
Advances in Komagataella phaffii Engineering for the Production of Renewable Chemicals and Proteins

 , and
, and
Abstract
1. Introduction
2. Komagataella Taxonomy and Diversity
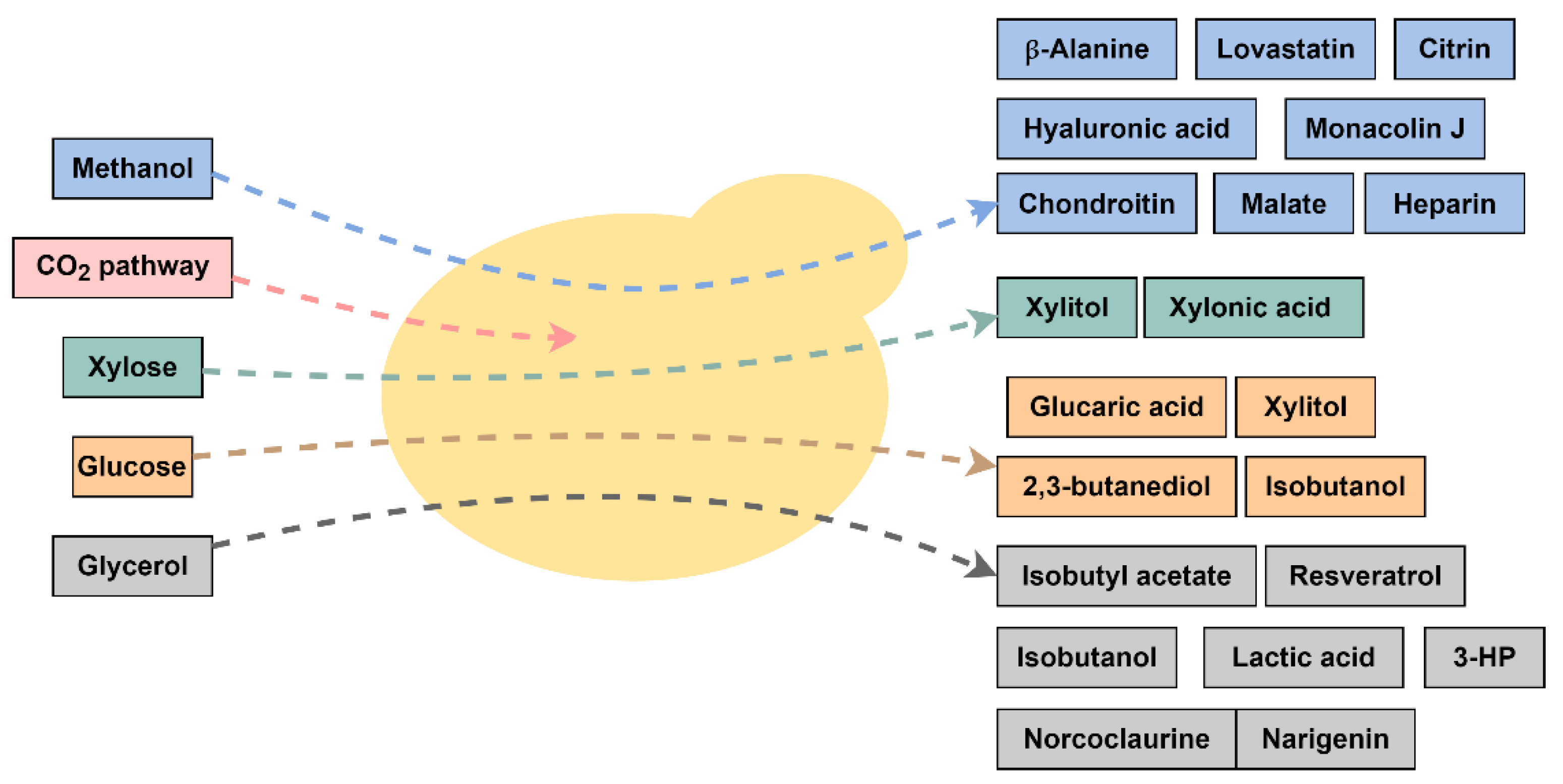
3. Production of Renewable Chemicals from Glycerol by K. phaffii
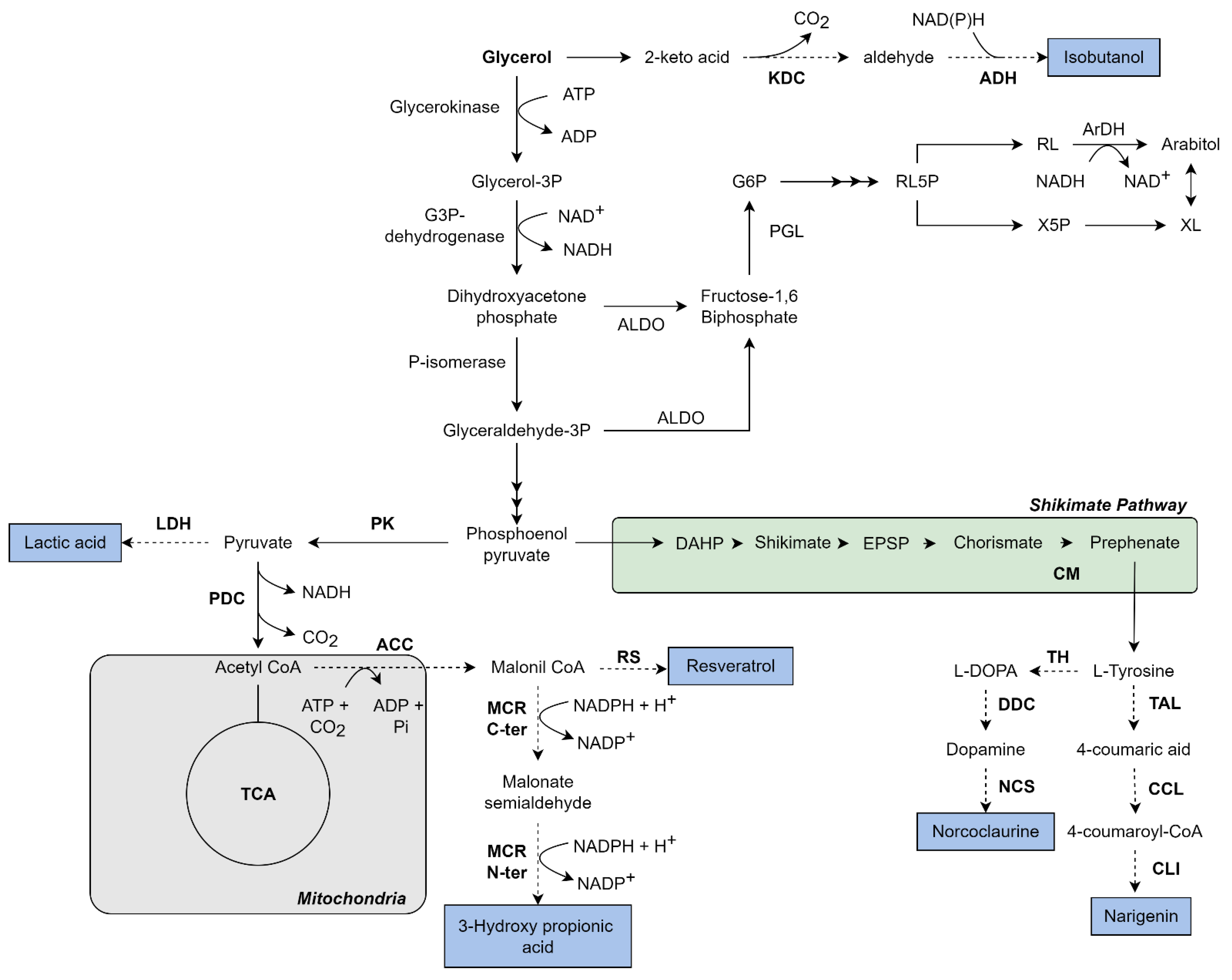
3.1. Metabolism of Glycerol
3.2. K. phaffii Engineering for Renewable Chemicals from Glycerol
3.2.1. Lactic Acid Production from Glycerol by K. phaffii
3.2.2. 3-Hydroxy-Propionic Acid (3-HP) Production from Glycerol in K. phaffii
3.2.3. Isobutanol and Isobutyl Acetate Production from Glycerol in K. phaffii
3.2.4. Aromatic Secondary Metabolite Production from Glycerol in K. phaffii
3.3. Glycerol Co-Utilization by K. phaffii
4. Renewables from Sugars and Lignocellulosic Hydrolysates
4.1. Metabolic Engineering of K. phaffii to Produce Renewables Chemicals from Glucose
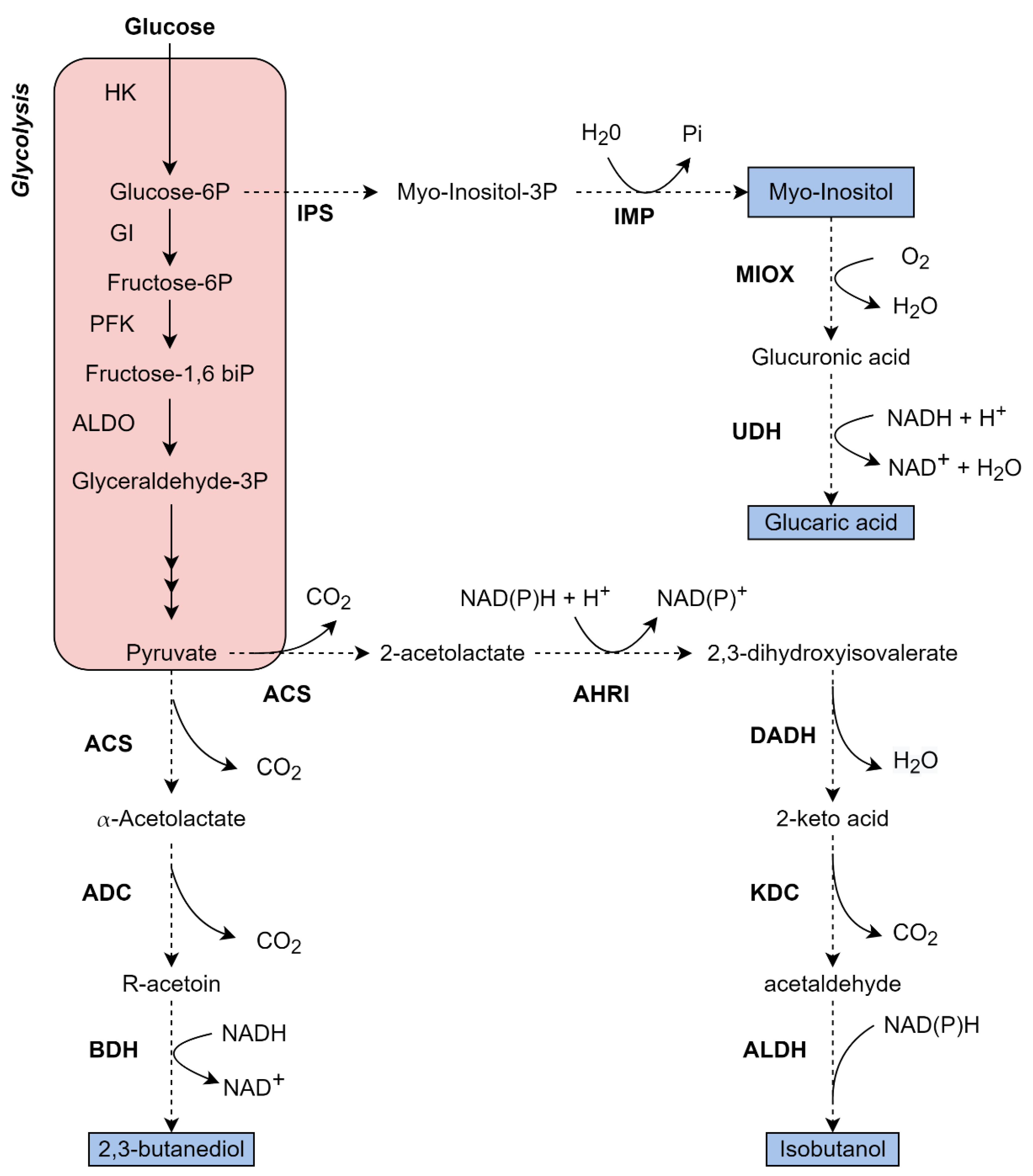
4.1.1. Isobutanol Production from Glucose in K. phaffii
4.1.2. 2,3-Butanediol Production from Glucose in K. phaffii
4.1.3. Inositol Production from Glucose in K. phaffii
4.1.4. Glucaric Acid Production from Glucose in K. phaffii
4.2. Metabolic Engineering of K. phaffii to Produce Renewables Chemicals from Xylose
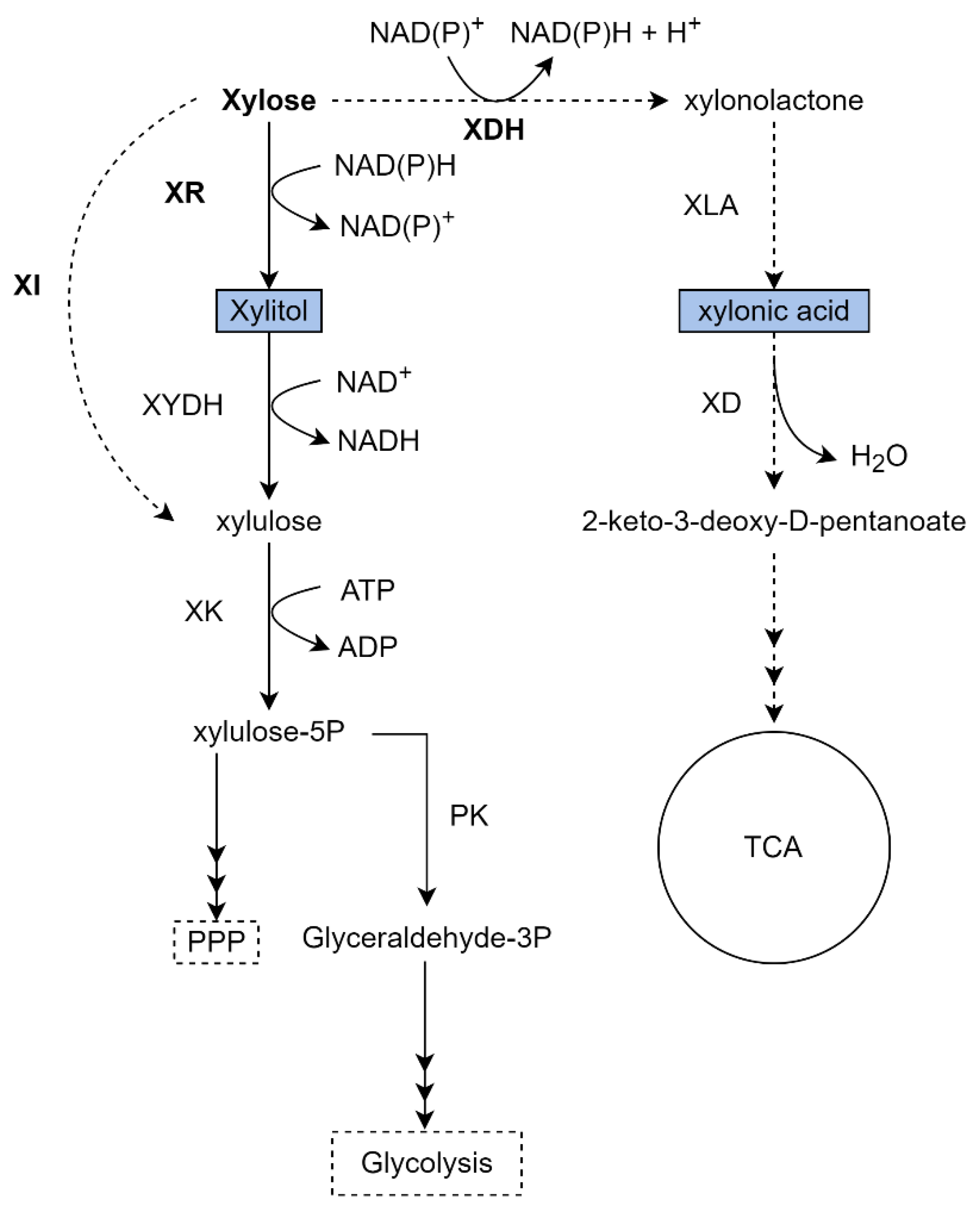
4.2.1. Xylitol Production from Xylose in K. phaffii
4.2.2. Xylonic Acid Production from Xylose in K. phaffii
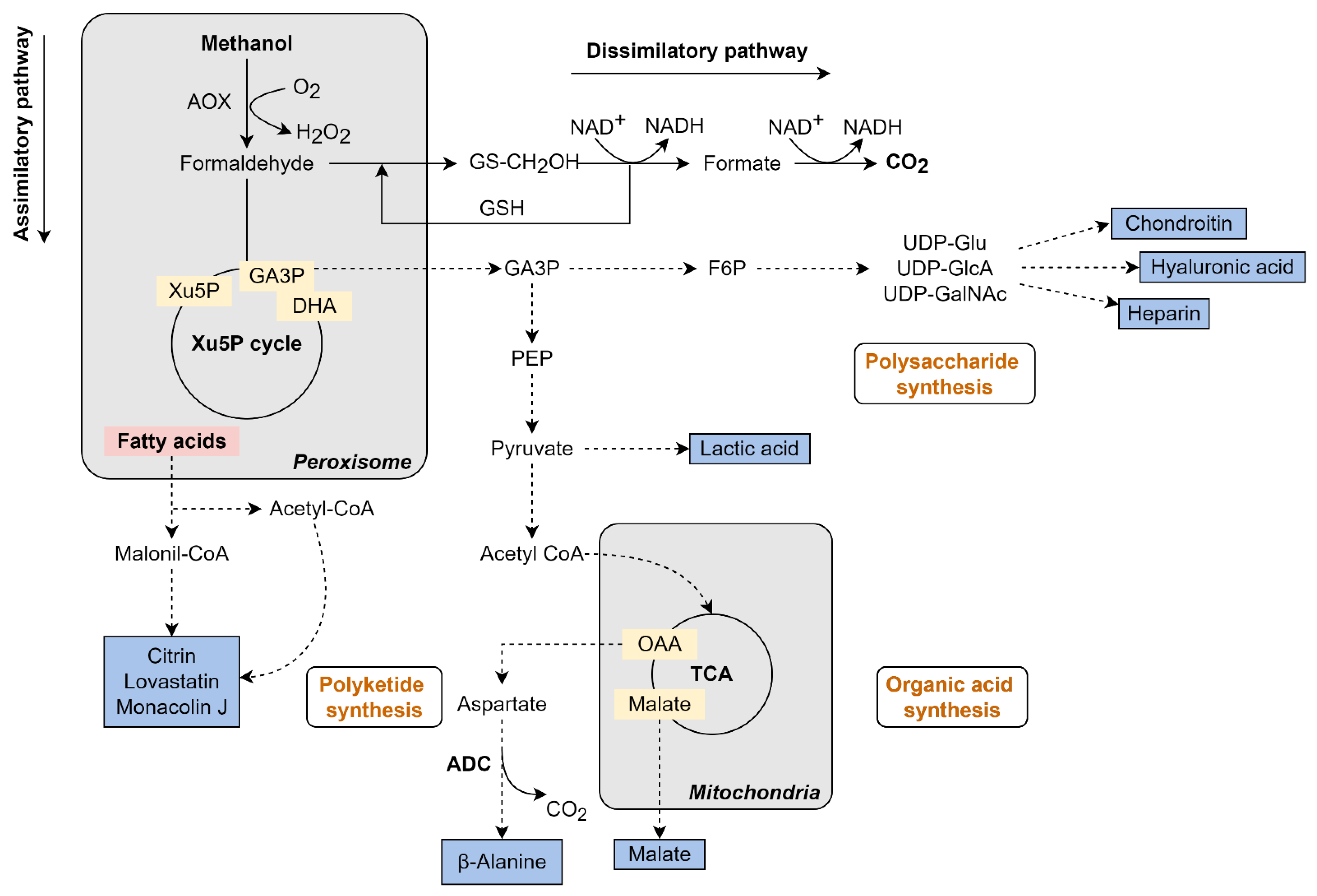
5. Renewables from Methanol and CO2
5.1. K. phaffii Engineering for CO2 Assimilation
5.2. Metabolic Engineering of K. phaffii for Production of Renewables from Methanol
5.2.1. β-Alanine Production from Methanol in K. phaffii
5.2.2. Organic Acids Production from Methanol in K. phaffii
5.2.3. Biopolymers Production from Methanol in K. phaffii
5.2.4. Polyketides Production from Methanol in K. phaffii
6. Komagataella phaffii Application in Protein Production
7. Strategies to Develop and Optimize Fermentative Processes
7.1. Fermentation Parameters
7.2. Operation Mode
7.3. Mathematical Models
8. Challenges for Industrial Processes with K. phaffii
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leong, H.Y.; Chang, C.K.; Khoo, K.S.; Chew, K.W.; Chia, S.R.; Lim, J.W.; Chang, J.S.; Show, P.L. Waste Biorefinery towards a Sustainable Circular Bioeconomy: A Solution to Global Issues. Biotechnol. Biofuels 2021, 14, 1–15. [Google Scholar] [CrossRef]
- Muscat, A.; de Olde, E.M.; Ripoll-Bosch, R.; Van Zanten, H.H.E.; Metze, T.A.P.; Termeer, C.J.A.M.; van Ittersum, M.K.; de Boer, I.J.M. Principles, Drivers and Opportunities of a Circular Bioeconomy. Nat. Food 2021, 2, 561–566. [Google Scholar] [CrossRef]
- Kardung, M.; Cingiz, K.; Costenoble, O.; Delahaye, R.; Heijman, W.; Lovrić, M.; van Leeuwen, M.; M’barek, R.; van Meijl, H.; Piotrowski, S.; et al. Development of the Circular Bioeconomy: Drivers and Indicators. Sustainability 2021, 13, 413. [Google Scholar] [CrossRef]
- Carneiro, C.V.G.; Silva, F.C.D.P.E.; Almeida, J.R. Xylitol Production: Identification and Comparison of New Producing Yeasts. Microorganisms 2019, 7, 484. [Google Scholar] [CrossRef]
- Trichez, D.; Carneiro, C.V.G.C.; Braga, M.; Almeida, J.R.M. Recent Progress in the Microbial Production of Xylonic Acid. World J. Microbiol. Biotechnol. 2022, 38, 127. [Google Scholar] [CrossRef] [PubMed]
- Ata, Ö.; Ergün, B.G.; Fickers, P.; Heistinger, L.; Mattanovich, D.; Rebnegger, C.; Gasser, B. What Makes Komagataella Phaffii Non-Conventional? FEMS Yeast Res. 2021, 21, foab059. [Google Scholar] [CrossRef]
- Bernauer, L.; Radkohl, A.; Lehmayer, L.G.K.; Emmerstorfer-Augustin, A. Komagataella Phaffii as Emerging Model Organism in Fundamental Research. Front. Microbiol. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Chiang, C.-J.; Ho, Y.-J.; Hu, M.-C.; Chao, Y.-P. Rewiring of Glycerol Metabolism in Escherichia Coli for Effective Production of Recombinant Proteins. Biotechnol. Biofuels 2020, 13, 205. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Z. Engineering Strategies for Enhanced Production of Protein and Bio-Products in Pichia Pastoris. Biotechnol. Adv. 2018, 36, 182–195. [Google Scholar] [CrossRef]
- Näätsaari, L.; Mistlberger, B.; Ruth, C.; Hajek, T.; Hartner, F.S.; Glieder, A. Deletion of the Pichia Pastoris KU70 Homologue Facilitates Platform Strain Generation for Gene Expression and Synthetic Biology. PLoS ONE 2012, 7, e39720. [Google Scholar] [CrossRef]
- Ito, Y.; Watanabe, T.; Aikawa, S.; Nishi, T.; Nishiyama, T.; Nakamura, Y.; Hasunuma, T.; Okubo, Y.; Ishii, J.; Kondo, A. Deletion of DNA Ligase IV Homolog Confers Higher Gene Targeting Efficiency on Homologous Recombination in Komagataella Phaffii. FEMS Yeast Res. 2018, 18, foy074. [Google Scholar] [CrossRef]
- Weninger, A.; Hatzl, A.-M.; Schmid, C.; Vogl, T.; Glieder, A. Combinatorial Optimization of CRISPR/Cas9 Expression Enables Precision Genome Engineering in the Methylotrophic Yeast Pichia Pastoris. J. Biotechnol. 2016, 235, 139–149. [Google Scholar] [CrossRef]
- Fischer, J.E.; Glieder, A. Current Advances in Engineering Tools for Pichia Pastoris. Curr. Opin. Biotechnol. 2019, 59, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Peña, D.A.; Gasser, B.; Zanghellini, J.; Steiger, M.G.; Mattanovich, D. Metabolic Engineering of Pichia Pastoris. Metab. Eng. 2018, 50, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Gasser, B.; Mattanovich, D. A Yeast for All Seasons—Is Pichia Pastoris a Suitable Chassis Organism for Future Bioproduction? FEMS Microbiol. Lett. 2018, 365, fny181. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Matsuda, M.; Maeda, K.; Mikata, K. The Phylogenetic Relationships of Methanol-Assimilating Yeasts Based on the Partial Sequences of 18S and 26S Ribosomal RNAs: The Proposal of Komagataella Gen. Nov. (Saccharomycetaceae). Biosci. Biotechnol. Biochem. 1995, 59, 439–444. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Identification and Phylogeny of Ascomycetous Yeasts from Analysis of Nuclear Large Subunit (26S) Ribosomal DNA Partial Sequences. Antonie Van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Guilliermond, A. Zygosaccharomyces Pastori, Nouvelle Espèce de Levures à Copulation Hétérogamique. Bull. Société Mycol. Fr. 1920, 36, 203–211. [Google Scholar]
- Heistinger, L.; Gasser, B.; Mattanovich, D. Microbe Profile: Komagataella Phaffii: A Methanol Devouring Biotech Yeast Formerly Known as Pichia Pastoris. Microbiology 2020, 166, 614–616. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. The Yeasts—A Taxonomy Study; Elsevier Science: Amsterdam, The Netherlands, 2011; Volume 5, ISBN 978-0-08-093127-2. [Google Scholar]
- Heistinger, L.; Dohm, J.C.; Paes, B.G.; Koizar, D.; Troyer, C.; Ata, Ö.; Steininger-Mairinger, T.; Mattanovich, D. Genotypic and Phenotypic Diversity among Komagataella Species Reveals a Hidden Pathway for Xylose Utilization. Microb. Cell Factories 2022, 21, 70. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P. Description of Komagataella Phaffii Sp. Nov. and the Transfer of Pichia Pseudopastoris to the Methylotrophic Yeast Genus Komagataella. Int. J. Syst. Evol. Microbiol. 2005, 55, 973–976. [Google Scholar] [CrossRef] [PubMed]
- De Schutter, K.; Lin, Y.-C.; Tiels, P.; Van Hecke, A.; Glinka, S.; Weber-Lehmann, J.; Rouzé, P.; Van de Peer, Y.; Callewaert, N. Genome Sequence of the Recombinant Protein Production Host Pichia Pastoris. Nat. Biotechnol. 2009, 27, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Love, K.R.; Shah, K.A.; Whittaker, C.A.; Wu, J.; Bartlett, M.C.; Ma, D.; Leeson, R.L.; Priest, M.; Borowsky, J.; Young, S.K.; et al. Comparative Genomics and Transcriptomics of Pichia Pastoris. BMC Genom. 2016, 17, 550. [Google Scholar] [CrossRef] [PubMed]
- Mattanovich, D.; Graf, A.; Stadlmann, J.; Dragosits, M.; Redl, A.; Maurer, M.; Kleinheinz, M.; Sauer, M.; Altmann, F.; Gasser, B. Genome, Secretome and Glucose Transport Highlight Unique Features of the Protein Production Host Pichia Pastoris. Microb. Cell Factories 2009, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.R.M.; Fávaro, L.C.L.; Quirino, B.F. Biodiesel Biorefinery: Opportunities and Challenges for Microbial Production of Fuels and Chemicals from Glycerol Waste. Biotechnol. Biofuels 2012, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- da Silva, G.P.; Mack, M.; Contiero, J. Glycerol: A Promising and Abundant Carbon Source for Industrial Microbiology. Biotechnol. Adv. 2009, 27, 30–39. [Google Scholar] [CrossRef]
- Tomàs-Gamisans, M.; Ødum, A.S.R.; Workman, M.; Ferrer, P.; Albiol, J. Glycerol Metabolism of Pichia Pastoris (Komagataella Spp.) Characterised by 13C-Based Metabolic Flux Analysis. New Biotechnol. 2019, 50, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Prielhofer, R.; Cartwright, S.P.; Graf, A.B.; Valli, M.; Bill, R.M.; Mattanovich, D.; Gasser, B. Pichia Pastoris Regulates Its Gene-Specific Response to Different Carbon Sources at the Transcriptional, Rather than the Translational, Level. BMC Genom. 2015, 16, 167. [Google Scholar] [CrossRef] [PubMed]
- Çelik, E.; Ozbay, N.; Oktar, N.; Çalık, P. Use of Biodiesel Byproduct Crude Glycerol as the Carbon Source for Fermentation Processes by Recombinant Pichia Pastoris. Ind. Eng. Chem. Res. 2008, 47, 2985–2990. [Google Scholar] [CrossRef]
- Kumokita, R.; Bamba, T.; Inokuma, K.; Yoshida, T.; Ito, Y.; Kondo, A.; Hasunuma, T. Construction of an l-Tyrosine Chassis in Pichia Pastoris Enhances Aromatic Secondary Metabolite Production from Glycerol. ACS Synth. Biol. 2022, 11, 2098–2107. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, D. Toward Glycerol Biorefinery: Metabolic Engineering for the Production of Biofuels and Chemicals from Glycerol. Biotechnol. Biofuels 2016, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Miao, J.; Luo, W.; Li, G.; Du, Y.; Yu, X. Crude Glycerol from Biodiesel as a Carbon Source for Production of a Recombinant Highly Thermostable β-Mannanase by Pichia Pastoris. Biotechnol. Lett. 2018, 40, 135–141. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Recent Advances in Lactic Acid Production by Microbial Fermentation Processes. Biotechnol. Adv. 2013, 31, 877–902. [Google Scholar] [CrossRef] [PubMed]
- Abedi, E.; Hashemi, S.M.B. Lactic Acid Production—Producing Microorganisms and Substrates Sources-State of Art. Heliyon 2020, 6, e04974. [Google Scholar] [CrossRef] [PubMed]
- de Lima, P.B.A.; Mulder, K.C.L.; Melo, N.T.M.; Carvalho, L.S.; Menino, G.S.; Mulinari, E.; de Castro, V.H.; dos Reis, T.F.; Goldman, G.H.; Magalhães, B.S.; et al. Novel Homologous Lactate Transporter Improves L-Lactic Acid Production from Glycerol in Recombinant Strains of Pichia Pastoris. Microb. Cell Factories 2016, 15, 158. [Google Scholar] [CrossRef]
- Melo, N.; Mulder, K.; Nicola, A.; Carvalho, L.; Menino, G.; Mulinari, E.; Parachin, N. Effect of Pyruvate Decarboxylase Knockout on Product Distribution Using Pichia Pastoris (Komagataella Phaffii) Engineered for Lactic Acid Production. Bioengineering 2018, 5, 17. [Google Scholar] [CrossRef]
- Tamires Moreira Melo, N.; Pontes, G.C.; Procópio, D.P.; de Gois e Cunha, G.C.; Eliodório, K.P.; Costa Paes, H.; Basso, T.O.; Parachin, N.S. Evaluation of Product Distribution in Chemostat and Batch Fermentation in Lactic Acid-Producing Komagataella Phaffii Strains Utilizing Glycerol as Substrate. Microorganisms 2020, 8, 781. [Google Scholar] [CrossRef]
- Fina, A.; Brêda, G.C.; Pérez-Trujillo, M.; Freire, D.M.G.; Almeida, R.V.; Albiol, J.; Ferrer, P. Benchmarking Recombinant Pichia Pastoris for 3-hydroxypropionic Acid Production from Glycerol. Microb. Biotechnol. 2021, 14, 1671–1682. [Google Scholar] [CrossRef]
- Siripong, W.; Wolf, P.; Kusumoputri, T.P.; Downes, J.J.; Kocharin, K.; Tanapongpipat, S.; Runguphan, W. Metabolic Engineering of Pichia Pastoris for Production of Isobutanol and Isobutyl Acetate. Biotechnol. Biofuels 2018, 11, 1. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Z. Production of (2R, 3R)-2,3-Butanediol Using Engineered Pichia Pastoris: Strain Construction, Characterization and Fermentation. Biotechnol. Biofuels 2018, 11, 35. [Google Scholar] [CrossRef]
- Liu, Y.; Gong, X.; Wang, C.; Du, G.; Chen, J.; Kang, Z. Production of Glucaric Acid from Myo-Inositol in Engineered Pichia Pastoris. Enzym. Microb. Technol. 2016, 91, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, X.; Luo, H.; Wang, Y.; Wang, Y.; Tu, T.; Qin, X.; Su, X.; Huang, H.; Yao, B.; et al. Metabolic Engineering of Pichia Pastoris for Myo-Inositol Production by Dynamic Regulation of Central Metabolism. Microb. Cell Factories 2022, 21, 112. [Google Scholar] [CrossRef] [PubMed]
- Louie, T.M.; Louie, K.; DenHartog, S.; Gopishetty, S.; Subramanian, M.; Arnold, M.; Das, S. Production of Bio-xylitol from Xylose by an Engineered P.Pastoris. Microb. Cell Factories 2021, 20, 50. [Google Scholar] [CrossRef] [PubMed]
- Ramos, T.G.S.; Justen, F.; Carneiro, C.V.G.C.; Honorato, V.M.; Franco, P.F.; Vieira, F.S.; Trichez, D.; Rodrigues, C.M.; Almeida, J.R.M. Xylonic Acid Production by Recombinant Komagataella Phaffii Strains Engineered with Newly Identified Xylose Dehydrogenases. Bioresour. Technol. Rep. 2021, 16, 100825. [Google Scholar] [CrossRef]
- Miao, L.; Li, Y.; Zhu, T. Metabolic Engineering of Methylotrophic Pichia Pastoris for the Production of β-Alanine. Bioresour. Bioprocess. 2021, 8, 89. [Google Scholar] [CrossRef]
- Yamada, R.; Ogura, K.; Kimoto, Y.; Ogino, H. Toward the Construction of a Technology Platform for Chemicals Production from Methanol: D-Lactic Acid Production from Methanol by an Engineered Yeast Pichia Pastoris. World J. Microbiol. Biotechnol. 2019, 35, 37. [Google Scholar] [CrossRef]
- Guo, F.; Dai, Z.; Peng, W.; Zhang, S.; Zhou, J.; Ma, J.; Dong, W.; Xin, F.; Zhang, W.; Jiang, M. Metabolic Engineering of Pichia Pastoris for Malic Acid Production from Methanol. Biotechnol. Bioeng. 2021, 118, 357–371. [Google Scholar] [CrossRef]
- Liu, Y.; Tu, X.; Xu, Q.; Bai, C.; Kong, C.; Liu, Q.; Yu, J.; Peng, Q.; Zhou, X.; Zhang, Y.; et al. Engineered Monoculture and Co-Culture of Methylotrophic Yeast for de Novo Production of Monacolin J and Lovastatin from Methanol. Metab. Eng. 2018, 45, 189–199. [Google Scholar] [CrossRef]
- Gao, L.; Cai, M.; Shen, W.; Xiao, S.; Zhou, X.; Zhang, Y. Engineered Fungal Polyketide Biosynthesis in Pichia Pastoris: A Potential Excellent Host for Polyketide Production. Microb. Cell Factories 2013, 12, 77. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, W.; Wang, Y.; Sheng, J.; Xu, R.; Li, J.; Du, G.; Kang, Z. Biosynthesis of Non-Animal Chondroitin Sulfate from Methanol Using Genetically Engineered Pichia Pastoris. Green Chem. 2021, 23, 4365–4374. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Zhou, Z.; Wang, P.; Xi, X.; Hu, S.; Xu, R.; Du, G.; Li, J.; Chen, J.; et al. Synthesis of Bioengineered Heparin by Recombinant Yeast Pichia Pastoris. Green Chem. 2022, 24, 3180–3192. [Google Scholar] [CrossRef]
- Jeong, E.; Shim, W.Y.; Kim, J.H. Metabolic Engineering of Pichia Pastoris for Production of Hyaluronic Acid with High Molecular Weight. J. Biotechnol. 2014, 185, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Ashok, S.; Park, S. Recent Advances in Biological Production of 3-Hydroxypropionic Acid. Biotechnol. Adv. 2013, 31, 945–961. [Google Scholar] [CrossRef] [PubMed]
- de Fouchécour, F.; Sánchez-Castañeda, A.-K.; Saulou-Bérion, C.; Spinnler, H.É. Process Engineering for Microbial Production of 3-Hydroxypropionic Acid. Biotechnol. Adv. 2018, 36, 1207–1222. [Google Scholar] [CrossRef]
- Tashiro, Y.; Rodriguez, G.M.; Atsumi, S. 2-Keto Acids Based Biosynthesis Pathways for Renewable Fuels and Chemicals. J. Ind. Microbiol. Biotechnol. 2015, 42, 361–373. [Google Scholar] [CrossRef]
- Weber, C.; Farwick, A.; Benisch, F.; Brat, D.; Dietz, H.; Subtil, T.; Boles, E. Trends and Challenges in the Microbial Production of Lignocellulosic Bioalcohol Fuels. Appl. Microbiol. Biotechnol. 2010, 87, 1303–1315. [Google Scholar] [CrossRef]
- Wess, J.; Brinek, M.; Boles, E. Improving Isobutanol Production with the Yeast Saccharomyces Cerevisiae by Successively Blocking Competing Metabolic Pathways as Well as Ethanol and Glycerol Formation. Biotechnol. Biofuels 2019, 12, 173. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, X.; Luo, H.; Wang, Y.; Wang, Y.; Tu, T.; Qin, X.; Huang, H.; Bai, Y.; Yao, B.; et al. Metabolic Engineering of Komagataella Phaffii for Synergetic Utilization of Glucose and Glycerol. Yeast 2022, 39, 412–421. [Google Scholar] [CrossRef]
- Wang, B.; He, M.; Wang, X.; Tang, H.; Zhu, X. A Multi-Model Predictive Control Method for the Pichia Pastoris Fermentation Process Based on Relative Error Weighting Algorithm. Alex. Eng. J. 2022, 61, 9649–9660. [Google Scholar] [CrossRef]
- Baruah, J.; Nath, B.K.; Sharma, R.; Kumar, S.; Deka, R.C.; Baruah, D.C.; Kalita, E. Recent Trends in the Pretreatment of Lignocellulosic Biomass for Value-Added Products. Front. Energy Res. 2018, 6, 141. [Google Scholar] [CrossRef]
- Zabed, H.; Sahu, J.N.; Suely, A.; Boyce, A.N.; Faruq, G. Bioethanol Production from Renewable Sources: Current Perspectives and Technological Progress. Renew. Sustain. Energy Rev. 2017, 71, 475–501. [Google Scholar] [CrossRef]
- Chen, H. Chemical Composition and Structure of Natural Lignocellulose. In Biotechnology of Lignocellulose; Springer: Dordrecht, The Netherlands, 2014; pp. 25–71. ISBN 978-94-007-6897-0. [Google Scholar]
- Chandel, A.K.; Garlapati, V.K.; Jeevan Kumar, S.P.; Hans, M.; Singh, A.K.; Kumar, S. The Role of Renewable Chemicals and Biofuels in Building a Bioeconomy. Biofuels Bioprod. Bioref. 2020, 14, 830–844. [Google Scholar] [CrossRef]
- Kim, D. Physico-Chemical Conversion of Lignocellulose: Inhibitor Effects and Detoxification Strategies: A Mini Review. Molecules 2018, 23, 309. [Google Scholar] [CrossRef]
- Liu, C.-G.; Xiao, Y.; Xia, X.-X.; Zhao, X.-Q.; Peng, L.; Srinophakun, P.; Bai, F.-W. Cellulosic Ethanol Production: Progress, Challenges and Strategies for Solutions. Biotechnol. Adv. 2019, 37, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Liu, D.; Zhao, X. Conversion of Lignocellulose to Biofuels and Chemicals via Sugar Platform: An Updated Review on Chemistry and Mechanisms of Acid Hydrolysis of Lignocellulose. Renew. Sustain. Energy Rev. 2021, 146, 111169. [Google Scholar] [CrossRef]
- Chen, H.; Liu, J.; Chang, X.; Chen, D.; Xue, Y.; Liu, P.; Lin, H.; Han, S. A Review on the Pretreatment of Lignocellulose for High-Value Chemicals. Fuel Process. Technol. 2017, 160, 196–206. [Google Scholar] [CrossRef]
- Almeida, J.R.; Modig, T.; Petersson, A.; Hähn-Hägerdal, B.; Lidén, G.; Gorwa-Grauslund, M.F. Increased Tolerance and Conversion of Inhibitors in Lignocellulosic Hydrolysates BySaccharomyces Cerevisiae. J. Chem. Technol. Biotechnol. 2007, 82, 340–349. [Google Scholar] [CrossRef]
- Sjulander, N.; Kikas, T. Origin, Impact and Control of Lignocellulosic Inhibitors in Bioethanol Production—A Review. Energies 2020, 13, 4751. [Google Scholar] [CrossRef]
- Paes, B.G.; Steindorff, A.S.; Formighieri, E.F.; Pereira, I.S.; Almeida, J.R.M. Physiological Characterization and Transcriptome Analysis of Pichia Pastoris Reveals Its Response to Lignocellulose-Derived Inhibitors. AMB Expr. 2021, 11, 2. [Google Scholar] [CrossRef]
- Bustos, C.; Quezada, J.; Veas, R.; Altamirano, C.; Braun-Galleani, S.; Fickers, P.; Berrios, J. Advances in Cell Engineering of the Komagataella Phaffii Platform for Recombinant Protein Production. Metabolites 2022, 12, 346. [Google Scholar] [CrossRef]
- Lamsen, E.N.; Atsumi, S. Recent Progress in Synthetic Biology for Microbial Production of C3–C10 Alcohols. Front. Microbiol. 2012, 3, 196. [Google Scholar] [CrossRef] [PubMed]
- Jong, E.d.; Stichnothe, H.; Bell, G.; Jørgensen, H. Task 42-Bio-Based Chemicals: A 2020 Update; IEA Bioenergy: Vienna, Austria, 2020. [Google Scholar]
- Zhao, E.M.; Zhang, Y.; Mehl, J.; Park, H.; Lalwani, M.A.; Toettcher, J.E.; Avalos, J.L. Optogenetic Regulation of Engineered Cellular Metabolism for Microbial Chemical Production. Nature 2018, 555, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Motta-Mena, L.B.; Reade, A.; Mallory, M.J.; Glantz, S.; Weiner, O.D.; Lynch, K.W.; Gardner, K.H. An Optogenetic Gene Expression System with Rapid Activation and Deactivation Kinetics. Nat. Chem. Biol. 2014, 10, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, N.M.; Binod, P.; Sindhu, R.; Awasthi, M.K.; Pandey, A. Microbial Engineering for the Production of Isobutanol: Current Status and Future Directions. Bioengineered 2021, 12, 12308–12321. [Google Scholar] [CrossRef] [PubMed]
- Hazeena, S.H.; Sindhu, R.; Pandey, A.; Binod, P. Lignocellulosic Bio-Refinery Approach for Microbial 2,3-Butanediol Production. Bioresour. Technol. 2020, 302, 122873. [Google Scholar] [CrossRef]
- Lian, J.; Chao, R.; Zhao, H. Metabolic Engineering of a Saccharomyces Cerevisiae Strain Capable of Simultaneously Utilizing Glucose and Galactose to Produce Enantiopure (2R,3R)-Butanediol. Metab. Eng. 2014, 23, 92–99. [Google Scholar] [CrossRef]
- Li, Y.; Han, P.; Wang, J.; Shi, T.; You, C. Production of Myo-inositol: Recent Advance and Prospective. Biotechnol. Appl. Biochem. 2022, 69, 1101–1111. [Google Scholar] [CrossRef]
- Braga, M.; Ferreira, P.M.; Almeida, J.R.M. Screening Method to Prioritize Relevant Bio-based Acids and Their Biochemical Processes Using Recent Patent Information. Biofuels Bioprod. Bioref. 2021, 15, 231–249. [Google Scholar] [CrossRef]
- Francois, J.M.; Alkim, C.; Morin, N. Engineering microbial pathways for production of bio-based chemicals from lignocellulosic sugars: Current status and perspectives. Biotechnol. Biofuels 2020, 13, 118. [Google Scholar] [CrossRef]
- Li, P.; Sun, H.; Chen, Z.; Li, Y.; Zhu, T. Construction of Efficient Xylose Utilizing Pichia Pastoris for Industrial Enzyme Production. Microb. Cell Factories 2015, 14, 22. [Google Scholar] [CrossRef]
- Araújo, D.; Costa, T.; Freitas, F. Biovalorization of Lignocellulosic Materials for Xylitol Production by the Yeast Komagataella Pastoris. Appl. Sci. 2021, 11, 5516. [Google Scholar] [CrossRef]
- Guo, Y.; Liao, Y.; Wang, J.; Ma, C.; Qin, J.; Feng, J.; Li, Y.; Wang, X.; Chen, K. Methylotrophy of Pichia Pastoris: Current Advances, Applications, and Future Perspectives for Methanol-Based Biomanufacturing. ACS Sustain. Chem. Eng. 2022, 10, 1741–1752. [Google Scholar] [CrossRef]
- Whitaker, W.B.; Sandoval, N.R.; Bennett, R.K.; Fast, A.G.; Papoutsakis, E.T. Synthetic Methylotrophy: Engineering the Production of Biofuels and Chemicals Based on the Biology of Aerobic Methanol Utilization. Curr. Opin. Biotechnol. 2015, 33, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Peiro, C.; Vicente, C.M.; Jallet, D.; Heux, S. From a Hetero- to a Methylotrophic Lifestyle: Flash Back on the Engineering Strategies to Create Synthetic Methanol-User Strains. Front. Bioeng. Biotechnol. 2022, 10, 907861. [Google Scholar] [CrossRef]
- Cotton, C.A.; Claassens, N.J.; Benito-Vaquerizo, S.; Bar-Even, A. Renewable Methanol and Formate as Microbial Feedstocks. Curr. Opin. Biotechnol. 2020, 62, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Rußmayer, H.; Buchetics, M.; Gruber, C.; Valli, M.; Grillitsch, K.; Modarres, G.; Guerrasio, R.; Klavins, K.; Neubauer, S.; Drexler, H.; et al. Systems-Level Organization of Yeast Methylotrophic Lifestyle. BMC Biol. 2015, 13, 80. [Google Scholar] [CrossRef]
- De, S.; Mattanovich, D.; Ferrer, P.; Gasser, B. Established Tools and Emerging Trends for the Production of Recombinant Proteins and Metabolites in Pichia Pastoris. Essays Biochem. 2021, 65, 293–307. [Google Scholar] [CrossRef]
- Zahrl, R.J.; Peña, D.A.; Mattanovich, D.; Gasser, B. Systems Biotechnology for Protein Production in Pichia Pastoris. FEMS Yeast Res. 2017, 17, fox068. [Google Scholar] [CrossRef]
- Gassler, T.; Sauer, M.; Gasser, B.; Egermeier, M.; Troyer, C.; Causon, T.; Hann, S.; Mattanovich, D.; Steiger, M.G. The Industrial Yeast Pichia Pastoris Is Converted from a Heterotroph into an Autotroph Capable of Growth on CO2. Nat. Biotechnol. 2020, 38, 210–216. [Google Scholar] [CrossRef]
- Gassler, T.; Baumschabl, M.; Sallaberger, J.; Egermeier, M.; Mattanovich, D. Adaptive Laboratory Evolution and Reverse Engineering Enhances Autotrophic Growth in Pichia Pastoris. Metab. Eng. 2022, 69, 112–121. [Google Scholar] [CrossRef]
- Zhang, C.; Ottenheim, C.; Weingarten, M.; Ji, L. Microbial Utilization of Next-Generation Feedstocks for the Biomanufacturing of Value-Added Chemicals and Food Ingredients. Front. Bioeng. Biotechnol. 2022, 10, 874612. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Ge, C.; Deng, L.; Tan, T.; Wang, F. C4-Dicarboxylic Acid Production by Overexpressing the Reductive TCA Pathway. FEMS Microbiol. Lett. 2015, 362, fnv052. [Google Scholar] [CrossRef] [PubMed]
- Ucm, R.; Aem, M.; Lhb, Z.; Kumar, V.; Taherzadeh, M.J.; Garlapati, V.K.; Chandel, A.K. Comprehensive Review on Biotechnological Production of Hyaluronic Acid: Status, Innovation, Market and Applications. Bioengineered 2022, 13, 9645–9661. [Google Scholar] [CrossRef]
- Xue, Y.; Kong, C.; Shen, W.; Bai, C.; Ren, Y.; Zhou, X.; Zhang, Y.; Cai, M. Methylotrophic Yeast Pichia Pastoris as a Chassis Organism for Polyketide Synthesis via the Full Citrinin Biosynthetic Pathway. J. Biotechnol. 2017, 242, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Karbalaei, M.; Rezaee, S.; Farsiani, H. Pichia Pastoris: A Highly Successful Expression System for Optimal Synthesis of Heterologous Proteins. J. Cell. Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef]
- Ahmad, M.; Hirz, M.; Pichler, H.; Schwab, H. Protein Expression in Pichia Pastoris: Recent Achievements and Perspectives for Heterologous Protein Production. Appl. Microbiol. Biotechnol. 2014, 98, 5301–5317. [Google Scholar] [CrossRef]
- Cereghino, J.L.; Cregg, J.M. Heterologous Protein Expression in the Methylotrophic Yeast Pichia Pastoris. FEMS Microbiol. Rev. 2000, 24, 45–66. [Google Scholar] [CrossRef]
- de Almeida, J.R.M.; de Moraes, L.M.P.; Torres, F.A.G. Molecular Characterization of the 3-Phosphoglycerate Kinase Gene (PGK1) from the Methylotrophic Yeast Pichia Pastoris. Yeast 2005, 22, 725–737. [Google Scholar] [CrossRef]
- Spiegel, H.; Schinkel, H.; Kastilan, R.; Dahm, P.; Boes, A.; Scheuermayer, M.; Chudobová, I.; Maskus, D.; Fendel, R.; Schillberg, S.; et al. Optimization of a Multi-Stage, Multi-Subunit Malaria Vaccine Candidate for the Production in Pichia Pastoris by the Identification and Removal of Protease Cleavage Sites: Optimization of a Malaria Vaccine Candidate. Biotechnol. Bioeng. 2015, 112, 659–667. [Google Scholar] [CrossRef]
- Tripathi, L.; Mani, S.; Raut, R.; Poddar, A.; Tyagi, P.; Arora, U.; de Silva, A.; Swaminathan, S.; Khanna, N. Pichia Pastoris-Expressed Dengue 3 Envelope-Based Virus-like Particles Elicit Predominantly Domain III-Focused High Titer Neutralizing Antibodies. Front. Microbiol. 2015, 6, 1005. [Google Scholar] [CrossRef]
- Saraswat, S.; Athmaram, T.N.; Parida, M.; Agarwal, A.; Saha, A.; Dash, P.K. Expression and Characterization of Yeast Derived Chikungunya Virus Like Particles (CHIK-VLPs) and Its Evaluation as a Potential Vaccine Candidate. PLoS Negl. Trop. Dis. 2016, 10, e0004782. [Google Scholar] [CrossRef] [PubMed]
- Farsiani, H.; Mosavat, A.; Soleimanpour, S.; Sadeghian, H.; Akbari Eydgahi, M.R.; Ghazvini, K.; Sankian, M.; Aryan, E.; Jamehdar, S.A.; Rezaee, S.A. Fc-Based Delivery System Enhances Immunogenicity of a Tuberculosis Subunit Vaccine Candidate Consisting of the ESAT-6:CFP-10 Complex. Mol. BioSyst. 2016, 12, 2189–2201. [Google Scholar] [CrossRef] [PubMed]
- Argentinian AntiCovid Consortium. Structural and Functional Comparison of SARS-CoV-2-Spike Receptor Binding Domain Produced in Pichia Pastoris and Mammalian Cells. Sci. Rep. 2020, 10, 21779. [Google Scholar] [CrossRef] [PubMed]
- Baeshen, M.N.; Bouback, T.A.F.; Alzubaidi, M.A.; Bora, R.S.; Alotaibi, M.A.T.; Alabbas, O.T.O.; Alshahrani, S.M.; Aljohani, A.A.M.; Munshi, R.A.A.; Al-Hejin, A.; et al. Expression and Purification of C-Peptide Containing Insulin Using Pichia Pastoris Expression System. BioMed Res. Int. 2016, 2016, 3423685. [Google Scholar] [CrossRef] [PubMed]
- Eissazadeh, S.; Moeini, H.; Dezfouli, M.G.; Heidary, S.; Nelofer, R.; Abdullah, M.P. Production of Recombinant Human Epidermal Growth Factor in Pichia Pastoris. Braz. J. Microbiol. 2017, 48, 286–293. [Google Scholar] [CrossRef]
- Li, P.; Yang, G.; Geng, X.; Shi, J.; Li, B.; Wang, Z.; Zhang, Q.; Yang, Y.; Xu, C. High-Level Secretory Expression and Purification of Recombinant Human Interleukin 1 Beta in Pichia Pastoris. Protein Pept. Lett. 2016, 23, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.-M.; Li, W.-J.; Shi, L.-Y.; Lv, Y.-J.; Sun, X.-Q.; Hu, J.-C.; Fan, Z.-C. Expression, Purification and Characterization of a Recombinant Antimicrobial Peptide Hispidalin in Pichia Pastoris. Protein Expr. Purif. 2019, 160, 19–27. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, M.; Chen, X.; Yang, G.; Yang, T.; Yu, L.; Hui, L.; Wang, X. Expression and Antibacterial Activity of Hybrid Antimicrobial Peptide CecropinA-Thanatin in Pichia Pastoris. Front. Lab. Med. 2018, 2, 23–29. [Google Scholar] [CrossRef]
- Popa, C.; Shi, X.; Ruiz, T.; Ferrer, P.; Coca, M. Biotechnological Production of the Cell Penetrating Antifungal PAF102 Peptide in Pichia Pastoris. Front. Microbiol. 2019, 10, 1472. [Google Scholar] [CrossRef]
- Xing, L.-W.; Tian, S.-X.; Gao, W.; Yang, N.; Qu, P.; Liu, D.; Jiao, J.; Wang, J.; Feng, X.-J. Recombinant Expression and Biological Characterization of the Antimicrobial Peptide Fowlicidin-2 in Pichia Pastoris. Exp. Ther. Med. 2016, 12, 2324–2330. [Google Scholar] [CrossRef]
- Zhu, W.; Gong, G.; Pan, J.; Han, S.; Zhang, W.; Hu, Y.; Xie, L. High Level Expression and Purification of Recombinant Human Serum Albumin in Pichia Pastoris. Protein Expr. Purif. 2018, 147, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Várnai, A.; Tang, C.; Bengtsson, O.; Atterton, A.; Mathiesen, G.; Eijsink, V.G. Expression of Endoglucanases in Pichia Pastoris under Control of the GAP Promoter. Microb. Cell Factories 2014, 13, 57. [Google Scholar] [CrossRef] [PubMed]
- Mellitzer, A.; Weis, R.; Glieder, A.; Flicker, K. Expression of Lignocellulolytic Enzymes in Pichia Pastoris. Microb. Cell Factories 2012, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ning, C.; Yuan, M.; Fu, X.; Yang, S.; Wei, X.; Xiao, M.; Mou, H.; Zhu, C. High-Efficiency Expression of a Superior β-Mannanase Engineered by Cooperative Substitution Method in Pichia Pastoris and Its Application in Preparation of Prebiotic Mannooligosaccharides. Bioresour. Technol. 2020, 311, 123482. [Google Scholar] [CrossRef]
- Rodrigues, K.B.; Macêdo, J.K.A.; Teixeira, T.; Barros, J.S.; Araújo, A.C.B.; Santos, F.P.; Quirino, B.F.; Brasil, B.S.A.F.; Salum, T.F.C.; Abdelnur, P.V.; et al. Recombinant Expression of Thermobifida Fusca E7 LPMO in Pichia Pastoris and Escherichia Coli and Their Functional Characterization. Carbohydr. Res. 2017, 448, 175–181. [Google Scholar] [CrossRef]
- Matsuyama, K.; Sunagawa, N.; Igarashi, K. Mutation of Cysteine Residues Increases Heterologous Expression of Peach Expansin in the Methylotrophic Yeast Pichia Pastoris. Plant Biotechnol. 2020, 37, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Miao, T.; Basit, A.; Liu, J.; Zheng, F.; Rahim, K.; Lou, H.; Jiang, W. Improved Production of Xylanase in Pichia Pastoris and Its Application in Xylose Production From Xylan. Front. Bioeng. Biotechnol. 2021, 9, 690702. [Google Scholar] [CrossRef]
- Juturu, V.; Aust, C.; Wu, J.C. Heterologous Expression and Biochemical Characterization of Acetyl Xylan Esterase from Coprinopsis Cinerea. World J. Microbiol. Biotechnol. 2013, 29, 597–605. [Google Scholar] [CrossRef]
- Zhang, S.-B.; Wang, L.; Liu, Y.; Zhai, H.-C.; Cai, J.-P.; Hu, Y.-S. Expression of Feruloyl Esterase A from Aspergillus Terreus and Its Application in Biomass Degradation. Protein Expr. Purif. 2015, 115, 153–157. [Google Scholar] [CrossRef]
- Zheng, F.; Liu, J.; Basit, A.; Miao, T.; Jiang, W. Insight to Improve α-L-Arabinofuranosidase Productivity in Pichia Pastoris and Its Application on Corn Stover Degradation. Front. Microbiol. 2018, 9, 3016. [Google Scholar] [CrossRef]
- Adina, S.R.; Suwanto, A.; Meryandini, A.; Puspitasari, E. Expression of Novel Acidic Lipase from Micrococcus Luteus in Pichia Pastoris and Its Application in Transesterification. J. Genet. Eng. Biotechnol. 2021, 19, 55. [Google Scholar] [CrossRef] [PubMed]
- Teng, X.; Chen, X.; Zhu, K.; Xu, H. Immunogenicity of Heparin-Binding Hemagglutinin Expressed by Pichia Pastoris GS115 Strain. Iran. J. Basic Med. Sci. 2018, 21, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-H.; Du, L.; Chag, S.M.; Ma, C.; Tricoche, N.; Tao, X.; Seid, C.A.; Hudspeth, E.M.; Lustigman, S.; Tseng, C.-T.K.; et al. Yeast-Expressed Recombinant Protein of the Receptor-Binding Domain in SARS-CoV Spike Protein with Deglycosylated Forms as a SARS Vaccine Candidate. Hum. Vaccines Immunother. 2014, 10, 648–658. [Google Scholar] [CrossRef]
- Chen, W.-H.; Chag, S.M.; Poongavanam, M.V.; Biter, A.B.; Ewere, E.A.; Rezende, W.; Seid, C.A.; Hudspeth, E.M.; Pollet, J.; McAtee, C.P.; et al. Optimization of the Production Process and Characterization of the Yeast-Expressed SARS-CoV Recombinant Receptor-Binding Domain (RBD219-N1), a SARS Vaccine Candidate. J. Pharm. Sci. 2017, 106, 1961–1970. [Google Scholar] [CrossRef]
- Markets Recombinant Proteins Market by Product, Application, End User-Global Forecasts (2022–2026). Available online: https://www.marketsandmarkets.com/Market-Reports/recombinant-proteins-market-70095015.html (accessed on 8 August 2022).
- Haon, M.; Grisel, S.; Navarro, D.; Gruet, A.; Berrin, J.-G.; Bignon, C. Recombinant Protein Production Facility for Fungal Biomass-Degrading Enzymes Using the Yeast Pichia Pastoris. Front. Microbiol. 2015, 6, 1002. [Google Scholar] [CrossRef]
- Steindorff, A.S.; Serra, L.A.; Formighieri, E.F.; de Faria, F.P.; Poças-Fonseca, M.J.; de Almeida, J.R.M. Insights into the Lignocellulose-Degrading Enzyme System of Humicola Grisea Var. Thermoidea Based on Genome and Transcriptome Analysis. Microbiol. Spectr. 2021, 9, e01088–e01121. [Google Scholar] [CrossRef]
- Liu, W.; Xiang, H.; Zhang, T.; Pang, X.; Su, J.; Liu, H.; Ma, B.; Yu, L. Development of a New High-Cell Density Fermentation Strategy for Enhanced Production of a Fungus β-Glucosidase in Pichia Pastoris. Front. Microbiol 2020, 11, 1988. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, U.; Sohail, M.; Ghanemi, A. Cellulases: From Bioactivity to a Variety of Industrial Applications. Biomimetics 2021, 6, 44. [Google Scholar] [CrossRef]
- Gaber, Y.; Rashad, B.; Hussein, R.; Abdelgawad, M.; Ali, N.S.; Dishisha, T.; Várnai, A. Heterologous Expression of Lytic Polysaccharide Monooxygenases (LPMOs). Biotechnol. Adv. 2020, 43, 107583. [Google Scholar] [CrossRef]
- He, J.; Yu, B.; Zhang, K.; Ding, X.; Chen, D. Expression of Endo-1, 4-Beta-Xylanase from Trichoderma Reesei in Pichia Pastorisand Functional Characterization of the Produced Enzyme. BMC Biotechnol. 2009, 9, 56. [Google Scholar] [CrossRef]
- Michelin, M.; Polizeli, M.d.L.T.M.; Ruzene, D.S.; Silva, D.P.; Vicente, A.A.; Jorge, J.A.; Terenzi, H.F.; Teixeira, J.A. Xylanase and β-Xylosidase Production by Aspergillus Ochraceus: New Perspectives for the Application of Wheat Straw Autohydrolysis Liquor. Appl. Biochem. Biotechnol. 2012, 166, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Pagnonceli, J.; Rasbold, L.M.; Rocha, G.B.; Silva, J.L.C.; Kadowaki, M.K.; Simão, R.C.G.; Maller, A. Biotechnological Potential of an Exo-polygalacturonase of the New Strain Penicillium Janthinellum VI2R3M: Biochemical Characterization and Clarification of Fruit Juices. J. Appl. Microbiol. 2019, 127, 1706–1715. [Google Scholar] [CrossRef] [PubMed]
- Sena, R.O.; Carneiro, C.; Moura, M.V.H.; Brêda, G.C.; Pinto, M.C.C.; Fé, L.X.S.G.M.; Fernandez-Lafuente, R.; Manoel, E.A.; Almeida, R.V.; Freire, D.M.G.; et al. Application of Rhizomucor Miehei Lipase-Displaying Pichia Pastoris Whole Cell for Biodiesel Production Using Agro-Industrial Residuals as Substrate. Int. J. Biol. Macromol. 2021, 189, 734–743. [Google Scholar] [CrossRef] [PubMed]
- García-Ortega, X.; Cámara, E.; Ferrer, P.; Albiol, J.; Montesinos-Seguí, J.L.; Valero, F. Rational Development of Bioprocess Engineering Strategies for Recombinant Protein Production in Pichia Pastoris (Komagataella Phaffii) Using the Methanol-Free GAP Promoter. Where Do We Stand? New Biotechnol. 2019, 53, 24–34. [Google Scholar] [CrossRef]
- Hill, P.; Benjamin, K.; Bhattacharjee, B.; Garcia, F.; Leng, J.; Liu, C.-L.; Murarka, A.; Pitera, D.; Rodriguez Porcel, E.M.; da Silva, I.; et al. Clean Manufacturing Powered by Biology: How Amyris Has Deployed Technology and Aims to Do It Better. J. Ind. Microbiol. Biotechnol. 2020, 47, 965–975. [Google Scholar] [CrossRef]
- Looser, V.; Bruhlmann, B.; Bumbak, F.; Stenger, C.; Costa, M.; Camattari, A.; Fotiadis, D.; Kovar, K. Cultivation Strategies to Enhance Productivity of Pichia Pastoris: A Review. Biotechnol. Adv. 2015, 33, 1177–1193. [Google Scholar] [CrossRef]
- Wehrs, M.; Tanjore, D.; Eng, T.; Lievense, J.; Pray, T.R.; Mukhopadhyay, A. Engineering Robust Production Microbes for Large-Scale Cultivation. Trends Microbiol. 2019, 27, 524–537. [Google Scholar] [CrossRef]
- Duman-Özdamar, Z.E.; Binay, B. Production of Industrial Enzymes via Pichia Pastoris as a Cell Factory in Bioreactor: Current Status and Future Aspects. Protein J. 2021, 40, 367–376. [Google Scholar] [CrossRef]
- Brooks, S.M.; Alper, H.S. Applications, Challenges, and Needs for Employing Synthetic Biology beyond the Lab. Nat. Commun. 2021, 12, 1390. [Google Scholar] [CrossRef]
- Crater, J.S.; Lievense, J.C. Scale-up of Industrial Microbial Processes. FEMS Microbiol. Lett. 2018, 365, fny138. [Google Scholar] [CrossRef]
- Safder, I.; Khan, S.; Islam, I.-u.; Ali, M.K.; Bibi, Z.; Waqas, M. Pichia Pastoris Expression System: A Potential Candidate to Express Protein in Industrial and Biopharmaceutical Domains. Biomed. Lett. 2018, 4, 1–14. [Google Scholar]
- Emenike, V.N.; Schenkendorf, R.; Krewer, U. Model-Based Optimization of Biopharmaceutical Manufacturing in Pichia Pastoris Based on Dynamic Flux Balance Analysis. Comput. Chem. Eng. 2018, 118, 1–13. [Google Scholar] [CrossRef]
- Azadi, S.; Mahboubi, A.; Naghdi, N.; Solaimanian, R.; Mortazavi, S.A. Evaluation of Sorbitol-Methanol Co-Feeding Strategy on Production of Recombinant Human Growth Hormone in Pichia Pastoris. Iran J. Pharm. Res. 2017, 16, 1555–1564. [Google Scholar] [PubMed]
- D’anjou, M.C.; Daugulis, A.J. A Rational Approach to Improving Productivity in RecombinantPichia Pastoris Fermentation. Biotechnol. Bioeng. 2001, 72, 1–11. [Google Scholar] [CrossRef]
- Moser, J.W.; Prielhofer, R.; Gerner, S.M.; Graf, A.B.; Wilson, I.B.H.; Mattanovich, D.; Dragosits, M. Implications of Evolutionary Engineering for Growth and Recombinant Protein Production in Methanol-Based Growth Media in the Yeast Pichia Pastoris. Microb. Cell Factories 2017, 16, 49. [Google Scholar] [CrossRef]
- Garcia-Ortega, X.; Adelantado, N.; Ferrer, P.; Montesinos, J.L.; Valero, F. A Step Forward to Improve Recombinant Protein Production in Pichia Pastoris : From Specific Growth Rate Effect on Protein Secretion to Carbon-Starving Conditions as Advanced Strategy. Process Biochem. 2016, 51, 681–691. [Google Scholar] [CrossRef]
- Garcia-Ortega, X.; Valero, F.; Montesinos-Seguí, J.L. Physiological State as Transferable Operating Criterion to Improve Recombinant Protein Production in Pichia Pastoris through Oxygen Limitation: Physiological State Control to Improve Protein Specific Productivity in Pichia Pastoris with Oxygen Limitation. J. Chem. Technol. Biotechnol 2017, 92, 2573–2582. [Google Scholar] [CrossRef]
- Rathore, A.S.; Mishra, S.; Nikita, S.; Priyanka, P. Bioprocess Control: Current Progress and Future Perspectives. Life 2021, 11, 557. [Google Scholar] [CrossRef]
- Saitua, F.; Torres, P.; Pérez-Correa, J.R.; Agosin, E. Dynamic Genome-Scale Metabolic Modeling of the Yeast Pichia Pastoris. BMC Syst. Biol. 2017, 11, 27. [Google Scholar] [CrossRef]
- Garrigós-Martínez, J.; Weninger, A.; Montesinos-Seguí, J.L.; Schmid, C.; Valero, F.; Rinnofner, C.; Glieder, A.; Garcia-Ortega, X. Scalable Production and Application of Pichia Pastoris Whole Cell Catalysts Expressing Human Cytochrome P450 2C9. Microb. Cell Factories 2021, 20, 90. [Google Scholar] [CrossRef]
- Nieto-Taype, M.A.; Garcia-Ortega, X.; Albiol, J.; Montesinos-Seguí, J.L.; Valero, F. Continuous Cultivation as a Tool Toward the Rational Bioprocess Development With Pichia Pastoris Cell Factory. Front. Bioeng. Biotechnol. 2020, 8, 632. [Google Scholar] [CrossRef] [PubMed]
- Gasset, A.; Garcia-Ortega, X.; Garrigós-Martínez, J.; Valero, F.; Montesinos-Seguí, J.L. Innovative Bioprocess Strategies Combining Physiological Control and Strain Engineering of Pichia Pastoris to Improve Recombinant Protein Production. Front. Bioeng. Biotechnol. 2022, 10, 818434. [Google Scholar] [CrossRef] [PubMed]
- Ponte, X.; Barrigón, J.M.; Maurer, M.; Mattanovich, D.; Valero, F.; Montesinos-Seguí, J.L. Towards Optimal Substrate Feeding for Heterologous Protein Production in Pichia Pastoris (Komagataella Spp) Fed-Batch Processes under PAOX1 Control: A Modeling Aided Approach: Optimal Feeding for Pichia Pastoris. J. Chem. Technol. Biotechnol. 2018, 93, 3208–3218. [Google Scholar] [CrossRef]
- Patra, P.; Das, M.; Kundu, P.; Ghosh, A. Recent Advances in Systems and Synthetic Biology Approaches for Developing Novel Cell-Factories in Non-Conventional Yeasts. Biotechnol. Adv. 2021, 47, 107695. [Google Scholar] [CrossRef]
- Berrios, J.; Flores, M.-O.; Díaz-Barrera, A.; Altamirano, C.; Martínez, I.; Cabrera, Z. A Comparative Study of Glycerol and Sorbitol as Co-Substrates in Methanol-Induced Cultures of Pichia Pastoris: Temperature Effect and Scale-up Simulation. J. Ind. Microbiol. Biotechnol. 2017, 44, 407–411. [Google Scholar] [CrossRef]
- Matthews, C.B.; Kuo, A.; Love, K.R.; Love, J.C. Development of a General Defined Medium for Pichia Pastoris. Biotechnol. Bioeng. 2018, 115, 103–113. [Google Scholar] [CrossRef]
- Liu, W.-C.; Inwood, S.; Gong, T.; Sharma, A.; Yu, L.-Y.; Zhu, P. Fed-Batch High-Cell-Density Fermentation Strategies for Pichia Pastoris Growth and Production. Crit. Rev. Biotechnol. 2019, 39, 258–271. [Google Scholar] [CrossRef]
- Nieto-Taype, M.A.; Garrigós-Martínez, J.; Sánchez-Farrando, M.; Valero, F.; Garcia-Ortega, X.; Montesinos-Seguí, J.L. Rationale-based Selection of Optimal Operating Strategies and Gene Dosage Impact on Recombinant Protein Production in Komagataella Phaffii (Pichia Pastoris). Microb. Biotechnol. 2020, 13, 315–327. [Google Scholar] [CrossRef]
- de Macedo Robert, J.; Garcia-Ortega, X.; Montesinos-Seguí, J.L.; Guimaraes Freire, D.M.; Valero, F. Continuous Operation, a Realistic Alternative to Fed-Batch Fermentation for the Production of Recombinant Lipase B from Candida Antarctica under the Constitutive Promoter PGK in Pichia Pastoris. Biochem. Eng. J. 2019, 147, 39–47. [Google Scholar] [CrossRef]
- Campbell, K.; Xia, J.; Nielsen, J. The Impact of Systems Biology on Bioprocessing. Trends Biotechnol. 2017, 35, 1156–1168. [Google Scholar] [CrossRef]
- Guirimand, G.; Kulagina, N.; Papon, N.; Hasunuma, T.; Courdavault, V. Innovative Tools and Strategies for Optimizing Yeast Cell Factories. Trends Biotechnol. 2021, 39, 488–504. [Google Scholar] [CrossRef] [PubMed]
- Abt, V.; Barz, T.; Cruz-Bournazou, M.N.; Herwig, C.; Kroll, P.; Möller, J.; Pörtner, R.; Schenkendorf, R. Model-Based Tools for Optimal Experiments in Bioprocess Engineering. Curr. Opin. Chem. Eng. 2018, 22, 244–252. [Google Scholar] [CrossRef]
- Hong, M.S.; Velez-Suberbie, M.L.; Maloney, A.J.; Biedermann, A.; Love, K.R.; Love, J.C.; Mukhopadhyay, T.K.; Braatz, R.D. Macroscopic Modeling of Bioreactors for Recombinant Protein Producing Pichia Pastoris in Defined Medium. Biotechnol. Bioeng. 2021, 118, 1199–1212. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, H.; Luna, M.F.; Stosch, M.; Cruz Bournazou, M.N.; Polotti, G.; Morbidelli, M.; Butté, A.; Sokolov, M. Bioprocessing in the Digital Age: The Role of Process Models. Biotechnol. J. 2020, 15, 1900172. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type Species | Strain | Genome Size (Mb) | Isolation | Origin | Carbon Sources | Ref |
|---|---|---|---|---|---|---|
| K. pastoris | CBS 704 | 9.6 | Aesculus species | France | Glucose Glycerol Methanol Ethanol Xylose | ASM170810v1 A |
| K. phaffii | CBS 7435 | 9.4 | Quercus velutina | California, USA | ASM170808v1 A | |
| K. ulmi | CBS 12361 | 9.6 | Ulmus americana | Illinois, USA | [21] | |
| K. kurtzmanii | CBS 12817 | 9.6 | Fir flux | Arizona, USA | ||
| K. mondaviorum | CBS 15017 | 9.5 | Populus deltoides | California, USA | ||
| K. pseudopastoris | CBS 9187 | 10.6 | Salix alba | Hungary | ||
| K. populi | CBS 12362 | 9.3 | Populus deltoides | Illinois, USA |
| Substrate | Product | Genetic modification | Process | Production (g·L−1) | Yield (g·g−1) | Productivity (g·L−1·h−1) | Reference |
|---|---|---|---|---|---|---|---|
| Glycerol | Lactic Acid | Expression of lactate dehydrogenase (LDH) | Batch and fed-batch fermentation | - | 0.70 | - | [36] |
| Lactic Acid | Deletion of ArDH gene | Batch fermentation | 24 | 0.85 | - | [38] | |
| 3-hydroxy propionic acid (3-HP) | Expression of mcr | Fed-batch | 24.75 | 0.13 | 0.54 | [39] | |
| Glycerol and glucose | Isobutanol | Over-expression of the keto acid degradation pathway and medium supplementation | Batch | 2.22 | - | - | [40] |
| Isobutyl acetate ester | Expression of an alcohol-O-acyltransferase enzyme | Batch | 0.051 | - | - | [40] | |
| Isopentyl acetate | Expression of an alcohol-O-acyltransferase enzyme | Batch | 0.024 | - | - | [40] | |
| Glucose | Isobutanol | Expression of LlkivD, ScADH7, PpIlv2, PpIlv3, PpIlv5, PpIlv6 | Shake-flask | 2.22 | 0.22 | - | [40] |
| 2,3-butanediol | Expression of alsS and alsD | 2-L bioreactor, fed-batch | 74.5 | 0.30 | 0.81 | [41] | |
| Glucaric acid | Expression of mMIOX and Udh | Shake flask; fed-batch | 6.61 | - | - | [42] | |
| Inositol | Overexpression of native inositol pathway | Fed-batch fermentation | 30.7 | - | - | [43] | |
| Xylose | Xylitol | Expression of PsXYL1 and gdh | - | 320 (mM) | 0.80 | 2.44 | [44] |
| Xylonic acid | Expression of XDH | 1-L bioreactor; batch | 37 | 0.96 | 0.41 | [45] | |
| Methanol | β-alanine | Overexpression of ADC and aspDH | 1-L bioreactor, fed-batch | 5.60 | - | - | [46] |
| Lactic acid | Multicopy integration of Ldh | Batch | 3.48 | 0.22 | 0.036 | [47] | |
| Malic acid | Overexpression of Mdh, Pyc and SpMae; Δgpi | Shake-flask; batch | 2.79 | - | - | [48] | |
| Lovastatin | Expression of LovB, LovC, LovG, NpgA, LovA, CPR, LovD, LovF | 5-L bioreactor, fed-batch | 0.250 | - | - | [49] | |
| Monacolin J | Expression of LovB, LovC, LovG, NpgA, LovA, CPR | 5-L bioreactor, fed-batch | 0.593 | - | - | [49] | |
| 6-Methylsalicylic acid | Overexpression of atX, npgA | 5-L bioreactor with 3-L operating volume, fed-batch | 2.2 | - | - | [50] | |
| Chondroitin sulfate | Expression of kfoA, kfoC, tuaD, C4ST, PAS_chr1-4_0253, PAS_chr3_0667 | 3-L bioreactor, fed-batch | 2.1 | - | - | [51] | |
| Heparin | Expression of tuaD, kfiC, kfiA, NDST, C5 epi, 2OST,3OST, 6OST | 3-L bioreactor, fed-bach | 2.08 | - | - | [52] | |
| Methanol and Glucose | Hyaluronic acid | Overexpression of xhasA2, xhasB, hasC, hasD, hasE | 2.5-L bioreactor with 1-L operating volume, fed-batch | 0.8–1.7 | - | - | [53] |
| Biomass | Solid Concentration | Biomass Composition (Dry Basis %) | Pretreatment | Hydrolysate Composition (g/L) | ||||
|---|---|---|---|---|---|---|---|---|
| Cellulose | Hemicellulose | Lignin | Acetic acid | Furans | Phenols | |||
| Sugarcane bagasse | 10% | 43.1 | 31.1 | 11.4 | Hot water | 1.1–3.4 | 0.5–5.1 | 1.4–2.4 |
| Corn stover | 10–20% | 37 | 22.7 | 18.6 | Hot water | 2.0–2.8 | 0.74–8.37 | 181–246 AU |
| Wheat straw | 30% | 30.2 | 21 | 17 | Steam explosion | 0.04–1.01 | 0.16–2.14 | nd |
| Maple | 23% | 41 | 15 | 29.1 | Hot water | 13.1 | 4.1 | 1.3 |
| Olive tree pruning | 20% | 25 | 11.1 | 16.2 | Steam explosion | 0.4–4.2 | 0–3.2 | nd |
| Protein | Expression Vector | Promoter | Production | Yield/Activity | Reference |
|---|---|---|---|---|---|
| Malaria vaccine candidate protein | pPICZαA | AOX | 3-L bioreactor, fed-batch | 62.2 g/L | [102] |
| Dengue vaccine candidate protein | pPICZ-A | AOX | Shake-flask; fed-batch | 15 mg/L | [103] |
| Chikungunya vaccine candidate protein | pPIC9K | AOX | Shake-flask; fed-batch | 60 mg/L | [104] |
| Tuberculosis vaccine candidate protein | pPICZαA | AOX | Shake-flask; fed-batch | 5 µg/mL | [105] |
| SARS-CoV-2 Spike RBD | pPICZαA | AOX | 7-L bioreactor, fed-batch | 45 mg/L | [106] |
| Human proinsulin | pPICZα | AOX | Shake-flask; fed-batch | 5 mg/L | [107] |
| Human epidermal growth | pPIC9K | AOX | Shake-flask; fed-batch | 2.27 µm/mL | [108] |
| Interleukin-1beta | pPICZαA | AOX | Shake-flask; fed-batch | 250 mg/L | [109] |
| Antimicrobial Hispidalin | pPICZαA | AOX | Shake-flask; fed-batch | 98.6 µg/mL | [110] |
| Antimicrobial CecropinA-thanatin | pPICZαA | AOX | Shake-flask; fed-batch | 1.061 µmol/L | [111] |
| Antimicrobial PAF102 peptide | pGAPZA | GAP | Shake-flask; batch | 180 mg/L | [112] |
| Antimicrobial fowlicidins | pPICZαA | AOX | Shake-flask; fed-batch | 85.6 mg/L | [113] |
| Human serum albumin | pPIC9K | AOX | Shake-flask; fed-batch | 8.86 g/L | [114] |
| Endoglucanase | pPink-GAP | GAP | 15-L bioreactor, fed-batch | 3 to 5 g/L | [115] |
| Cellobiohydrolase | pPpB1 | AOX | 1-L bioreactor, fed-batch | 6.55 g/L | [116] |
| β-glucosidases | pPIC3.5K | AOX | 5-L bioreactor, fed-batch | 403 mg/L | [117] |
| LPMO | pPICZαA | AOX | Shake-flask; fed-batch | - | [118] |
| Expansin | pPICZαA | AOX | 5-L bioreactor, fed-batch | 4.3 mg/L | [119] |
| Xylanase | pPICZαA | AOX | 7.5-L bioreactor, fed-batch | 2503 U/mL | [120] |
| β-xilosidase | pPICZαA | AOX | Shake-flask; fed-batch | 0.22 mg/L | [121] |
| Feruloyl esterase | pGAPZαA | GAP | Shake-flask; fed-batch | - | [122] |
| Acetyl xylan esterase | pPICZαA | AOX | Shake-flask; fed-batch | 1.5 mg/L | [121] |
| α-L-arabinofuranosidase | pPICZαA | AOX | 7.5-L bioreactor, fed-batch | 164 U/mL | [123] |
| Mannase | pPIC9K | AOX | 10-L bioreactor, fed-batch | 10.47 g/L | [117] |
| Lipase | pPICZαA | AOX | Shake-flask; fed-batch | 145.4 U/mg | [124] |
| Heterologous Product | Optimization Strategy | Observed Improvement | Ref |
|---|---|---|---|
| Erythropoietin (biopharmaceutical product) | Dynamic flux balance analysis (elementary process function integrated with FBA) | The maximum productivity obtained in optimization is 66% higher than the benchmark experimental study | [146] |
| Recombinant Human Growth Hormone (rhGH) | Study of sorbitol-methanol co-feeding strategy and results compared with the basic feeding protocol | Under optimal conditions, cell biomass, total protein, and rhGH concentration increased 15%, 99.5%, and 99.4% | [147] |
| Sea raven antifreeze protein | A model-based approach to optimize qV in a glycerol/methanol mixed-feed continuous stirred-tank reactor | In the optimized conditions, qV was 2.2 mg/L.h, representing a tenfold increase compared with an initial strategy | [148] |
| Growth and AOX-promoter based recombinant protein expression | Adaptive laboratory evolution to improve growth and recombinant protein production in methanol-based growth media | Evolved populations showed increased µ. A selected clone showed increased product titers ranging from a 2.5-fold increase in shake flask batch culture to a 1.8-fold increase in fed-batch cultivation | [149] |
| Human 2F5 antigen-binding fragment (Fab) | Development of a novel operational strategy, with carbon-starving periods and elucidation of the µ effects on the protein secretion | Increments up to 50% of both yields and total production were observed. High µ presented an increment up to 8-fold on the production rates | [150] |
| Human 2F5 antigen-binding fragment (Fab) | Evaluation of a wide range of oxygen-limiting conditions in chemostat cultivations | Specific conditions that lead to the maximum productivity of the process were determined and resulted in an increase of up to 3-fold in qV and YP/X | [151] |
| Human recombinant alpha 1-antitrypsin (A1AT) | A new control system designed for maintaining the μopt during the induction phase. The neural network was applied to adjust and optimize the performance of the robust control system | The newly designed μ-stat control technique enhanced production by up to 1.5 and 2.1 folds in comparison with oxygen-limited fed-batch feeding and mixed feed methods, respectively | [152] |
| Human Serum Albumin (HSA) | Dynamic genome-scale metabolic model for glucose-limited, aerobic cultivations for batch and fed-batch cultures | The model suggested that implementation of a decreasing µ during the feed phase of fed-batch culture results in a 25% increase in qV | [153] |
| Fab fragment (anti-HIV antibody 2F5) | Model for product accumulation in fed-batch, based on iterative calculation, to optimize the time course of the media feed to maximize qV | Good correlation to the optimized model data, and a 2.2-fold improvement of the volumetric productivity in fed-batch optimal profile | [138] |
| Cytochrome P450 enzymes (CYPs) | Rational optimization criteria to optimize production kinetics in bioreactors | Carbon-limiting strategy at the highest µ maximized qP. In the optimum condition, up to threefold increases in terms of qV and yield were achieved in comparison with initial tests | [154] |
| Recombinant hepatitis B small surface antigen (rHBsAg) | Optimization of continuous process efficiency by evaluation of D, and comparison with well-established fed-batch mode | Continuous process reaches similar levels of product titer with qV and qP, respectively, about 1.5 and 1.3 times higher than in fed-batch mode | [155] |
| Candida rugosa lipase | A combination of strain and bioprocess engineering: different gene dosages compared in chemostat cultures with different oxygen-limiting conditions and hypoxic conditions in carbon-limited fed-batch cultures | Increases of up to 9-fold in the production rates were reached when both strain and bioprocess engineering were improved | [156] |
| Candida antartica lipase | Trade-off between fed-batch and continuous. The influence of the μ was examined on various key bioprocess parameters | In continuous mode, the overall production was 5.8 times greater than the fed-batch process | [155] |
| Streptomyces ghanaenis L-glutamate oxidase | Study of the effect of feeding strategy on cell growth and enzyme production | The cell density and total enzyme activity were 210 g/L and 118 U/mL, respectively, which represent a 3-fold and 36-fold increase relative to shake flask experiments | [142] |
| Trichoderma reesei xylanase | HCDF under optimal parameters | HCDF strategy boosts the amount of enzyme by 40.1-fold in comparison to the shake flask fermentation | [142] |
| Rhizopus oryzae lipase | A numerical optimization of mathematical model which includes cell substrate and product kinetics | The optimal profiles were defined, and bioprocess efficiency improvement was confirmed in terms of a 2.2-fold higher final titer and 3.4-fold higher productivity | [157] |
| Applicable to different products | Combination of three metabolic models optimized with constraint-based FBA | Simulated values were highly comparable with existing experimental results that outperformed each model. This similarity can be useful to reduce experimental work and costs in optimization process | [158] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carneiro, C.V.G.C.; Serra, L.A.; Pacheco, T.F.; Ferreira, L.M.M.; Brandão, L.T.D.; Freitas, M.N.d.M.; Trichez, D.; Almeida, J.R.M.d. Advances in Komagataella phaffii Engineering for the Production of Renewable Chemicals and Proteins. Fermentation 2022, 8, 575. https://doi.org/10.3390/fermentation8110575
Carneiro CVGC, Serra LA, Pacheco TF, Ferreira LMM, Brandão LTD, Freitas MNdM, Trichez D, Almeida JRMd. Advances in Komagataella phaffii Engineering for the Production of Renewable Chemicals and Proteins. Fermentation. 2022; 8(11):575. https://doi.org/10.3390/fermentation8110575
Chicago/Turabian StyleCarneiro, Clara Vida Galrão Corrêa, Luana Assis Serra, Thályta Fraga Pacheco, Letícia Maria Mallmann Ferreira, Lívia Teixeira Duarte Brandão, Mariana Nogueira de Moura Freitas, Débora Trichez, and João Ricardo Moreira de Almeida. 2022. "Advances in Komagataella phaffii Engineering for the Production of Renewable Chemicals and Proteins" Fermentation 8, no. 11: 575. https://doi.org/10.3390/fermentation8110575
APA StyleCarneiro, C. V. G. C., Serra, L. A., Pacheco, T. F., Ferreira, L. M. M., Brandão, L. T. D., Freitas, M. N. d. M., Trichez, D., & Almeida, J. R. M. d. (2022). Advances in Komagataella phaffii Engineering for the Production of Renewable Chemicals and Proteins. Fermentation, 8(11), 575. https://doi.org/10.3390/fermentation8110575