar]
- Ferreira, R.B.; Piçarra-Pereira, M.A.; Monteiro, S.; Loureiro, V.B.; Teixeira, A.R. The wine proteins. Trends Food Sci. Technol. 2001, 12, 230–239. [Google Scholar] [CrossRef]
- Claus, H.; Mojsov, K. Enzymes for Wine Fermentation: Current and Perspective Applications. Fermentation 2018, 4, 52. [Google Scholar] [CrossRef] [Green Version]
- Toohey, J.I. Sulphane sulphur in biological systems: A possible regulatory role. Biochem. J. 1989, 264, 625–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustafa, A.K.; Gadalla, M.M.; Sen, N.; Kim, S.; Mu, W.; Gazi, S.K.; Barrow, R.K.; Yang, G.; Wang, R.; Snyder, S.H. H2S Signals Through Protein S-Sulfhydration. Sci. Signal. 2009, 2, ra72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dóka, É.; Pader, I.; Bíró, A.; Johansson, K.; Cheng, Q.; Ballagó, K.; Prigge, J.R.; Pastor-Flores, D.; Dick, T.P.; Schmidt, E.E.; et al. A novel persulfide detection method reveals protein persulfide- and polysulfide-reducing functions of thioredoxin and glutathione systems. Sci. Adv. 2016, 2, e1500968. [Google Scholar] [CrossRef] [Green Version]
- Bekker, M.Z.; Nandorfy, D.E.; Kulcsar, A.C.; Faucon, A.; Bindon, K.; Smith, P.A. Comparison of remediation strategies for decreasing ‘reductive’ characters in Shiraz wines. Aust. J. Grape Wine Res. 2021, 27, 52–65. [Google Scholar] [CrossRef]
- Waterhouse, A.L.; Sacks, G.L.; Jeffery, D.W. Appearance of Reduced Aromas during Bottle Storage. In Understanding Wine Chemistry; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; ISBN 978-1-118-73072-0. [Google Scholar]
- Lajin, B.; Goessler, W. Exploring the sulfur species in wine by HPLC-ICPMS/MS. Anal. Chim. Acta 2019, 1092, 1–8. [Google Scholar] [CrossRef]
- Roullier-Gall, C.; Hemmler, D.; Gonsior, M.; Li, Y.; Nikolantonaki, M.; Aron, A.; Coelho, C.; Gougeon, R.D.; Schmitt-Kopplin, P. Sulfites and the wine metabolome. Food Chem. 2017, 237, 106–113. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).